Looks like no one added any tags here yet for you.
Jaké jsou dvě formy organizace chemických reakcí v buňce?
Soustředění enzymů do proteinových komplexů a soustředění metabolických dějů do organel.
Jaké jsou výhody kompartmentalizace v buňce?
- V buňkách probíhá mnoho protichůdných chemických procesů, jejich oddělení zabrání chaosu.
- Vytvoření vhodného chemického mikroprostředí pro daný typ chemických procesů (řeší pomalost difúze substrátů, obsah iontů, pH) – reakce probíhají snadněji a rychleji.
- Izolace potenciálně nebezpečných rozkladných dějů
- Dělba práce mezi organelami
Které organely obsahují hydrolytické enzymy?
Lysosomy.
Které organely obsahují oxidativní enzymy?
Peroxisomy.
Jaký je rozdíl mezi cytoplazmou a cytosolem?
Cytoplazma zahrnuje cytosol a organely, zatímco cytosol je omezen plazmatickou membránou.
Nevýhody kompartmentalizace
- Dělba práce má mnoho výhod, ale také různé nároky.
- Biosyntéza organel
- Nutnost koordinace
- Třídění proteinů pro různé destinace...
Membránové organely - Prokaryotická buňka:
bez specializovaných vnitřních oddílů – jediný oddíl obklopený membránou = cytosol
Membránové organely - Eukaryotická buňka
- bohatě strukturována do samostatných oddílů s vyhraněnou funkcí
- Vysoká organizace na podbuněčné úrovni
- Další organizační (nadbuněčné) úrovně: buňky – tkáně – orgány – orgánové systémy - organismy
Cytoplazma
tekutina složená z cytosolu a organel (vše mezi plazmatickou membránou a jádrem)
Cytosol
omezen plazmatickou membránou a vnějším povrchem organel
Evoluce organel
- Předchůdci prvních eukaryot: mikroorganismy podobné bakteriím (žádné vnitřní membrány)
- Plazmatická membrána zřejmě zajišťovala všechny membránové funkce (vč. syntézy ATP)
- Umožněno malou velikostí původních buněk
- Velké eukaryotické buňky jsou závislé na vnitřních membránách
vznik membrány jádra, ER, GA, endozomů a lysozomů
- atrně vznikly vchlípením plazmatické membrány
- Vzájemná komunikace prostřednictvím váčků
- Vchlipování membrány pozorováno u dnešních fotosyntetizujících bakterií
vznik mitochondrií a chloroplastů
- Mitochondrie a chloroplasty mají vlastní DNA – podobnost s bakteriálním genomem
- Patrně se vyvinuly z bakterií pohlcených primitivními eukaryotickými buňkami
- Nezapojí se do vezikulárního transportu
Zvětšování organel
- Běžné v průběhu buněčného cyklu
- Přidávání nových molekul: přísun lipidů a proteinů
- Nutnost správného třídění materiálu
Třídění proteinů
- Nastává po dokončení nebo v průběhu jejich syntézy
- Posttranslační transport z cytosolu do mitochondrií, chloroplastů, peroxisomů, vnitřního prostoru jádra
- Kotranslační transport přes ER do Golgiho aparátu, lysosomů, endosomů, jaderné membrány, plazmatické membrány, extracelulárního prostoru
Jaké jsou tři hlavní mechanismy importu proteinů do organel?
- Syntéza všech proteinů začíná na ribozomech cytosolu (s výjimkou několika mitochondriálních a chloroplastových proteinů)
- Do vnitřního prostoru jádra: přes jaderné póry
- Do mitochondrií, chloroplastů, peroxisomů: přes proteinové translokátory
- Do ER, Golgiho aparátu, endosomů, lysosomů, membrán, vnějšího prostoru: vezikulárním transportem
Translokace proteinů z cytosolu do organel
- Import proteinů do organel určuje adresová sekvence – 15-60 aminokyselin - je součástí proteinu
- u ER zvaná signální sekvence
- Po navedení proteinů do cílových struktur odštěpena
- Absence adresové sekvence: protein zůstává v cytosolu
- Umělá výměna adresové sekvence mění lokalizaci proteinu
Jaderná membrána
Dvě vrstvy
Vnitřní: vazebná místa pro chromozomy a jadernou laminu
Jaderná lamina: síť proteinových vláken poskytující vnitřní oporu
Vnější: složením velmi příbuzná membráně ER, ve kterou přechází
perinukleární prostor kontinuálně přechází v lumen ER
Jak probíhá transport jadernými póry?
- Pór prostupuje vnější i vnitřní jadernou membránou
- Funguje jako selektivní branka, která aktivně přenáší specifické makromolekuly a zároveň umožňuje volnou difúzi menších molekul oběma směry
- Z cytosolu do jádra: jaderné proteiny, transkripční faktory, atd.
- Z jádra do cytosolu: RNA, ribosomální podjednotky (nesestřižená pre-mRNA jádro neopouští)
- Větší molekuly a makromolekulární komplexy nemohou póry přecházet bez signálu (tzv. jaderného lokalizačního signálu, NLS)
- Na NLS se vážou pomocné cytosolové proteiny (importiny), které navádějí protein k póru a zajišťují jeho translokaci
- Nutná energetická podpora hydrolýzou GTP
- Importiny se pórem vracejí do cytosolu
Struktura jaderného póru
- Velký počet proteinových vláken na obou stranách póru
- Vlákna neomezují průchodnost
- Nevyžadována změna konformace přenášených molekul
Co je signální sekvence pro ER?
15-60 aminokyselinová sekvence, která určuje transport proteinu do endoplazmatického retikula.
export RNA z jádra
- rRNA, tRNA, mRNA vznikají v jádře, ale fungují v cytoplazmě
- Jadernou membránou prostupují póry, nutná energie
- Přenášené RNA se spojují s proteiny obsahujícími adresovou sekvenci pro export z jádra
- Translokace membránou probíhá v komplexu s proteiny (exportiny)
translokace do mitochondrií a chloroplastů
- Obě organely specializovány na syntézu ATP
- Vymezeny vnitřní a vnější membránou
- U chloroplastů navíc třetí membránový systém – thylakoidní membrána
- Proteiny importované z cytosolu mají na N-konci adresovou sekvenci k přesunu dochází ve speciálních místech vzájemného dotyku membrán
- Během transportu se proteiny rozvíjejí
- Adresová sekvence se odštěpuje signální peptidázou
- Obnovení konformace uvnitř organely napomáhají chaperony
Endoplazmatické retikulum
- Cytoplazmatický systém membránově uzavřených tubulů a váčků
- Napojení na jadernou membránu
- Největší organela většiny eukaryotických buněk (10% celkového objemu)
- Tvoří přibližně polovinu všech buněčných membrán
Hladké ER
Není spojeno s ribozomy, tvorba a odbourávání cukrů
Metabolismus cholesterolu a lipoproteinů
Detoxifikační reakce
Místo syntézy membránových a zásobních lipidů, a dalších lipofilních sloučenin (steroidů)
Zásobárna Ca2+
Drsné ER
Na vnějším povrchu pokryto ribozomy, které syntetizují proteiny pro ER, Golgiho aparát, lysosomy, plazmatickou membránu a sekreční proteiny
Vstup proteinů do endoplazmatického retikula
- Nastává ještě před dokončením jejich syntézy - kotranslačně
- Podmínkou je přítomnost signální sekvence pro ER na N-konci
- Spojen s translací
- Ribozom syntetizující příslušný protein musí být v kontaktu s membránou ER
- Vznikající protein proniká membránou ER a nemůže se vrátit zpět do cytosolu
Z cytosolu se do ER přenášejí 2 druhy proteinů:
- rozpustné: jsou do lumen ER přeneseny úplně (určeny k sekreci nebo pro lumen ER nebo jinou organelu spojenou se sekreční drahou)
- Membránové: zůstávají zanořeny v membráně ER (transmembránové proteiny ER, jiných organel nebo plazmatické membrány)
Proteosyntéza na ribozomech drsného ER
- Volné a ribozomy vázané na ER jsou nerozlišitelné, liší se jen proteiny, které zrovna syntetizují
- Syntéza všech proteinů je zahájena na volných ribozomech
- Signální sekvence na N-konci vznikajícího polypeptidu přitahuje ribozom k drsnému ER
- Signální sekvenci tvoří krátký úsek hydrofobních AK, který se odštěpuje během přenosu do lumen ER
Navádění ribozomů k membráně ER
- Zapojeny jsou nejméně dva proteiny:
- SRP („signal recognition particle“) - přítomen v cytosolu
- Váže se k signální sekvenci
- Receptor pro SRP
- Zanořen v membráně ER
- Přitahuje komplex ribozom/peptid/SRP k membráně ER
Přenos proteinů do ER
- Translací vzniká polypeptidový řetězec se signální sekvencí na N-konci
- Signální sekvence je rozeznána a vázána SRP
- Zpomalení syntézy daného proteinu
- Komplex SRP-ribozom se váže k receptoru SRP v membráně ER
- Po uvolnění SRP (recyklace) signální sekvence otvírá translokační kanál, kterým se protein přesunuje přes membránu
- Zároveň se obnovuje jeho syntéza
- Po splnění své naváděcí role je signální sekvence odštěpena signální peptidázou
Syntéza transmembránových proteinů
- Nepřecházejí do lumen úplně, ale zůstávají zanořeny v membráně ER
- Translokace je přerušena pomocnou sekvencí hydrofobních aminokyselin, která ukotvuje protein v membráně (sekvence zastavující přenos)
- Po odštěpení signální sekvence a umístění proteinu v lipidové dvojvrstvě proteosyntéza pokračuje na cytosolové straně membrány
Vezikulární transport
- Obousměrný dopravní systém mezi ER a plazmatickou membránou
- Zajištěn váčky, které pučí z membrán a zase s nimi fúzují
- Forma komunikace mezi vnitřkem buňky a okolím
- Sekreční dráha (z ER -> GA ->) a endocytóza (do buňky)
Sekreční dráha:
ER – Golgi – lysosom/membrána
- Biosyntéza proteinů, vstup do ER
- Přesun do Golgiho aparátu k buněčnému povrchu nebo přes endosomy k lysosomům
- Vstup proteinů do ER je hlavní křižovatkou v dopravě proteinů eukaryotickou buňkou
Endocytická dráha:
- membrána - lysosom
- Molekuly z vnějšího prostředí uzavřeny do váčků plazmatické membrány
- Přesun k raným endosomům a lysosomům
- Při „dálkovém“ transportu jsou váčky vedeny pomocnými proteiny, které se pohybují podél vláken cytoskeletu, jinak difúze
- Hydrolytické enzymy v lysosomech zajistí rozklad přijatého materiálu
transportní váčky
- Malé organely vymezené lipidovou membránou, které se pohybují buňkou
- Zajišťují přepravu materiálu mezi oddíly sekreční/endocytické dráhy
- Molekuly se dostávají do váčků odvozených z vnitřních organelových membrán (ER, Golgi) (sekreční dráha) nebo z plazmatické membrány, která zaškrcením obklopuje část mimobuněčného prostoru (endocytóza)
- Náklad se vyloží v jiném buněčném oddílu splynutím membrány váčku s membránou cílové organely nebo membránou plazmatickou
- Systém je využíván membránovými lipidy i protein
Specifita váčků
- Přijímají jen proteiny určené pro příslušný cíl
- Fúzují jen s příslušnou cílovou membránou
- Podmínka: každá organela udržuje identitu – má charakteristické zastoupení proteinů a lipidů v plazmatické membráně
- Mezi organelami zajišťují kyvadlovou dopravu různé typy váčků, každý z nich nese specifický náklad
Vchlipování váčků
- Poháněno skládáním proteinového pláště
- Specifický proteinový plášť obklopuje váčky na cytosolové straně
- Po ukončení vchlípení váček plášť ztrácí (usnadnění následné fúze s membránou cílové organely)
- Existují různé typy opláštěných váčků se specifickými obaly
Dvě hlavní funkce plášťů:
- Tvarování membrány do váčku
- Zachytávání molekul určených k transportu
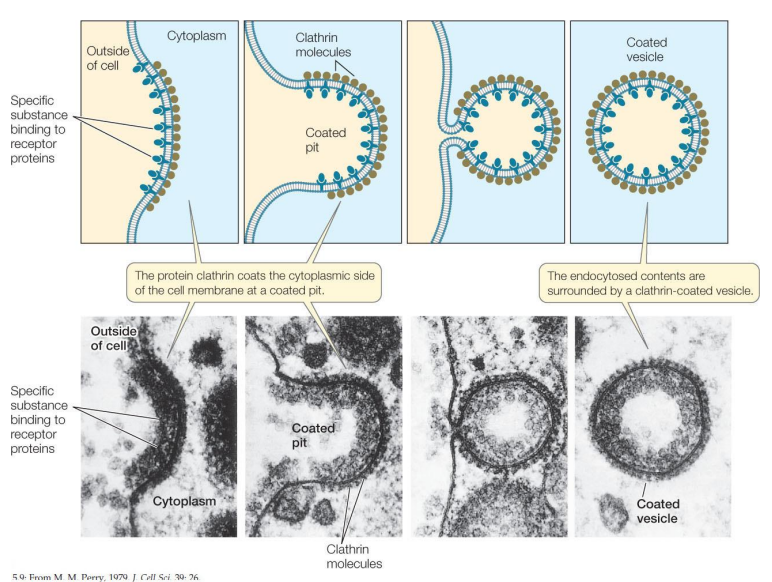
Co jsou klathrinové váčky?
nejlépe popsanél
Pučí z Golgiho aparátu v rámci sekreční dráhy a z plazmatické membrány na počátku endocytické dráhy
Hlavní plášťový protein – klathrin
Molekuly klathrinu se na cytosolové straně membrány skládají do sítě
Membránu tak nutí k zaškrcení do podoby váčku
Klathrinové váčky pučící z vnitřního povrchu plazmatické membrány kožních buněk
Vazbu klathrinu k membráně zajišťují adaptiny
Malý GTP-vázající protein dynamin se obtáčí kolem vchlípeného váčku
Hydrolýzou GTP se dynamin utáhne a odškrtí váček od membrány
Specifita klathrinových váčků
- Vyplývá z existence specifických transportních signálů na molekulách určených k přepravě a receptech v membráně, které k nim mají afinitu
- Receptory pro molekuly nákladu jsou ve spojení s klathrinem prostřednictvím adaptinu
Typy váčků
- klathrinové s nejméně dvěma typy adaptinu
- Váčky COP („coat proteins“ - coatomer COPI, COPII)
Pohyb váčků
- Na krátké vzdálenosti (ER - Golgi): prostá difúze
- Delší vzdálenosti (Golgi – plazmatická membrána u velkých buněk): využití motorových proteinů pohybujících se podél vláken cytoskeletu
Rozeznání cílové struktury
- Zprostředkováno molekulárními značkami, které transportní váčky nesou na svém povrchu
- Označují původ váčku a náklad
- Tyto značky musí být rozeznány receptory na cílových membránách
- Klíčovými složkami jsou transmembránové proteiny SNARE
- Proteiny SNARE váčků (v-SNARE; vesicle) interagují s komplementárními proteiny SNARE na povrchu cílových membrán (t-SNARE; target)
- Každá organela má svou typickou variantu proteinu SNARE
Splynutí membrán
SNARE zajišťují rozpoznání, nikoliv fúzi
Ke splynutí membrán nedochází okamžitě po navázání váčku
Požadováno velmi těsné přiblížení
Z hydrofilního povrchu membrány musí být odstraněna voda (energeticky nevýhodné)
Fúze katalyzována zvláštními proteiny, které se musí seskupit do komplexu a zajistit energeticky náročnou fúzi
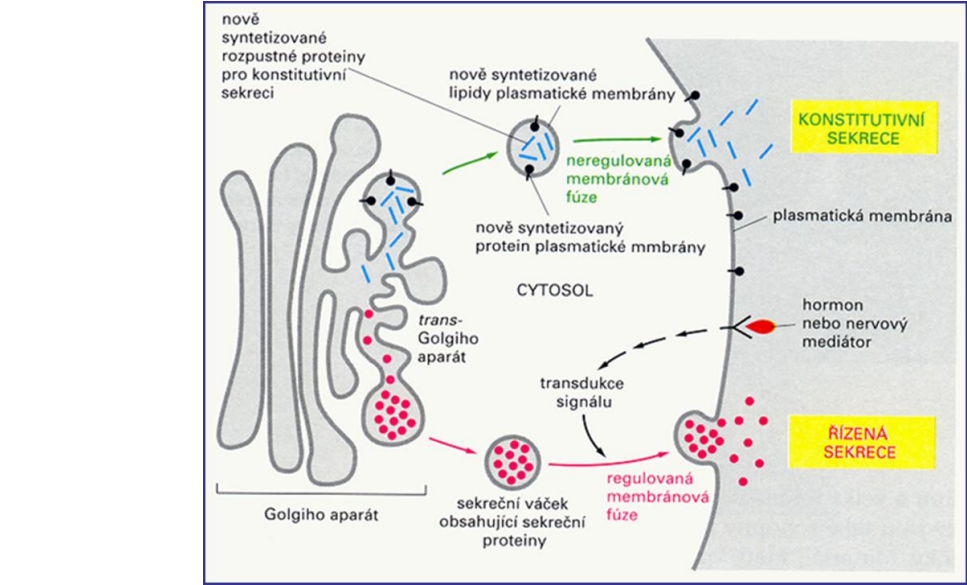
Sekreční dráha
Zajišťuje uvolnění materiálu do mimobuněčného prostředí (exocytózu)
Proteiny, lipidy a sacharidy určené k sekreci procházejí drahou ER Golgiho aparát – plazmatická membrána
Průběžná modifikace přenášených molekul, např. v lumen ER dochází díky oxidaci postranních řetězců cysteinu k tvorbě disulfidových můstků (v redukujícím prostředí cytosolu k této modifikaci nedochází)
V Golgiho aparátu – proteolýza, glykosylace..
Následkem je vyšší stabilita molekul a odolnost ke změnám pH ve vnějším prostředí
Řízená sekrece z buňky
funguje jen v buňkách zaměřených na sekreci (např. hormonů, trávicích enzymů, atd.)
Transportní váčky se zvětšují v sekreční váčky
Sekreční váčky se shromažďují v blízkosti plazmatické membrány
Přenášená molekula se uvolní jen po přijetí signálu
Signál je zachycen receptorem
Přijetí signálu vyvolá exocytózu
Konstitutivní (neřízená) sekrece
Neustálý proud váčků, které pučí z Golgiho aparátu a plazmatickou membránou (základní dráha exocytózy)
Dodává nově tvořené lipidy a proteiny do plazmatické membrány
Zodpovídá za růst plazmatické membrány před buněčným dělením
Sekretované proteiny přilnou k membráně z vnějšku nebo se stávají součástí extracelulární matrix
Běžná u všech buněk eukaryot
Endocytické dráhy
- Způsob příjmu kapaliny spolu s molekulami z vnějšího prostředí
- Pro malé molekuly využívají všechny eukaryotické buňky
- Příjem velkých částic nebo celých buněk zajišťují specializované fagocytické buňky
Průběh endocytózy:
- Obklopení pohlcovaného materiálu plazmatickou membránou
- Vchlípení membrány dovnitř buňky spolu s pohlcovaným materiálem
- Odškrcení membrány a vznik nitrobuněčného endocytického váčku
- Předání pohlceného materiálu lysosomům, kde je stráven
- Metabolity vzniklé trávením v lysosomu se přenášejí do cytosolu, kde je buňka může dále využít
Jaké typy endocytózy existují?
- Dle velikosti endocytických váčků
- Fagocytóza: trávení velkých částic, např. mikroorganismů a zbytků buněk prostřednictvím velkých váčků zvaných fagosomy
- Pinocytóza: pohlcování kapaliny a molekul malými váčky
Fagocytické buňky pohlcují velké částice
Ochrana proti infekci
Zajištují např. makrofágy, neutrofily
Mechanismus fagocytózy:
Na povrch infekční částice se vážou protilátky
Infekční částice s navázanými protilátkami se váže na povrch fagocytické buňky a aktivuje ji
Následkem je tvorba listovitých výběžků plazmatické membrány – pseudopodií, které částici pohltí
Vzniká fagosom
Lysosomy: organely pro buněčné trávení
Membránové váčky s hydrolytickými enzymy
Zajišťují řízené nitrobuněčné trávení původně mimobuněčných materiálů a opotřebovaných organel
Lysosomy: trávení zajišťují enzymy
Obsahují asi 40 typů hydrolytických enzymů (kyselé hydrolázy) odbourávajících proteiny, nukleové kyseliny, oligosacharidy a fosfolipidy
Optimální aktivita v kyselém prostředí (pH 5), které je udržováno uvnitř lysosomů – bezpečnost pro cytozolové struktury
Kyselé pH udržuje vodíková pumpa poháněná ATP, která do lysosomu čerpá H+
Složky lysosomální membrány jsou chráněny před vlastními enzymy glykosylací
Lysosomy odbourávají materiál z různých zdrojů
Větší extracelulární částice jsou přijímány fagosomy, které s lysosomy fúzují
Extracelulární kapalina je přijímána menšími endocytickými váčky, které je předávají lysosomům přes endosomy
Lysosom někdy tráví i složky vlastní buňky (autofagie)
Jaký je význam SRP v procesu transportu proteinů?
SRP navádí ribozom k membráně ER.
Co je úlohou Golgiho aparátu v buňce?
Úprava, třídění a transport proteinů.
Jaký je proces fagocytózy?
Pohlcování velkých částic makrofágy a neutrofily.
Jaké enzymy se nacházejí v lysosomech?
Hydrolytická enzymy (kyselé hydrolázy), odbourávající biomolekuly.
Jak se v buňkách transportují proteiny?
Pomocí transportních váčků nebo translokátorů.
Jaký je úkol membránové pumpy v lysosomech?
Udržovat kyselé pH pro optimální aktivitu enzymů.
Co je autofagie?
Proces, kdy lysosomy tráví vlastní složky buňky.
Jak se proteiny importují do mitochondrií?
Přes proteinové translokátory.
Jaké signály jsou potřebné k transportu váčků?
Specifické transportní signály na povrchu váčků.
Jak funguje exocytóza?
Uvolnění sekrečních vesikul z buňky po signalizaci.
Jaké je rozdělení organel v prokaryotických buňkách?
Nemají specializované oddíly, cytosol je jediným oddílem.
Jaký je úkol chaperonů v transportních procesech?
Pomoc při obnovení konformace proteinů uvnitř organel.
Jak se udržuje identita organel?
Každá organela má specifické proteiny a lipidy v membráně.
Kde se nachází signální sekvence na proteinech pro ER?
Na N-konci proteinu.
Jak se přenáší metabolity z lysosomů do cytosolu?
Po trávení se uvolňují do cytosolu, kde jsou dále využity.
Shrnutí
Vnitřní prostor eukaryotických buněk je bohatě strukturován
Do každého oddělení buňky se transportují specifické proteiny
Dopravu zajišťují transportní váčky nebo proteinové translokátory
Materiály přijímané z mimobuněčného prostředí jsou přenášeny k lysosomům, kde jsou podrobeny enzymatickému rozkladu