Transmission genetics
genetic processes that occur within individuals and how genes are passed from one individual to another
Molecular genetics
molecular structure of heredity, DNA and how biochemical process of the cell transfer genes to phenotype
Quantitative genetics
transmission of traits in large groups of individuals, where traits are simultaneously determined by many genes
Population genetics
transmission of traits in large groups of individuals, where traits are determined at one or a few genetic loci. Studies the changes in genetic patterns of populations in time and space. study the gene pool of a population, NOT the genotypes of individuals.
Population
: a group of individuals (usually large) that occur in the same place and are of the same species. Individuals in a population interbreed and share a common set of genes.
Gene pool
The genes shared by individuals in a population-some alleles may be very common, but others can be quite rare
frequency
a proportion (genotype or alleles) that always ranges between 0 and 1
Scarlet tiger moth frequency example
So the proportion of genotype (or allele) A + proportion of genotype (or allele) B = 1
To calculate the genotype frequency, simply count the number of individuals with a given genotype and divide by total number of individuals
Scarlet tiger moths: 452 BB, 43 Bb, 2 bb
f(BB) = 452/497 = 0.909
f(Bb) = 43/497 = 0.087
f(bb) = 2/497 = 0.004
TOTAL = 1.0
why is allele frequency important?
In sexually reproducing organisms, alleles not genotypes are carried on chromosomes during meiosis. There are always fewer alleles than genotypes, so it is easier to track population changes.
allele frequency- gene counting
1. Numbers of genotypes- gene counting
Allele frequency = number of copies of a given allele/
sum of counts of all alleles in population
Example: if we have a population of 1,000 sexually reproducing, diploid ladybugs
Let's say 300 AA, 500 Aa, and 200 aa
Frequency of A = (2 X # of homozygotes = 600) + (# of heterozygotes = 500) = 1100
Divide by total number of alleles (2000) to get 1100/2000 = 0.55
allele frequency- genotype frequency
2. Genotype frequency
Example: if we have a population of 1,000 sexually reproducing, diploid bugs
Let's say 300 AA, 500 Aa, and 200 aa
f(AA) = 300/1000 = 0.30
f(Aa) = 500/1000 = 0.50
f(aa) = 200/1000 = 0.20
p = f(A) = (frequency of AA homozygote) + (1/2 frequency of Aa heterozygote)
0.30 + 0.25
p = f(A) = 0.55
q = f(a) = (frequency of aa homozygote) + (1/2 frequency of Aa heterozygote)
0.20 + 0.25
q = f(a) = 0.45
NOTICE that p + q = 1
Multiple allele example
Let's say we have three alleles, A1, A2, and A3
Again, we can use gene counting to get the answer:
p = f(A1) = (2 x # of A1A1) + (# of A1A2) + (# of A1A3)
(2 x total number of individuals)
q = f(A2) = (2 x # of A2A2) + (# of A1A2) + (# of A2A3)
(2 x total number of individuals)
r = f(A3) = (2 x # of A3A3) + (# of A1A3) + (# of A2A3)
(2 x total number of individuals)
Notice in these formulas that we only include a heterozygote if it includes the allele we are calculating
Snapdragon example
100 individual snapdragons
25 red
50 pink
25 white
Calculate the genotype and allele frequencies for each allele
Genotype frequency: simply count the number of individuals with a given genotype and divide by total number of individuals
f(AA) = 25/100 = 0.25
f(Aa) = 50/100 = 0.50
f(aa) = 25/100 = 0.25
Allele frequencies can be calculated from genotype frequencies:
p = f(A) = (frequency of AA homozygote) + (1/2 frequency of Aa heterozygote)
q = f(a) = (frequency of aa homozygote) + (1/2 frequency of Aa heterozygote)
p = 0.25 + 0.25 = 0.50 q = 0.25 + 0.25 = 0.50 p + q = 1
genetic drift
random fluctuations in the frequencies of alleles or haplotypes, often leading to some alleles being fixed (100% frequency)- it is a form of non-adaptive evolution, and is a consequence of chance
KEY CONCEPT: smaller the population, bigger the effect of drift
genetic drift example
Example: large, finite population of A1, single new mutant A2
Parental mating: A1A1 X A1A2
Probability that that one surviving offspring is A1A1 = ½
Probability that two surviving offspring are A1A1 = ½ X ½ = ¼
Probability that A2 is lost by chance immediately = ¼
Probability that A2 is lost after 127 generations = 98.5%
KEY CONCEPT: smaller the population, bigger the effect of drift
why do we care about genetic drift?
Many genetic diseases are recessive, we want to know the likelihood of carriers (we only see diseased and affected)
Also, want to know the total of genetic diversity in a population
Allows us to track genetic changes (i.e., evolution) of a population over time and space
The Paradox of Brachydactyly
The geneticist Reginald Punnett told mathematician G. H. Hardy about the problem of brachydactyly in the British
Punnett asked why the frequency of brachydactyly wouldn't increase with every generation if it is a dominant trait
Hardy came up with a formula to explain why bracydactyly allele frequency stays stable over generations, or reaches equilibrium
Key to his formula is the assumption that natural selection does not favor one phenotype (brachydactyly vs. normal) over another
A few weeks after Hardy published his formula, a German physicist named Wilhelm Weinberg published the same idea
Brachydactyly
a genetic defect causing shortening of the fingers, and it is a dominant trait
Assumptions of Hardy-Weinberg Principle
-Mating is random (for locus in question), or panmictic
-The population is infinitely large, and thus genetic drift is not likely
-Genes are not added from outside the population (gene flow or migration)
-Mutation does not occur
-All individuals have equal probabilities of survival and reproduction
If we find a population is NOT in Hardy Weinberg equilibrium, one or more of these assumptions is violated
Symbols for allele frequency:
Freq of A (dominant allele) = p
Freq of a (recessive allele)= q
First, p + q = 1
Therefore p = (1 - q)
In our simple example
p = 0.5 and therefore, q = 0.5
Derivation of Hardy-Weinberg Formula
Population genotype frequencies reaches Hardy-Weinberg equilibrium after 1 generation of random mating.
1 = p2 + 2pq + q2
Note we can also derive HW from:
(p + q)2
Probability of the homozygous dominant genotype:
product of the two dominant allele frequencies
(the AND rule with probability)
Probability of the heterozygous genotype:
either a recessive sperm and a dominant egg
OR a recessive egg and a dominant sperm
(p X q) + (p X q) = 2pqal
Probability of the homozygous recessive genotype:
product of the two recessive allele frequencies
allele frequencies
1 = p + q
p = f(A)
q = f(a)
Concealed genetic variation
when homozygous recessive alleles are very rare (because of dominant allele), almost all of the carriers are heterozygotes
Sometimes a dominant allele can be rare in a population
If HW assumptions are met, then:
Allele frequencies will not change
Genotype frequencies will stabilize at p2:2pq:q2 (HW equilibrium) AFTER ONE GENERATION
Subsequent generations will remain at HW equilibrium
What will genotype and allele frequencies of offspring look like IF mating is random?
If mating is random, offspring allele frequencies do not change from that of parents, and are thus at Hardy-Weinberg equilibrium
Genotype frequency stabilizes at HW equilibrium after 1 generation- must compare HW values to original genotype frequencies
NOTE: probability is mathematical prediction; frequency is observed rate in the population
Note that genotype frequencies sum to 1, and allele frequencies sum to 1
We have a population of 2,000 Gaboon vipers. A genetic difference for venom toxicity exists at a single locus, and the alleles have incomplete dominance.
If we observe 100 individuals homozygous for the t allele (tt = low toxicity), 800 heterozygotes (Tt = medium toxicity), and 1,100 homozygous dominant (TT = if you get bit you die).
A) What is the frequency of the t allele in the population?
B) Are the genotypes in Hardy-Weinberg equilibrium?
A) Allele frequency of t = (2 x tt homozygotes) + (Tt heterozygotes) / (total number of alleles in population)
(2 x 100) + (800) / 4000 = 0.25 = q
B) If q = 0.25, then p = 1 – 0.25 = 0.75
So p2 = (0.75)2 = 0.5625
2pq = 2 x 0.75 x 0.25 = 0.375
q2 = (0.25)2 = 0.0625
Genotype frequencies:
TT = 1100/2000 = 0.55
Tt = 800/2000 = 0.40
tt = 100/2000 = 0.05
A Χ2 test says no HW equilibrium, but it’s close
We have a population of 3800 dragons- if they are homozygous recessive for the u allele, they are red- heterozygotes and UU are normal green dragons.
If we observe 2000 red dragons homozygous for the u allele, 1000 heterozygotes (Uu), and 800 homozygous dominant (UU), tell me:
A) What is the frequency of the u allele in the population?
B) Are the genotypes in Hardy-Weinberg equilibrium?
A) Allele frequency of u = (2 x uu homozygotes) + (Uu heterozygotes) / (2 x number of allele copies in population)
(2 x 2000) + (1000) / 7600 = 0.6579 = q
B) If q = 0.6579, then p = 1 – 0.6579 = 0.3421
So p2 = (0.3421)2 = 0.1170
2pq = 2 x 0.3421 X 0.6579 = 0.4501
q2 = (0.6579)2 = 0.4328
Genotype frequencies:
UU = 800/3800 = 0.2105
Uu = 1000/3800 = 0.2632
uu = 2000/3800 = 0.5263
HW frequencies are different from initial genotype frequencies, so the dragons are NOT in HW equilibrium
Allopatric
populations that do not overlap, and are often separated by a distance
Sympatric
populations that overlap each other
Parapatric
populations that do not overlap but are just adjacent to each other (not separated by a distance)
Hybrid zone
a region in which genetically distinct parapatric populations (often species) interbreed
Cline
a gradual change in a character or allele frequencies over a geographic distance- often indicative of adaptive geographic variation
Reinforcement
Hybrids are less fit than either pure breed species. The species continue to diverge until hybridization can no longer occur.
Fusion
Reproductive barriers weaken until the two species become one.
Stability
Fit hybrids continue to be produced.
Bottleneck
a circumstance that decreases the size of a population
Founder effect
a special type of bottleneck in which a small number of individuals establishes a new population. The new population will experience genetic drift because of small population size, and may lose some rare alleles from the original population
The smaller the founder bottleneck, the faster heterozygosity decreases
After numerous generations, the population size increases, and mutation and recombination restore heterozygosity
Coalescence
Concept that all gene copies in a population are derived from a common ancestor
The smaller the population, the less time required for coalescence
IMPORTANT: By chance, a FINITE population (violation of HW) will eventually become monomorphic for one allele. The probability of a neutral allele (no selective advantage to phenotype) becoming fixed is equal to its initial frequency.
Deme
small independent populations of a species
metapopulation
several proximate demes
Random walk
equally likely consecutive changes that result in fixation or loss of an allele
Random fluctuation example
p = 0.5 in a deme, and by chance, becomes 0.47 in next generation. In next generation will become higher or lower- but both outcomes equally likely
Some demes will either lose or become fixed for an allele- this probability increases with time
Important Concepts of Genetic Drift
Allele or haplotype frequencies fluctuate randomly in a deme, and eventually one allele or haplotype will become fixed
Because of #1, genetic variation at a locus declines with time, the frequency of heterozygotes [H = 2p(1–p)] declines, and this decline can be a proxy for genetic drift in the population
Let’s say p = 0.5, then H = 2 X 0.5 (1–0.5) = 0.5
But then, if p = 0.8, then H = 2 X 0.8 (1–0.8) = 0.32
An allele’s probability of becoming fixed equals its frequency at that time, regardless of past changes in its frequency
Demes (or populations) with the same initial allele frequency (p) undergo random walk, where a proportion (p) will become fixed for one form of the allele, and a proportion of others (1–p) become fixed for the alternate allele
The frequency of a new mutation that is represented by one gene copy in a population with 2N gene copies is:
pt = 1/2N = probability of p becoming fixed
notice as population size (N) increases, pt decreases
As seen in #5, genetic drift occurs faster in smaller populations. In a diploid population, the average time to fixation of a new, neutral mutant allele (assuming it becomes fixed) is 4N generations
Among demes in a metapopulation, the average allele frequency ( p )
changes little (small number of demes) or not at all (large number of demes), but as allele frequencies in each deme become 0 or 1 over time, the frequency of heterozygotes (H) declines to 0 in each deme, AND in the metapopulation as a whole
Census size (aka absolute population size)
actual number of adults in a population
Effective population size (Ne)
number of individuals in an ideal population (one where every adult mates) in which the rate of genetic drift (measured by decline in heterozygosity) would be the same as it is in the actual population
Example: 10,000 adults in a population, but only 1,000 breed, then genetic drift proceeds at same rate as a population of 1,000 (Ne)
As Ne decreases, genetic drift and loss of heterozygosity increases
This is bad because it can lead to decreased viability and/or inbreeding depression
Effective Population Size can be reduced in 5 ways:
Variation in the offspring produced by females, males, or both
A sex ratio that differs from 1:1
Natural selection increases variation in offspring of certain phenotypes
Example: Selection favors larger individuals, so smaller individuals contribute less to gene pool (complete set of unique alleles in a population)
If generations overlap, probability of inbreeding increases
Fluctuations in population size, especially when population sizes are small
Inbreeding depression
the reduction of fitness (survival and/or reproductive output) resulting from an increase in deleterious homozygous recessive alleles in inbred individuals- big problem with endangered species captive breeding programs
Genetic rescue
introduction of new genes to inbred population
Inbreeding coefficient (F)
measure of probability of two alleles being identical by descent. Ranges from 0 (random mating) to 1 (all alleles identical by descent)
what occurs during inbreeding?
When inbreeding is occurring, heterozygote proportion decreases by 2Fpq, and half of this value is added to the proportion of each homozygote
what happens when population size increases?
As population size (N) increases, genetic drift has less of an effect, whereas the mutation rate (µ) has more of an effect
Thus heterozygosity increases with mutation rate and population size
FST = fixation index
measures variation in a locus for two alleles among populations- ranges from 0 (no variation among populations- lots of gene flow) to 1 (populations fixed for different alleles- no gene flow)
If Nm = number of immigrants per generation,
Nm = (1/FST) – 1 If Nm = 1 (one breeding individual per 4 population is an immigrant), then FST = 0.20.
So, even a little gene flow keeps heterozygosity high
Remember the rate at which genetic drift causes alleles to become fixed is inversely proportional to the effective population size (Ne) BUT, genetic drift can be counteracted by gene flow
Earth Forms
Our solar system and Earth formed at the same time, about 4.6 billion years ago
Earth formed by collision of smaller bodies into one larger one, releasing huge amounts of heat
As Earth cooled, a solid crust formed, gases were released (little oxygen), formed water vapor
Oceans and protocontinents by 4 bya
Large land masses 3 bya
First Prokaryotes
First fossils of life are stromatolites (layered mounds of bacteria) 3.5 billion years old
In 2016 possible stromatolites from Greenland up to 3.9 bya, BUT in 2018, fossils became doubtful
Little oxygen in atmosphere, so first bacteria were probably anaerobic
Around 2.5-3 bya, photosynthetic cyanobacteria evolved, started increasing oxygen in atmosphere
Origin of Eukaryotes
Eukaryotic organisms may have arisen 2.7 bya, but first fossils are 1.5 bya
Have a nucleus with chromosomes, mitotic spindle, undergo meiosis for sexual reproduction
Most eukaryotes remained unicellular for a billion years after their origin, but…
Many have mitochondria or chloroplasts
Endosymbiont Theory
Eukaryotic cells started out as anaerobic organisms in primordial Earth
1-1.5 bya, a eukaryotic cell engulfed a photosynthetic bacteria without killing it, and a symbiotic relationship occurred
As Earth’s atmospheric oxygen levels rose, the bacteria lost its photosynthetic properties, but continued to provide the cell with biochemical advantages- led to mitochondria
Why are chloroplast and mitochondria independent?
Chloroplasts in plant cells evolved in a similar way to the endosymbiont theory
Chloroplasts and mitochondria are now dependent on their cell homes, and vice versa
Mitochondria and chloroplasts are ancient descendants of ingested bacteria. This is why they have separate genomes.
Mitochondria
organelles involved in cellular respiration, which converts biochemical energy from nutrients to ATP
ATP
is a molecule of stored energy that can be used for several crucial cellular processes
Oxidative phosphorylation
the electron transport chain system of the mitochondria transport ions to the intermembranal space, creating a gradient
What processes make ATP?
When ions are allowed to flow down the gradient, ATP synthase creates ATP from ADP + phosphate
Substrate-level phosphorylation (Krebs Cycle): also makes ATP through a series of chemical reactions in the matrix of mitochondria
Mitochondrial Genome
Most mitochondrial genomes are circular, double-stranded and supercoiled DNA, but no histone proteins
Linear mt genomes in protozoa (often unicellular eukaryotes) and fungi
One mitochondrion can contain several copies of the genome in multiple nucleoid regions- can be many mitochondria per cell
The genome in plants, animals and fungi is similar in the type of genes, but genome size can vary widely- humans about 20kb but some plants are 2 million
mt Genome Replication
Replication of mtDNA is semiconservative like it is in the nuclear genome
Unlike genomic DNA, mtDNA can replicate at any phase of the cell cycle
Bases not equally distributed on both strands- heavy and light strands because one strand will have many purines (two rings- heavy) and other will have complementary pyrimidines (one ring- light)
A gene called ori for origin of replication opens “D loop” region where H strand undergoes synthesis first, followed by L strand
Mitochondrial Genes
Most proteins of mitochondria are encoded by nuclear genes, but some on mt itself
Ribosomal proteins are coded by nuclear genes, but ribosomal RNA is coded by the mitochondria
Transcription is unusual because a single RNA molecule is made, and cut into smaller pieces
tRNA’s are cut out of the single transcript by specific enzymes, freeing the mRNA and rRNA
No 5’ caps on mitochondrial mRNA, but a poly-A tail is added to the 3’ end as in nuclear mRNA
No introns in animal mtDNA, but they are found in yeast and plants
Translation in Mitochondria
Mt start codons are close to the 5’ end, which have no leader sequences
A special tRNA called fMet-tRNA is used to initiate protein synthesis- also unique mt initiation factors, elongation factors and release factors
Only plant mitochondria use the universal genetic code of the nuclear genome
Animal and fungal mitochondria have a slightly different code for translation
Genetic Relationships via mtDNA
mtDNA can be used to trace familial relationships in humans
Different mt genes can be used to reconstruct the evolutionary history of many different organisms
Phylogenies from mtDNA are revolutionizing evolutionary biology
Chloroplast Genomes
Organelles for site of photosynthesis in plants, green algae and photosynthetic protists
Like mtDNA, cpDNA is double-stranded, circular, without proteins, and supercoiled- multiple copies of genome in each chloroplast nucleoid
Genome size is between 80kb and 600 kb
Gene organization is similar to mtDNA- genes for chloroplast rRNA, tRNA’s, and some genes for photosynthesis, transcription and translation, but many proteins of chloroplast come from nuclear DNA
Translation is similar to system we studied for prokaryotes- code is standard, universal code
Summary of mtDNA and cpDNA:
Both organelles are co-dependent with eukaryotic cells:
Mitochondria make ATP for cell energy, but require proteins made in cytoplasm to function
Chloroplasts make ATP (light reactions) and sugars (dark reactions) during photosynthesis for plant cells- like mitochondria they require proteins made in cytoplasm to exist
Mitochondria have their own modified genetic code, but chloroplasts use standard genetic code
Mitochondria are inherited maternally (most cases), chloroplasts maternally in flowers, other plants can be paternally
Rules of Non-Mendelian Inheritance
Mendelian ratios are not found with extranuclear genes because meiosis-based segregation does not occur
Reciprocal crosses in eukaryotes are not the same because inheritance is biased towards one sex and there is no meiosis-based segregation
Uniparental inheritance: genes inherited through mtDNA or cpDNA are the same in all progeny as ONE of the parents- usually this is maternal inheritance because mother’s cytoplasm in her gamete usually exceeds father’s sperm
Extranuclear genes can not be mapped to the chromosomes of the nucleus because they are not inherited in the same way
Non-Mendelian inheritance is not affected by substituting a nucleus with a different genotype
Why don’t male mitochondria contribute?
the “midpiece” section of the sperm contains huge numbers of mitochondria (or one large one in some species) that provide ATP for the tail, or flagellum
During fertilization, the acrosome releases enzymes that allows one single “winner” sperm to enter the egg
The nucleus contains the genetic material needed to make the zygote- only the head of the sperm is drawn into the cytoplasm of the egg
Maternal inheritance
The genotype of mom is what is important
-Many maternally inherited traits are influenced by extranuclear genes
Mitochondrial inheritance
Sometimes cytoplasmic DNA/RNA
Mitochondria are in the egg and none of the mitochondria come from Dad
species
a population or a group of populations whose members have the potential to interbreed in nature and produce viable, fertile offspring.
Criterion
Potential reproductive compatibility
Prezygotic Isolation
Prezygotic barriers to fertilization between two species
the separation of different species to keep them from creating offspring by preventing the gametes from forming a zygote.
Habitat isolation, Temporal isolation, Behavioral isolation, Mechanical isolation, Gametic isolation
Postzygotic Isolation
barriers that occur after zygote formation such as organisms that die as embryos or those that are born sterile
Example: Donkey x Horse
= Sterile Mule
reduced hybrid viability, reduced hybrid fertility, hybrid breakdown
Does reproductive isolation cause speciation?
NO- reproductive isolation & speciation are a by-product of genetic changes that occur during changes of populations for other reasons
So what processes drive speciation?
Genetic changes within populations as a consequence of:
Genetic drift and/or mutation
Natural selection
Polyploidy and hybridization
Nictitating membrane
useful to everything from sharks to cats, but vesitigial (lacrimal caruncula) in humans
Charles Darwin (1809–1882): Theory of Evolution
All species have descended (with modification) over [a LONG] time from one or a few common ancestors through the process of natural selection. THE unifying theme to explain the diversity of life on Earth.
when does evolution occur?
Evolution only occurs when there is a change in gene frequency within a population (a group of organisms in the same species) over time.
These genetic differences are heritable and can be passed on to the next generation—which is what really matters in evolution: long term change.
descent with modification
genetic changes in a population over time
Common Ancestry
different species share a common ancestor, and ALL life can be traced back to one common ancestor
allopatric speciation
the evolution of genetic reproductive barriers between populations that are geographically separated by a physical barrier
Most prevalent mode of speciation in animals
Populations that begin process of speciation in allopatry can come back into contact, and either interbreed freely at hybrid zones, or not at all
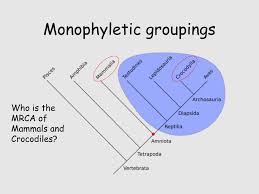
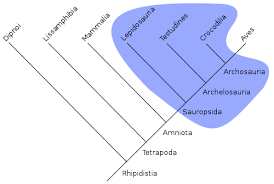
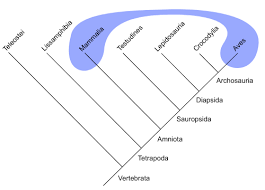
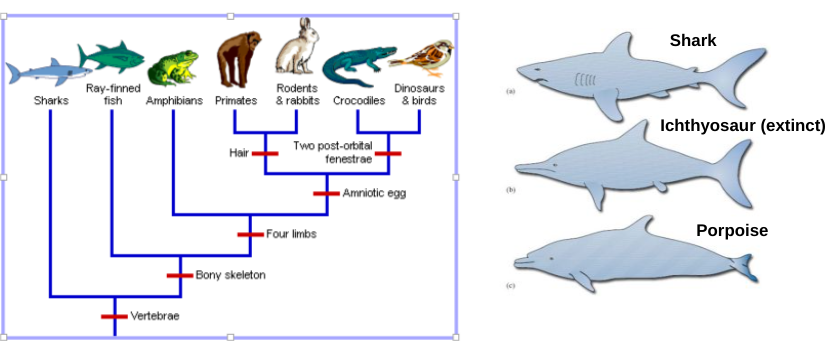
Clade
monophyletic group
node
common ancestor
branch
lineage
monophyletic
paraphyletic
polyphyletic
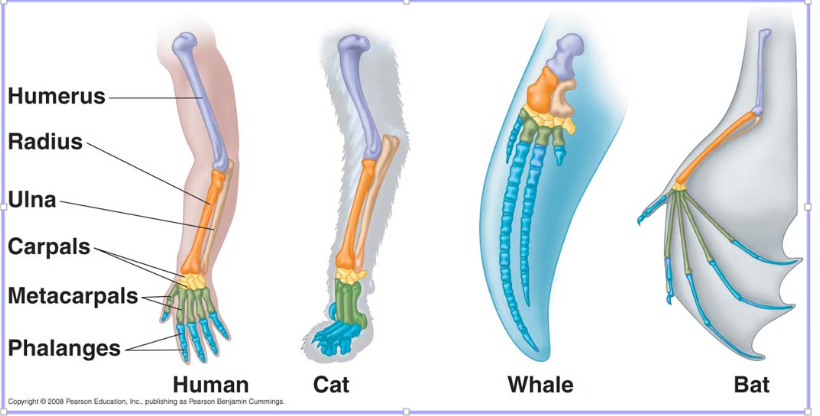
Homology
a character state shared by taxa without modification from a common ancestor
Homoplasy
a character state that has independently evolved two or more times, or similarity that is NOT from common ancestry (often convergent evolution)
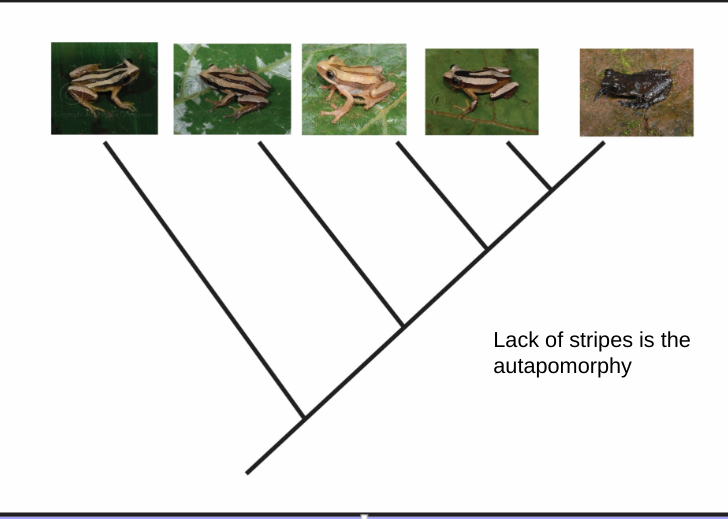
Autapomorphy
derived character state present in a single lineage
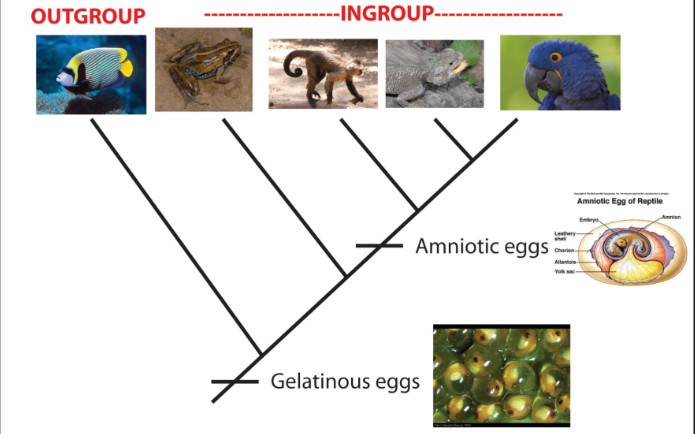
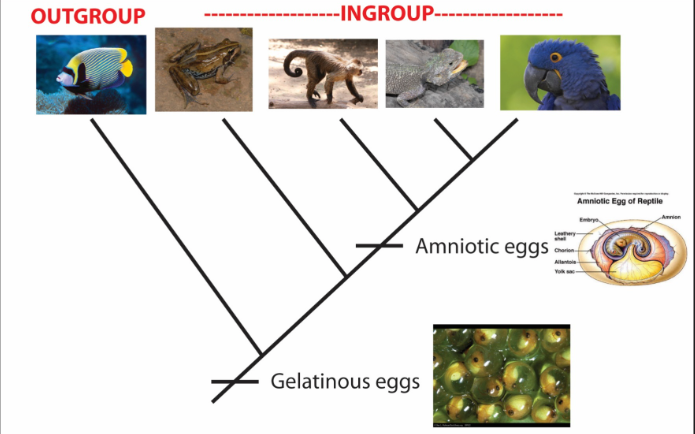
Plesiomophy
ancestral character state (e.g., gelatinous eggs)
Synapomorphy
a homologous character state that supports the monophyly of a group through common ancestry (e.g., amniotic eggs)
Genetic changes in closely related species
Small changes in sequences such as
base changes and small deletions
Duplications of genes and DNA
Rearrangements on all scales
Acquisition of foreign DNA and genes
Loss of DNA and genes
Notice in our DNA comparison that alignment of homologous characters is key
Homologous sequences
Descended from same ancestral sequence through DNA replication across generations.
Usually for our purposes if there is “significant” similarity we assume that the sequences are from a common ancestor
This is why BLAST works for us
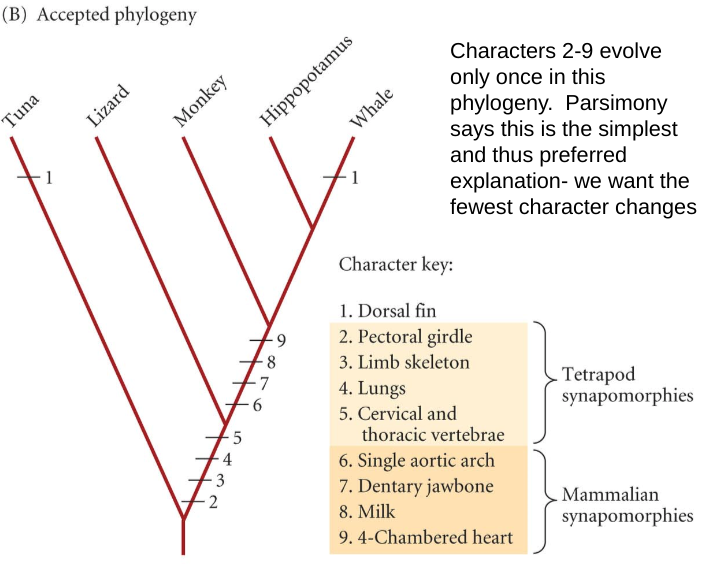
Principle of Parsimony, or Occam’s Razor
the simplest explanation is preferred
Maximum Parsimony
all mutations are equally likely
The problem… With DNA sequence data, Maximum Parsimony weights all character state changes (mutations) as equal, but in reality, they are biased
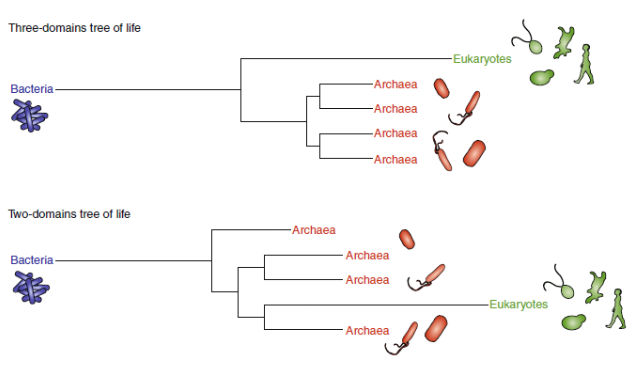
Long-branch attraction
Species with many autapomorphies will be chosen as sister groups, when in reality they are not
How many domains of life are there?
New study from December 2019 suggests there are only two domains of life, and that previous results supporting 3 domains of life were misguided by long branch lengths