09/10, Estelle Raffin
1/27
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No study sessions yet.
28 Terms
Les rôles multiples de la motricité
La motricité désigne l’ensemble des processus nerveux et musculaires qui permettent la production de mouvements. Elle est essentielle à la survie, à la communication et à l’adaptation à l’environnement.
a) Fonctions biologiques vitales et de reproduction
La motricité permet d’assurer les fonctions physiologiques indispensables :
S’alimenter (mouvements de préhension et de mastication)
Respirer (la ventilation dépend de muscles squelettiques tels que le diaphragme et les intercostaux)
Fuir ou se défendre en cas de danger
b) Fonctions de relation avec l’environnement
La motricité joue également un rôle fondamental dans les interactions sociales et émotionnelles :
Langage corporel : positions du corps, gestuelle (ex. positions de « high power » et « low power », Amy Cuddy, Harvard University)
Communication non verbale : mouvements oculaires, expressions faciales
Expression émotionnelle à travers la posture et les mimiques
Ainsi, la motricité est omniprésente dans la vie quotidienne, et permet une interaction constante avec l’environnement.
Le système moteur ne sert pas uniquement à agir sur le monde, mais aussi à communiquer et décoder les messages non verbaux produits par autrui.
Motricité et cognition : une relation réciproque
Les activités motrices permettent d’explorer, de manipuler et de comprendre le monde.
Elles sont donc à la base de l’acquisition d’informations sensorielles et du développement des capacités cognitives.
En retour, nos capacités cognitives influencent nos comportements moteurs.
Il existe une complémentarité étroite entre le système moteur et le système sensoriel, donnant lieu à une intégration sensorimotrice essentielle à la production de mouvements coordonnés.
La complémentarité sensorimotrice
a) Le système sensoriel
Le système sensoriel fournit au cerveau :
une représentation du monde extérieur,
et des informations internes sur la position et l’état du corps.
Il transforme des énergies physiques (lumière, son, pression, etc.) en flux organisés de potentiels d’action interprétables par le système nerveux.
Principales modalités sensorielles :
Vision
Audition
Toucher
Olfaction
Gustation
La complémentarité sensorimotrice
b) La perception de soi dans l’espace
La perception de la position et des mouvements du corps repose sur plusieurs sources d’information :
Système vestibulaire : ≈ 20 %
Récepteurs musculaires et articulaires (propriocepteurs) : ≈ 70 %
Vision : ≈ 10 %
Ces signaux convergent vers le système nerveux central pour permettre la stabilisation posturale, l’équilibre et la coordination motrice.
Organisation du système moteur
Le système moteur génère des potentiels d’action conduisant à la contraction musculaire, transformant ainsi l’information nerveuse en énergie mécanique (force).
Les réponses motrices assurent :
le maintien de la posture,
l’équilibre,
et la réalisation de mouvements dirigés.
Organisation du système moteur
On distingue trois grandes catégories de mouvements, qui se différencient par leurs circuits neuronaux et leur niveau de contrôle volontaire :
a) Les mouvements réflexes
Réponses automatiques, involontaires et innées, déclenchées par des récepteurs sensoriels musculaires, articulaires ou cutanés.
Ils régulent le tonus musculaire et assurent la posture.
L’intégration se fait au niveau de la moelle épinière (ex. réflexe myotatique ou tendineux).
b) Les activités motrices rythmiques
Comportements présentant une composante réflexe et une composante volontaire de moindre importance.
Exemples : respiration, marche, mastication.
Ces activités dépendent de circuits situés dans la moelle épinière et le tronc cérébral, tout en pouvant être modulées par des commandes corticales.
c) Les mouvements volontaires
Mouvements délibérés et finalisés, planifiés et exécutés sous le contrôle du cortex moteur.
Le cortex envoie des commandes descendantes via les voies cortico-spinales, qui activent les motoneurones de la moelle épinière.
Ces circuits permettent le contrôle précis et contextuel des muscles squelettiques.
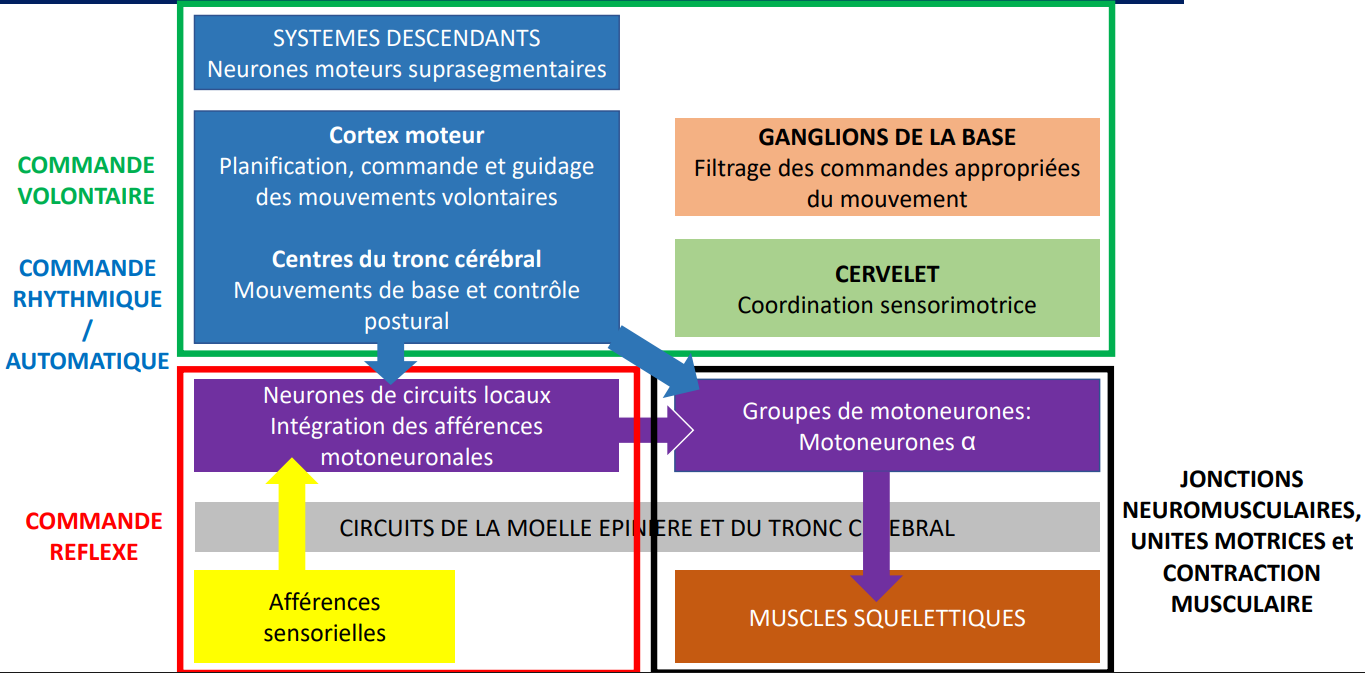
Organisation hiérarchique du contrôle moteur
Les réflexes sont gérés localement par la moelle épinière.
Les activités rythmiques impliquent le tronc cérébral et la moelle épinière.
Les mouvements volontaires dépendent du cortex cérébral, qui exerce un contrôle descendant sur les structures inférieures.
Points essentiels à retenir
Le système moteur et le système sensoriel travaillent conjointement pour produire des mouvements efficaces et adaptés.
La motricité sert à la fois des fonctions vitales, sociales et cognitives.
Le contrôle moteur est organisé de manière hiérarchique (moelle → tronc cérébral → cortex).
Les niveaux de contrôle de la motricité
La motricité repose sur une organisation hiérarchique du système nerveux impliquant trois grands niveaux :
la moelle épinière,
le tronc cérébral,
et les aires corticales motrices.
Ces structures sont interconnectées : chacune peut agir de manière autonome, mais également moduler ou être modulée par les autres.
1. La moelle épinière
a) Organisation et structure
La moelle épinière, protégée par la colonne vertébrale, s’étend du tronc cérébral jusqu’à la vertèbre L1.
Elle constitue un centre d’intégration réflexe et un relais de communication entre le cerveau et le reste du corps.
Elle contient :
une substance grise centrale (corps cellulaires neuronaux) ;
une substance blanche périphérique (axones assurant la circulation de l’information nerveuse).
La moelle comporte 31 paires de nerfs spinaux :
8 cervicaux (C1–C8)
12 thoraciques (T1–T12)
5 lombaires (L1–L5)
5 sacrés (S1–S5)
1 coccygien
Elle se termine à L1 car sa croissance s’arrête vers 4 ans, tandis que la colonne vertébrale continue de s’allonger jusqu’à environ 18 ans.
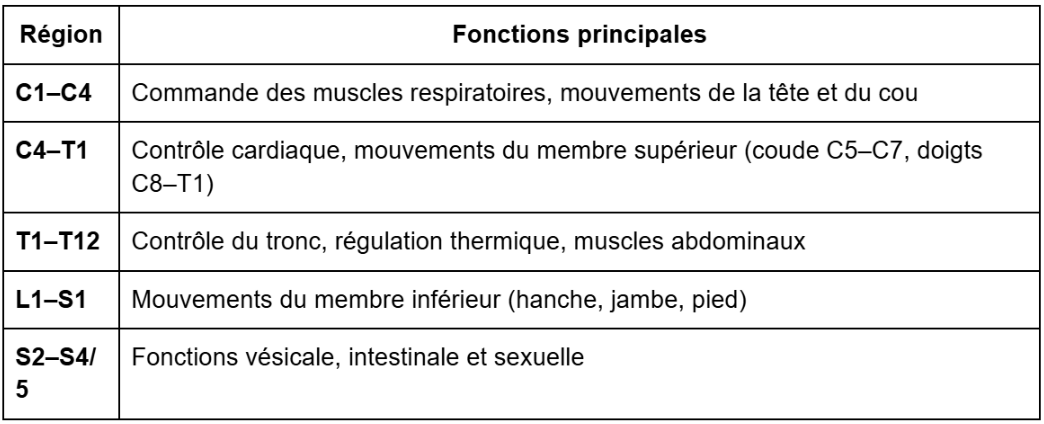
Les niveaux de contrôle de la motricité
1. La moelle épinière
b) Fonctions principales
Les plexus cervical et brachial sont particulièrement importants sur le plan clinique, car les lésions à ces niveaux peuvent avoir de lourdes conséquences fonctionnelles.
c) Rôle moteur : les réflexes et l’innervation réciproque
La moelle épinière assure la mise en œuvre de réflexes moteurs reposant sur des connexions mono- ou polysynaptiques.
Ces réflexes permettent :
la régulation du tonus musculaire,
les réflexes de retrait en réponse à des stimulations nociceptives.
Un principe fondamental est celui de l’innervation réciproque :
lorsqu’un réflexe provoque la contraction d’un muscle agoniste, il entraîne simultanément le relâchement du muscle antagoniste, grâce à un jeu d’excitation et d’inhibition coordonné.
Ainsi, la moelle épinière fonctionne comme un centre réflexe intégré, capable de produire des réponses automatiques et coordonnées sans intervention directe du cerveau.
Les niveaux de contrôle de la motricité
2. Le tronc cérébral
Le tronc cérébral, situé entre la moelle épinière et le cerveau, est composé de :
la moelle allongée (bulbe rachidien),
le pont (protubérance),
et le mésencéphale.
Fonctions :
Régulation du tonus musculaire et de la posture, grâce à ses projections descendantes sur la moelle épinière.
Contrôle automatique de fonctions vitales : respiration, rythme cardiaque, cycles veille-sommeil, etc.
Voie de passage essentielle pour les influx sensoriels ascendants et les commandes motrices descendantes.
Le tronc cérébral constitue donc un centre moteur automatique qui ajuste en permanence l’activité de la moelle épinière.
Les niveaux de contrôle de la motricité
3. Le cortex cérébral
Le cortex cérébral représente le niveau le plus élevé de la hiérarchie motrice.
Il comprend plusieurs aires motrices corticales (primaire, prémotrice, aire motrice supplémentaire) ayant des rôles et des connexions distincts.
Le cortex moteur primaire (M1) envoie des commandes directes aux motoneurones de la moelle via le faisceau corticospinal (ou pyramidal).
D’autres aires motrices se projettent sur les noyaux du tronc cérébral, modulant ainsi les fonctions automatiques et posturales.
Le cortex est donc responsable de la planification, de la commande et du contrôle volontaire des mouvements.
Résumé
Trois niveaux hiérarchiques : moelle épinière, tronc cérébral, cortex moteur.
La moelle assure les réflexes et la transmission des commandes.
Le tronc cérébral régule le tonus, la posture et les automatismes vitaux.
Le cortex commande la motricité volontaire.
Le système moteur combine un contrôle hiérarchique et un fonctionnement parallèle pour une exécution fluide et adaptative du mouvement.
Les niveaux de contrôle de la motricité
4. Une organisation hiérarchique et parallèle
La motricité est dite hiérarchique, car :
la moelle épinière fonctionne de manière autonome pour les réflexes ;
le tronc cérébral module l’activité de la moelle ;
le cortex cérébral exerce un contrôle descendant sur les deux niveaux inférieurs.
Cependant, cette organisation est également parallèle :
Les différents niveaux peuvent agir simultanément et interagir entre eux.
Par exemple, une commande volontaire du cortex peut activer un motoneurone tout en inhibant localement un réflexe via un circuit spinal, afin d’ajuster le mouvement.
Ainsi, la motricité humaine repose à la fois sur un fonctionnement top-down (descendant) et sur des mécanismes locaux intégrés, garantissant la souplesse, la coordination et la précision des actions motrices.
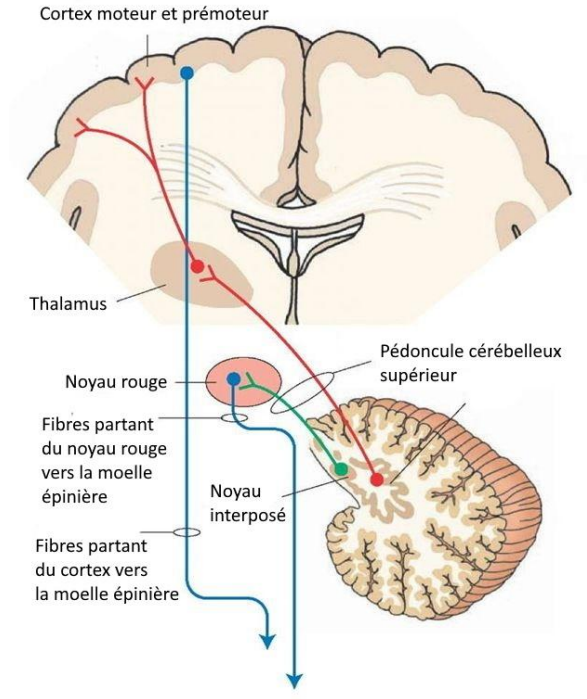
Autres structures impliquées dans le contrôle moteur
En complément de la moelle épinière, du tronc cérébral et du cortex moteur, deux structures jouent un rôle essentiel dans la coordination et la modulation des mouvements volontaires :
→ les noyaux gris centraux (ou ganglions de la base)
→ le cervelet
Ces deux systèmes n’initient pas directement le mouvement, mais ajustent et harmonisent son exécution selon le contexte et les besoins.
Le cervelet
Structure et connexions
Le cervelet est constitué de deux hémisphères, séparés par une structure médiane appelée vermis, et divisés en lobe antérieur et lobe postérieur.
Il présente une surface plissée semblable à celle du cortex et contient plusieurs noyaux profonds, dont le noyau dentelé, particulièrement impliqué dans la motricité.
Il reçoit un grand nombre de fibres afférentes provenant de presque toutes les régions du cerveau (notamment des aires frontales et pariétales) ainsi que de la moelle épinière.
Ses fibres efférentes, issues des cellules de Purkinje, se projettent principalement vers le cortex prémoteur et d’autres régions motrices, formant une boucle cortico-cérébelleuse essentielle pour le contrôle fin du mouvement.
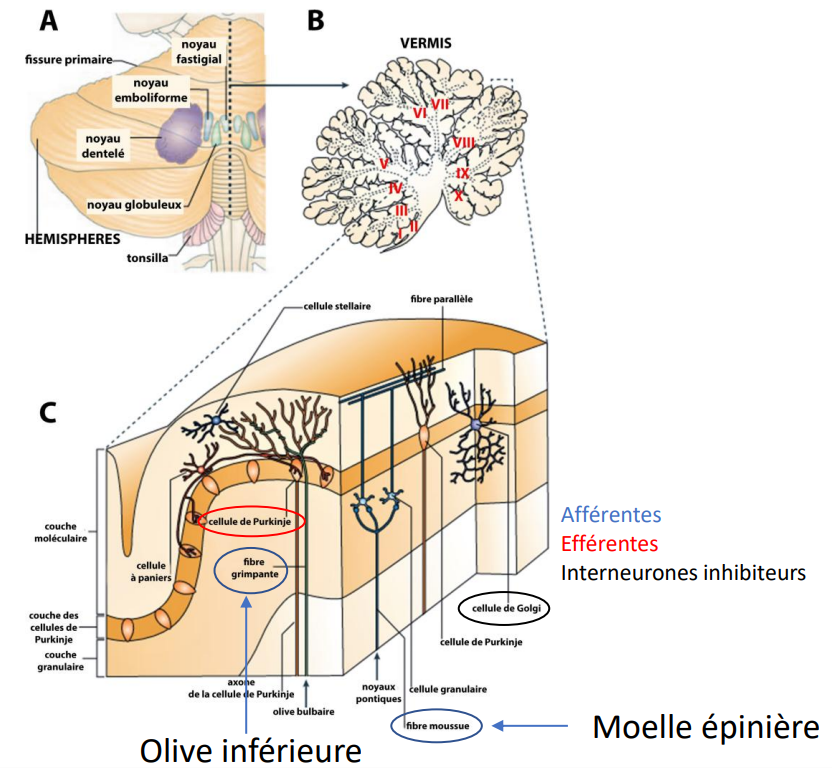
Autres structures impliquées dans le contrôle moteur
Le cervelet
Rôle fonctionnel
Le cervelet agit comme un comparateur :
il reçoit à la fois une copie efférente des commandes motrices envoyées aux muscles par le cortex moteur,
et des informations sensitives de la moelle épinière indiquant la position réelle du corps.
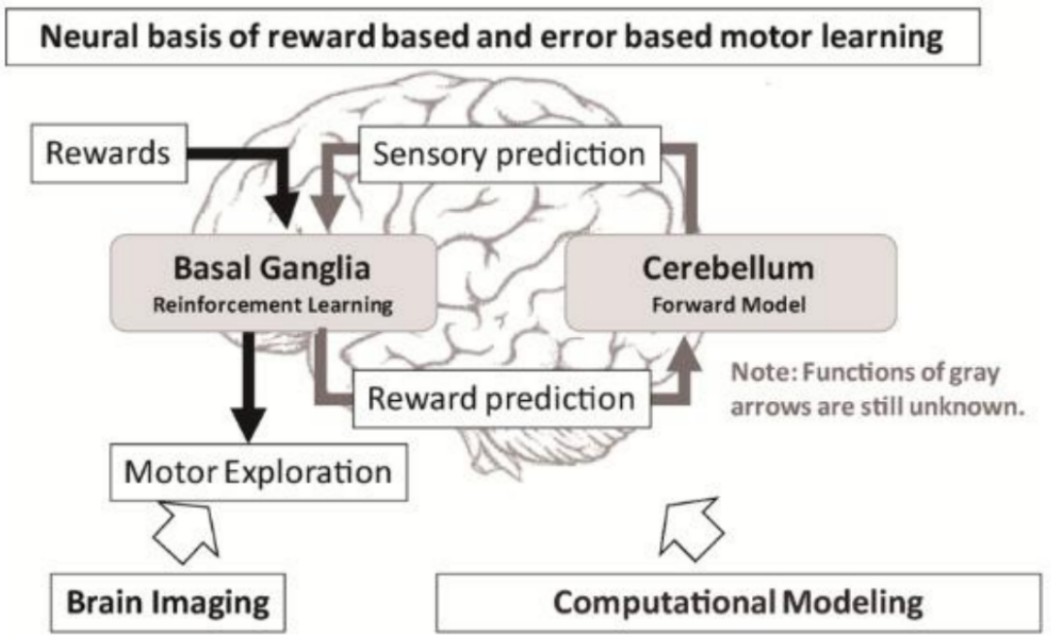
Ce mécanisme lui permet de détecter les écarts entre le mouvement prévu et le mouvement réalisé, et de corriger les erreurs en temps réel pour assurer la précision spatiale et temporelle des gestes.
Il joue donc un rôle central dans :
la coordination et l’harmonisation des mouvements,
la correction des erreurs motrices,
et surtout l’apprentissage moteur par rétroaction sensorielle (apprentissage par l’erreur).
Un exemple classique est celui des tâches d’adaptation visuomotrice (pointer une cible décalée artificiellement) : le cervelet permet la correction progressive du geste grâce à la comparaison entre prédiction et retour sensoriel.
Autres structures impliquées dans le contrôle moteur
Le cervelet
Dysfonctionnements
Les syndromes cérébelleux illustrent l’importance de cette structure :
les mouvements deviennent maladroits, tremblés et mal synchronisés.
Les patients présentent des difficultés à activer correctement les muscles agonistes et antagonistes, entraînant une perturbation des phases d’accélération et de freinage du geste.
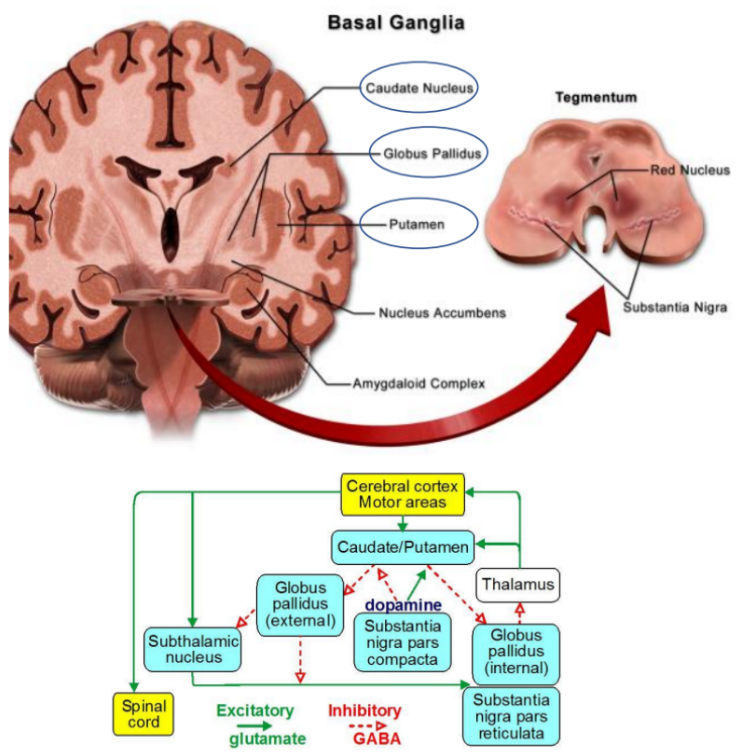
Les ganglions de la base
Organisation et connexions
Les ganglions de la base regroupent plusieurs structures sous-corticales interconnectées :
le striatum (noyau caudé + putamen),
le globus pallidus,
le noyau sous-thalamique,
la substance noire,
et leurs connexions avec le thalamus et le cortex frontal.
Ils reçoivent des afférences de l’ensemble du cortex cérébral et renvoient leurs projections vers le cortex prémoteur et moteur via le thalamus, formant ainsi une boucle cortico-striato-thalamo-corticale.
Autres structures impliquées dans le contrôle moteur
Les ganglions de la base
Rôle fonctionnel
Les ganglions de la base jouent un rôle majeur dans :
la sélection et l’initiation des mouvements volontaires,
la régulation des schémas moteurs automatiques et habituels,
et l’apprentissage moteur par renforcement (apprentissage basé sur la récompense).
Le striatum contient des neurones dopaminergiques sensibles aux signaux de récompense, qui codent la valeur et la pertinence d’une action.
Ainsi, cette structure participe à la motivation du mouvement et à l’ajustement du comportement moteur selon les conséquences (positives ou négatives) de l’action.
Dysfonctionnements
Les troubles des ganglions de la base se traduisent par des altérations du contrôle moteur :
Dans la maladie de Parkinson, la dégénérescence dopaminergique de la substance noire perturbe l’activité du striatum. Cela entraîne une akinésie (difficulté à initier le mouvement) et des dyskinésies (mouvements involontaires).
Les patients présentent également un apprentissage par récompense altéré, en lien avec la perturbation des circuits dopaminergiques.
Ainsi :
Les ganglions de la base interviennent dans la préparation et la sélection de l’action (motricité anticipée, pro-active).
Le cervelet intervient dans la correction et l’adaptation dynamique du geste (motricité rétroactive).
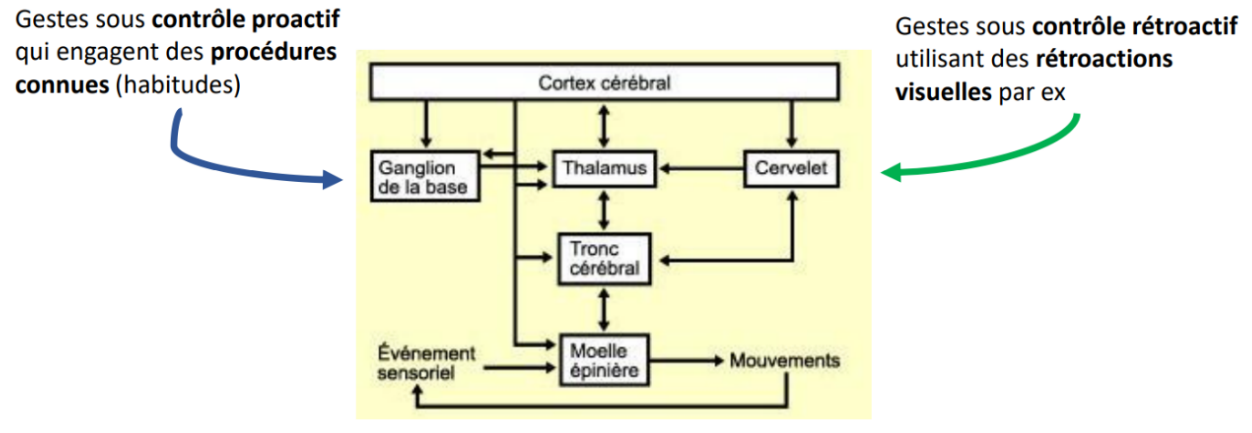
En résumé
Le cervelet assure la coordination, la précision temporelle et spatiale, et l’apprentissage par l’erreur.
Les ganglions de la base contrôlent la sélection des actions, les automatismes moteurs et l’apprentissage par renforcement.
Ces deux systèmes modulent les circuits moteurs du tronc cérébral et du cortex selon des principes complémentaires, garantissant la fluidité, la souplesse et la pertinence du mouvement.
La jonction neuromusculaire et le système musculo-articulaire
Organisation fonctionnelle du système musculo-articulaire
Le système musculo-articulaire comprend l’ensemble des structures impliquées dans le mouvement :
les os,
les muscles,
les tendons,
les ligaments,
les articulations,
le cartilage,
et d’autres tissus conjonctifs assurant le soutien et la transmission des forces mécaniques.
Le muscle présente une organisation hiérarchique (ou fractale) : il est constitué de faisceaux de fibres musculaires, eux-mêmes formés de myofibrilles, dont l’unité fonctionnelle est le sarcomère — la plus petite structure contractile du muscle.
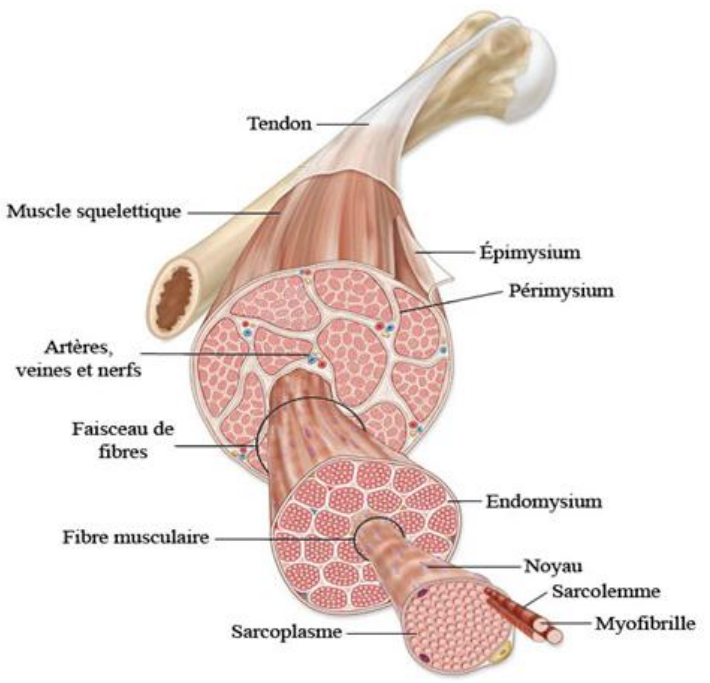
La jonction neuromusculaire et le système musculo-articulaire
Les types de fibres musculaires
On distingue trois grands types de muscles selon leur structure et leur mode de contraction :
Muscles striés squelettiques
Responsables des mouvements volontaires (marcher, saisir un objet, déglutir, etc.).
Présentent une organisation striée correspondant à la répétition des sarcomères le long des myofibrilles.
Muscles lisses
Assurent la motricité involontaire (péristaltisme intestinal, vasoconstriction, dilatation).
Contraction lente et soutenue, indépendante de la volonté.
Muscle cardiaque (myocarde)
Contraction involontaire et rythmique, sous le contrôle du système nerveux autonome.
Organisation striée mais réseau cellulaire spécifique (disques intercalaires).
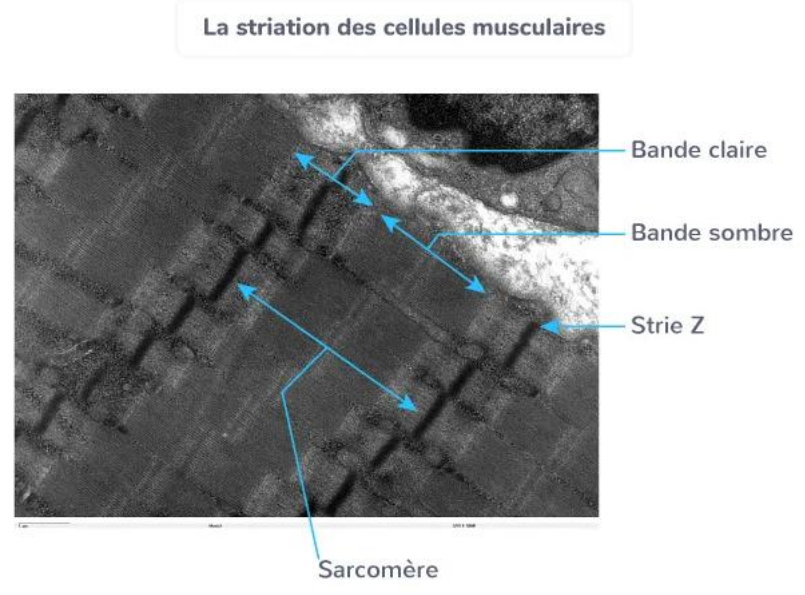
La jonction neuromusculaire et le système musculo-articulaire
Les sarcomères : unité fonctionnelle de contraction
Le sarcomère est l’unité contractile fondamentale du muscle strié.
Il est constitué de :
filaments fins d’actine,
filaments épais de myosine,
et de protéines régulatrices (troponine, tropomyosine).
Lors de la contraction, les filaments d’actine glissent entre les filaments de myosine, provoquant le raccourcissement du sarcomère et donc la contraction du muscle.
La succession de ces sarcomères le long des myofibrilles explique l’aspect strié du muscle squelettique.
La commande nerveuse du muscle : les motoneurones
Rôle et types de motoneurones
Deux sous-types :
Motoneurones α (alpha) : innervent les fibres musculaires extrafusales, responsables de la contraction effective du muscle.
Motoneurones γ (gamma) : innervent les fuseaux neuromusculaires (fibres intrafusales) impliqués dans la régulation du tonus et de la posture.
Le sarcolemme désigne la membrane plasmique de la fibre musculaire, interface entre le signal nerveux et la réponse contractile.
La commande nerveuse du muscle : les motoneurones
Rôle et types de motoneurones
La contraction musculaire est déclenchée par les motoneurones, cellules nerveuses qui transmettent les ordres du système nerveux central (SNC) vers les fibres musculaires.
On distingue :
Les motoneurones supérieurs (ou centraux)
Situés dans le cortex moteur et les centres moteurs du tronc cérébral.
Contrôlent les motoneurones inférieurs.
Les motoneurones inférieurs (ou périphériques)
Localisés dans la moelle épinière.
Leurs axones myélinisés sortent par les racines ventrales pour innerver directement les fibres musculaires.
Représentent la voie finale commune de la motricité.
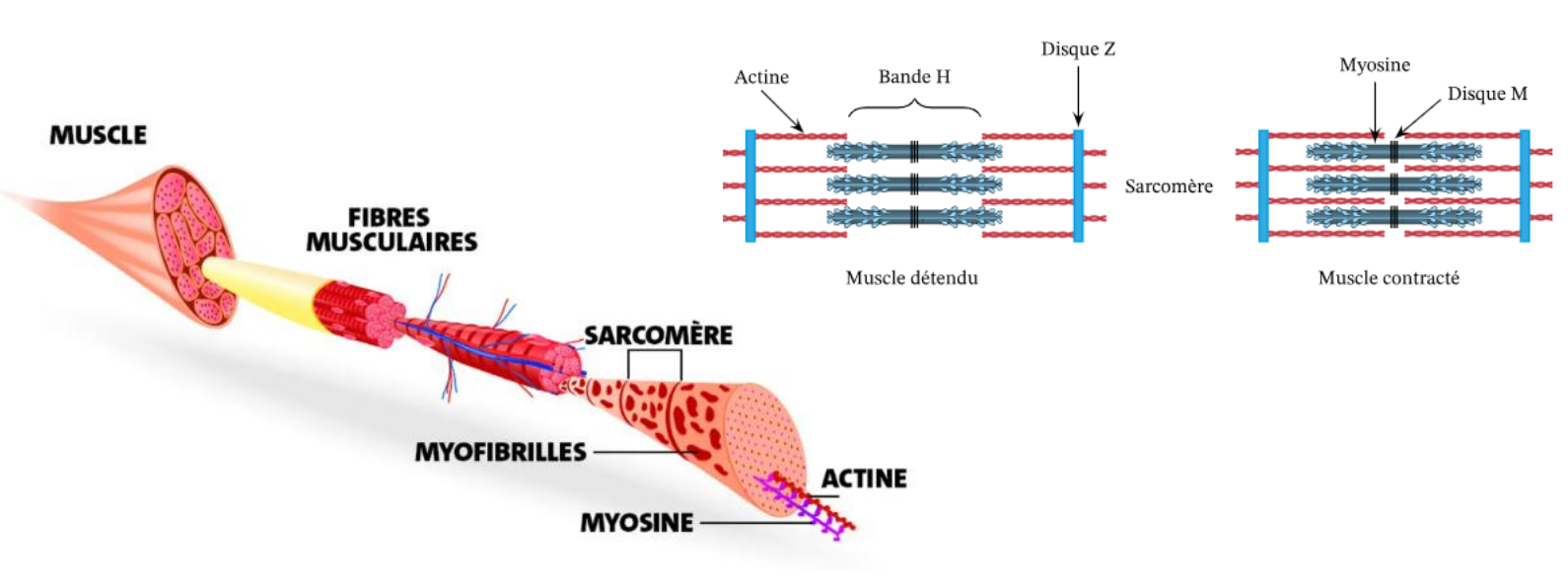
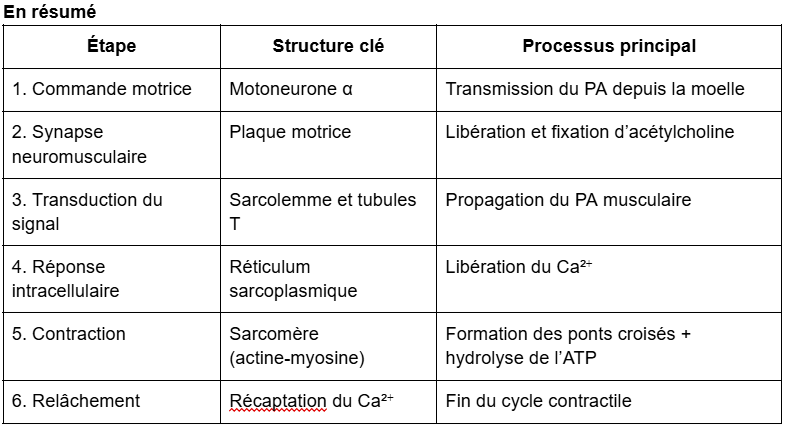
De l’influx nerveux à la contraction musculaire
I. Transmission du signal électrique à la jonction neuromusculaire
Lorsque le potentiel d’action (PA) arrive à la terminaison axonale du motoneurone (bouton synaptique) :
Il provoque l’ouverture de canaux calciques voltage-dépendants.
L’entrée de Ca²⁺ dans le bouton synaptique déclenche la fusion des vésicules synaptiques contenant l’acétylcholine (ACh).
L’ACh est libérée dans la fente synaptique et diffuse vers la plaque motrice de la fibre musculaire.
Elle se fixe sur les récepteurs nicotiniques situés sur le sarcolemme, déclenchant la dépolarisation de la membrane musculaire.
Cette synapse neuromusculaire est dite cholinergique, car son neurotransmetteur est l’acétylcholine.
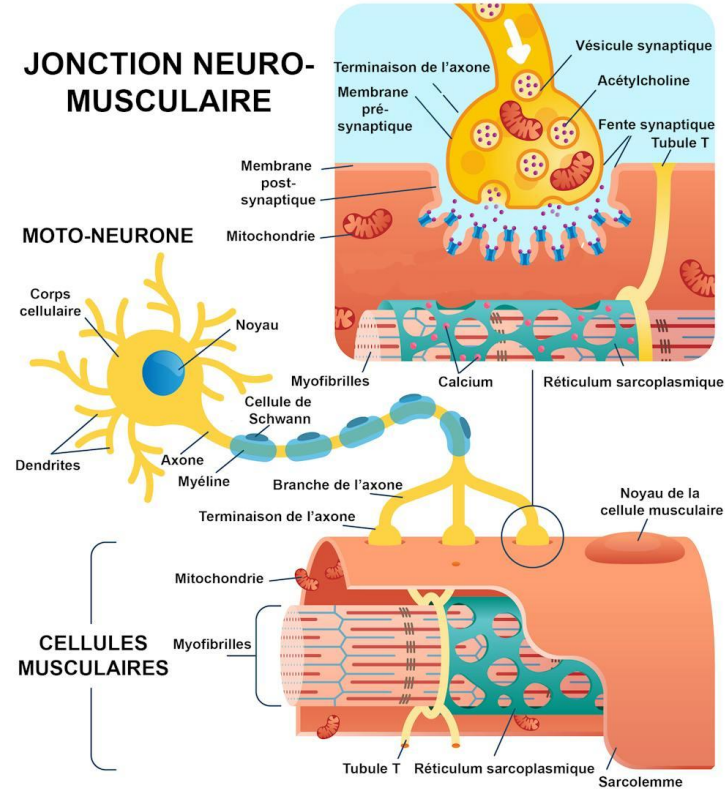
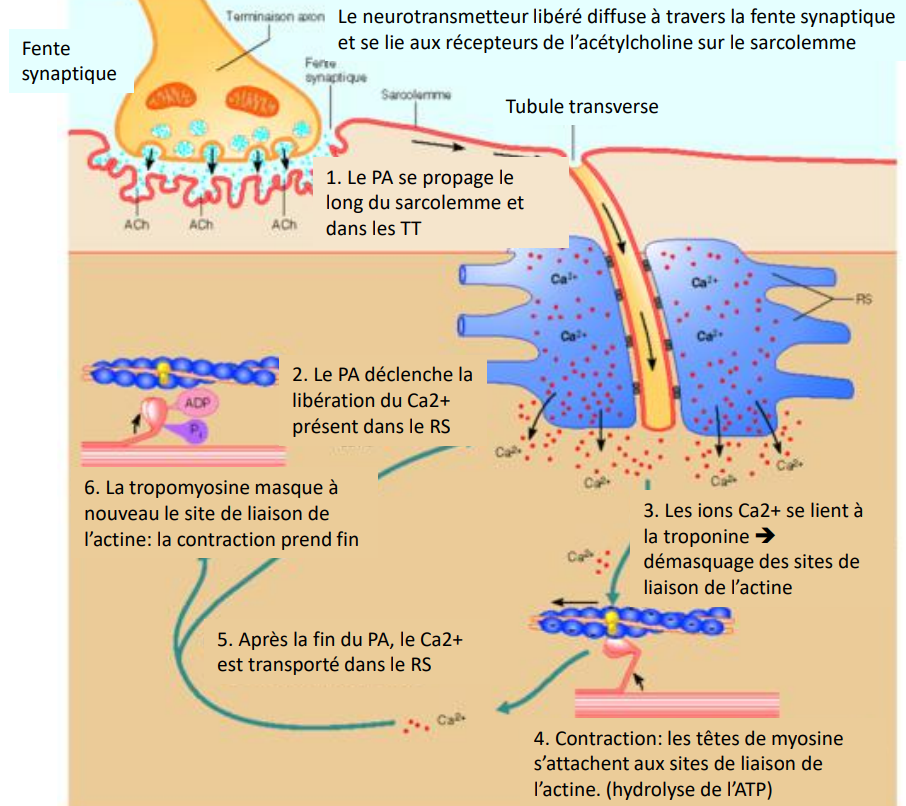
De l’influx nerveux à la contraction musculaire
II. Transformation du signal électrique en signal chimique puis mécanique
La fixation de l’ACh sur ses récepteurs induit un potentiel d’action musculaire qui se propage le long du sarcolemme et dans les tubules T.
Les tubules T transmettent le signal au réticulum sarcoplasmique (RS), provoquant la libération de Ca²⁺ dans le sarcoplasme.
Le Ca²⁺ se lie à la troponine, ce qui entraîne un déplacement de la tropomyosine et dévoile les sites de liaison de l’actine.
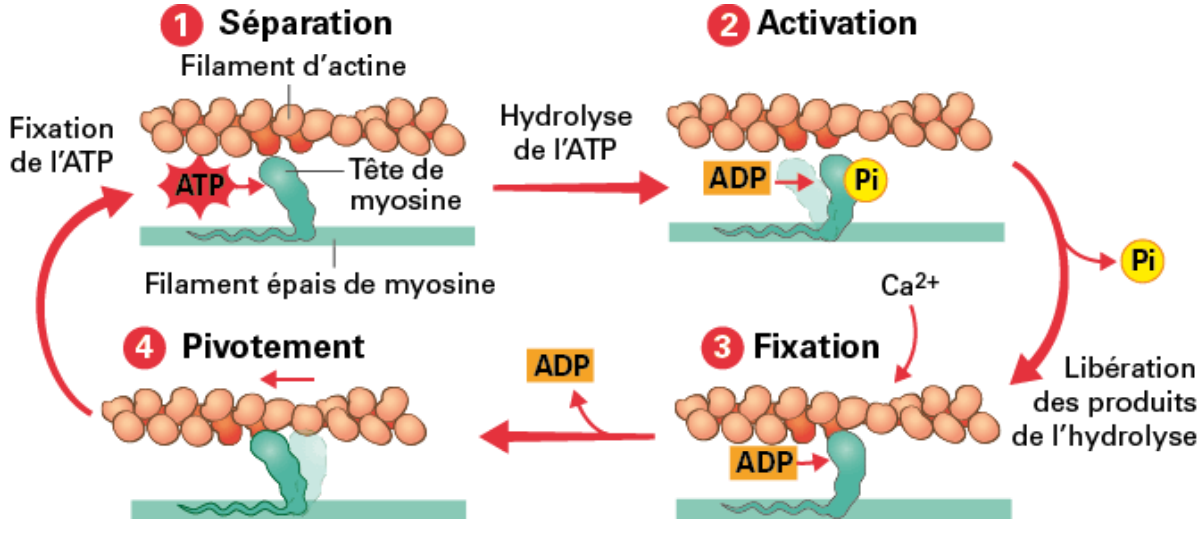
Les têtes de myosine peuvent alors se fixer sur l’actine et effectuer un mouvement de pivotement : c’est le cycle des ponts d’union (cross-bridges).
Chaque cycle consomme une molécule d’ATP, hydrolysée en ADP + Pi, fournissant l’énergie nécessaire à la contraction.
À la fin du PA, le Ca²⁺ est recapté par le RS ; la tropomyosine reprend sa position initiale, le muscle se relâche.
Les 5 étapes clés de la contraction musculaire
Libération de l’ACh dans la fente synaptique.
Dépolarisation du sarcolemme et propagation du PA dans les tubules T.
Libération du Ca²⁺ par le réticulum sarcoplasmique.
Interaction actine–myosine et hydrolyse de l’ATP → génération de la force.
Recaptation du Ca²⁺ → relâchement du muscle.
L’unité motrice : base du recrutement musculaire
Une fibre musculaire est innervée par un seul motoneurone.
Un motoneurone peut, en revanche, innerver plusieurs fibres musculaires.
Cette association (un motoneurone + l’ensemble des fibres qu’il contrôle) forme une unité motrice, qui constitue l’unité fonctionnelle du recrutement musculaire.
→ Plus le ratio d’innervation est faible, plus le mouvement est fin et précis (ex. muscles oculaires).
→ Les muscles posturaux, en revanche, présentent un ratio élevé, adapté à des contractions soutenues et globales.
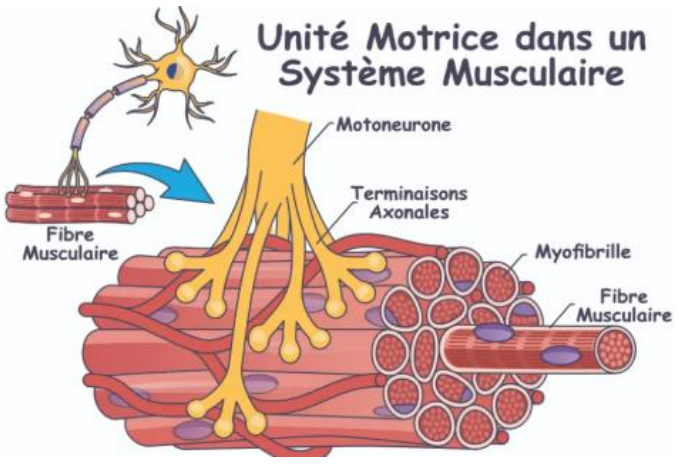
Unités motrices
Une unité motrice correspond à un motoneurone et à l’ensemble des fibres musculaires qu’il innerve.
Chaque fibre musculaire n’est innervée que par un seul motoneurone, mais un motoneurone peut innerver un nombre variable de fibres musculaires, ce qui détermine le rapport d’innervation.
Ce rapport influence la précision ou la force d’un mouvement :
Dans les muscles fins (ex. muscles extraoculaires), le rapport est faible : un motoneurone n’innerve que quelques fibres, ce qui permet un contrôle très précis.
Dans les muscles posturaux ou des membres inférieurs, un motoneurone innerve de nombreuses fibres, favorisant une production de force importante plutôt qu’une finesse motrice.
Ainsi, plus un mouvement exige de la précision, plus le rapport d’innervation est faible.
Types d’unités motrices
On distingue deux grands types d’unités motrices selon leurs caractéristiques fonctionnelles :
Unités motrices lentes (type I) : fibres musculaires lentes, contractions faibles mais soutenues, grande résistance à la fatigue, métabolisme aérobie.
Unités motrices rapides (type II) : fibres rapides, contractions puissantes et brèves, fatigabilité élevée, métabolisme anaérobie.
Principe de fonctionnement
Contraction synchrone : lorsqu’un motoneurone décharge, toutes les fibres musculaires qu’il innerve se contractent simultanément.
Régulation de la force : pour des mouvements précis ou faibles, seules quelques unités lentes sont recrutées. Si la force nécessaire augmente, des unités rapides sont progressivement ajoutées.
Hiérarchie de recrutement (principe de Henneman) : les unités lentes sont toujours recrutées avant les unités rapides, ces dernières étant plus coûteuses sur le plan énergétique.
Génération et modulation de la force
La force musculaire peut être modulée de deux façons :
Recrutement spatial : activation d’un nombre croissant d’unités motrices au sein d’un même muscle.
→ Plus la stimulation est forte, plus le nombre d’unités recrutées augmente, jusqu’à atteindre un plateau.Recrutement temporel : augmentation de la fréquence des décharges d’un même motoneurone.
→ Lorsque les stimulations se succèdent rapidement, le muscle n’a plus le temps de se relâcher : les contractions se sommant donnent un tétanos, c’est-à-dire une contraction soutenue maximale.On parle de tétanos parfait lorsque la tension atteint un plateau stable sans relâchement.
Le rapport entre ces deux modes de recrutement varie selon le type de muscle :
Motricité fine (main) : prédominance du recrutement spatial, le recrutement temporel n’intervenant qu’à haute intensité.
Muscles puissants (jambes) : prédominance initiale du recrutement temporel, avant l’ajout d’un recrutement spatial à haute intensité.
Fatigabilité
Les unités motrices rapides, plus puissantes, sont aussi plus fatigables : elles cessent de se contracter plus tôt que les unités lentes, qui sont moins énergivores et adaptées aux efforts prolongés (ex. sports d’endurance).
Adaptations à l’entraînement
Lors d’un entraînement de force, les premières adaptations observées sont nerveuses :
amélioration de la coordination entre motoneurones et fibres musculaires,
optimisation du recrutement des unités motrices.
Ce n’est qu’ensuite qu’apparaît une hypertrophie musculaire (augmentation de la masse musculaire).
Points essentiels à retenir
La jonction neuromusculaire est une synapse cholinergique entre le motoneurone et la fibre musculaire.
La contraction musculaire résulte du glissement des filaments d’actine et de myosine au sein du sarcomère.
L’unité motrice, formée par un motoneurone et les fibres musculaires qu’il innerve, représente l’unité fonctionnelle de base du mouvement.
Chaque muscle contient un grand nombre d’unités motrices, dont le mode de recrutement et de décharge détermine la force et la précision du mouvement.
Les différents types de contractions musculaires
La contraction musculaire correspond à une modification de la longueur des fibres musculaires sous l’action du système nerveux. Selon la relation entre tension développée et mouvement produit, on distingue plusieurs types de contractions :
Contraction isométrique :
Le muscle développe une tension sans modification de sa longueur.
Exemple : tenir une bouteille d’eau sans bouger. Le muscle est contracté mais ne se raccourcit pas — il maintient simplement la position contre la gravité.
Contraction concentrique :
Elle correspond à un raccourcissement des fibres musculaires, le muscle produisant une force supérieure à la résistance externe.
Exemple : porter la bouteille d’eau à la bouche. Le muscle se raccourcit pour rapprocher les segments articulaires.Contraction excentrique :
Elle implique un allongement du muscle sous tension, lorsque la résistance externe dépasse la force développée par le muscle.
Exemple : reposer la bouteille sur la table. Le muscle contrôle le mouvement tout en s’allongeant.Contraction isocinétique (plus rare) :
Il s’agit d’une contraction où la vitesse d’allongement ou de raccourcissement reste constante, souvent observée dans des conditions expérimentales ou en rééducation motrice.
Maladies neuromusculaires (MNM)
Une maladie neuromusculaire désigne toute affection altérant le fonctionnement du muscle et donc la motricité, que l’origine soit musculaire ou liée à un autre élément de l’unité motrice.
Les structures pouvant être atteintes sont :
le motoneurone (corps cellulaire dans la corne antérieure de la moelle épinière),
la jonction neuromusculaire,
ou les fibres musculaires elles-mêmes.
Selon le site de l’atteinte, on distingue plusieurs types de pathologies :
Myopathies : atteinte directe du muscle,
Myasthénie : atteinte de la jonction neuromusculaire,
Neuropathies périphériques : atteinte des nerfs périphériques,
Sclérose latérale amyotrophique (SLA) et amyotrophies spinales : atteinte du motoneurone.
Les causes peuvent être :
Primitives : d’origine génétique ou auto-immune,
Secondaires : liées à une infection, une maladie endocrinienne ou une exposition toxique.