Topic 8.1 response to the environment
1/44
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced |
|---|
No study sessions yet.
45 Terms
How are changes in environment detected
Receptors are specialised cells that can generate an electrical impulse in a sensory neurone when stimulated by a particular stimulus e.g. light receptors are stimulated when light falls on them
Receptor cells send signals via either the nervous system or the hormonal system to the body's co-ordination centres in the brain or spinal cord
Signals are then sent on to the parts of the body which respond, known as the effectors
The human nervous system
The human nervous system consists of
Central nervous system (CNS) – the brain and spinal cord
Peripheral nervous system (PNS) – all of the nerves in the body
The nerves connect the receptors in the sense organs with the CNS, and connect the CNS with effectors
The CNS acts as a central coordinating centre for the impulses that come in from, and are sent out to, any part of the body
Sympathetic vs Parasympathetic
The periferal nervous system (PNS) is split into the somatic nervous system (conscious control) and the autonomic nervous system (subconscious control)
The autonomic nervous system is divided into the sympathetic and parasympathetic nervous systems
Sympathetic Nervous System
Prepares the body for action ('fight or flight' response) and increases the heart rate during exercise
Parasympathetic Nervous System
Calms the body down after action ('rest and digest' response) and decreases the heart rate after exercise
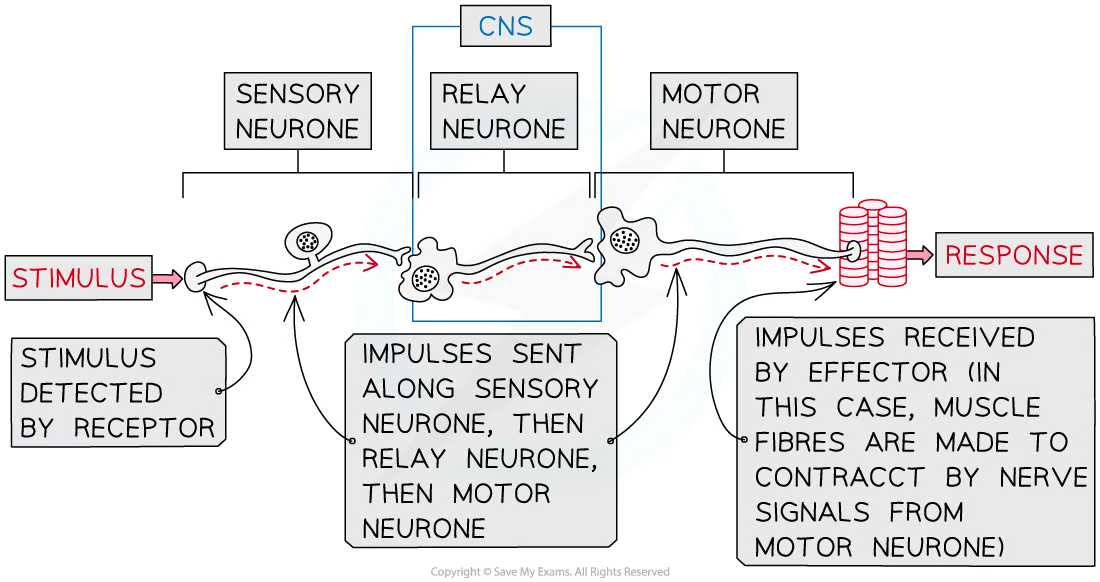
Nerve impulse pathway
stimulus > receptor > sensory neurone > CNS > motor neurone > effector
The hormonal system
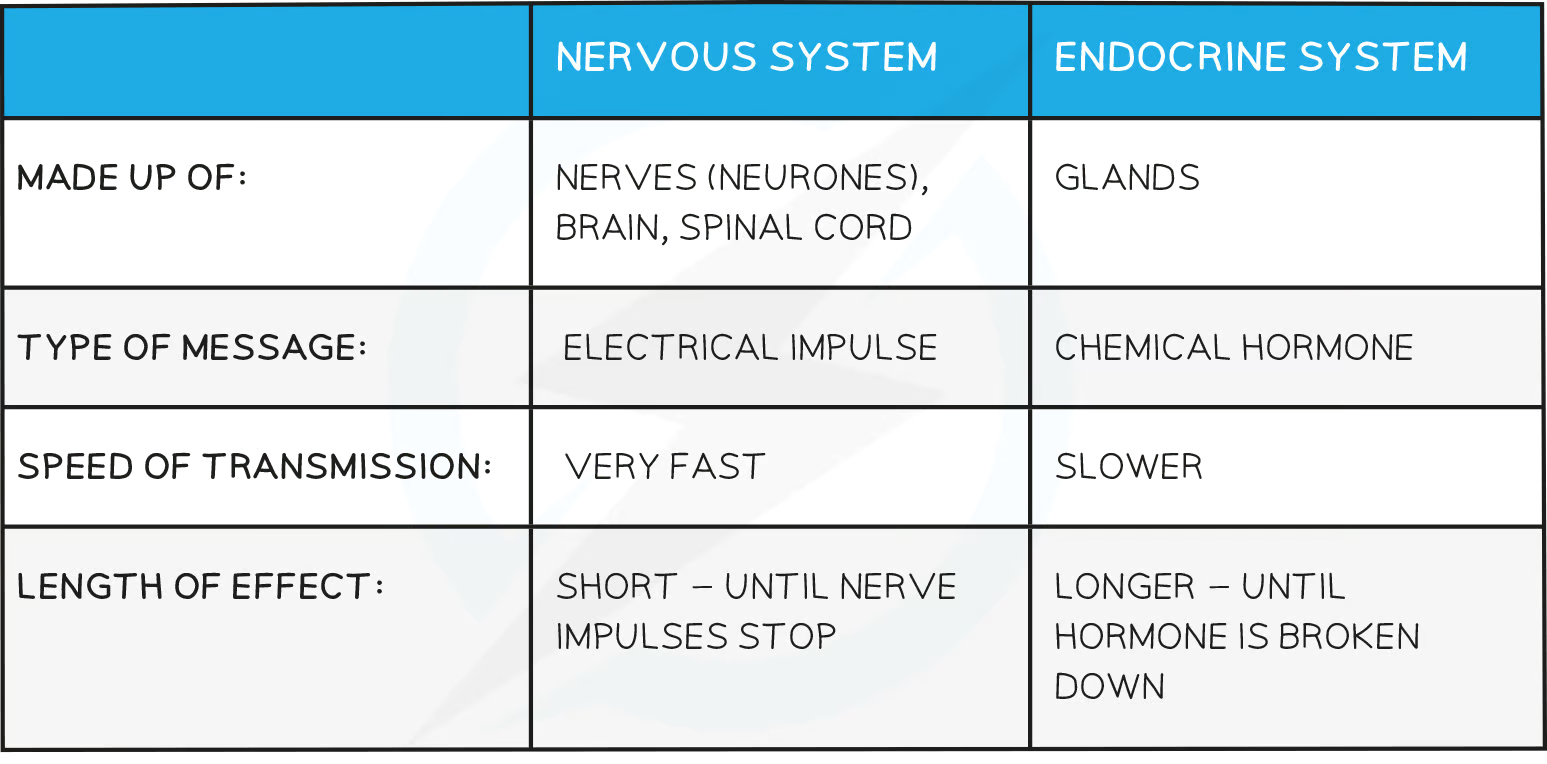
Hormones are chemical substances produced by endocrine glands and carried by the blood
Hormones transmit information from one part of an organism to another and bring about change by altering the activity of one or more specific target organs
Hormones are slower in action than nerve impulses and are therefore used to control functions that do not need instant responses
Endocrine system
Endocrine glands that produces hormones in animals are known collectively as the endocrine system
Endocrine glands secrete hormones directly into the blood
Endocrine glands can be stimulated to secrete hormones by the action of another hormone or by the arrival of a nerve impulse
Hormone pathway
stimulus > receptor > hormone > effector
nervous vs endocrine
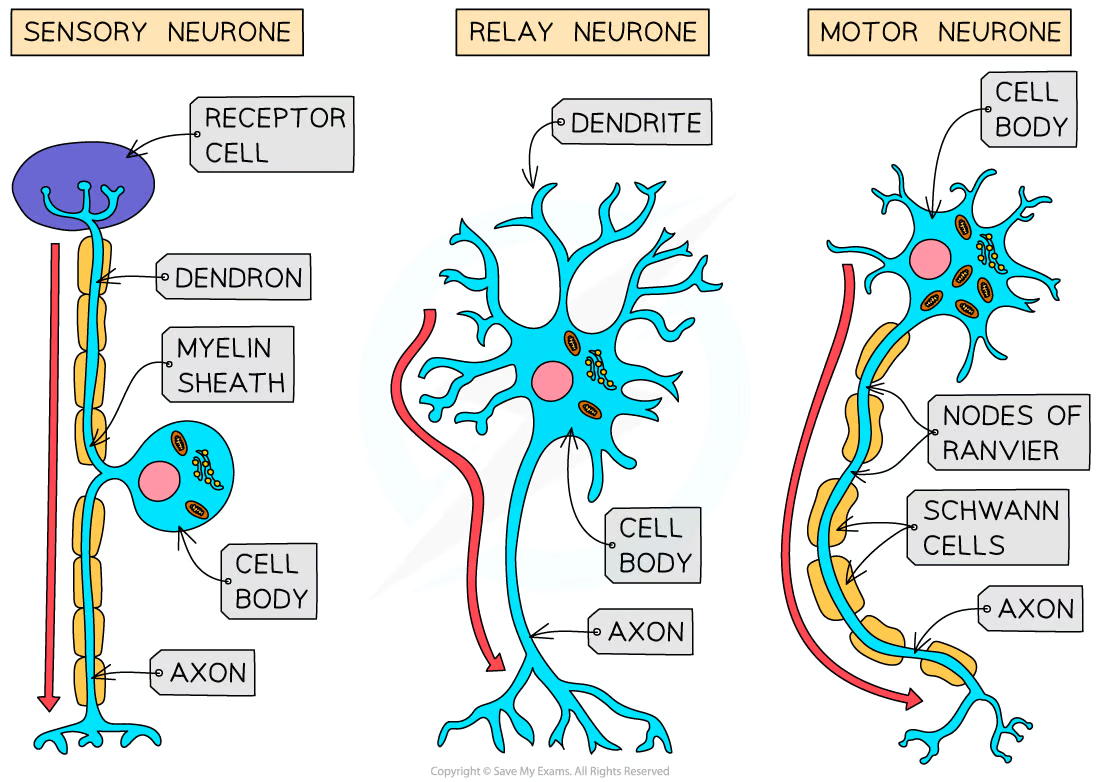
Neurones
A bundle of neurones is a nerve
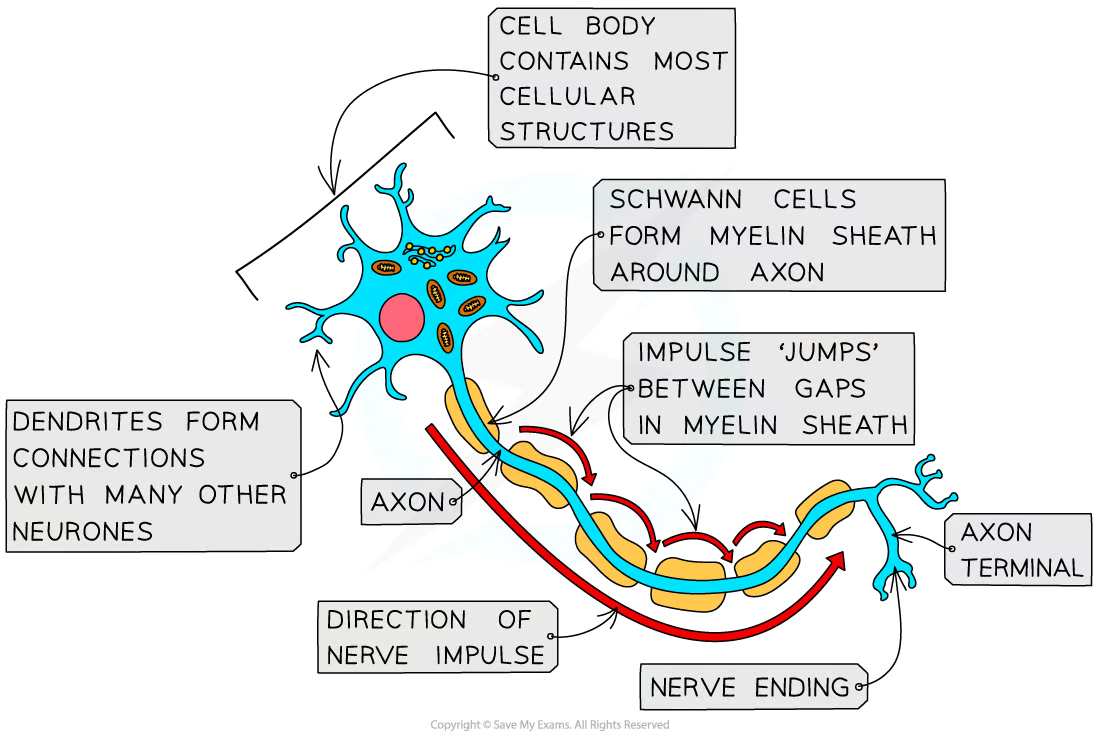
A neurone has
A long fibre known as the axon
A cell body that contains the nucleus and other cellular structures
axon terminal which has many nerve endings which allow neurones to connect to and receive impulses from other neurones, forming a network for easy communication
Myelinated neurones
Axon is insulated by a fatty layer known as the myelin sheath
Myelin sheath is made up of specialised cells called Schwann cells which wrap themselves around the axon
The uninsulated gaps between the Schwann cells are known as the nodes of Ranvier
Electrical impulses in myelinated neurones do not travel down the whole axon but instead jump from one node to the next, speeding up impulse transmission
Sensory neurones
Sensory neurones carry impulses from receptors to the brain and spinal cord in the CNS
A cell body that branches off in the middle of the axon
The dendrites are attached to a receptor cell
The section of neurone that links the dendrites attached to the receptor cell with the cell body is known as a dendron; it delivers the electrical impulse to the cell body
The section of neurone that connects the cell body with the CNS is the axon
Motor neurones
Motor neurones carry impulses from the CNS to effector muscles or glands
A large cell body at one end that lies within the spinal cord or brain
Many highly-branched dendrites extending from the cell body, providing many connections with the axon terminals of other neurones
Relay neurones
Relay neurones are found entirely within the CNS and connect sensory and motor neurones
Short neurones with axons and highly branched dendrites
Pupil response
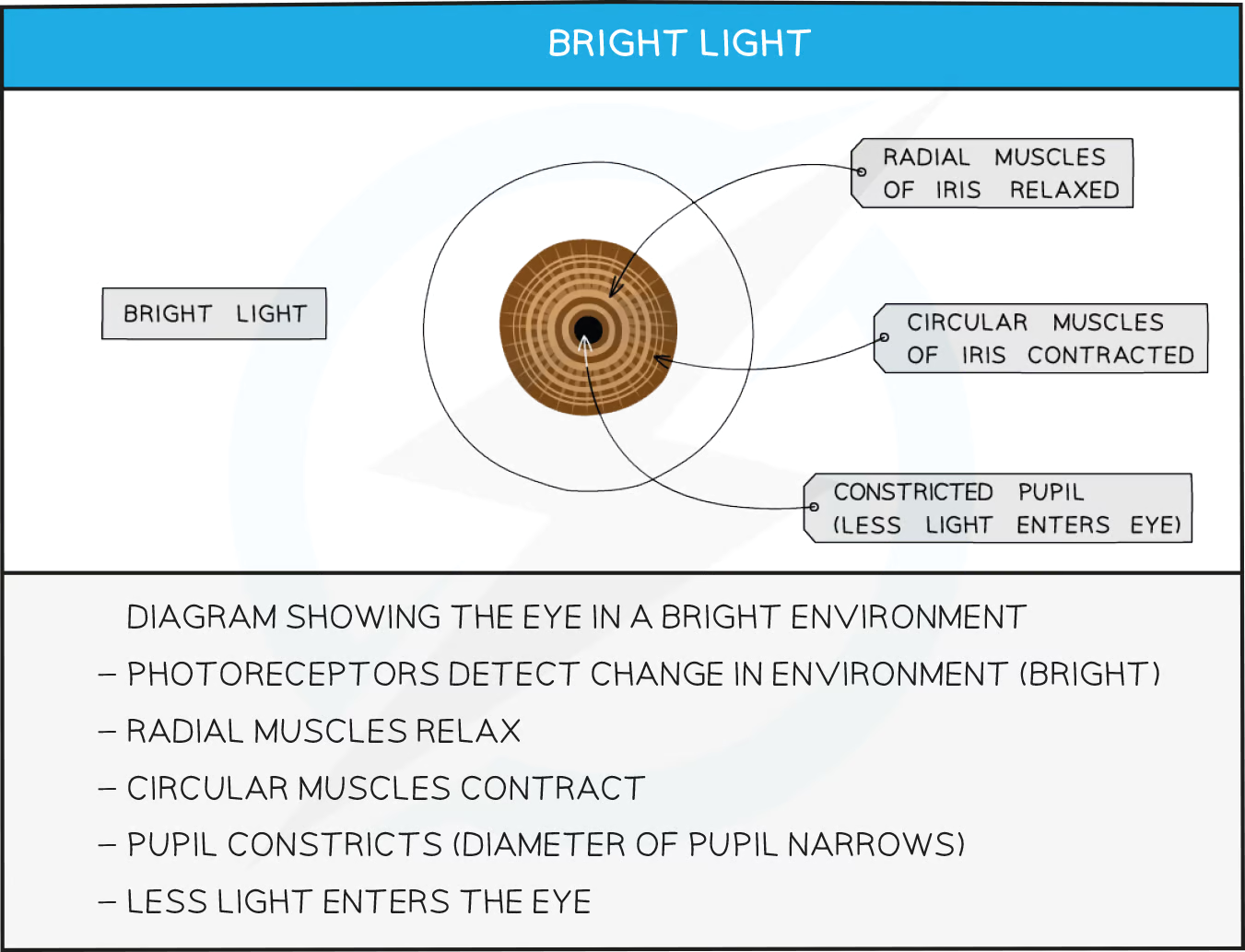
Changing pupil diameter enables the eye to control the amount of light hitting the retina
The diameter of the pupil in the eye is determined by two sets of muscles
The circular muscles contract to constrict the pupil
The radial muscles contract to dilate the pupil
The two sets of muscles work antagonistically, meaning that when one set of muscles contracts the other relaxes, and vice versa
Pupil response in bright light
bright light > light receptors in eyes > sensory neurone > CNS > motor neurone > circular muscles in iris contract
Contraction of the circular muscles in the iris of the eye causes the pupil to constrict
This limits the amount of light entering the eye and prevents damage to the retina
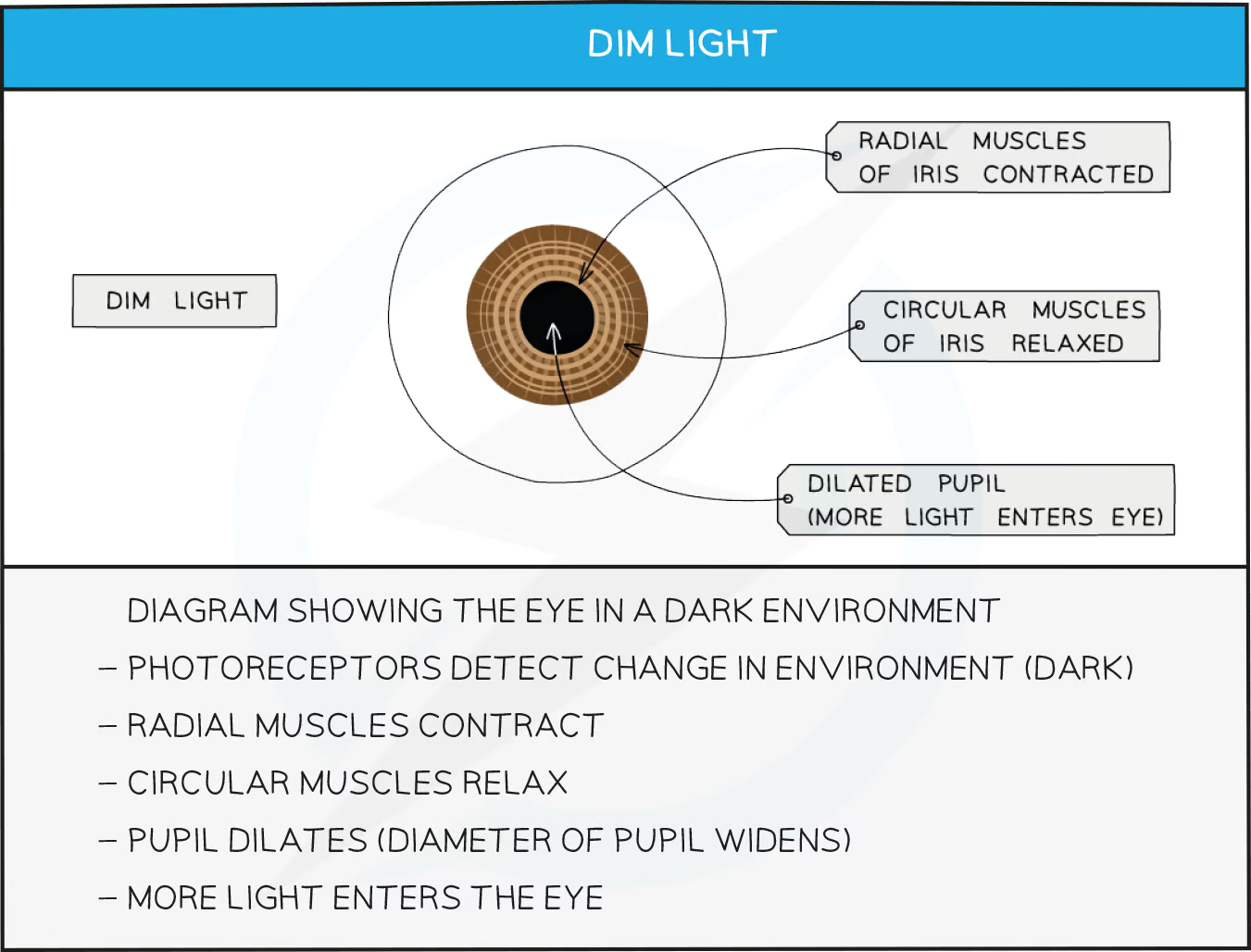
Pupil response in low light
low light > light receptors in eyes > sensory neurones > CNS > motor neurone > radial muscles in iris contract
Contraction of the radial muscles in the iris of the eye causes the pupil to dilate
This maximises the amount of light entering the eye, improving vision
resting potential
Sodium-potassium pumps use ATP to actively transport 2 potassium ions in to the axon for every 3 sodium ions pumped out of the axon.
This creates a concentration gradient across the membrane for both sodium and potassium ions
The neurone cell surface membrane has sodium ion channels and potassium ion channels to allow the two types of ions to move across the membrane by facilitated diffusion.
The neurone membrane is less permeable to sodium ions than potassium ions so potassium ions diffuse out at a faster rate than sodium ions can diffuse back in (more K+ leak channels than sodium)
The negative state in the axon produces an electrochemical gradient causing potassium ions to be attracted to the inside of the axon.
However the potassium ion concentration force is greater than the electrochemical force, which results in an overall movement of potassium ions out of the axon causing the outside to be more positive than inside.
When there is no further movement of potassium ions the potential difference across the axon is -70mV. The axon is said to be polarised
action potential (Depolarisation)
A stimulus causes the neurone membrane at one part of the neurone to increase in permeability to sodium ions.
Sodium ions begin to move into the axon down their concentration gradient
If the potential difference reaches around -55 mV, known as the threshold potential, sodium voltage-gated channels open leading to a further influx of sodium ions
Once the charge has been reversed from -70 mV to around +30 mV the membrane is said to be depolarised and an action potential has been generated
action potential (Repolarisation)
1ms after an action potential is generated all the voltage-gated sodium ion channels in this section of the membrane close
Voltage gated potassium channels in this section of axon membrane now open, allowing the diffusion of potassium ions out of the axon down their concentration gradient
If ion channels open this means the permeability of that part of the membrane to those ions (sodium/potassium) has increased
action potential (Hyperpolaristation)
Hyperpolaristation
There is a short period during which the membrane potential is more negative than resting potential; this is known as hyperpolarisation
The period during which the membrane is hyperpolarised is known as the refractory period
The membrane is unresponsive to stimulation during the refractory period, so a new action potential cannot be generated at this time
This makes the action potentials discrete events and means the impulse can only travel in one direction

transmission of an action potential
Once an action potential has been generated it can be propagated, or transmitted, along the length of the axon
The depolarisation of the membrane at the site of the first action potential causes sodium ions to diffuse along the cytoplasm into the next section of the axon, depolarising the membrane in this new section, and causing voltage gated sodium channels to open
This triggers another action potential in this section of the axon membrane
This process then repeats along the length of the axon
In myelinated neurons the sodium ions channels are located at the nodes of Ranvier as the sodium ions cannot diffuse through the myelinated parts of the axon.
Myelination and action potentials
By insulating the axon membrane myelin increases the speed at which action potentials can travel along the neurone
In sections of the axon that are surrounded by a myelin sheath membrane depolarisation cannot occur, as the myelin sheath stops the diffusion of sodium and potassium ions
Action potentials can only occur at the nodes of Ranvier
Nodes of Ranvier are the gaps between the Schwann cells that make up the myelin sheath
The membrane at the nodes of Ranvier depolarises when the sodium ions arrive
The diffusion of sodium ions in this way is known as local currents, or local circuits
The action potential therefore appears to ‘jump’ from one node to the next; this is known as saltatory conduction
Saltatory conduction allows the impulse to travel much faster than in an unmyelinated axon of the same diameter
preventing impulse transmission
Such drugs may bind to sodium ion channels, preventing them from opening and therefore preventing an influx of sodium ions when an axon is stimulated
Preventing sodium ion influx prevents membrane depolarisation and an action potential cannot be generated
all or nothing principle
An impulse is only transmitted if the initial stimulus is sufficient to increase the membrane potential above a threshold potential
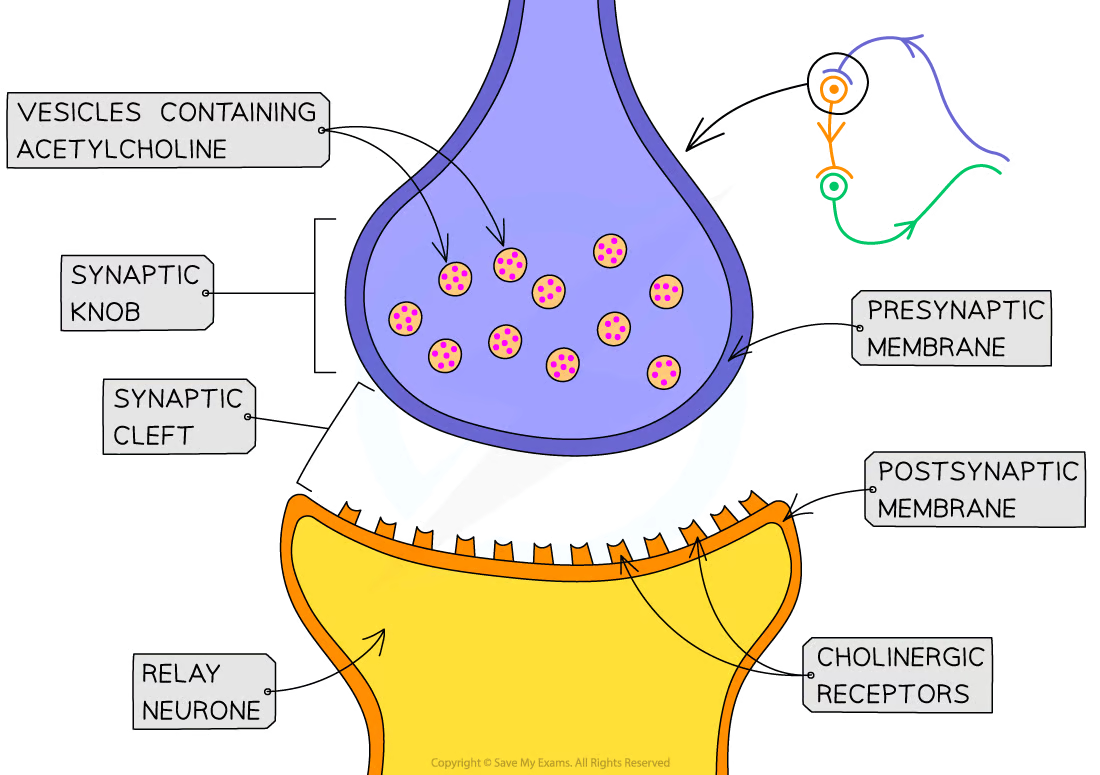
Synapses intro
Gap between the neurones is known as the synaptic cleft
presynaptic neurone has a rounded end known as the synaptic knob
Nerve impulses are passed across the synaptic cleft by the diffusion of chemicals known as neurotransmitters e.g. acetylcholine
Neurotransmitters are contained within vesicles in the synaptic knob
Synaptic transmission
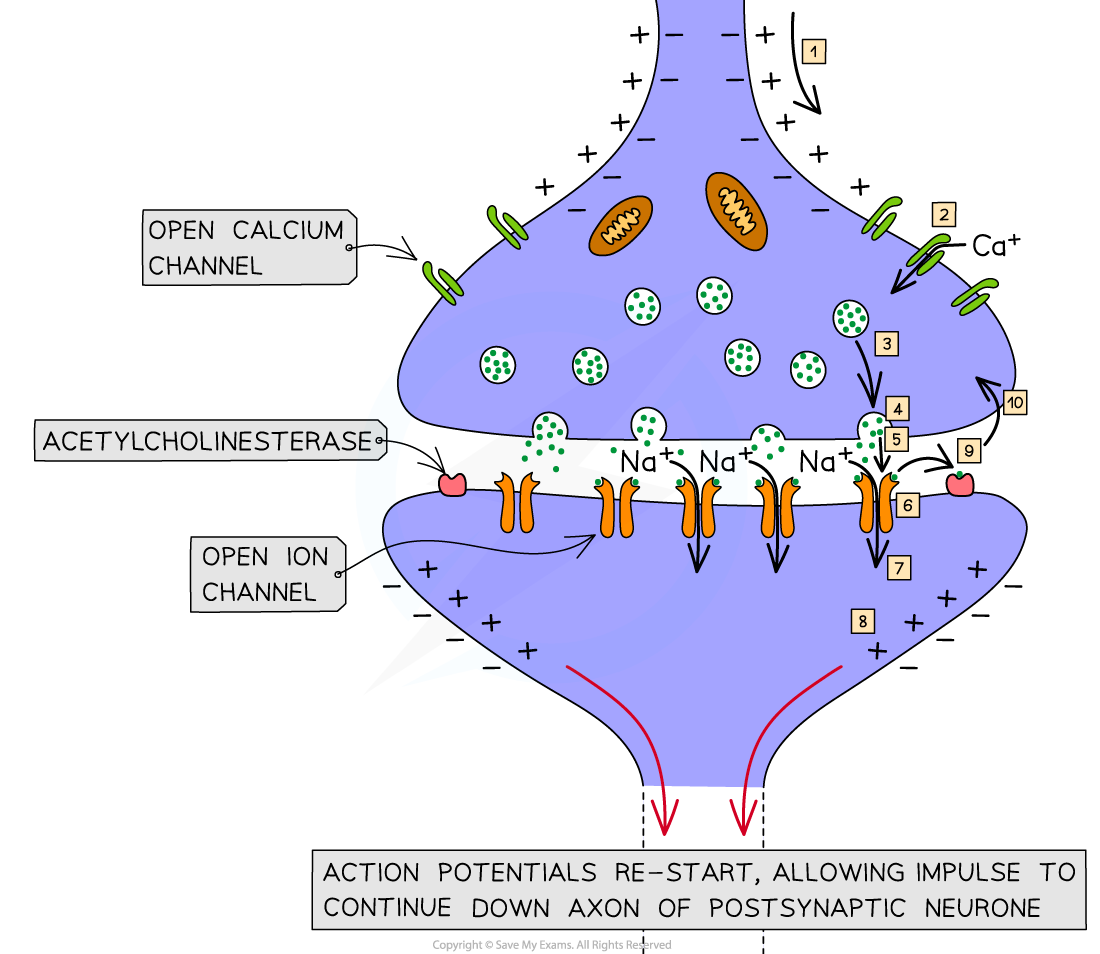
When an action potential arrives at the end of the axon of the presynaptic neurone the membrane becomes depolarised, causing voltage gated calcium ion channels to open
Calcium ions diffuse into the synaptic knob via calcium ion channels in the membrane
The calcium ions cause vesicles in the synaptic knob to move towards the presynaptic membrane where they fuse with it and release chemical messengers called neurotransmitters into the synaptic cleft by exocytosis
The neurotransmitters diffuse across the synaptic cleft and bind with receptor molecules on the postsynaptic membrane; this causes associated sodium ion channels on the postsynaptic membrane to open, allowing sodium ions to diffuse into the postsynaptic cell
If enough neurotransmitter molecules bind with receptors on the postsynaptic membrane then an action potential is generated, which then travels down the axon of the postsynaptic neurone
what determines whether an action potential will be generated in the post synaptic neurone
Whether or not an action potential is generated depends on whether or not threshold potential is reached, which in turn depends on the number of action potentials arriving at the presynaptic knob
Many action potentials will cause more neurotransmitter to be released by exocytosis
A large amount of neurotransmitter will cause many sodium ion channels to open
Many sodium ion channels opening will allow a large influx of sodium ions, increasing the likelihood of threshold being reached
The neurotransmitters are then broken down to prevent continued stimulation of the postsynaptic neurone
The enzyme that breaks down acetylcholine is acetylcholinesterase
Synapses and unidirectionality of impulse transmission
Synapses ensure the one-way transmission of impulses
Impulses can only pass in one direction at synapses because neurotransmitter is released on one side and its receptors are on the other; chemical transmission cannot occur in the opposite direction
Synapses and divergence
One neurone can connect to several other neurones at a synapse, allowing nerve signals to be sent in several directions from a single presynaptic neurone
Synapses and amplification
Amplification of nerve signals by summation
When an impulse arrives at a synapse it does not always cause an impulse to be generated in the next neurone; a single impulse that arrives at a synaptic knob may be insufficient to generate an action potential in the post-synaptic neurone
Only a small amount of acetylcholine may release into the synaptic cleftA small number of sodium ion channels are opened in the postsynaptic axon membrane
An insufficient number of sodium ions pass through the membrane
The threshold potential is not reached
The effect of multiple impulses can be added together to overcome this in a process known as summation
Summation can be achieved by
Several presynaptic neurones converging to meet a single postsynaptic neurone
This is known as synaptic convergence
Many action potentials arriving at a postsynaptic knob in quick succession
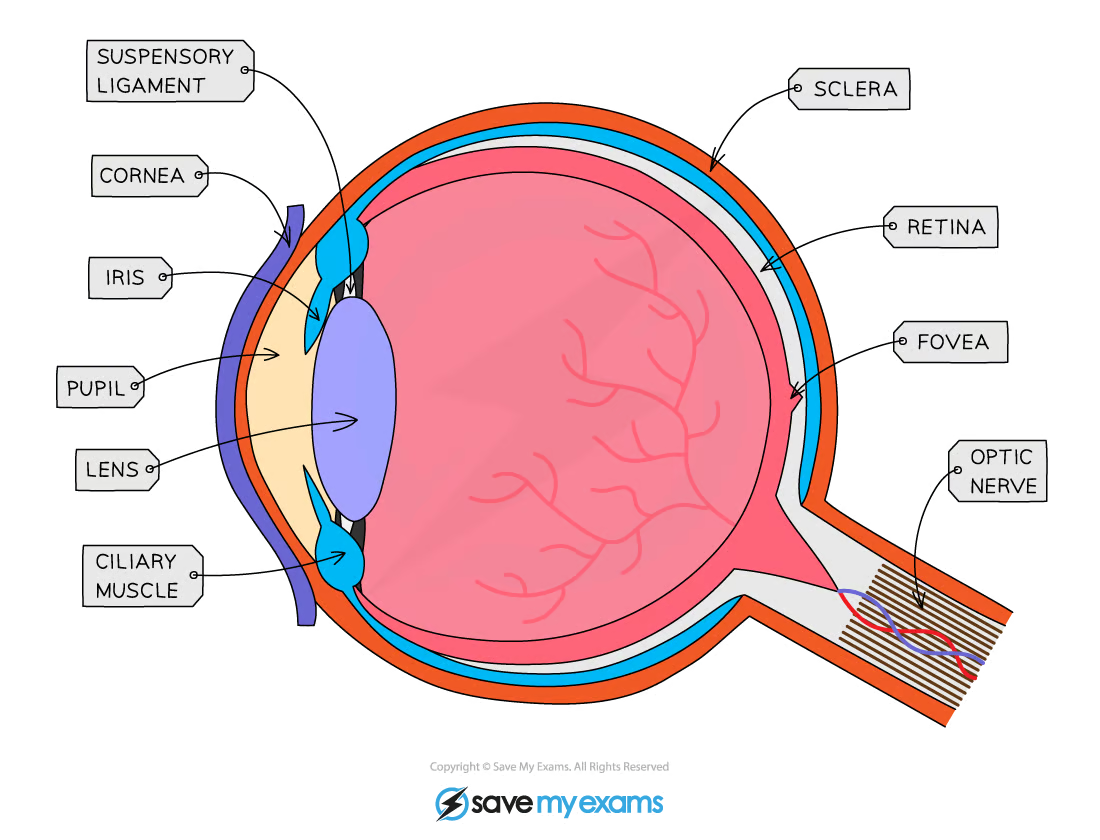
The eye intro
The eye is a sense organ containing receptors sensitive to light intensity and wavelength
Light enters the eye throught the pupil and is focused onto a region of the retina called the fovea
The amount of light that enters the eye is controlled by the muscles of the iris
Light is focused using the lens, the shape of which is controlled by ciliary muscles attached to the lens by suspensory ligaments
The muscles change the shape of the lens to allow it to focus light reflected from objects at different distances from the eye
The fovea contains many light receptors, or photoreceptors
Action potentials generated in the photoreceptor are transmitted to the brain via the optic nerve
The optic nerve leaves the back of the eye from a region known as the blind spot
The blind spot contains no photoreceptors
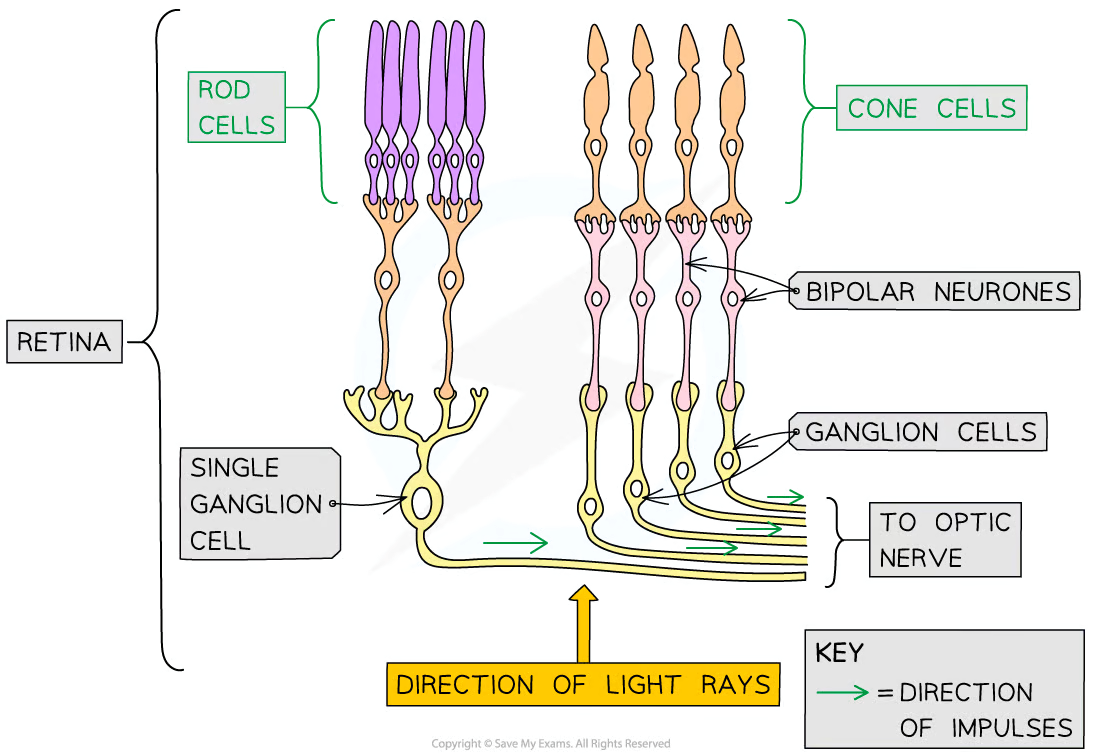
Rod Cells
One of the 2 types of photoreceptors in the retina
Primarily located around the outer retina
Rod cells are sensitive to light intensity
so can detect the presence of brightness of light
Images generated using information from only rod cells is black and white
A number of rod cells are attached to a single bipolar cell (retinal convergence)
Rhodopsin is the pigment found in rod cells
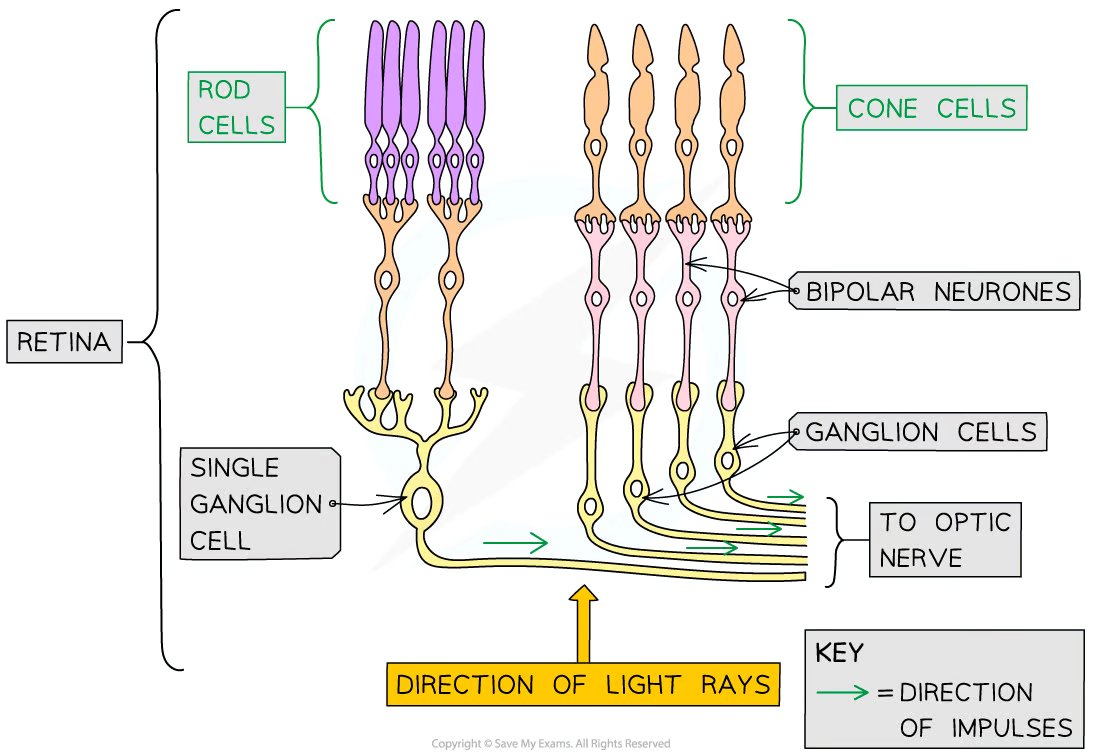
Cone Cells
Mostly found grouped together in the fovea
Sensitive to different wavelengths of visible light and so detect colour
Cone cells can be red-sensitive, green-sensitive, or blue-sensitive
The number of red-, green-, and blue-sensitive cone cells stimulated will determine the colours seen
Images generated using information from cone cells will be in colour
Iodopsin is the pigment found in cone cells
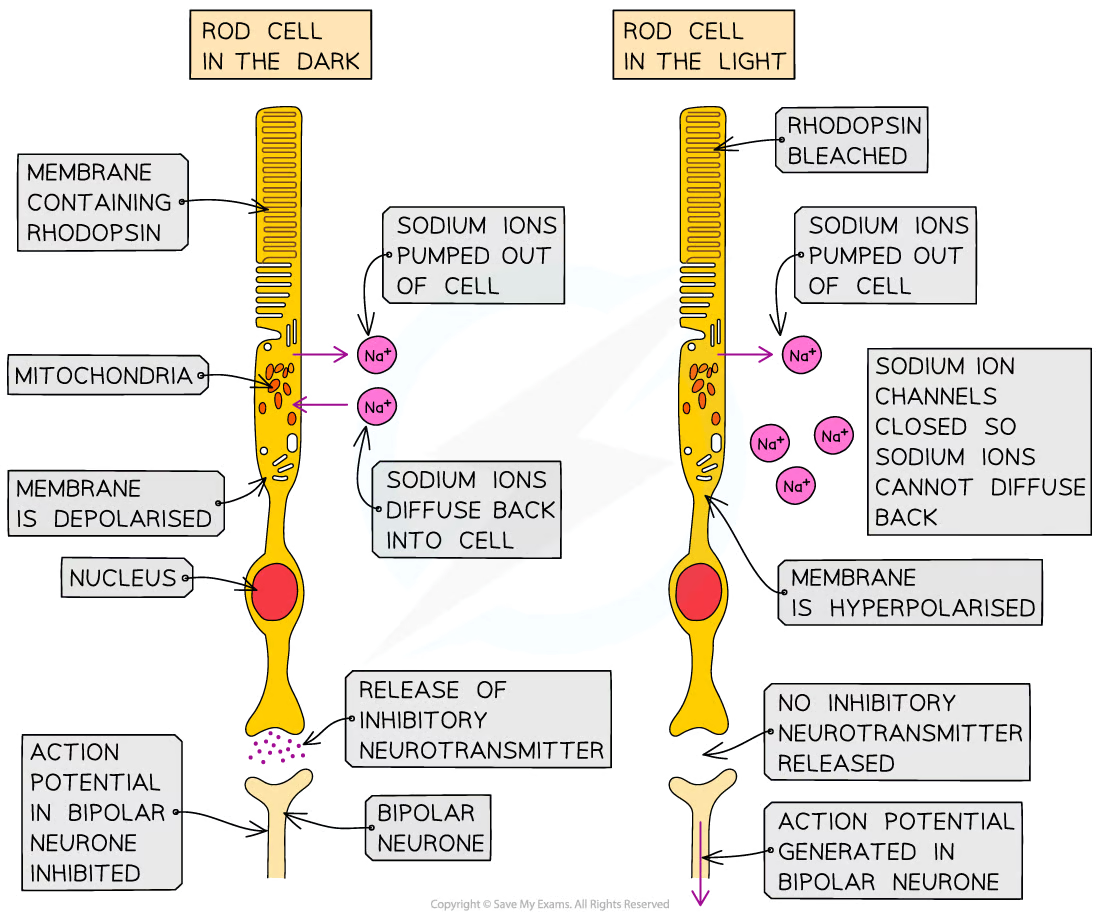
Signal transduction in rod cells in the dark
Sodium ions are actively pumped out of the rod cell, generating a concentration gradient
The sodium ions then diffuse back down this concentration gradient via the sodium ion channels
This means the cell maintains a constant state of depolarisation which means the synaptic vesicles at the end of the cell are always fusing with the membrane and neurotransmitter is always being released into the synaptic cleft
The neurotransmitter released is glutamate which binds to the post synaptic membrane of the bipolar cell and inhibits the generation of an action potential, preventing a nerve impulse from being sent to the optic nerve.
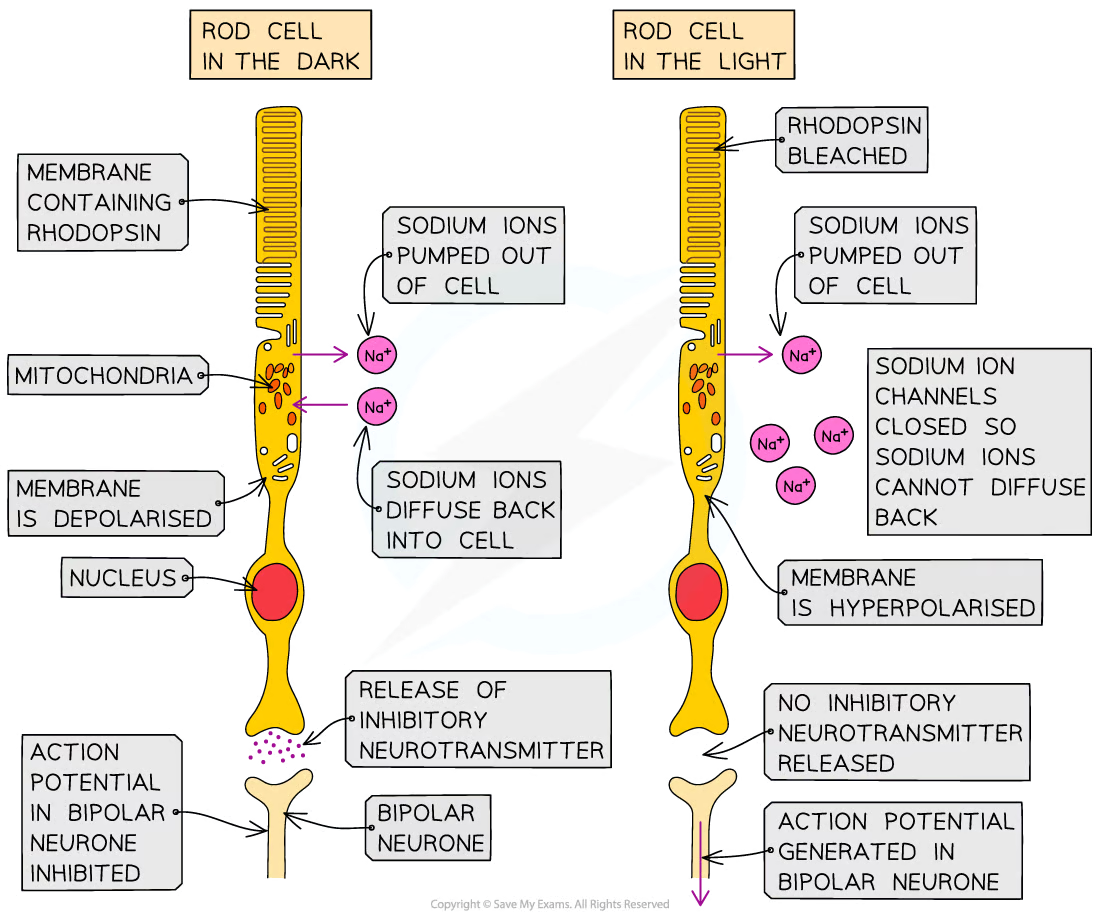
Signal transduction in rod cells in light
Light bleaches rhodopsin causing it to break into retinal (non-protein component) and opsin (protein component)
The bleaching of rhodopsin results in a signal cascade that causes the sodium channels to close preventing sodium ions from diffusing back into the rod cell
The lack of positively charged ions entering the rod cell causes its interior to become more negative until it reaches a hyperpolarised state
The hyperpolarised rod cell stops releasing the inhibitory neurotransmitter glutamate, so the generation of an action potential in the neighbouring bipolar neurone is no longer inhibited
An action potential is generated in the bipolar neurone attached to the rod cell and an impulse is sent to the optic nerve
Colour Blindness
These people do lack either red/blue/green sensitive cones but those colour sensitive cone they lack are stimulated by a different colour sensitive cone e.g. green stimulated cones being stimulated by red light
Tropism
Plants can respond to stimuli in various ways, including by altering their growth
E.g. a seedling will bend and grow towards the light because there is more growth on the shaded side than on the illuminated side
This type of directional growth response is referred to as a tropism
Phototropism is a growth response to light
Geotropism is a growth response to gravity
The response to gravity is also known as gravitropism
Tropisms can be positive or negative
Positive tropisms involve growth towards a stimulus. E.g. positive phototropism is a growth response towards light
Negative tropisms involve growth away from a stimulus. E.g. negative geotropism is a growth response away from gravity i.e. upwards
Plant growth factors
Plant growth factors are sometimes referred to as plant hormones as they are chemical messengers
Growth factors are produced in the growing parts of a plant before moving from the growing regions to other tissues where they regulate cell growth in response to a directional stimulus
E.g. auxin is a growth factor that stimulates cell elongation in plant shoots and inhibits growth in cells in plant roots
Indoleacetic acid (IAA)
IAA is a type of auxin
It is thought that IAA brings about plant responses such as phototropism by altering the transcription of genes inside plant cells
Altering the expression of genes that code for proteins involved with cell growth can affect the growth of a plant
IAA is produced by cells in the growing parts of a plant before it is redistributed to other plant tissues
IAA can be transported from cell to cell by diffusion and active transport
Transport of IAA over longer distances occurs in the phloem
The redistribution of IAA is affected by environmental stimuli such as light and gravity, leading to an uneven distribution of IAA in different parts of the plant
This brings about uneven plant growth
Phototropism in a flowering plant
Cells in the tip of the shoot produce IAA
IAA is then transported evenly down the shoot
The concentration of IAA determines the rate of cell elongation within the stem
A higher concentration of IAA causes an increase in the rate of cell elongation
If the concentration of IAA is not uniform across the stem then uneven cell growth can occur
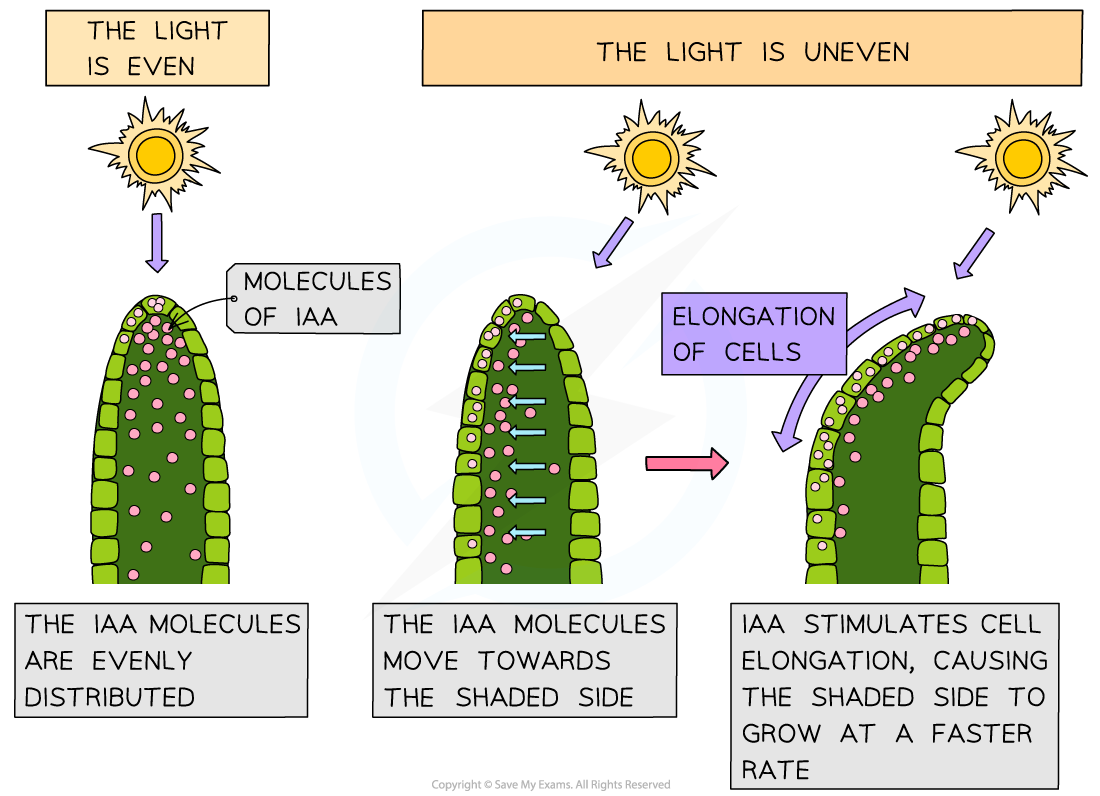
When light shines on a stem from one side, IAA is transported from the illuminated side of a shoot to the shaded side
An IAA gradient is established, with more on the shaded side and less on the illuminated side
This results in a faster rate of cell elongation on the shaded side of the root and so the shoot bends towards the source of light
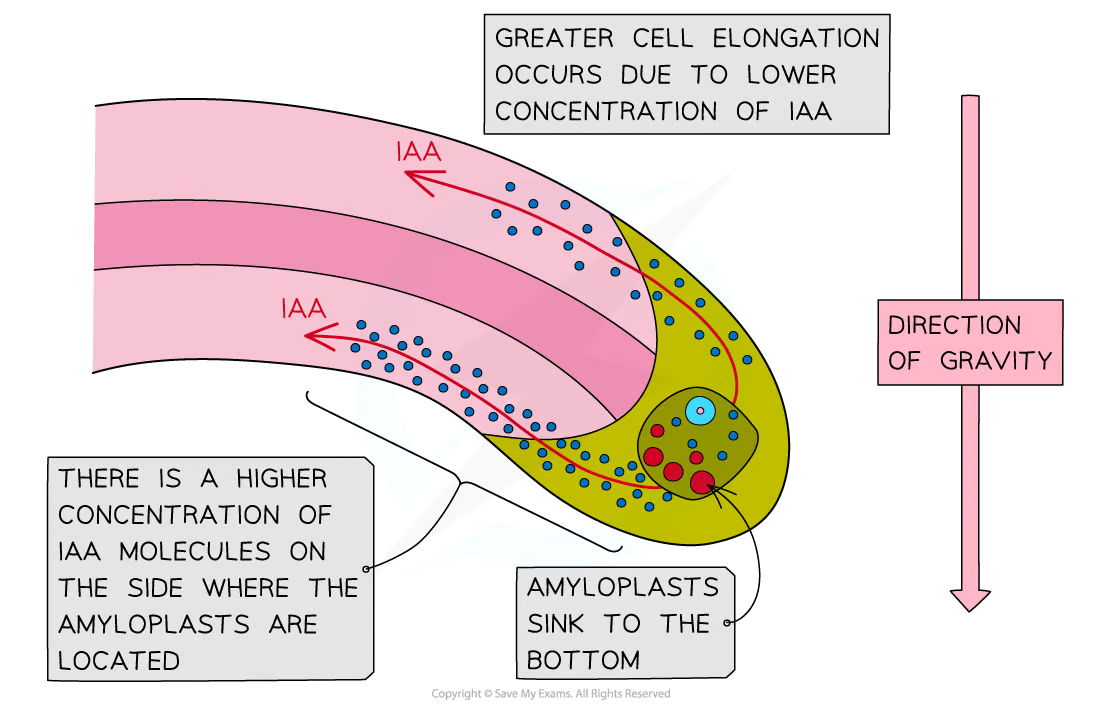
IAA in roots
Roots respond to gravity in a response known as gravitropism
In roots, IAA concentration also affects cell elongation, but higher concentrations result in a lower rate of cell elongation
IAA is transported towards the lower side of plant roots
The resulting high concentration of auxin at the lower side of the root inhibits cell elongation
As a result, the lower side grows at a slower rate than the upper side of the root, causing the root to bend downwards
What is flowering in plants controlled by?
Flowering in plants is controlled by the stimulus of night length
Some plants flower when nights are short and some flower when nights are long
When the nights reach a certain length, genes that control flowering may be switched on or off, leading to the activation or inhibition of flowering
Genes that are switched on are expressed, leading to production of the polypeptides for which they code, while genes that are switched off are not expressed, so the polypeptides for which they code are not produced
The length of night can be detected by a plant because it determines the quantities of different forms of a pigment called phytochrome in the leaf
Phytochromes
Consists of a protein component bonded to a non-protein light absorbing pigment molecule
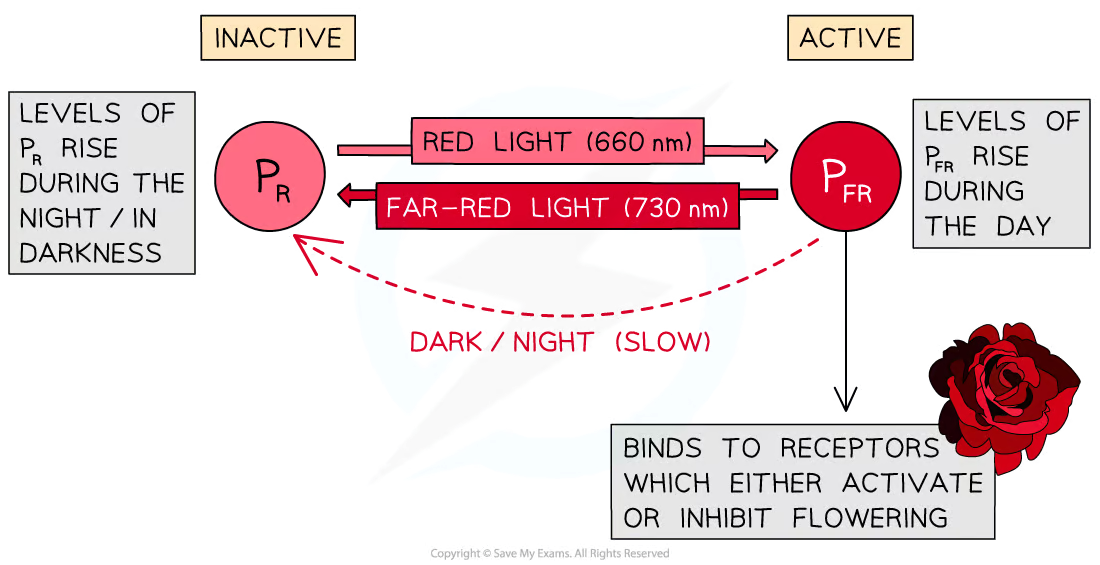
The phytochrome pigment exists in two forms
PR is the inactive form of phytochrome, it absorbs light from the red part of the spectrum (wavelength 660 nm)
PFR is the active form of phytochrome, it absorbs light from the far red part of the spectrum (wavelength 730 nm)
Absorption of different wavelengths of light causes a reversible conversion between the PR and PFR forms of phytochrome
When PR absorbs red light (660 nm) it is converted into PFR
When PFR absorbs far red light (730 nm) it is converted back into PR
In the absence of red light, the unstable PFR gradually converts back into PR
Phytochrome during the day and night
During the day levels of PFR rise
Sunlight contains more wavelengths at 660 nm than 730 so the conversion from PR to PFR occurs more rapidly in the daytime than the conversion from PFR to PR
During the night levels of PR rise
Red light wavelengths are not available in the darkness and PFR converts slowly back to PR
Short and long day plants
Long day plants flower when nights are short e.g. summer
High levels of PFR in long day plants activate/promote flowering
In short-day plants, the active form of phytochrome (PFR) suppresses flowering. During long periods of darkness (long nights), PFR is converted to PR. With PFR no longer present, flowering is not suppressed, and short-day plants flower.