Why can’t noble gases form any bonds?
Because their electron shells are completely filled.
What determines the shape of a molecule?
The shape of a molecule is determined by the shape of the orbitals that make up the covalent bonds, e.g. methane looks like a tetrahedron.
What is a polar covalent bond?
When the electrons are evenly distributed ex: H-H. This is not true in water: there, electrons spend most of their time close to oxygen, because oxygen is more electronegative.
What is electronegativity?
Electronegativity is a measure of how strongly electrons are attracted to the nucleus. C-H bond is regarded as non-polar
Hydrogen bonds
The biggest consequence of polar covalent bonds is the hydrogen bond, the electrostatic attraction between the partial positive and negative charges of two water molecules.
Properties of Water
Cohesion explaining how water can rise in trees; meniscus and surface tension; liquid water is denser than ice allowing survival of larger organisms in lakes; water moderates temperature, because hydrogen bond formation or disruption buffers heat energy. A last property of water is that it facilitates chemical reactions, because it is such a good solvent.
Polar molecules
dissolve easily in water; they are called hydrophilic
Non-polar molecules
do not dissolve in water and are called hydrophobic. The hydrophobic interaction describes nonpolar molecules being forced together in water, because being together minimizes the disruption of hydrogen bonding in the surrounding water. It is a weak force and a little heat disturbs it.
van der Waals force
the attraction of nonpolar molecules to each other because of transient dipoles (caused by the random localization of electrons in different areas of their orbitals). Occurs between all molecules but is only relevant for nonpolar molecules.
Ionic bonds
Ionic bonds relevant in biological settings are those within proteins and are relatively weak; ionic bonds in salt crystals are much stronger, similar to the covalent bond or even stronger.
Acids and bases
important because most biologically important compounds are acids and bases, e.g. nucleic acids, amino acids, phospholipids, many metabolic compounds. Acids release/donate H+ ions (protons) in solution. Weak acids like the carboxyl group are most important in biology. They dissociate partially and reversibly.
Bases accept H+ (protons) in solution, and therefore release OH-. The amino group is a weak base and the most important in biology. It partially and reversibly accepts protons.
In summary, both acids and bases can be characterized by the amount of protons in solution.
Buffers
Why dissociated acetic acid (acetate) is also called the conjugate base, because it can accept protons. So weak acids act as buffers, because over a certain range the pH does not change much, even though you add a lot of base. As can be seen from the titration curve, a weak acid on its own can only buffer added base. If you want to make a buffer that buffers in both directions, you have to add equal amounts of weak acid and its conjugate base to reach the half- equivalence point of the titration curve. Buffers are very important in biology, because it is important for the function of many molecules that the pH is constant. That’s why our blood and intracellular fluids are buffered.
Hydroxyl
Functional group - OH
Carboxyl
Functional group - C=O (Carboxylic acid COOH)
Amino
Functional group - NH3
Macromolecules
We usually call proteins, nucleic acids and carbohydrates macromolecules, because they can form large polymers.
Condensation
formation of a polymer linked by covalent bonds, releasing one water molecule with each monomer added. This reaction is anabolic, requires energy input. Examples are DNA replication, protein synthesis, making of starch.
Hydrolysis:
Breaking of covalent bonds with the help of water to transform a polymer into its constituent monomers. This reaction is catabolic, releases energy. Example: digestion of food molecules for energy generation.
ATP hydrolysis
ATP hydrolysis releasing energy or breaking a starch polymer into monomers releasing energy, means the entire chemical reaction, which involves at least 2 covalent bonds being broken and 2 covalent bonds being formed. Breaking a single covalent bond requires energy, whereas forming a single covalent bond releases energy.
Proteins
Proteins are the machines that do all of the work in a cell: build structures like hair, replicate DNA, catalyze metabolic reactions, transport materials inside cells and across the membrane, etc. Proteins are made of amino acids, which are ionized at neutral pH.
Amino Acids
There are 20 amino acids, which are classified according to the properties of their side chains: nonpolar, hydrophobic amino acids; polar amino acids; charged amino acids. Peptide bond formation (a condensation reaction) occurs between the carboxyl group of one amino acid and the amino group of the next amino acid generating a peptide backbone consisting of NCCNCCNCCNCC repetitions (in this example there are four amino acids covalently bonded. An amino acid always starts with an amino group [N-terminus] and ends with a carboxyl group [C-terminus]). Amino acids are abbreviated with a three-letter code or a one letter code: proline = Pro = P
Primary structure
The number of amino acids used and the sequence in which they are arranged (covalently bonded to each other) is called the primary structure. The primary structure determines all the properties of the resulting protein.
Peptide bond formation requires energy
Secondary Structure
As the polypeptide backbone consists of single covalent bonds, it is pretty flexible (rotation about the single bonds is possible, full rotation for the C-C bond, less so for the N-C bond). This allows polypeptides to fold into proteins. This occurs in two steps. First, hydrogen bonds form within the polypeptide backbone between the O of a carboxyl group and the H of an amino group. This can give rise to exactly two secondary structures that form very quickly after a polypeptide has been made: the alpha-helix and the beta-pleated sheet. Note that these hydrogen bonds do not involve the side chains at all! In the alpha-helix, hydrogen bonds form in the direction of the helix, generating a stable rod-like structure. Side chains point outwards, away from the helix. In the beta-pleated sheet, hydrogen bonds form within the plane of the sheet, generating a stable sheet. Side chains point away from the plane, up or down. There are no obvious rules, where you’ll find alpha helices and beta sheets within a protein. Protein structure determination is the only way to confirm the presence or absence of secondary structures within a protein.
Releases energy ie occurs automatically
Proline
Proline is a unique amino acid, because the side chain is covalently bonded to both the C and the N atom of the peptide backbone. This generates a kink in the peptide because the ring prevents free rotation of the N-C bond. Also, the backbone hydrogen bond cannot form, because the backbone N lacks a hydrogen. Therefore, formation of secondary structures (alpha helix, beta pleated sheet) is impossible. A proline is often the last amino acid of an alpha helix.
Tertiary Structure
Tertiary structure finishes the folding of a polypeptide and is mediated by side chain interactions.
ionic bonds
hydrogen bonds
disulphide bonds
hydrophobic interactions
Releases energy ie occurs automatically
Types of interaction contribute to tertiary structure formation
Four types of interaction contribute to tertiary structure formation: ionic bonds, hydrogen bonds, disulfide bonds, and hydrophobic interactions, all of them typically between amino acid side chains in the interior of the protein.
Disulfide bonds (Curling/uncurling hair)
Curling/Uncurling your hair: reduce/break disulfide bonds in keratin; then shape hair as you wish; finally oxidize/reform disulfide bonds to keep hair in new shape. In the keratin protein, you find coiled coils forming through hydrophobic interaction as discussed above, and additionally, you find disulfide bonds between alpha-helices to provide additional strength.
Hydrophobic interactions
Hydrophobic interactions are the most important determinant of protein folding/tertiary structure, because stretches of hydrophobic amino acids automatically rearrange towards the interior of a protein (away from water), whereas hydrophilic amino acids rearrange to be on the outside interacting with water. Important for many structural proteins is the coiled coil, which are two alpha- helices wrapped around each other. Hydrophobic amino acids at every 4th position generates a band of hydrophobicity running along the length of the alpha-helix and slowly rotating around it. Hydrophobic interaction then ensures that two such alpha helices come together just at that band resulting in the coiled coil (exactly like a rope that consists of intertwined strands). This structure occurs in proteins giving strength to tendons, hair, feathers. It can be a tertiary structure, if both alpha-helices are from the same polypeptide; if the two
alpha-helices are from two polypeptides, it would be called quaternary structure
Quaternary structure
Quaternary structure indicates several polypeptides interacting to form a bigger protein complex, e.g. hemoglobin consists of four individual polypeptides, the coiled-coil keratin consists of three alpha-helices. Many proteins, however, consist of only one polypeptide, that is, they are fully folded after tertiary structure formation.
Protein turnover
Proteins have a half-life ranging from minutes to weeks (average about 2 days). Protein turnover is very important, because proteins often get damaged: fever, pH change (lemon juice in milk changes the protonation state of COOH and NH4 groups; milk proteins precipitate, same happens in cheese production), other chemical changes. Example: egg becomes hard by boiling because proteins denature and get all entangled when they try to renature.
Protein Chaperones
Chaperones help proteins to fold properly after synthesis or after stress-related unfolding.
Nucleic acids
Nucleotides make RNA and DNA and serve functions in signaling and energy storage as monomers. A nucleotide is made up of a 5-carbon sugar, a nitrogenous base and a phosphate group. 4 nucleotides in RNA: cytosine, uracil (pyrimidines); guanine, adenine (purines). In DNA uracil is replaced by thymine. DNA lacks the hydroxyl group at carbon-2 (deoxyribose).
Nucleotide polymerization
Nucleotides polymerize via phosphodiester linkages (condensation reaction), with the 3’ hydroxyl group of the polymer forming a covalent bond with the 5’ phosphate group of the incoming nucleotide (5’ and 3’ refers to the position of the carbon atom, where the functional group is attached). This means that a long RNA/DNA molecule always starts with a 5’ phosphate group and ends with a 3’ hydroxyl, we say, polymerization occurs in 5’ to 3’ direction.
DNA
made of two antiparallel strands held together by hydrogen bonds between the bases.
Purine bases can only pair with pyrimidine bases, and more specifically, guanine (G) with cytosine (C), and adenine (A) with thymine (T). G-C forms three hydrogen bonds, and is therefore more stable than A-T, which forms two hydrogen bonds.
double helix with the sugar-phosphate backbone on the outside and the bases on the inside arranged like rungs on a ladder.
has a major groove and a minor groove. In the major groove, the two sugar-phosphate backbones are more widely spaced allowing DNA-binding proteins to recognize the bases in the interior. Very important for transcription factors, restriction enzymes etc., which recognize a unique DNA sequence.
carries the genetic information.
Base pairing and the double helical structures makes DNA much more stable than RNA.
RNA
came first in evolution,
can store and execute information.
can work like an enzyme catalyzing certain reactions, because it can fold into complicated 3D structures similar to proteins. The folding occurs because nucleotide bases within the same macromolecule pair via hydrogen bonds (in contrast to DNA, where nucleotide bases of different macromolecules pair).
A particularly important RNA structure found in many mRNAs is the stem-loop structure, which contributes to the regulation of mRNA function.
Anabolic reactions
Anabolic reactions require energy input. Examples are DNA replication, protein synthesis, making of starch. All condensation reactions are anabolic.
Catabolic reactions
Catabolic reactions release energy. Example: digestion of food molecules for energy generation. All hydrolysis reactions are catabolic.
Entropy
The tendency of energy to become evenly distributed or dispersed over time. Energy dispersal from being highly localized to being widely dispersed. The overall disorder in the universe increases as they proceed. The fact that all these processes only proceed in one direction is described in the second law of thermodynamics and is based on probability. The dispersing energy is the driving force for energy conversions! Energy conversions always result in a state of higher probability (a more disordered state).
Two ways a cell can release free energy/drive a chemical reaction?
with a chemical reaction creating disorder in the cell (e.g. digesting a polymer). This is a change in entropy in a closed system, called . Note, does not refer to the overall change of entropy in the entire universe.
with a chemical reaction releasing heat (enthalpy = - ), which generates disorder/disperses energy in the surrounding environment. In this case, the entropy outside the closed system increases.
What happens if H is negative and S is positive?
Heat is released ( H negative ) and disorder is increased (T positive ): this reaction is always spontaneous (exergonic), because G is always negative: .
What happens if H is negative and S is negative?
Heat is released (negative ), but disorder decreases (negative ): For example, in protein folding, heat is released, because favourable ionic bonds and other side-chain interactions occur, but the disorder decreases, because you get a nicely folded protein; because S depends on temperature, this process occurs only below a certain temperature; above a certain temperature, T becomes bigger than H, and then G is overall positive, therefore no protein folding above, say 50 degrees.
What happens if H is positive and S is positive?
Heat is used (positive ), but disorder increases (positive ): spontaneous above a certain temperature, e.g. dissolving NaCl in water. Heat is sucked away from the environment, that’s why the glass gets cold; heat is needed to break the strong crystal bonds; it’s the generation of disorder that drives the reaction.
What happens if H is positive and S is negative?
Heat is used and disorder decreases: never spontaneous (endergonic): . This applies to most anabolic reactions. Therefore, anabolic reactions can ONLY occur by coupling them to exergonic reactions to make the overall negative!
Thermodynamics
most reactions are reversible
adding different concentrations speeds up the forward/reverse reaction
At point of chemical equilibrium, rate of forward and reverse reactions are equal
The concentrations have to be taken into account to calculate the actual G in most cases
Direction of a reaction can be predicted if ∆G is known, but not the rate of reaction
Exergonic reactions need an activation energy
Standard free energy (delta G)
applies to 25 degrees Celsius and [1M]
Transfering energy in cells
All living cells use ATP for capture, transfer and storage of energy
Some free energy released by exergonic reactions is captured in ATP which can then drive endergonic reactions
ATP is particularly useful as energy currency because its ∆G is an intermediate between what you gain in respiration and what you expend in anabolism
polymerization
(e.g. peptide bond formation) requires energy input,
ATP hydrolysis
ATP+H20 = ADP + Pi + free energy
(breaking the bond between the 2nd and 3rd phosphate in ATP) releases energy.
reaction is very exergonic because ADP is constantly removed either by reforming ATP of by hydrolysis to AMP
Catalysis
a catalyst is any substance that speeds up a chemical reaction without itself being used up
most biological catalysts are enzymes
Only reactions with over -∆G can be catalyzed
∆G is not changed, rather an alternate pathway with lower activation energy is provided
Model of enzyme action
Initiation: reactants bind to the active site in a specific orientation, forming an enzyme-substrate complex
Transition state facilitation: Interactions between enzyme and substrate lower the activation energy required
Termination: products have lower affinity for active site and are released. Enzyme is unchanged after reaction
Inducing this transition state can happen by binding substrates in just the correct orientation, by exposing the reactants to altered charge environments that promote catalysis, or by inducing a strain on the substrate that facilitates breaking of a covalent bond (breaking polymers)
Enzyme cofactors
Some enzymes require cofactors (anything that is not an amino acid) in order to function: can be metal ions, small organic molecules
Saturated enzyme
Enzymes are saturated when all active sites are occupied; a further increase in substrate concentration will no longer increase the rate of product formation; this is the maximum speed of reaction or turnover rate, which varies widely from enzyme to enzyme.
Enzyme inhibition
Enzymes can be regulated competitively, with the regulator binding the active site, or allosterically, with the regulator binding somewhere else on the enzyme (allosteric = other site). Allosteric inhibition is much more efficient, because less inhibitor molecules are required (you need more competitor molecules than substrate molecules for effective competitive inhibition, but you only need more allosteric regulators than enzymes, because nothing else binds at the allosteric site).
Competitive inhibition
regulator binding to active site instead of substrate
Allosteric Regulation
regulator binding somewhere else on the enzyme
Activation: the active site becomes available to the substrates when a regulatory molecule binds to a different site on the enzyme
Inhibition: the active site becomes unavailable to the substrates when a regulatory molecule binds to a different site on the enzyme
(allosteric = other site).
Allosteric inhibition is much more efficient, because less inhibitor molecules are required (you need more competitor molecules than substrate molecules for effective competitive inhibition, but you only need more allosteric regulators than enzymes, because nothing else binds at the allosteric site).
Carbohydrates
Monosaccharides
Disaccharides
Polysaccharides
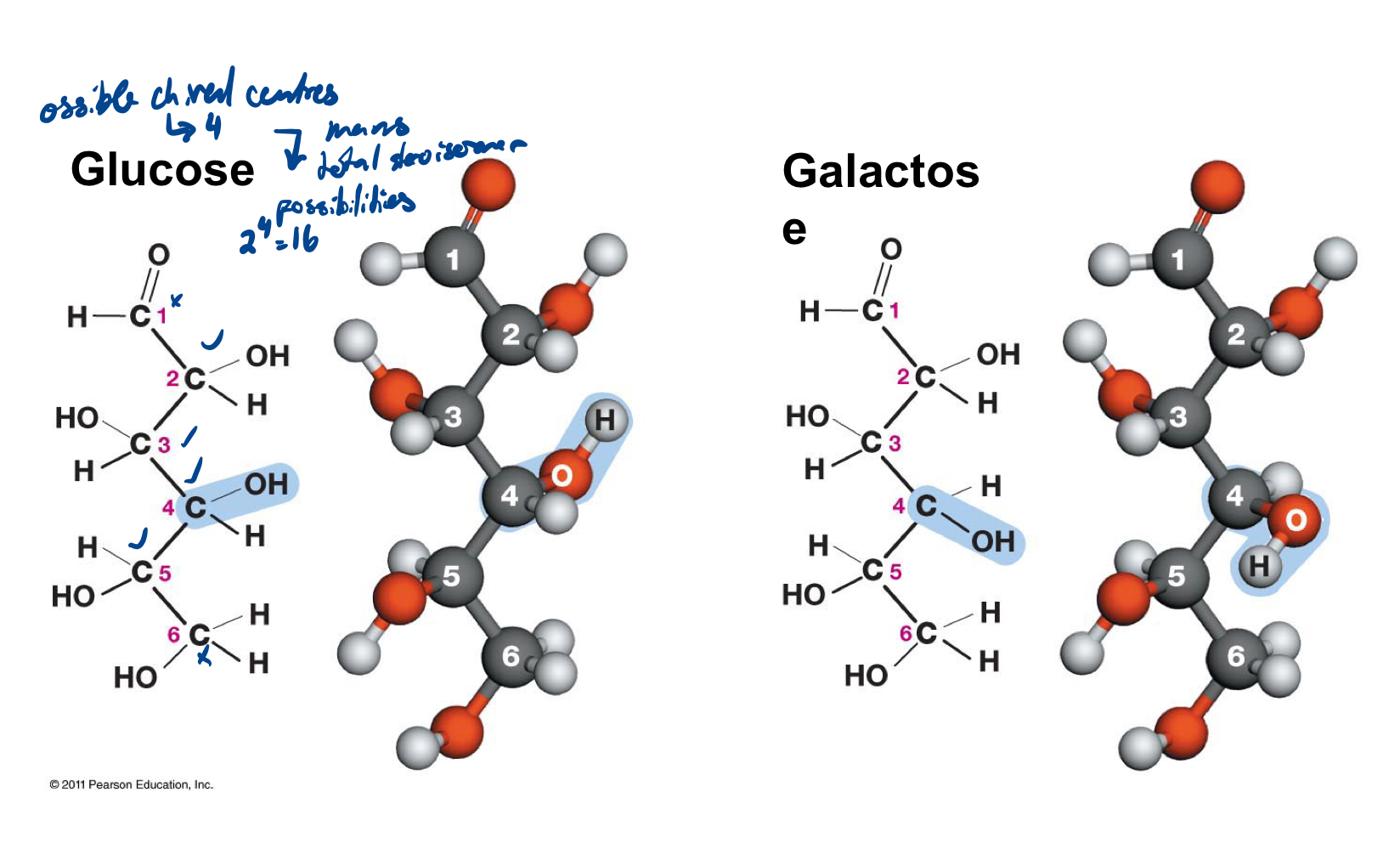
The typical structure is a multiple of CH2O, e.g. C3H6O3. You always find two functional groups in a sugar: one carbonyl group and several hydroxyl groups. Aldose and ketose are isomers, same formula, but different structure. Important to keep in mind: all isomers differ in their covalent bonds (that is, optical isomers also have a different structure).
Optical isomers (stereoisomers) - mirror images, must have chiral centre
Glucose and galactose
Glucose and galactose are two different optical isomers of this 6-carbon sugar.
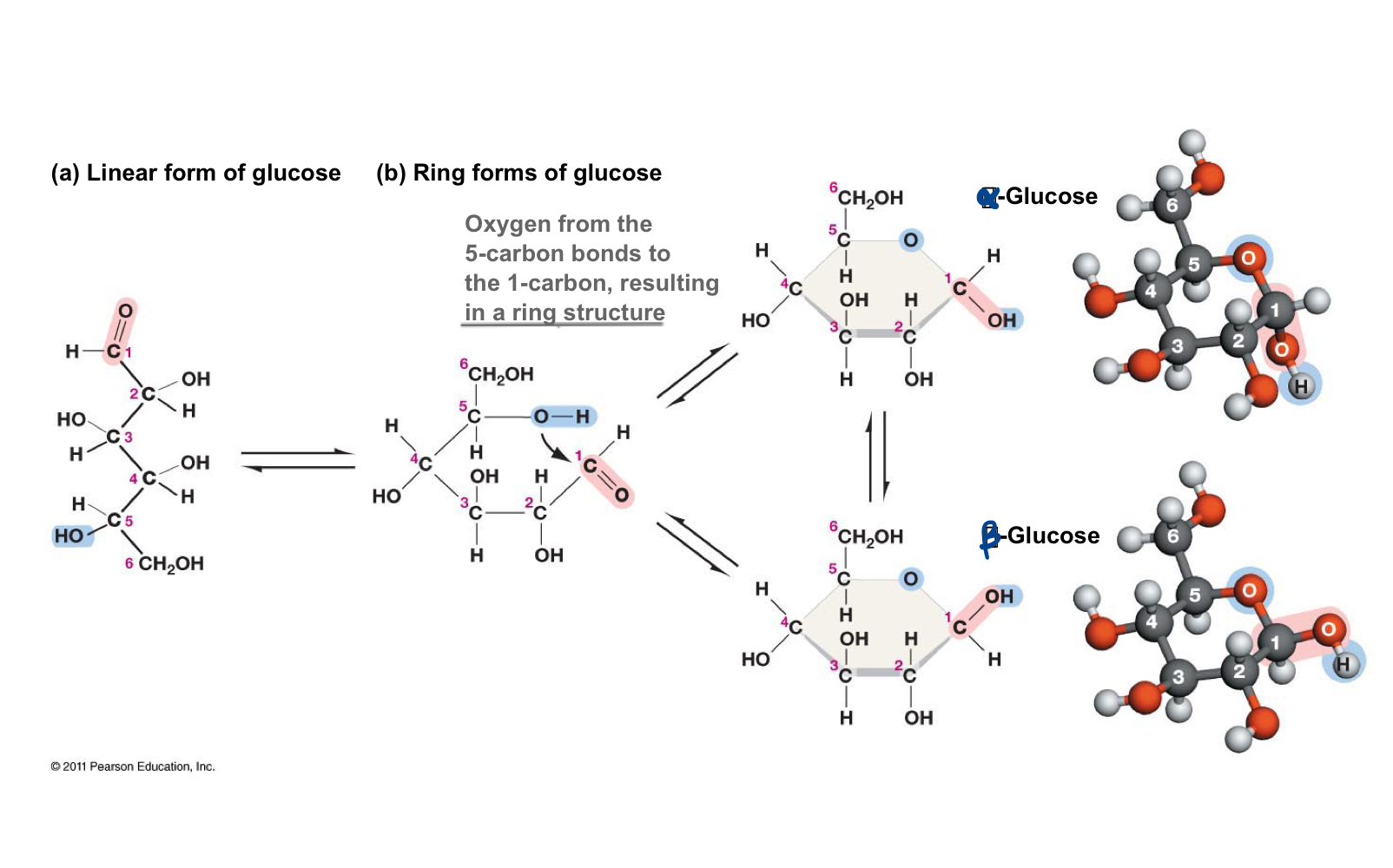
Glucose linear form to ring form
In solution, straight-chain glucose forms another covalent bond to become the ring form of glucose. Because this converts C-1 from a symmetric to an asymmetric carbon atom (C-1 in the ring form has four different groups attached to it, versus 3 in the straight chain form), you can get two glucose isomers, alpha- and beta- glucose.
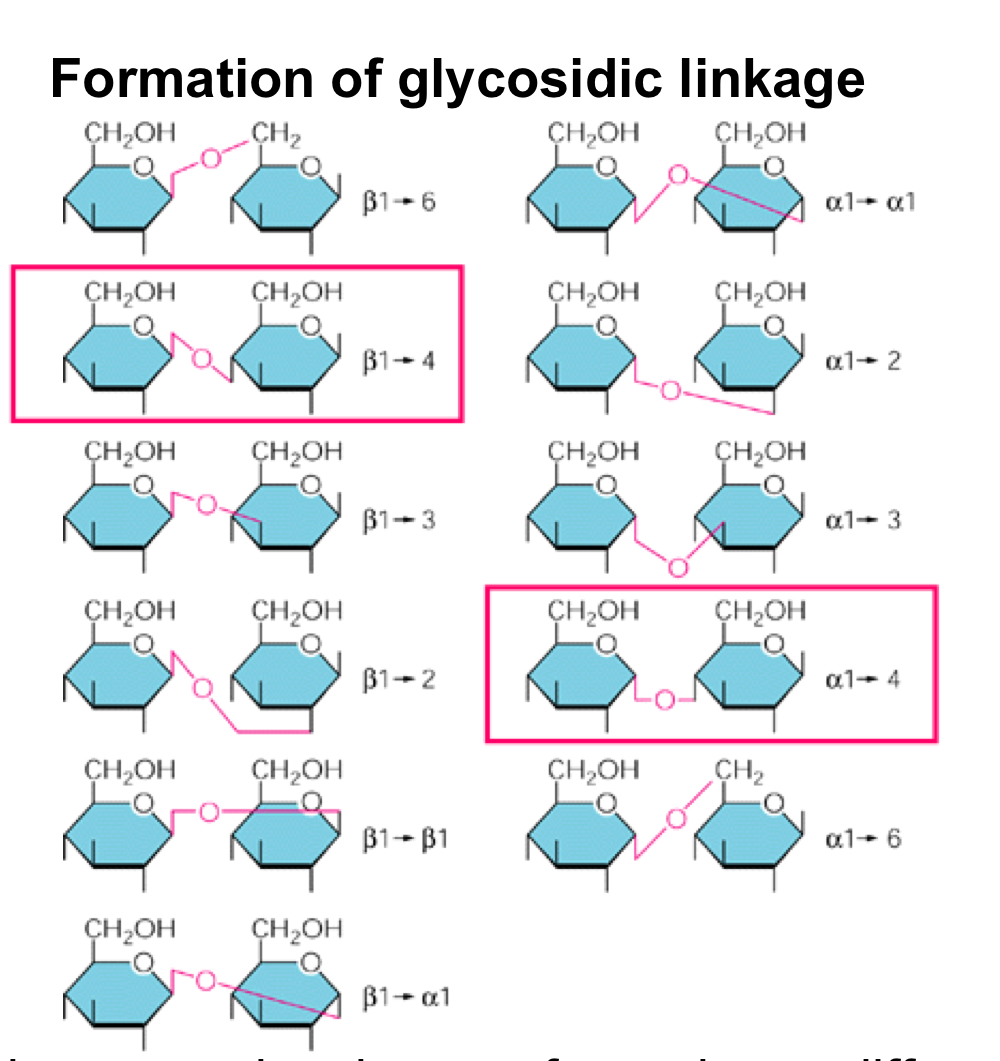
Glycosidic linkage
Sugar polymers (polysaccharides) are formed by covalent bonds called glycosidic linkages between the C-1 of one sugar with any OH-group of the second sugar, but only two links are interesting to us: alpha1,4 giving rise to maltose and eventually starch, and beta1,4 giving rise to cellobiose, and eventually cellulose.
In starch, bulky CH2OH groups are all on the same side, bending the polymer resulting in the shape of a spiral.
When starch is unbranched is is amylose in plants; moderately branched = amylopectin in plants; highly branched = glycogen in animals
On the other hand, the second glucose in cellobiose is flipped (with the CH2OH group pointing downwards) compared to starch, which has huge structural implications. You now get a very symmetrical, straight molecule = better hydrogen bonding. Cellulose = always unbranched, parallel strands joined by hydrogen bonds
Lipids
insoluble in water
Roles:
energy storage! (fats and oils)
cell membranes! (phospholipids)
Capture of light energy (carotinoids)
Hormones and vitamins (steroids and modified fatty acids)
thermal insulation
electrical insulation of nerves
water repellency (waxes and oils)
Fats and oils
The most important lipids are fats (solid) and oils (liquid), which consist of three fatty acids and one glycerol connected by covalent bonds.
Fatty acids on their own have one carboxyl group and a long hydrocarbon chain.
Fatty acids are amphiphilic (hydrophilic carboxyl group plus hydrophobic hydrocarbon chain) and are found in soap.
Fats provide more energy per weight than starch but take longer to mobilize; long- term storage.
Fats form via dehydration reactions
Phospholipids
Phospholipids are also amphiphilic: a hydrophilic head group and two hydrophobic fatty acid tails attached to glycerol. Phospholipids have the unique capability to self- assemble into lipid bilayers with the hydrophobic fatty acid tails pointing inward away from water.
Lipid bilayer
Made from phospholipids
To avoid exposed edges, lipid bilayers will assemble into a globular compartment. This is energetically most favourable, as no more hydrophobic parts are exposed to water.
cell membranes
very fluid, with phospholipids in constant lateral motion
Unsaturated fatty acids in lipid bilayers increase the fluidity and permeability of the membrane.
Unsaturated Fatty Acids
Some fatty acids contain double bonds; then they are called unsaturated fatty acids. Double bonds cause kinks in fatty acid tails. Therefore, they cannot be packed as closely together as straight, saturated fatty acids. This is the reason butter is solid at room temperature (only straight-chain fatty acids in butter fat), and oils are fluid (many unsaturated fatty acids: double bonds making kinks).
Membranes
very fluid in order to allow proteins inserted into the membrane to interact with each other.
peripheral membrane protein on outside, integral/transmembrane proteins on inside
Usually membrane proteins are one or multiple alpha-helices.
need a stretch of hydrophobic proteins long enough to cross the lipid bilayer
This works if all amino acids making up the alpha-helix are hydrophobic, because their side chains are pointing outward interacting with the lipids, whereas the hydrogen bonds needed to stabilize the alpha-helix are all along the length of the alpha-helix cylinder.
The insertion of membrane proteins into the lipid bilayer introduces an asymmetry, because integral membrane proteins can only move laterally, not vertically. This means the side of the protein pointing outward never changes, the protein just moves around within the plane of the membrane (like phospholipids).
Freeze Fracture
Freeze fracture splits membranes into two lipid leaflets Because the interior becomes a weak point with only LDF holding the two leaflets together
Selective permeability
membranes are selectively permeable
Freeze fracture splits membranes into two lipid leaflets Because the interior becomes a weak point with only LDF holding the two leaflets together
main function of membranes is to serve as a barrier and selectively transport molecules across the membrane
Gases like oxygen, and small polar molecules like water pass freely across the membrane. Large charged molecules can barely cross the membrane, and ions not at all.
Diffusion
the passive mixing of substances resulting in net transport along a concentration gradient
Reason: random walk (Brownian motion) of individual molecules due to thermal motions and collisions
rate is determined by temp, size of molecule, and steepness of concentration gradient
Osmosis
diffusion of water across a selectively permeable membrane
water moves from regions of low solute concentration to high solute concentration
Hypertonic solution
higher solute concentration outside membrane
net flow of water out
Ocean water cannot quench your thirst, because water will flow out of your cells to dilute the ocean water.
Hypotonic solution
higher solute concentration inside membrane
net flow of water in
Isotonic solution
same solute concentration as inside the cell, therefore no net water flow
Passive transport: Facilitated diffusion
depends on two types of membrane proteins: channel proteins and carrier proteins
electrochemical gradient (high concentration of Na)
gated channel proteins (always gated: closed as default state): ions flow along their electrochemical gradient when the channel is open
Carrier protein Glucose carrier: binding of sugar to either side causes a conformation change in the carrier protein resulting in transport of sugar to the other side. Sugar can be transported in both directions (reversible)! Carrier proteins can be saturated because they must bind their substrate, i.e. at one point the rate of diffusion cannot increase further because all carrier proteins are occupied.
Both channel proteins and carrier proteins (all facilitated diffusion) display saturation kinetics, that is, there is a concentration at which maximal transport is reached. Note, that saturation occurs much earlier in a carrier than in a channel protein.
Active transport
moves substances against their concentration gradient, and therefore requires energy.
Primary active transport
directly relies on ATP hydrolysis to overcome the +∆G of transporting something against its concentration gradient,
Most important example: Sodium potassium pump in animals to counteract hypotonic drinking water, not found in plants, because they have a stable cell wall that prevents bursting of cells upon water inflow. The sodium potassium pump moves three sodium ions out and two potassium ions in (a net movement of one ion out). This controls osmolarity, generates the resting potential, and sets up ion gradients.
Secondary Active transport
uses the energy from the concentration gradient set up by primary active transport.
Secondary active transport systems use gradients established by the sodium potassium pump. An important example is the sugar sodium co-transporter, which uses the energy from sodium inflow to transport sugar into cells with a high interior concentration of sugar. The best example is the cells lining the gut, whose task is to take up nutrients from the food we eat. To maximize the uptake of sugar, these cells use a sodium sugar co-transporter to take up every sugar molecule from the gut lumen (where the digesting food passes by). On the other side the sugar crosses the membrane into the extracellular fluid with a sugar carrier, because these sugar molecules are constantly removed/shipped to the rest of our body.
Cells in human body
about 40 trillion: 4x1013
Why are cells small?
because most chemical reactions in cells require diffusion
diffusing molecules don’t travel long distances because of their random walk
distance traveled is proportional to square root of time
Everything inside a cell has to diffuse across the cell membrane, therefore a substance like glucose would take too long to encounter the first glycolytic enzyme, if the cell volume were big. (reason why eukaryotes are bigger than prokaryotes). Eggs (only exception to size rule) can be big because they just store stuff
Prokaryotes
can live in any environment, oxidize anything, have the greatest metabolic diversity of all organisms, and are much smaller than eukaryotes.
Their DNA sits in a nucleoid, which is not surrounded by a membrane.
no nucleus and few membrane-enclosed compartments
Eukaryote
Eukaryote means “real nucleus” and indicates the main distinguishing feature of eukaryotes, a nucleus surrounded by a nuclear envelope consisting of two membranes.
Compartmentalization is the key to eukaryotes’ ability to have much bigger cells.
Many internal membrane systems that create very small, specialized spaces where diffusion can again happen very quickly.
Compartmentalization is largely mediated by the endomembrane system giving rise to several organelles: nucleus, ER, Golgi, vesicles, and lysosomes.
Organelles in Animal Cells
Unique: centrioles
Organelles in Plant Cells
Unique: cell wall (cellulose), chloroplast
Endo-membrane system
Note that the interior of vesicles, Golgi, ER (their lumen), is equivalent to the extracellular space (see colour code in figure).
Orientation of membrane is always preserved: This can be understood by looking at how vesicles bud off from organelles and fuse with the cell membrane (the membrane leaflet facing the cytosolic side always faces the cytosolic side, in ER, Golgi, vesicles, and cell membrane).
Nucleus
surrounded by nuclear envelope made up of two membranes.
contains our DNA stored as chromosomes and the nucleolus, where ribosomes are made from rRNA and proteins (ribosomal proteins are made in cytoplasm and brought back to the nucleolus to be assembled).
Assembled ribosome subunits and mRNA are transported through nuclear pores to the cytoplasm.
Nuclear proteins like transcription factors are brought into the nucleus through nuclear pores.
rRNA is transcribed into mRNA or rRNA by RNA polymerases
rRNA is not translated; it directly folds into a 3D structure
Ribosomes are made of 4 rRNAs and about 80 proteins in the nucleolus (RNAs largely determine structure and function)
The Rough ER
close to the nucleus, and its membranes are contiguous with the nuclear envelope.
Membrane proteins are made at the rough ER.
rough because of the ribosomes attached to the outside of the ER membrane. Ribosomes become attached when they start translating a membrane protein. The first few amino acids encode a signal sequence that targets the ribosome to the rough ER, ensuring that the freshly made membrane protein is immediately inserted into the ER membrane during translation (with the help of other proteins).
Secreted proteins are released into the ER lumen (the interior of the ER).
Signal hypothesis
Signal sequence is synthesized
signal binds to SRP
SRP binds to receptor
protein is synthesized into ER
protein synthesis is complete
The Smooth ER
All membrane lipids are made in the smooth ER.
Detoxification also occurs in the smooth ER and usually means oxidation, for example adding hydroxyl groups that make a molecule more hydrophilic, such that it can be excreted. (more smooth ER in liver cells of drug addicts)
has tubules to increase surface area
Golgi
stack of membranes generated by vesicles coming from the ER and fusing on the cis face with the Golgi, and other vesicles budding off on the trans face.
While proteins and lipids are in the Golgi, they are further modified, e.g. sugar side chains are added, and proteins are sorted to reach their proper final destination.
The Golgi is a distribution centre. Proteins and lipids move through the Golgi from the cis to the trans side, because vesicles continuously fuse on the cis side (coming from the ER), and continuously bud off from the trans side (to move on to the cell membrane, e.g.).
trans side faces plasma membrane
Secretory pathway
Ribosome deposits protein in ER
Protein exits ER
Protein enters Golgi for processing
Protein exits Golgi
Protein exits cell (exocytosis)
Pulse Chase experiment
demonstrated the traveling of proteins through the endomembrane system.
add radioactively labeled amino acids for a short time like 1 min to the culture medium (the pulse),
then wash the cells and add unlabeled medium for various amounts of time (the chase) before looking at the cell by electron microscopy.
Immediately after labeling, radioactively labeled proteins will show up in the rough ER, later on in the Golgi, secretory vesicles, and eventually in the extracellular fluid.
Receptor-mediated endocytosis
for moving bigger stuff like lipid vesicles or entire proteins into the cell
Macromolecules bind to receptors
early endosome forms
processing
digestive enzymes received
mature lysosome
Phagocytosis
for moving entire organisms into the cell
Detection (bigger thing ex: bacteria)
Phagosome formation
Delivery to lysosome and digestion
Release of particles into cytosol
Autophagy
Self-eating (in case of damage or starvation)
organelle surrounded by membrane
delivery to lysosome
recycling
Protein sorting and vesicle transport
proteins are tagged
proteins are sorted
vesicles bud
proteins interact with receptors
delivery
Happens in Golgi
Mitochondrion
most of ATP is produced here
the citric acid cycle occurs in the matrix
respiratory enzymes are integral membrane proteins located in invaginations of the inner membrane. These invaginations (called cristae) maximize the area for inserting membrane proteins.
Chloroplasts
convert light energy to chemical energy in photosynthesis.
carbon fixation occurs in the stroma yielding sugars, amino acids, and all fatty acids
the light reactions of photosynthesis take place in stacks of membranes called thylakoids, which have separated from the inner membrane, they are full of enzymes.
Plastid
Mitochondria and Chloroplasts
Both have outer and inner membranes
The space enclosed by the inner membrane (called matrix in mitochondria and stroma in chloroplasts) is where you can find DNA (a small circular genome), RNA and ribosomes.
descendants of bacteria that were at one point taken up by an ancestral eukaryotic cell through phagocytosis. Evidence for this is the double membrane. They also have their own genome with genes more similar to bacterial genes, and own ribosomes more similar to bacterial ribosomes. They also have a unique system for protein and lipid import again suggesting they evolved separately from the endomembrane system.
Cytoskeleton
consists of structural elements important for cell shape and movement.
The most important structural proteins are actin filaments and microtubules.
Both animal and plant cells have a cytoskeleton.
Crowded!
Actin Filaments
polar with a plus and minus end,
they polymerize and depolymerize from monomers through non- covalent protein-protein interactions. (monomer refers to a single actin protein, one polypeptide)
Many actin- binding proteins regulate actin stability and polymerization.
Actin filaments are found just below the cell membrane (cortically) and are responsible for cell shape and cell shape changes.
Together with myosin, they also mediate muscle contraction, cell shape changes, cytoplasmic streaming and cytokinesis. This works because myosin II forms bipolar complexes with myosin heads at opposite ends. These myosin heads walk toward the plus end (and only the plus end) of actin filaments. A bipolar myosin II complex bound to antiparallel actin filaments leads to contraction of these filaments.
Actin-myosin contraction
actin and myosin interact to cause movement
myosin “head” attached to actin and moves, and the filament slides
coiled-coil tail domains of myosin II can interact to form antiparallel bipolar complexes
these may contain many myosin molecules, as in thick filaments of skeletal muscle, or as few as 2 myosins
myosin heads can move along actin filaments in only one direction
antiparallel actin filaments can “contract”
Ex: cytokinesis in animals (pinching membrane in two); cytoplasmic streaming in plants (move cytoplasm around cell)
Microtubules
much bigger in diameter and form a cylinder of alpha beta tubulin dimers. But like actin filaments, microtubules are polar with a plus and minus end, and there are many microtubule-binding proteins that regulate growth or shrinkage of microtubules.
organize cells in that they move organelles and provide tracks for intracellular transport.
Vesicles move along MTs to their destination, an extreme example is vesicles moving along axonal MTs to the synapse of nerve cells.
MTs also move chromosomes during cell division.
In contrast to actin, MTs have two different motor proteins, the kinesin motor walking in one direction and the dynein motor walking in the opposite direction, a necessary requirement for efficient intracellular transport.
MTs also form cilia and flagella made up of 9 doublet MTs and 2 central MTs.
Kinesin
has a tail, stalk and head
walks along a microtubule track
every step requires energy
(dynein walks in opposite direction!)