Forest Ecology Exam 2
1/171
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
172 Terms
Community Ecology (Synecology)
-Community ecology is the studies involving the description and quantification of some aspect of a natural assemblage of different species of organisms.
-E.g.: the study, classification, and mapping of forest plant associations, or forest types (the description of the animal community in a small lake; or study of the change in plant and animal communities in an area over time). Community ecology is the association of populations of two or more different species occupying the same geographical area and in a particular time.
-No longer talking about population ecology (dynamics of species populations and how these populations interact with the environment. It is the study of how the population sizes of species change over time and space.)
-Community Ecology is community integrated (united) or not integrated (not united)
-Specific objectives of community ecology including understanding forest dynamics
Understanding Forest Dynamics
-The study of how plant communities originate, develop, and maintain themselves.
-Why and how forests change in their structures and species composition
-E.g.: concept, data collection, data analysis, interpretation of data.
-General explanations of forest dynamics include: critical evaluation of alternative theories or models that have been proposed
-Consider how certain premises (assumptions) related to general theories and concepts influence contemporary debates about forest management.
-E.g.: Given global environmental change, how do we understand what is happening, what do we know about ecosystem structure and function? What is the nature of plant community? Are there general models of special patterns of biotic (living) communities?
Communities
-Communities vary in their composition and structure according to variations in the physical environment
-Local and regional gradients (categories) of physical factors are associated with characteristic patterns of ecosystem types and biomes.
Coenocline (Community Gradient)
-A coenocline is the sequence of biotic communities along an environmental gradient.
-The assemblage of physical environmental factors that change as one moves along a community gradient is called a complex gradient.
-The combined community-environment gradient is called an ecocline.
Mutual exclusion
-In logic and probability theory, two propositions (or events) are mutually exclusive or disjoint if they cannot both be true (occur).
-Strong competition between each of the dominant species (tree, shrub, herb, moss) leads to strong mutual exclusion (so in competition, if one species wins, the other can not also win because if they are mutually exclusive, they can't both win).
Possible Patterns of Species Distribution
-Figure 13.1 shows 7 hypothetical patterns (a-g) of distribution of plant species along an environmental gradient such as moisture or elevation.
-There are several possible ways plants could distribute themselves along environmental gradients.
-Figure 13.1 shows a line for species of: tree, shrub, herb, and moss. (The importance value of each species changes as the environmental gradient changes).
-It IS possible for different ecosystems to be in different regions (no ecosystem is always the same)
-All species arise and grow together. They are either tightly integrated or individual.
(a.) Steep environmental gradients
(a.) Walking through discrete communities, there is tight integration.
-Mutually exclusive associations with little overlap.
-There are mutually exclusive (disjoint and cannot both be true/occur) associations because there is strong competition between the domination species.
-Each dominate species creates a particular habitat where a characteristic group of subordinate (lower ranked) species develops.
-This grouping represents how the subordinate vegetation adapted to the presence of the dominate species (Tree is on top, then shrub, then herb, then moss).
-Each dominate species occupies a particular section of the complex gradient.
-There is a change in environmental conditions over relatively short distances (there is tight integration).
(b.) Gentle environmental gradients
-Mutually exclusive associations with broad overlap (more spread out).
-Similar to (a.) but there is broad overlapping ranges because of less intense competition and less competitive exclusion among the dominate species.
-There are identifiable groupings involving all layers.
(c.) Complex gradient and interlayer competition
-When species in any layer compete strongly with other species in that layer, but show a weak response to species in other layers (or species in other layer are responding to different environmental gradients)
-This leads to very little overlap in the ranges of species within a layer.
-This also leads to little or no grouping of species between layers.
-Therefore, there is little association between species in different layers which means no clearly identifiable plant associations when looking along the gradient.
(d.) Some mutual exclusion, no obvious associations of species in different layers
-Similar to relationship in (c.) but with more broadly overlapping ranges of species with any layer.
-There is some degree of mutual exclusion within a layer but no obvious associations of species in different layers.
(e.) Few species in the dominant layer (responding to climatic factors) and many species in the lower layers (responding to a complex of local factors)
-The dominant tree species do not show mutual exclusion, but subordinate species do show mutual exclusion.
-Species (tree canopy) in upper layer have broadly overlapping distributions along the complex gradient.
-Species in lower layers show marked grouping.
-This occurs if: there are relatively few species in the dominant layer (all responding primarily to factors relating to climate that vary gradually along a major complex gradient).
-And if there are many species in the lower layers [all responding primarily to a complex of local site factors (like soil characteristics) that vary considerably over short distances].
(f.) Continuum of vegetation change along environmental gradient
-If all species distribute themselves along the environmental gradient simply in response to their individual environmental tolerances (independent of the presence of other species).
-Then, species in all layers appear and drop out again in an irregular manner when moving along the complex gradient.
-There is neither identifiable grouping between layers nor sharp exclusion of species within a layer.
-There is a continuum (continuous sequence) of vegetation change along the environmental gradient.
(g.) Bimodal or multimodal distribution
-The distribution of species along the gradient can display various combinations of the foregoing patterns at different locations on the gradient.
-The distribution of species along the gradient can display a bimodal or multimodal distribution.
Physiognomy
- The overall size and shape of an organism.
-Descriptions such as 'trees', 'shrubs', and 'herbs' are frequently used to characterize the general appearance of the vegetation of a region.
-The variation in physiognomy along such gradients occurs with variations in species composition (the 2 variations go together).
-As physiognomy varies, species composition (makeup) varies.
1. Can discrete (separate) units of vegetation be clearly defined?
-Can this be true, or is there continuous, gradual variation over space in the attributes of vegetation?
-When plant populations are analyzed according to their quantitative distribution along an environmental gradient, each species forms a bell-shaped curve.
-The curves are distributed along the gradient as a series with overlapping ranges, but no two curves have identical ranges or optima (best).
-No clusters of species are found (representing similar species behavior relative to the gradient) -This leads to recognition of a series of discrete units.
-This suggests that species populations form a shifting series of combinations along environmental gradients.
-This leads to the interpretation of vegetation as a complex and largely continuous population pattern
2. How integrated (merged) is a plant community?
-How is this, and does a plant community function as a coordinated, harmonious whole?
-Each community is characterized by a particular species composition, vertical structure, patterns of change over time, biomass, energy flow, and nutrient cycling.
-It is the biotic component of the ecosystem.
-Community has no implicit definition of spatial extent or boundaries.
-This is a question of interdependence (2 or more things being dependent on each other) or functional integrity (the state of being whole/unity and undivided). in
-A community is a multispecies aggregation with varying degrees of integration (a more or less specific composition)
-A community is a multi-species aggregation (formation of things in a cluster) with some degree of repeatability and consistency from place to place (a formation of clusters of many species that can repeat and can be consistent).
European association concept (concerning species distribution)
(1.) The individual species in the association are, to some extent, adapted to each other.
(2.) The association is made up of species that have similar habitat requirements.
(3.) The association has some degree of integration.
Fredrick Clements (organismal view)
-Fredrick Clements proposed a high degree of integration, a plant community is an organic entity (a complex organism).
-Development of the analogy between the community and the individual organism is largely attributed to Clements.
-He believed that a community, like an organism, is born, grows, matures, reproduces, and dies. Organismal view of a community. A community is like an organism.
Implications of the organismal view
-Implications: discrete, repeatable vegetation units can be recognized and classified.
-E.g.: A cell is a part of an organism just as an organism is part of a cell. A cell has predictable stages, just like an ecosystem has predictable stages.
-Succession constitutes the series of life history stages of the complex organism (progression amounts to the stages of an organisms life history).
Quasi-organismal view
-Concept of the community as a quasi-organism: says that there are certain similarities and parallels between these 2 different levels of biological organization.
-A plant community differs from a true organism in the following ways:
-In a community, there are more than one (made up of multiple organisms) different from an organism which is made up of cells that are all genetically identical.
-Communities lack genetic identity because organisms are all different/not all the same.
-The source of energy is usually fixed when talking about just one organism.
-But in a community, there are different variables that determine the amount of energy (so the sources is no longer fixed).
-A community has boundaries that are not as discrete (distinct) as that of an organism.
Alex Tansely (quasi-organismal view)
-Developed the ecosystem concept in 1935.
-Tansely's points include:
-Lack of clear delimitations (determining boundaries)
-Lack of genetic unity (same community types might develop differently/have different life histories)
-Lack of coordinated reproduction
-Lack of structural integrity
-However, Tansley saw a strong enough similarity between an organism and a community, so that an analogy is valid (distinction between the literal similarities and the analogy).
Ramensky and Gleason
-Different concept of vegetation was developed independently by these 2 plant ecologists (1920s).
-Consider that species are distributed along their environmental gradients according to their individual adaptations and tolerances.
-The community of plants observed in a particular habitat consists of populations of those species that are able to invade, survive, and reproduce successfully in that environment.
-Because no 2 species are alike in distribute, the centers of species distributions along the gradient is scattered with a broad overlap.
Individualistic and Continuum hypothesis
-Individualistic and continuum hypothesis of a region as a series of plant distributions views the vegetation of a region as a series of plant species populations distributed independent of each other along the physical gradients of the environment. (f.)
-Gleason was looking at the individual concept of the plant association.
-Supporting evidence for validity of Gleason's concept: Quantitative studies of floristic composition and vegetation gradients.
-Independent migration of species as shown by paleo-ecological studies.
-Implications: vegetation varies gradually in space and discrete boundaries between different vegetation units are rare.
-Due to the uniqueness of associations, vegetation cannot be perfectly classified.
Henry Gleason (individualistic concept of the plant association)
-Gleason thought we should think about individuals responding to environmental changes.
-Based this on 3 simple premises:
(1) Dispersal of propagules (a vegetative structure that can become detached from a plant and give rise to a new plant, e.g., a bud, sucker, or spore) occurs at different rates. Therefore, propagules arrive at different sites at different times.
(2) The site (operational environment) acts as a filter so that only certain species can survive at a particular site.
(3) Sites vary over space and time. Therefore, species composition at any particular site will be unique because of chance dispersal and the independent distribution of each species.
What is the convenience of being a super organism?
-If our worldview is structures into distinct units then our world view is more simple.
-Just like if we look at each organism separately, we will have a better understanding of each one and therefore, a better understanding of the ecosystem as a whole.
-Once you can categorize a site, then you can be prescriptive (narrow) (I understand what this super organism is, what it looks like, and what it does).
Super-organism Concept
-Although there are connections between an individual and an ecosystem, the differences are too great to make inferences.
-This is easy and more convenient. If you look at how policy is structured, the super-organism ideas are still there.
-The concept of the community as a supra-organism has never received widespread acceptance because of the numerous fundamental differences between a community and an individual organism.
Continuum/Gradient Concept
-Views vegetation of a region as a series of plant species populations distributed independent of each other along the physical gradients of the environment.
-Stressed the idea that plant species populations are independently distributed along environmental gradients (elevation, moisture) so that plant communities change gradually in their species composition.
-We are now more concerned with what is happening over time (puts us closer to the bottom of the figure, (f.) continuum of vegetation change along environmental gradient).
-E.g.: If you hike a mountain, generally tree species composition changes gradually, because tree distribution is relative to temperature and elevation, this changes over spacial distribution. More abrupt changes in shrub distribution is due to the soil composition (soil composition is a main factor that can change suddenly).
Population ecology
-Most living organisms exist for most of their lives as members of a population: a group of interacting individuals of the same species.
-There are both advantages and disadvantages to an individual organism as a member of a population.
-The study of the abundance, distribution, productivity, and or dynamics of a group or organisms of the same type (a single-species population).
-E.g.: Investigating the competition for light and nutrients in a pine plantation, the role of disease in controlling the numbers of insects on a tree, or the rate of growth and mortality of the individuals in a salmon population.
Spacial Arrangement of Populations
-How populations are structures in space.
-We can put patterns into 3 possible categories: random, clustered, and regular.
(1) A common pattern in nature is random.
(2) There could also be clustered populations (a fine spacial scale may be random, but looking at a larger scale, it can be clustered).
(3) Another pattern in nature is regular, which maximizes the space between trees.
Univariate
-One variable
-There are 3 possible univariate groups: random, clustered, and regular.
Bivariate point patterns
-Two variables
-There are 2 basic patterns, either positive or negative.
-Positive association: the occurrences of A increases the likelihood of B.
-Negative association: the presence of A decreases the probability of finding B.
Morphological causes of special patterns CH14
-Example: dead adult trees and new sprouts.
-What bivariate patterns would you expect to see in a burned forest?
-The burned forest would be clustered. There would be clumps of live regeneration of sprouts (because there is a positive association between the sprouts and the dead parent trees).
-Were there any dead trees that you would expect to see more sprouts coming out of? (positive association)
-There would be positive bivariate special pattern between dead trees and sprouts.
Morphological example of spacial patterns
-There would be a positive bivariate association between adult trees and new trees with large seeds (because large seeds can't travel far).
-Some trees regenerate almost entirely through the help of bird species.
Community causes of spacial patterns
-Regular for area with trees planted in rows and evenly separated (people did this to minimize competition between the individual organisms).
Community example of spacial patterns
-A natural forest stand that reestablished itself after a forest fire. It has a relatively regular special pattern as a result of competition (post disturbance stands generally have a regular special pattern because only the trees that do best will survive, so they have less competition).
Clustered spacial patterns
-Clustered for the single tree with other bush type species, because the 2 species rely on each other.
-You tend to find clustered patterns in areas with established organisms in harsh environments.
Wind as a mechanical force
-Uprooting, and wind-snap. Regeneration and ecosystem structure after the disturbances will be clumped in the area of the canopy of the disturbed trees.
Disturbance
-Large patches in the landscape created by disturbance. Once regeneration is in full swing, patches of regeneration will be clumped as entire area of blown down stands.
What has been important in getting an ecosystem to how it is today?
-Ask about the univariate and bivariate special patterns. From this we can infer regeneration patterns and understand dynamics within a stand.
-This will tell us what the ecosystem was like in the past.
Population vs. community
-Groups of similar organisms vs. groups of dissimilar organisms.
-A population is a group of a member of species living in a specific area.
What is the unit of a population?
-The units of a population are what organisms make up that population (e.g. a plant)
-A genet is a unit (plant) in the population arising from a seed from sexual reproduction (e.g. a seedling). This is genetically unique.
-A ramen is a unit (plant) in the population arising from a vegetative reproduction (e.g. one that comes from asexual reproduction).
-These are the same population but have different development aka different tree seedling recruitment and establishment (long term population development can be more similar).
Population ecology
-Population ecology is the study of the abundance, distribution, productivity, and/or dynamics of a group of organisms of the same type (a single-species population).
-An example would be the rate of growth and mortality of the individuals in a salmon population.
-This is the study of the abundance and dynamics of species populations.
Tree seedling recruitment/establishment example
-Figure 14-14 represents the seed rain from Picea engelmannii in three forest clearings in Utah as a function of distance from the stand.
-Seed rain is the production at and dispersal of seeds to a site. The seed rain varies as a function of distance from the plant.
-Figure 14-14: The y-axis is the uncut forest and the x-axis is the distance from the uncut forest. This shows the total amount of seed rain arriving on the ground as a function of distance from the uncut forest.
-The greater the distance from the uncut forest, the less seed rain (aka as you move away from the uncut forest, the total amount of seed rain decreases.
Recruitment of new plants into a population
-Recruitment of new plants into a population can occur in several ways as a result of either sexual reproduction or vegetative reproduction.
-This is how regeneration following some disturbance occurs.
-The production at and dispersal of seeds to a site occur in 4 ways: seed rain, seed bank, bud bank, and seedling bank.
-Regeneration that relies on seedling bank is quicker than other forms because they are already established, they just need to take advantage of their resources (light, water, nutrients).
Seed rain
-This is the production at and dispersal of seeds to a site. This can be compared to the processes of birth rates and dispersals. This is how seeds begin dispersing and arriving at a site.
-Plants have a genetically determined ability to produce an abundance of offspring.
-The seed rain varies as a function of distance from the plant (this distance can be caused by things like wind).
-The larger the disturbance (all other things equal, only thinking about seed rain), the longer the regeneration will take because there is a larger area that is far away from the uncut forest (it will take a while to get the seeds to be dispersed throughout the disturbed area).
-The shape of the tree makes a difference when thinking about seed rain.
-Regeneration will vary with tree shape. A circular shape vs a semicircle.
-For a semicircle shaped patch, there is a larger proportion of undisturbed forest closer to the disturbed area (easier dispersal).
-The more circular the patch, the longer the regeneration time because the disturbed area is further from the undisturbed area.
Seed bank
-Seed rain contributes to the seed bank.
-A seed bank is the population of living but ungerminated seeds contained in the soil (live viable seeds that are in the soil/already on site after a disturbance).
-The ratio of viable seeds of different species changes between the soil surface and deeper soil layers so removal of different layers will alter the species composition of germinates.
-The larger the seed bank, the better the regeneration.
-If there is a disturbance event that eliminates soil, then the seed bank, nutrients, and water is also removed. If soil is removed, the shape and area of the forest becomes more important when thinking about regeneration because now seed bank isn't important, but seed rain is.
-If soil is intact, the seed bank is there still.
-Some fires are so hot that the upper part of the soil is eliminatied/destroyed and the seed bank is compromised. If disturbances get larger and more intense, then there is a higher reliance on seed rain which takes longer and depends on the size and shape of the live/undisturbed forest.
Bud bank
-A bud bank is the population of buds on the lower stem; the root crown; or roots, rhizomes, or other underground organs.
-This refers to the seedlings and saplings that were in tact before the disturbance and survived the disturbance (small trees that were already established).
-The region of the plant containing these buds remains dormant due to hormones produced by the live aboveground biomass.
-When this biomass is killed, this region of the plant becomes active and new shoots are produced.
-E.g. If you have a high intensity fire vs a high intensity wind storm: the seedling bank would be more important after a high intensity wind storm rather than following a fire disturbance because the fire would also destroy the seedling bank.
Seedling bank
-Seedlings of shade-tolerant tree species can remain in the understory as surpassed seedlings or saplings for decades.
-These are capable of responding to increased light and reduced competition for soil resources after disturbance to the understory canopy.
-If there is a well established seedling bank that survives a stand-replacing disturbance, it may dominate the seed rain, seed bank, and bud bank in determining the composition of the post disturbance community of trees.
-Regeneration that relies on a seedling bank is quicker than other forms because they are already established, they just need to take advantage of their resources (light, water, nutrients).
Difference between fire and wind disturbances
-It is important to know the difference between fire and wind because when wind destroys forests, we try to fix it but we end up causing more destruction and end up slowing down the regeneration.
-Distinction between wind and fire: wind usually only kills tree canopy, and fire either kills floor vegetation or all vegetation.
Young forests vs. old forests
-Old forests are more heterogeneous (different/diverse).
-Older forests are more susceptible to wind because of the micro gusts and the different level of the canopy causes more damage.
-The old forest have a larger seedling bank because of the different canopy level which leads to a quicker regeneration rate.
Regeneration Niche
-This refers to the sum of everything a plant needs to establish and to grow.
-Environmental conditions are susceptible to growth.
-Example of regeneration niche: there is more regeneration closer to rocks and fallen logs because they modulate temperature and protect against wind.
-Regeneration on top of fallen logs is different depending on moisture and dryness.
Variations in the physical environment
-Dispersal of seeds can result in either a random or uniform distribution of plant propagules over a large area.
-Only the plant propagules that land on a suitable substrate will become established.
-For moisture-requiring plant, only the propagules that land on moist soil will survive, which will lead to clumping of these species on this particularly moist site.
Geometric Population Growth
-If a reproductive unit of a species with a short regeneration time is placed in an environment that provides all it needs to live, then the abundance of the organism will increase over time. (Figure 14-4)
-This pattern of population change is called geometric/exponential growth. It starts of exponentially but as it approaches the carrying capacity it levels off (e.g. elk herds get too large until they cant support themselves any longer).
-The abundance of the population shows an increase with time.
-If an organism has discrete/non-overlapping generations (annual plants w/ a single generation per year), then N(t+1)=R(sub0)*N(subt)
-Aka: The population size in the subsequent generation (N(t+1)) is equal to the net reproductive rate (Rsub0) times the population size at generation (Nsubt).
-The higher the reproductive rate, the more rapid the population growth.
-There are usually constraints that make this logic non-realistic. The reason this doesn't apply is because there are different.
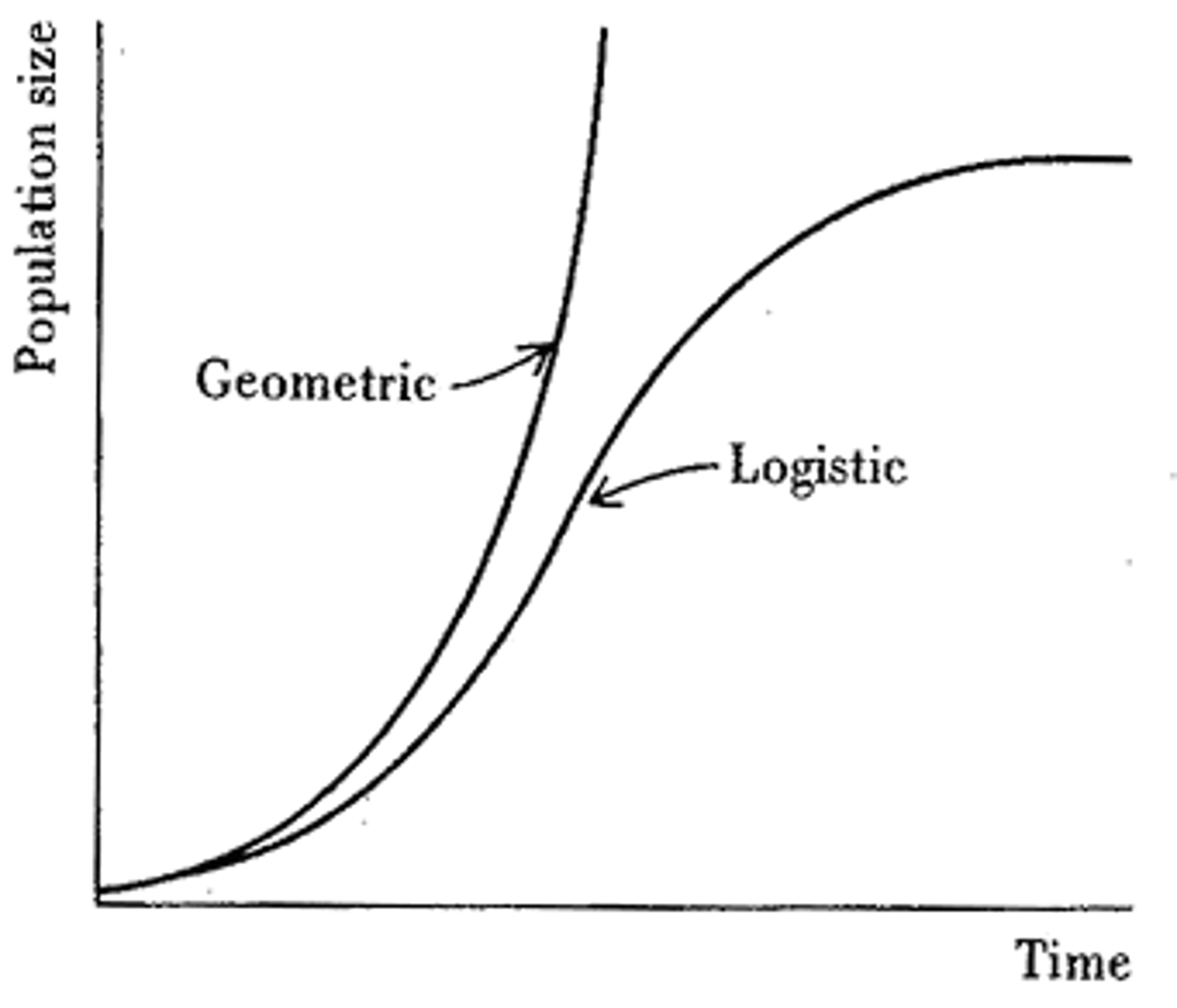
Geometric vs. Logistic Growth
-Geometric population increase is characteristic of the early phases of population growth (growth of a population that has recently escaped from the traditional population control e.g. the human population).
-Geometric growth ignores the reality that the early growth of a population changes (the populations develop over time).
-Despite the enormous reproductive potentials, populations of some organisms grow at geometric rates for only a brief period of time (ignoring disturbances, they generally don't show sustained growth over many generations).
-Logistic growth considers the lack of sustained growth over many generations.
Logistic population growth
-There is a lack of sustained growth because as a population grows in numbers, its negative feedback (environmental resistance) also grows due to competition, disease, stress.
-As a population increases, so does the environmental resistance which steadily reduces the rate of population growth until the resistance is so great that population increase is no longer possible.
-The population grows slowly at first, then there is a period of rapid increase, then the population growth rate slows down as it approaches the carrying capacity (this is where the environment is full of a particular species).
-The population can come crashing down because the more a population overreaches the carrying capacity, the more it will quickly shoot down.
Life table
-This provides a summary of the schedule of deaths in a population and has the causes of mortality in each age group.
-The number of survivors at the start of each age interval.
Survivorship curves
-Look at the age distribution of a population. We can say there are a certain number of people that are young and a certain number that are old, then assume that the age structure is a predictor of mortality.
-There are a large variety of survivorship curves that can be found in nature, but there are 4 major types.
-There are two categories of a survivorship curve:
1. the cohort or dynamic type
2. the stationary age distribution, or static type.
-Survivorship curves are not constant they change with removal of predators.
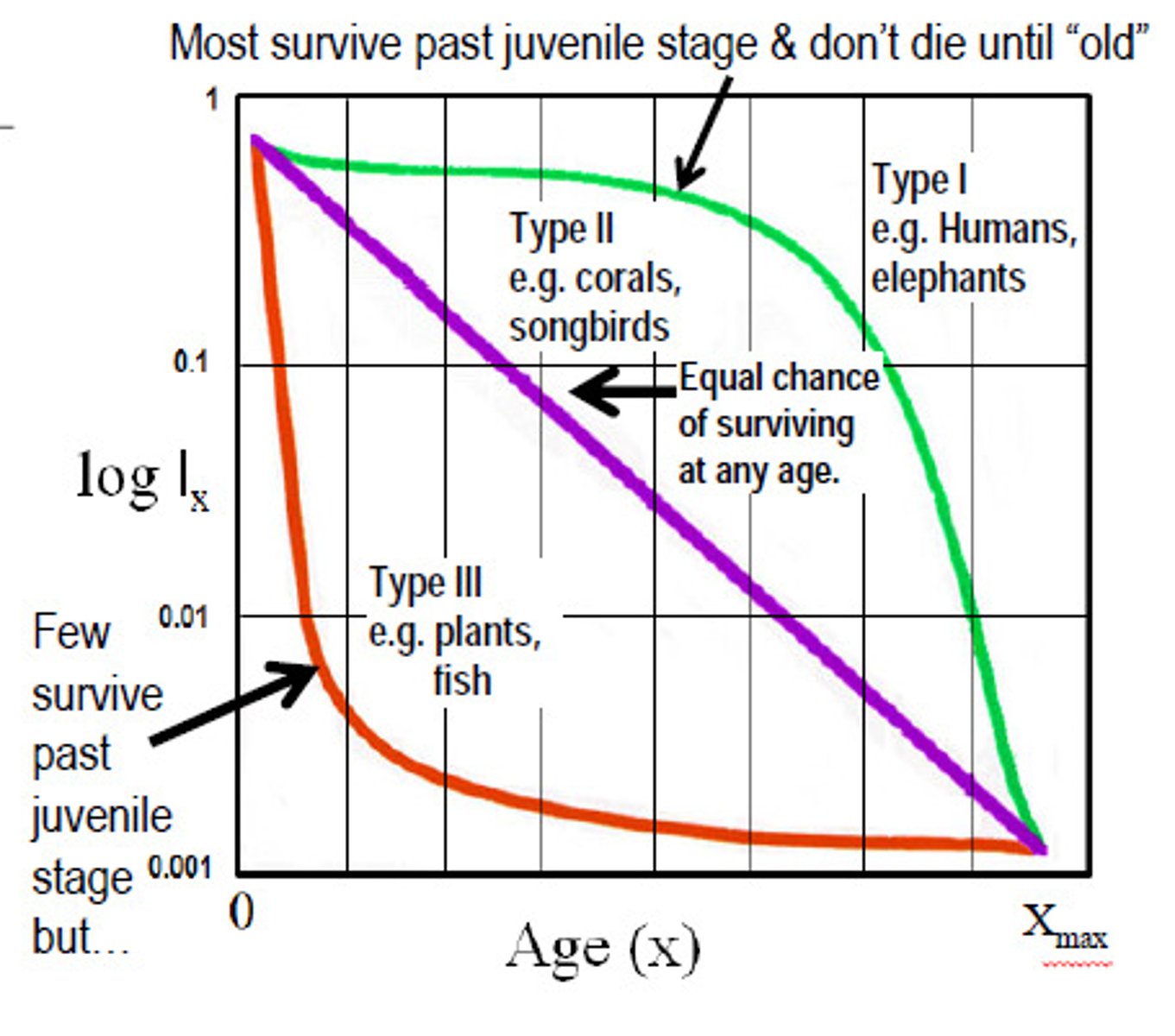
Survivorship curve type 1
-The population has very low mortality until near the end of their life span
-Most dying organisms are old.
-Humans
-Its the top curve
Survivorship curve type II
-Mortality in type II populations is distributed evenly across all age classes
-Its the straight line
-There is a constant number of deaths per unit time.
-Mortality is independent of age
-Birds tend to have this.
Survivorship curve type III
-In type III, a constant percentage of the survivors die in each time interval.
-There is high initial mortality then lower mortality at old age
-This is common in plants because as
-Type IV is similar but its more heavy juvenile mortality followed by low and fairly constant mortality for the rest of the life span.
Selection patterns
-Either r-selected or K-selected.
-For r-selected, the mortality is density independent, it has a type 3 survivorship curve, there is low competitive ability, a short life span, and rapid rate of growth.
-For K-selected, the mortality is density dependent, the survivorship curve is types 1 and 2, the competitive ability is high, the life span is long, and the rate of growth is slow.
r-selection
-This happens in environments that are patchy and unpredictable, there is high risk to any individual, there is very little competition from other organisms.
-The optimum evolutionary strategy is to produce large numbers of small offspring.
-These species are r-strategists and have been produced by r-selection.
-Rapid rate of increase in early stages of succession.
-They have initially high survivorship curves, life span is short but growth rate is rapid.
K-selection
-Species in a more predictable and uniform environment with high levels of inter and intraspecific competition.
-They are selection to produce fewer, but larger offspring with greater competitive abilities.
-The K is the same K as carrying capacity but is associated with more stable populations.
Static vs. cohort (dynamic) life tables
-Static life tables: we use static life tables to assume the future. You have to make assumptions about survival and mortality (e.g. you could assume there would be more fir than spruce in the future, but survivorship curve would have to stay the same to assume this. This can be difficult because survivorship curves to NOT hold constant).
-Cohort (dynamic) life table: if you follow a population over a time period (e.g. you see a cohort of trees establish, come back and see and record which trees have survived) The advantage of this is you can see the actual development, however this is very impractical.
Static life table
-These are made by examining the age distribution of the population at the time of a census.
-e.g. the age distribution of trees in a 1 ha Douglas-fir forest at the time of measurement
Density-dependent vs. density-independent regulation (mortality)
-Two major properties of populations: variation in mean abundance and variation in temporal fluctuation.
-These are types f mortality
-Theories grouped into schools of thought: the biotic (density-dependent) school, and the abiotic (density-independent).
Density-dependent mortality
-With density dependent feedback mechanisms, populations can avoid increasing to levels where they destroy their habitat or supply of resources, or they decrease to extinction. It matters how dense the trees are when thinking about mortality.
-r-selected species tend to have a density
-Even mortality driven by external forces has a component of density dependance.
-E.g. We think of drought as an external factor. There is still a density component.
-E.g. Parasites and predators (but there is a lag in the response time, so there is less balance).
-E.g. Competition for food or other resources.
Density-independent mortality
-Suggests that because mortality affects individual organisms rather than populations, investigations of animal distribution and abundance should focus on the individual
-There are other factors that influence the survival and reproduction of individuals: weather, food, other organisms, and shelter.
-E.g. Droughts or natural disturbances are not dependent on how dense a forest stand is.
-r-selected species tend to have a density independent mortality.
What is ecological succession?
-Ecosystems change for a variety of reasons: change in climate, soil conditions, change as a result of interactions between the members of a living community, and change caused by disturbances like wind and fire.
-The process of change after disturbance is called ecological succession.
-Over time, there is a series of living communities- plants, animals, and microbes- that successively occupy and are replaced on a particular area of land, with accompanying changes in soil and microclimatic conditions.
-Ecological succession is a directional (non cyclical, there is some difference in species that occur in latter and before), non-seasonal(longer time periods, decades to centuries ago), cumulative change in the types of plant species that occupy a given area though time.
Difference between stand structure and ecological succession
-Stand structure is the distribution of trees by species and size within a stand. The structure is the result of several factors: Growth habit of the tree species, especially the degree of shade tolerance.
-Ecological succesion describes what happens over time to species composition. (how it changes after a disturbance)
-A stand structure is more general terms.
Why does ecological succession occur?
-Under the changed conditions of the environment, the previously dominant species may fail and another species may become ascendant. Ecological succession may also occur when the conditions of an environment suddenly and drastically change.
-Either allogenic or autogenic succession occurs.
-The driving force behind succession, the reason why the change occurs, is not always the same.
-We can classify according to the driving force with 3 main categories of succession: autogenic succession, allogenic succession, and biogenic succession.
-Describes what happens to species composition over time.
Autogenic succession
-In many cases, the replacement of one community by the next happens because of changes in the physical, chemical and biotic environment that have been produced by the resident organisms.
-These changes often leave the site less optimal for the organism that produce the change and more optimal for those organisms that replace them.
-The stimulus for change is an internal one (comes from inside the ecosystem) (e.g., gradual soil improvements could allow a new species to develop).
Allogenic succession
-Allogenic secessione occurs when physical processes that are independent and external to the biotic community cause changes in the physical environment, which leads to changes in biota.
-E.g. filling in a lake with sediment to form a bog will change the biota.
-Stimulus for change is an external one (comes from outside the ecosystem).
Biogenic succession
-Occurs when there is sudden interference with an autogenic or allogenic succession by a living organism that temporarily becomes the major agent of successional change.
Types of succession
-Ecological succession, the process of ecosystem development, occurs in virtually every type of environment found on earth. The details of succession vary according to the type of ecosystem.
-Primary or secondary succession.
-Succesion begins after a disturbance, whether it is at a slow rate (primary succession) or a fast rate (secondary succession).
Primary succesion
-This is when succession begins in environments that lack organic matter and that have not yet been altered in any way by living organisms.
-Sites where that was not previously vegetated or all sources of vegetation have been removed.
-E.g. lava flow that covers and eliminated seeds, or glacial retreat: may have been vegetation before, but there is no more remnants of the vegetation. Severe fires could also cause this.

Secondary succession
-Where succession begins in an environment that has already been more or less modified by a period of occupancy by living organisms.
-This is more common, and refers to areas that were previously vegetated.
-Secondary succession is faster because the soil already has the nutrients and it doesn't have to be recreated.
-If the soil is still intact, post-disturbance regeneration will be much quicker because the soil is still there.
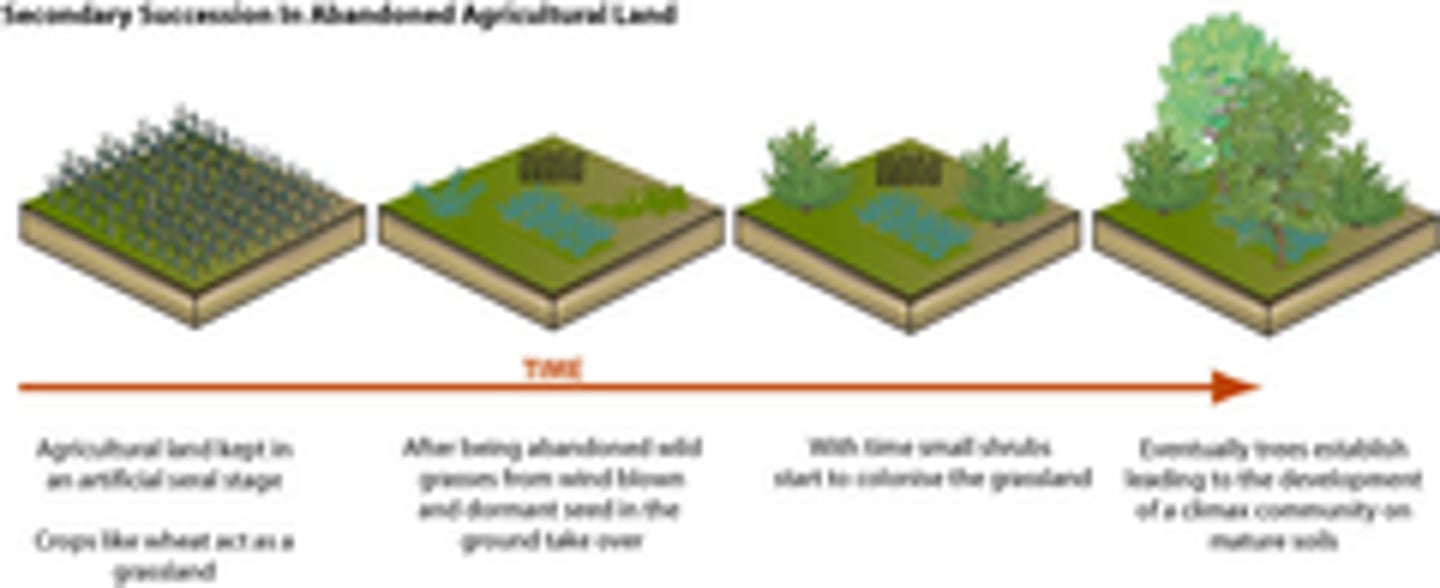
Relay floristics
-Two principles are considered as being involved in vegetation development on abandoned agricultural lands.
-The first, called Relay Floristics, involves a succession of incoming and outgoing plants, each group invading the land, driving out its predecessor, and in turn preparing the site so as to be driven out itself.
-Entry and growth of the latter species depends on earlier species.
-Establishment occurs in stages over time. first there were crops then weeds came after, then grassland, shrubland and finally forrest came after respectively.
-Sequence of species arriving later and the alteration of the site by the early colonizers.
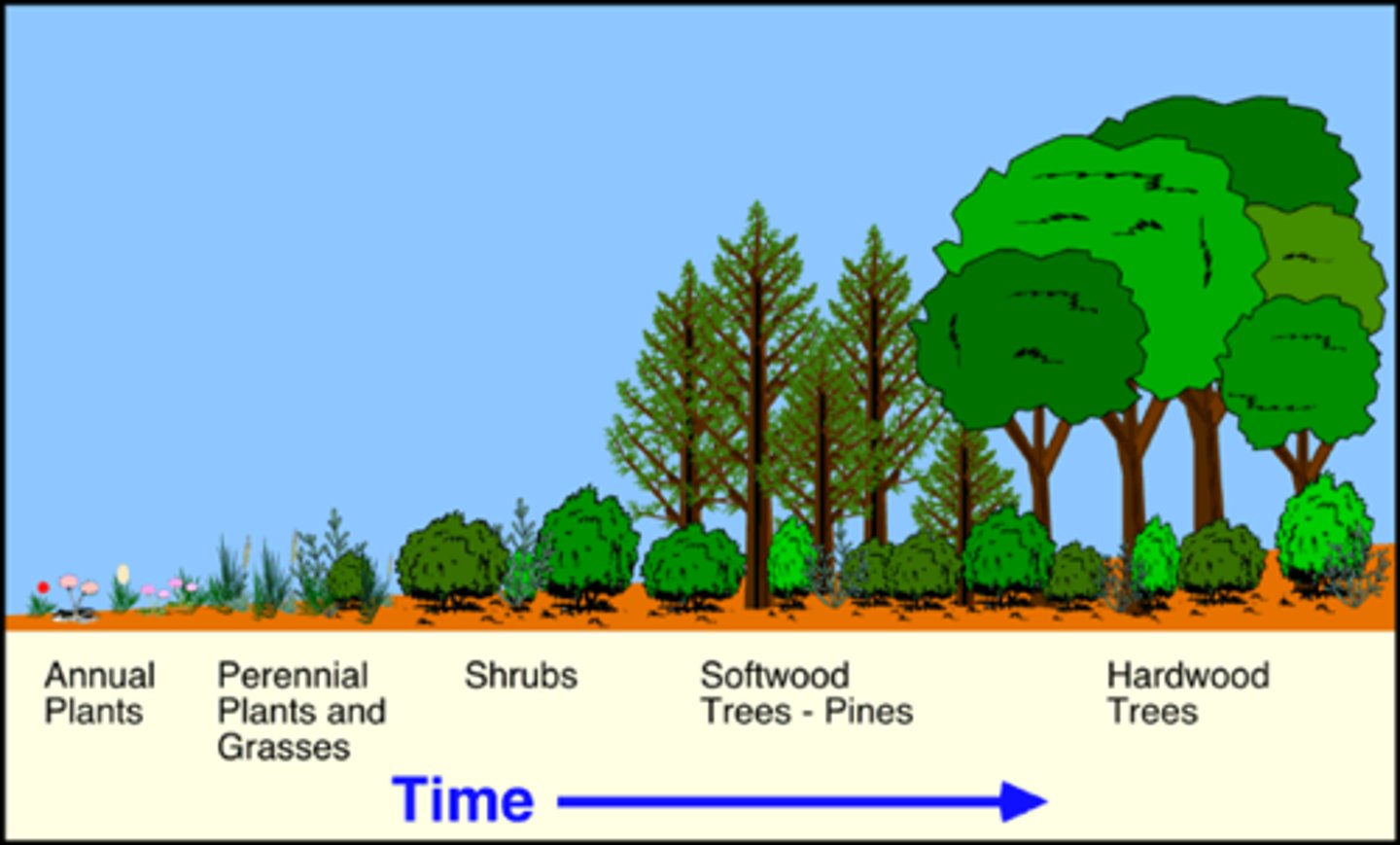
Initial floristics
-Part of the individualistic/stochastic view of succession by Gleason (part of the mono climax theory of succession).
-Eglar proposed that the succession of plant species in abandoned agricultural fields depends on relative growth rates and shade tolerance of the species arriving (succession was dependent on who gets there first and their relative growth rates).
-All species establish soon after disturbance.
-Dominance (not establishment) occurs in stages over time.
Mechanisms, pathways, and models of succession
-Mechanisms: a process or interaction that contributes to successional change (e.g., competition, predation, change in soil moisture and nutrients availability).
-Pathways: the temporal pattern of ecosystem change (e.g., the sequence of plant communities and changes in physical and other biotic components of the ecosystem over time).
-Models: some abstract conceptual representation used to describe and or explain successional pathways in terms of successional mechanisms that result in successional change.
-Ecologists have been struggling to find a general explanation of the mechanisms and pathways of ecological succession.
Facilitation
-Environmental alteration by pioneer species is frequently a necessary condition for successful establishment of later successional species.
-Facilitation model -like clemencies model, follows the relay floristics model
-Each wave of colonists "prepares the way" for the next group.
-Biotic modification changes conditions to favor later stages.
-More likely to occur in primary succession.
-Facilitation would be more important during primary succession because when you start at the base level, with nothing, the gradual growth of the first species that establish in this area provide a more available area to thrive.
Inhibition
-All species resist invasion of competitors. Can make the soil more or less available to next species.
-It is much more common, given a seed source, there is competition and inhibition.
-This looks like the relay floristics model, however, early seral stages prevent establishment of later stages.
-Competitive inhibition of juvenile stages of later seral species.
-Early seral stages act as habitat for consumers of later seral species
How do early successional species differ from late successional species?
-We can classify species as being early of late successional species based on their seed size.
-Seed dispersal matters, because some seeds are extremely small and other plants force larger seeds.
-Early successional species need to establish in areas that are free from competition, so the smaller and the lighter the seed is, the easier it is for the seed to do this.
-Late successional species have more intense conditions and greater competition, so the larger seeds are more successful (there are less larger seeds, but these few large seeds have low mortality rates).
Stress tolerance/competitive ability
-Early stages of succession are relatively free of competition because their focus is on growing. In early stages, the efficiency with which solar energy enters the ecosystem is low.
-In late stages of succession, there is more competition.
Life history "strategies" or "vital attributes"
-Vital attributes are 3 types of knowledge that allow for predicting the direction and patterns of succession in a disturbed ecosystem.
1. Method of persistence (the method of invasion and colonization and the method of persistence of propagules after disturbance).
2. Conditions for establishment (the conditions required for establishment and growth to maturity).
3. Critical life history events (time taken to reach critical stages in the life history).
-E.g.: seed dispersal characteristics of dominant plants, photosynthetic efficiency of dominant plants at low light, competitive ability, plant longevity, r-selected or K-selected, plant biomass, rate of Net primary productivity, site of nutrient storage.
Climax
-Climax is a self-replacing seral stage that is relatively stable (in the sense that it persists for a long time- several to many generations of dominant plant species-relative to other seral stages).
Seral
-A seral community (or sere) is an intermediate stage found in ecological succession in an ecosystem advancing towards its climax community. In many cases more than one seral stage evolves until climax conditions are attained.
Monoclimax Theory (Cements)
-This is a classical theory of succession.
-This theory states that the species composition and structure of the end community (what the dynamics of the area und up looking like aka the climax) are determined by the regional macroclimate.
-The theory only requires a definite linear development toward the climatic climax.
-Clements had the idea of monoclimax (one climate): the view was a convergence (intersection) of floristic composition (vegetation structure and plant community types) that's determined primarily by climate.
-If you know the climate, you can determine what type of species will be there. This theory shows stability of the endpoint of succession. There is a strong role of climate control which is seen as the dominate factor determining what type of organisms.
-This theory states: There is a strong role of climatic control of the climax; ensures stability of the endpoint of succession (permanence of climax); reaction (community control, dominance of autogenic change) as the mechanism (proximate cause) of change.
Supraorganisms
-Monoclimax theory is linked to the idea that the climax is comparable to the supra organisms.
-The term "supraorganism" refers to a collection of individuals which behave as a single unit with enhanced function.
-(we prefer to the more common but slightly less informative "superorganism")
-Biologist Mobius recognized the intimate association among oysters and the algae they fed on, the parasites that prey on them, and the other organisms they compete with.
-Recognition of the integration of the components of a biotic community into a functional system led to idea of the community as a supra organism.
Issues with Monoclimax theory (theoretical explanation)
-Life history (the series of changes undergone by an organism during its lifetime) of the climax formation as a theoretical explanation of succession: if you know previous climate, it can explain the succession.
-This theory is false.
-There is microclimatic variability these could affect vegetation.
-If you apply this view, it won't be accurate because ecosystems are changing either gradually or abruptly.
-This view also ignores the role of disturbances as an agent of change.
-This view also says ecosystems are the base of environmental movements.
-Also says that everything is stable and unchanging.
Early critiques (Gleason)
-Gleason discussed some of the problems with mono climax theory.
-He pointed out that it ignores climatic variability and disturbances as ecosystems change.
-These people worked in the 20s and 30s.
-Early theories of succession ignores the importance of disturbances and climatic variation (must consider disturbance and climate change).
-Disturbances were seen as abnormal which led to problems of management.
Polyclimax Theory (Tansely)
-During succession, floristic convergence (intersecting of vegetation) is only partial (incomplete).
-There may be more than one type of stable endpoint.
-Alternative stable endpoints are controlled by local factors such as topographic position or soil type (consequently, called topographic climax, edaphic climax, etc. as apposed to climatic climax).
-This theory points out that there are many factors that can intervene to prevent and area from reaching the climatic climax condition.
-In addition to the climatic climax, there are other environmental factors including: pyral climax (fire), edaphic climax (soil), and biotic climax (animal influences).
-Polyclimax theory: the vegetation of a region is viewed as a pattern of communities at different stages of succession, some of which may reach climatic climax relatively rapidly (only a few centuries).
-The idea of many climaxes (not only climate factor).
-This view reduces the scale and looks at finer spatial patterns.
-Also recognized that there may be more than one type of stable endpoint that are controlled by local factors.
Issues with Polyclimax theory
-This is also a false theory because this idea is only talking about climax.
-This theory doesn't fit reality because we need to recognize disturbances as normal, usual occurrences, not as rare events.
Assumption of climax theories
-Figure proposes mono climax theories.
-It is looking at ecosystems being more or less integrated or individual.
-Shows that ecosystems are coherent (logical and consistent) units.
-It's important to recognize that individual species can respond individually to different environmental conditions.
Patterns Climax Theory (Whittaker)
-This theory does not require discrete (separate) units.
-Landscape consists of environmental gradients.
-Tendency for a stable type of vegetation to develop will be different at each point along these gradients.
-Therefore, the climax vegetation will be a spatial pattern of vegetation which reflects the spatial variation in the underlying physical environment.
-Work in the 50s proposed that we view ecosystems using a continuum and recognize that ecosystems change along the continuum.
-He put forth the last climax theory which was a combination of individualistic concept of plant association and the continuum/gradient concept.
-Whittaker was trying to get us away from viewing ecosystems as being stable.
-This is a combination of the continuum/gradient concept and the individualistic concept of the plant association.
-In areas of relatively homogeneous regional climate, Whittaker recognized the most common climax type as the prevailing (current/existing) climax.
Advantage of this climax theory
-It doesn't require discrete units.
-It recognizes space over time and continuous change (this is closer to reality).
-Landscape consists of environmental gradients.
-There is a movement away from trying to think of ecosystems as superorganism and looking more at patterns or process.
Disadvantages of climax theory
-These people came up with the ideas that shaped ecology.
-All theories except Whittaker's have been discredited because no scientists talk about climax systems anymore.
-If you look at forests that haven't been disturbed for many years, you will see characteristics of the region, however, you cannot assume that this is normal.
-When there is a disturbance we need to see it as a normal event in order to have correct management responses.
-The climax concepts carried so much baggage because it was so difficult to save some of these theories that they just discredited all of them.
-Some of this viewpoint still exist today and if we recognize these, we can see why some policies are wrong.
Steady-state concepts (Whittaker)
-This concept talks about literally what is happening in ecosystems.
-A steady-state ecosystem is a constant biomass and nutrient content over time (there are inputs and outputs, but over time/decades there is no huge change).
-Steady-state populations have constant population sizes.
-Steady-state communities (aka compositional equilibrium) has species persistence.
-In steady-state, there is no endpoint, it's just saying over centuries and decade, things haven't changed much.
-The relative proportion of species isn't changing that much. This allows for change more than the climax theory.
-In practice, the steady-state concepts displaced Whittaker's patterns climax theory.
-E.g.: to interpret whether it is a steady-state community, you have to know the past of the specific ecosystems and the past species composition.
Disturbance
-Any relative discrete event (forest fires, wind storms, logging operations, insect outbreak) in time (there is a clear beginning and a clear end) that disrupts ecosystem, community, or population structure and changes resources, substrate availability or the physical environment (nothing has been destroyed or ruined, just changed).
-This is a very literal description of change. Looking at this from an ecological perspective, it may or may not be a bad thing (makes us think neutrally).
-There is a book that changed collective ecological thinking towards the idea that ecological disturbances are the norm.
Patch Dynamics
-The patchiness in vegetation is both due to patchiness in the underlying physical environment and due to vegetation responses to disturbances.
-They situated disturbances fundamentally in vegetation structure.