H1 Baena- Introduction to the organelles of the eukaryotic cell
1/5
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No study sessions yet.
6 Terms
why do mitochondria and chloroplasts require a specialised transport system?
after the endosymbiotic events, huge amounts of gene transfer occurred from the endosymbionts to the nucleus so that they could be controlled
chloroplasts and mitochondria have highly reduced genomes, so they can’t produce all the proteins they need, which must be made by the nucleus
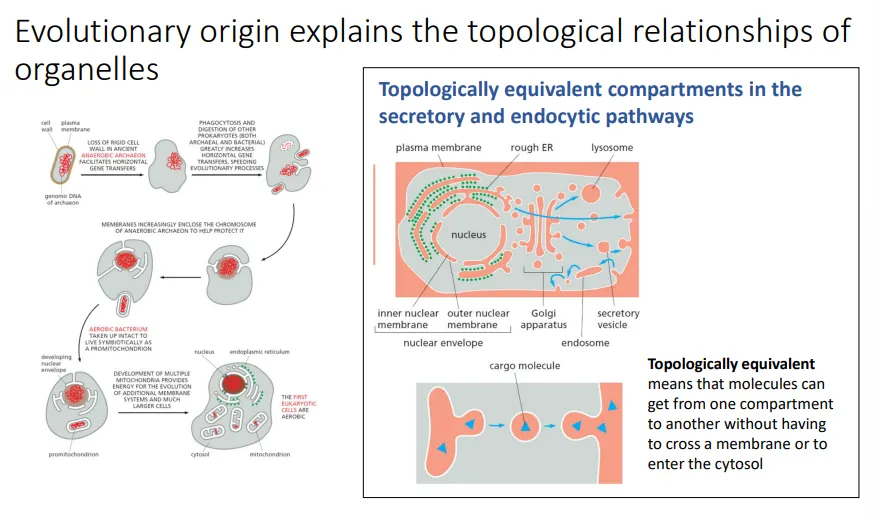
the other cell organelles are topologically similar, and so they can interconnect, integrate and exchange molecules easily without them crossing membranes
however, organelles of endosymbiotic origin are isolated from this vesicular traffic system because they evolved separately
this means they need their own transport system
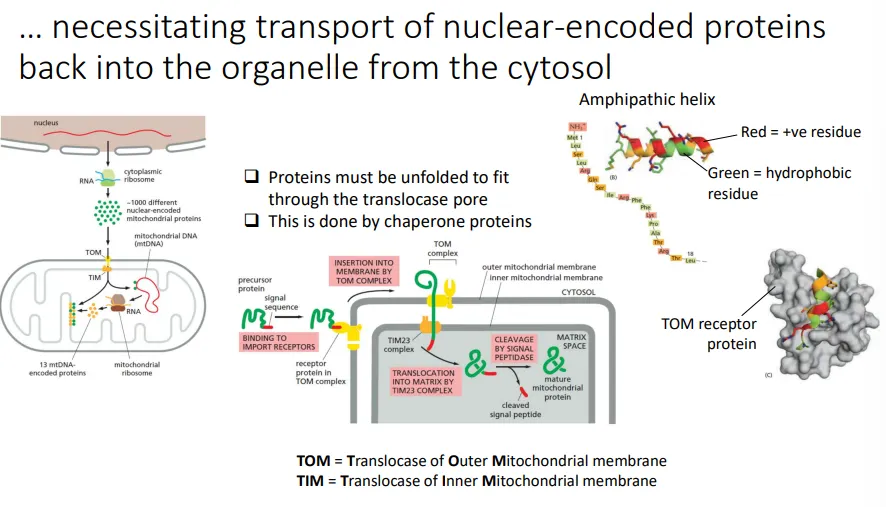
how are proteins imported into the mitochondria?
the mitochondrial proteins that must be imported from the cytosol (originating from genes in the nucleus) have a signal peptide sequence at the N-terminus
this is a recognition motif for the TOM (outer membrane) and TIM (inner membrane) protein complexes
when the sequence is recognised, it triggers the transient linearisation of the precursor peptide so that it can be translocated across the double membrane into the mitochondrial matrix
then the signal peptide is cleaved off by signal peptidase to produce the mature protein
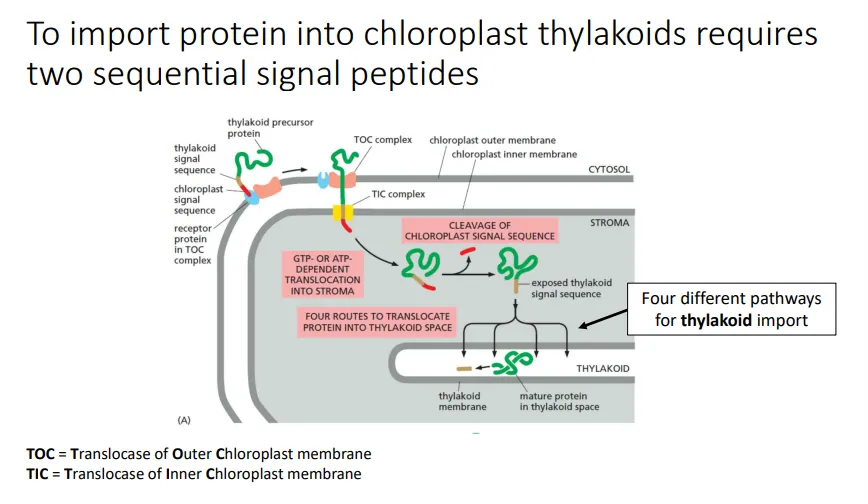
how are proteins imported into the chloroplast thylakoids?
the chloroplast proteins that must be imported from the cytosol (originating from genes in the nucleus) into the thylakoid lumen have two signal peptide sequences at the N-terminus
the first chloroplast signal sequence is a recognition motif for the TOC (outer membrane) and TIC (inner membrane) protein complexes
when the sequence is recognised, it triggers the transient linearisation of the precursor peptide so that it can be translocated across the double membrane into the chloroplast stroma
then the chloroplast signal peptide is cleaved off by signal peptidase, which exposes the second thylakoid signal sequence
this is recognised and allows the protein to be imported into the thylakoid lumen
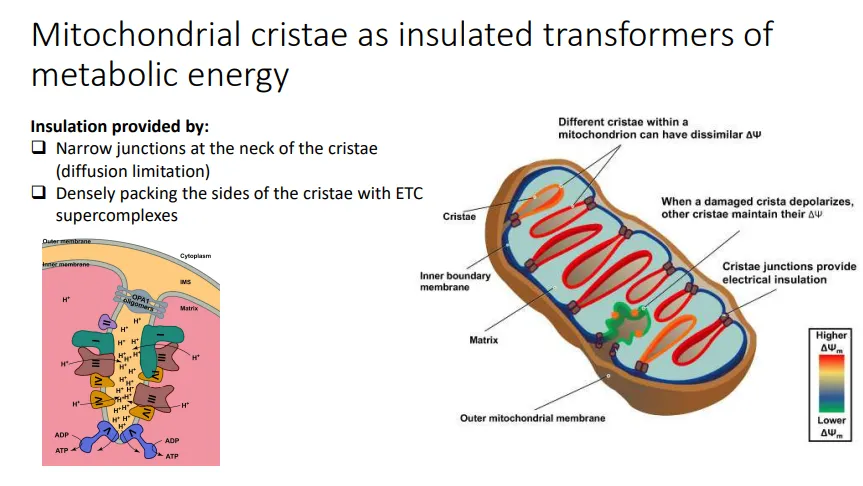
how are invaginations in mitochondria organised?
MICOS and OPA1 oligomers are protein complexes that create the junction between the cristae and the rest of the intermembrane space, at the ‘neck’
this insulates the invagination so that the proton gradients are maximised, as protons can’t diffuse into the rest of the IMS
this also means that if one cristae is damaged, the rest of the cristae don’t get depolarised, so the mitochondrion can remain functional
ATP synthase dimers are located at the tip, which results in the curvature of the membrane
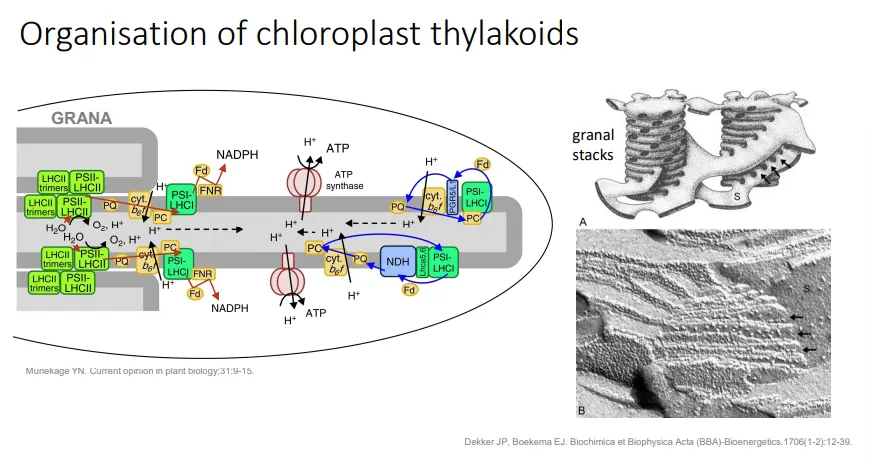
how are invaginations in chloroplasts organised?
thylakoids are stacked into grana, which maximises the absorption of light
the granal stacks are connected by single thylakoids, because some sections of the thylakoid membrane must be exposed to the stroma
this allows the ETC and ATP synthase to release NADPH and ATP into the stroma for the calvin-benson cycle to occur
what are peroxisomes?
peroxisomes are organelles that metabolise fatty acids by beta-oxidation when glucose is scarce (which can go through the glycolytic pathway instead)
they use oxygen to remove electrons from the substrate, producing harmful hydrogen peroxide, which they then detoxify using catalase
the fatty acids are attached to coenzyme A, oxidised by using FAD to transfer 2 electrons and 2 protons to oxygen, and converted into acetyl CoA (which can enter the TCA cycle) by a series of enzymes
mammals evolved an electron transfer flavoprotein in their mitochondria, which is used instead of FAD and oxygen, passing the electrons directly to the ETC instead of producing hydrogen peroxide (though they still also use peroxisomes)
they are also involved in detoxifying the compounds produced in photorespiration, a wasteful process where O2 competes with CO2 as a substrate for RuBisCo (as it evolved in anaerobic conditions), so that they can be used for sugar production