Week 9: Species Abundance Distribution + The Latitudinal Diversity Gradient (LDG)
1/64
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced |
|---|
No study sessions yet.
65 Terms
Biological structure of a community
the mix of species, including both their number and relative abundance
Species richness
number of species that occur within a community
Relative abundance
percentage each species contributes to the total number of individuals
Rank- abundance plot
graphical way to show relative abundance
Species evenness
equitable distribution of individuals among species
Why are large carnivores/ predators rare?
loss of energy/ inefficient transfer→ differences between trophic levels
a histogram showing the number of individuals per species on the x axis (arithmetic scale) against the number of species in that class on the y axis
hollow curve pattern
few common species → many species have a low abundance
many rare species → few species have a high abundance
one of ecologies universal laws
Rank-abundance plot (Whittaker plot)
most common = rank 1
least common = rank n (n = number of species)
Log Series
Fisher R. A. et al. 1943
Ronald Fisher → analysed species-abundance
moths → light traps
hollow curve pattern can be seen
uneven distribution of abundance
larger alpha = more evenly distributed abundance
N+ alpha on slide not N- alpha
According to the log-series, singleton species are the modal class

What did Frank Preston argue?
log-series like distribution is a sampling artefact due to failure to sample the rarest species
most datasets represent samples rather than complete enumerations
Preston, F.W. (1948) The commonness, and rarity, of species. Ecology, 29, 254–283.
Log normal
Preston, 1948
Plotting well sampled communities with the x-axis on a log-scale reveals an intermediate peak
modal class of intermediate abundance: log-normal
normally distributed on log scale + sampling enough→ log normal
octaves
each class double the number as before
species consisting of 1 individual less than species consisting of 2-3
Preston veil line
More recent study backing up log normal distribution
Conolly et al (2005) Community Structure of Corals and Reef Fishes at Multiple Scales, Science 309: 1363
increase in sampling → log normal distribution emerges
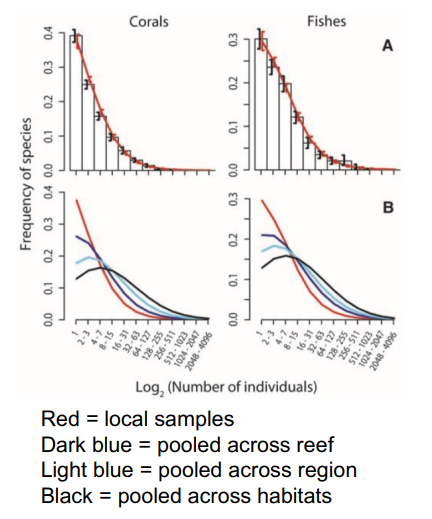
Variation in shape of SAD in tropical communities vs. temperate
tropical rain forest trees look log -normal but look at whole rainforest → hollow curve
tropics → more species, more evenly distributed
opposite to temperate zones
tropical forest trees on completely sampled plots log-normal like
understory plants in Irish conifer plantation log-series like
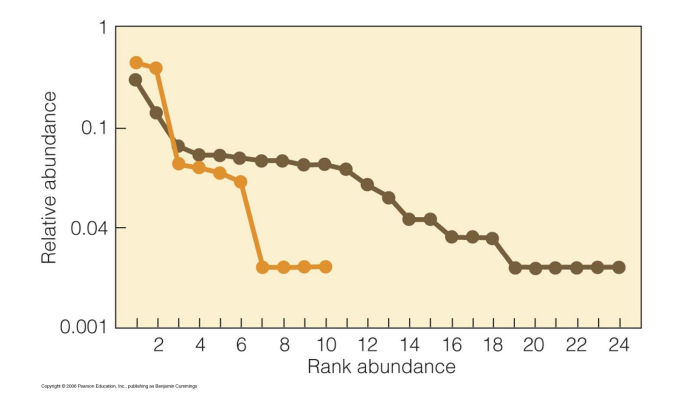
What did Alroy show in a 2015 paper?
Alroy 2015 The shape of terrestrial abundance distributions. Science Advances
variation in both richness and evenness among communities
tropical communities (black) have shallower rank abundance plots -> more even distribution of abundance
temperate communities (grey) dominated by a few species
What did Anne Magurran show in Magurran & Henderson (2003) Nature?
more than one pattern
Estuarine fish, Bristol, 21 years
Discontinuity between persistence and maximum abundance
Ever present core species: Log-Normal
1 group → always there, high abundance → log modal
Transient: Log-series
other group→ not consistently there → log series
Potential Explanations
abundance of rare species controlled by random dispersal events
other group → controlled by availability of resources
Why does detecting abundance matter?
impacts of human activity on biodiversity
abundance in disturbed vs. undisturbed
Link between disturbance and SAD
Hill et al. (1995) J. Applied Ecology, 32754-760
Butterfly species across forest transects in Indonesia
undisturbed: Log-normal like (solid symbols)
disturbed: Log-series like (open symbols)
disturbed → more uneven distribution
Estimating Diversity of Floral Species in the Amazon basin
ter Steege, H. et al. 2013 Hyperdominance in the Amazonian Tree Flora
Estimates of local tree abundance from 1195 plot samples: sample includes ~5,000 species
Compared 16 different statistical SAD models: log-series fit best
Use log-series to predict the unsampled species as well as their abundance
Estimated 16,000 species across the Amazon basin
227 (1.4%) account for half of all individual trees
The rarest 5800 species have estimated population sizes of <1000→ species may never be discovered
Why is it important to estimate the floral diversity in the Amazon?
important finding as forests are carbon sinks
→ conserve the few 100s of tree species + understand response to environmental change for most of the forest
What processes explain the shape of SADs?
Central Limit Theorem
Niche apportionment models
Neutral theory
Example of population growth being determined by the multiplicative (synergistic) effect of different resources
Elser, J. J., et al. 2007. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecology Letters
Addition of both nitrogen and phosphorus results in higher plant productivity than expected based on the effect of each resource in isolation
What is the central limit theorem?
May, R. M. 1975. Patterns of species abundance and diversity
The multiplicative effect of many independent variables acting on population growth would lead to a ‘lognormal’ SAD
Nekola and Brown 2007 The wealth of species: ecological communities, complex systems and the legacy of Frank Preston
The abundance of many things in nature, not just species abundance, show lognormal patterns
Niche apportionment models
Tokeshi (1990) Niche apportionment or random assortment: species abundance patterns revisited
The local abundance of a species depends on the amount of resources available to that species
Relative abundance thus reflects resource division among species •
Imagine a resource pool (we might call this ‘niche space’) utilized by a species. A new species invades the community and takes a fraction of these resources, and this process is repeated
Leads to a ‘hollow curve’ pattern
Many different ways of theoretically dividing niche space that lead to different shaped SADs
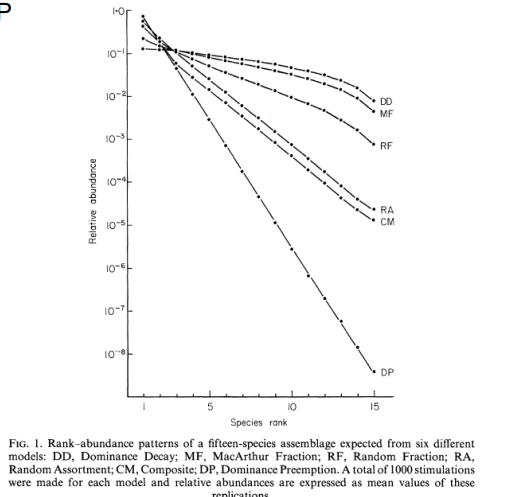
Dominance pre-emption (DP) model
Each species arrives and takes up a constant proportion of the remaining resources
K=0.5
First ranked 50% of total abundance/biomass
Second ranked 50% of remaining 50% = 25%
Third ranked = 12.5% of total abundance
Generates a ‘geometric series’ distribution
Characterized by a straight line (with slope k) on the rank-abundance diagram
why 50%
→ no theoretical explanation for the fraction of resources species should take
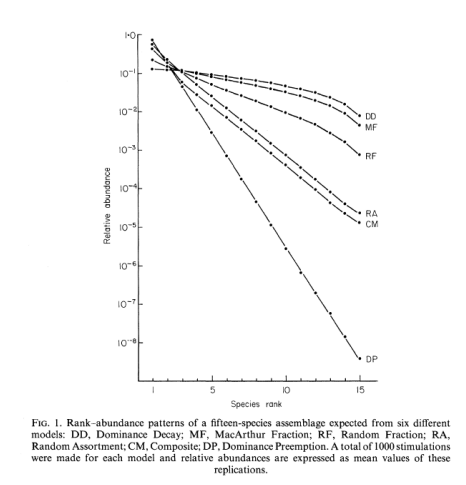
Dominance decay (DD) model
Dominance decay (DD) is the inverse of DP
top line of the graph
It is always the largest niche that is divided
Results in a more even distribution
Random assortment (RA) niches are selected at random and split at random
Intermediate level of evenness
log-normal like
How did neutral theory begin?
The unified neutral theory of biodiversity and biogeography
started as a theory in genetics →most mutations neither deleterious/ selected for
no positive + negative effect → explanation for huge variation
Rosindell et al 2001 The unified neutral theory of biodiversity and biogeography at age ten.
What did Stephen Hubbell (2001) argue?
too many tree species to be explained by the partitioning of niche space given all plants depend on a few limiting resources (light, water, phosphorus, nitrogen)
How can so many species co-exist? (Stephen Hubbell)
niche partitioning → are there so many different niches?
Tree species coexist, not because their niches are different, but because the have equal fitness (ability to grow + reproduce)
similarity in fitness allows them to co-exist → one species can’t out-compete another because same fitness
How does neutral theory work e.g within a tree community?
‘Zero sum game’ i.e. total number of individuals in local community and in the region is fixed. As one individual dies it is replaced by another (of the same or different species)
All individuals, regardless of species identity, are identical in terms of their ecology and fitness and have equivalent probabilities of death, producing an offspring or dispersing i.e. the model is neutral at the individual level
These random processes cause ‘ecological drift’, as the abundance of each species randomly fluctuates over time
Eventually each species will drift to extinction until there is only one species remaining
Individuals randomly mutate to become new species (speciation), balancing extinction -> dynamic equilibrium
Zero sum game
plot with fixed number of individuals → zero sum game
Shape of abundance distribution: Neutral Theory
J (size of regional pool)
v (speciation rate)
Number of sites (size of local community)
θ = 2Jv
Small ~ geometric series of abundances
Medium ~ log-Normal distribution
Large ~ flat (even) distribution
Fits to rain forest data
Comparing neutral and niche models
Volkov et al. 2003 compared niche (log normal model) and neutral model in predicting the abundance of trees on Barro Colorado island, Panama
Neutral model (green) fit better than the niche (black) model
Relative abundance of species due to neutral processes?
But similar predictions despite fundamentally different assumptions
Abundance patterns alone insufficient to resolve the processes structuring communities
What should happen if abundance is set by niches?
Ricklefs and Renner (2012)
if abundance is set by niches then families of trees abundant on one continent should also be abundant on other continents (if we assume they have retained similar niches)
Continents have been separated for tens of millions of years and so these correlations are not predicted by the random drift in abundance that occurs in the absence of niches (i.e. neutral theory)
Stronger tests of niche and neutral models
Harpole & Tilman (2006), Non-neutral patterns of species abundance in grassland communities
Neutral + niche models provide a similar fit to the relative abundance of plant species in the Cedar Creek experiment
Neutral theory predicts that abundance is independent of the niche
Harpole and Tilman (2006) found a –ve relationship between R* and biomass
R* = level to which species reduce soil nitrate when grown in monoculture
Lower R* = Higher competitive ability
Is neutral theory a sufficient explanation for patterns in species abundance?
probably not
But the theory highlighted the importance of regional processes (speciation, dispersal) and chance events in structuring communities
Resulted in stronger interrogation of empirical data and realization that the shape of SADs alone are not sufficient to discriminate between different theories
One hectare of tropical rainforest can contain…
400 tree species (Kraft 2008
900 plant species (Balslev 1998)
More than all tree species in Europe
~16,000 tree species in Europe
What is the Latitudinal Diversity Gradient (LDG)
Most species on Earth live in the tropics
The number of species found in a given area declines from the tropics to the poles
This pattern is repeated across many different types of organisms, from plants to birds, and ants to fish
the first order biodiversity pattern
Historical Observations of LDG
The much greater biological diversity of the tropics compared to temperate zone was clear to early naturalists, like Darwin, Bates, Humboldt and Wallace
Bates (1892) wrote of collecting more than 700 species of butterflies within an hour's walk of his home in Para, Brazil (compared to ~60 resident species in the UK)
Papers showing LDG
Economo et al. 2018 → LDG in ants
Davies et al. 2008 → LDG in mammals
Papers showing LDG in aquatic animals
Collen et al. 2014. Global patterns of freshwater species diversity, threat and endemism
freshwater biodiversity = highest in amazon basin
Tittensor et al. 2010. Global patterns and predictors of marine biodiversity across taxa
marine organisms → highest diversity = coral triangle
What drives these large scale gradients in biodiversity?
No comprehensive answers
Mix of abiotic factors (e.g. energy, heterogeneity, stability, area), chance (e.g. mid-domain effect), historical events (e.g. ice ages) and biotic interactions (e.g. competition)
Why is it hard to find out what causes LDG?
can’t easily do (manipulative) experiments on large scales
long to wait for results e.g of evolutionary processes driving these gradients
just have a single planet → can’t compare with other places
Theory: Energy Availability
Curvature of the Earth results in greater solar energy at the tropics, driving higher net primary productivity (production of plant biomass) where water is not limiting
Plant species richness strongly predicted by AET (the quantity of water (mm/yr) removed from a surface by evaporation and transpiration)
Statistically explains >70% of the variation in richness
higher input of energy cascades up food chains to support more species at higher trophic levels
Currie, D. J. (1991)
AET
Actual evapotranspiration
The quantity of water that is removed from a surface due to the processes of evaporation and transpiration
Correlation between bird species richness and AET
Storch et al 2007
positive correlation
energetic limits to species richness
lots of energy → lots of food
support a higher density of individuals
More individuals hypothesis (MIH)
The local density and species richness of birds increases with energy availability across N America and Europe
With a higher total number of individuals, rarer species are supported at a higher abundance, reducing rates of stochastic extinction
Higher energy → more individuals → more species
Mönkkönen, Forsman and Bokma, (2006)
Exceptions that prove the rule: Endothermic mammals and ectothermic marine predators
looking at smaller taxonomic levels → counter example e.g penguin species richness
can be informative when thinking about the mechanisms driving the patterns
Ectothermic marine predators show expected LDG, but richness of endothermic birds and mammals peaks at high latitudes
metabolic asymmetry
Metabolic asymmetry
at high latitudes species able to maintain a higher body temperature can attain higher swimming speeds making them better able to capture sluggish ectotherm prey, increasing energy availability to endotherms
if ability of other fish/ prey you are catching declines = zero sum game
endotherms → can maintain body temp + swimming speed across different water temps
Grady et al 2019 Metabolic asymmetry and the global diversity of marine predators
Exceptions that prove the rule: brittle star species richness
Shallow water species: strong LDG predicted by temperature gradients. Higher temperatures provide more kinetic energy for prey capture
Deep water species: uniform temperature. Peak in richness at high latitudes and continental margins where more chemical energy
More individuals hypothesis (MIH): Manu National Park, Peru vs. Hubbard brook, USA
Terborgh et al 1990
Species richness: 4-5 x higher (160 species at a single point)
Biomass: ~5 x higher (190kg/km2 )
Abundance: almost identical! (1920 vs ~2000 individuals km2)
Perhaps MIH only explains richness gradients at low-intermediate levels of energy availability?
larger amounts of energy supporting higher biomass → supporting larger birds not higher density of birds
The Mid-Domain Effect
If species geographic ranges were randomly placed in a bounded domain then richness would be expected to peak at the centre of the domain ‘the pencil box effect’
Has been suggested as a possible null model for the LDG
but species have to arise randomly on the surface of the Earth for this to work → speciation more likely in some places than others
Colwell and Lees (200) The mid-domain effect: geometric constraints on the geography of species richness
Why mountain passes are higher in the tropics?
Janzen, 1967
the tropics highlands are inhospitable throughout the year for a lowland species (and vice versa) whereas at high latitudes species can cross these barriers during some seasons
temperate regions → summer/ winter → habitats moving up and down mountain → don’t get strong geographic isolation
Thermal niches
Tewksbury, Huey & Deutsch 2008 Ecology. Putting the heat on tropical animals
Stable temperatures enable greater specialisation of species thermal niches
climate stable in tropics → species evolved to have narrow niches
temperate species operate over larger temperature ranges
Example of non-random speciation: Lupinus
Hughes and Eastwood (2006) Island radiation on a continental scale: Exceptional rates of plant diversification after uplift of the Andes
idea that climatic stability → leads to species narrow niches that are stratified at different elevations and that promotes geographical isolation and speciation
Lupinus, diverse genus of plants. Andean species arose in the last ~1.5 million years
Given rise to 81 species, amongst the fastest rates of speciation ever inferred.
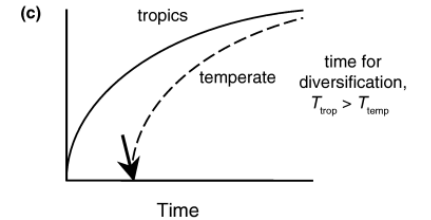
Theory: More time for evolution in the tropics
For most of Earth’s history the climate has existed in a greenhouse state, substantially warmer than the present
Cold environments, now found at high latitudes, have arisen relatively recently
last 60 million years = progressive cooling
cold environments only ‘recently’ originated → over the last 40-50 million years
Less time for species to accumulate there?
Latitude correlated with multiple factors – energy and time. Challenge of only having a single planet and not being able to perform experiments
But we can test the effect of time using phylogenetic data
‘Tropical niche conservatism hypothesis’
Wiens and Donogue 2004 Historical biogeography, ecology and species richness
Species inherit their niche from their ancestor
tropics older → ancestors of clades occurred in tropics
It takes time for species to adapt and spread into new climates
colonsing new latitudes hard → evolve new traits / evolutionary innovation - takes time
e.g. herbaceous growth, deciduous leaves, and narrow water-conducting cells are adaptations to freezing tolerance in plants.
Evidence supporting tropical niche conservatism hypothesis
Economo et al 2018 Macroecology and macroevolution of the latitudinal diversity gradient in ants
Phylogenetic tree of ~15000 ant species and subspecies
Ants arose in the tropics >80mya and only spread out of the tropics <40mya
Species have accumulated at similar rates – no evidence for differences in rates of speciation or extinction
ant richness increasing at similar rate in extratropical regions compared to tropics but starting later
LDG in the fossil record
Mannion, P (2020) A deep-time perspective on the latitudinal diversity gradient
The shape and strength of the LDG has varied over geological time e.g. relatively flat LDG for dinosaurs during Cretaceous
Modern LDG developed over last ~50 million years
Strong diversity peak in the tropics during ice-house periods and flattening during greenhouse periods?
but patchy fossil record, sampling bias
Inverse latitudinal gradient in speciation rates for marine fishes
Rabosky et al. 2018
Speciation rates not highest in tropics for fish → highest at high latitudes
Potential hypothesis
Rates of speciation in fish fastest a high latitudes where species richness is lowest
Perhaps tropical communities have reached a ‘carrying capacity’ and ecological niches are full, while the diversity of polar regions is still increasing to fill empty ecological niches
Which 3 fundamental processes control the number of species in a region?
Immigration
Speciation
Extinction
Diversification = Speciation - Extinction
The LDG must be driven by some combination of:
Differences in the rates of any, or all, of these processes
Differences in the time available for speciation (or immigration)
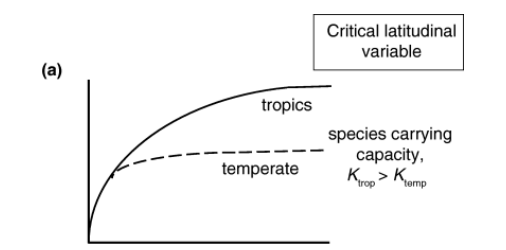
Differences in carrying capacity regulating these processes
Differences in carrying capacity
Negative feedback of standing diversity on diversification and or immigration. e.g. as diversity increases rates of speciation slow down or rates of extinction increase
The tropics can support more species at equilibrium
Equilibrium model.
Mittlebach et al 2007
Differences in rates
Faster rates of speciation, and/or slower rates of extinction and/or faster immigration into the tropics
Non-equilibrium/historical model
Mittlebach et al 2007
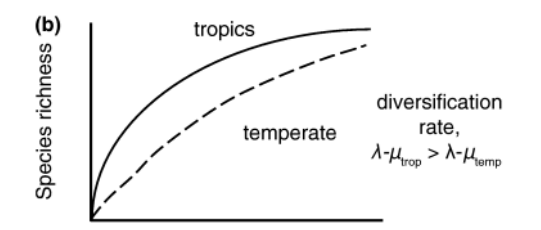
Differences in time
Tropical climates are older, providing more time for speciation
Non-equilibrium/historical model
Mittlebach et al 2007