Cartões: Fisiologia sensorial | Quizlet
1/78
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced |
|---|
No study sessions yet.
79 Terms
Sistema Sensorial
Deteta e transmite informação quer do meio externo, quer do meio interno. Esta deteção pode ser feita por neurónios, neurónios estes que se especializam na deteção de informação periférica, ou por células sensoriais, que não são neurónios mas comunicam com eles.
Se a informação for detetada por estas células, há uma sinapse entre esta célula especializada e o neurónio, sendo o neurónio a enviar informação até ao SNC.
É um sistema constituído por células sensoriais recetoras, vias nervosas aferentes e pelo encéfalo.
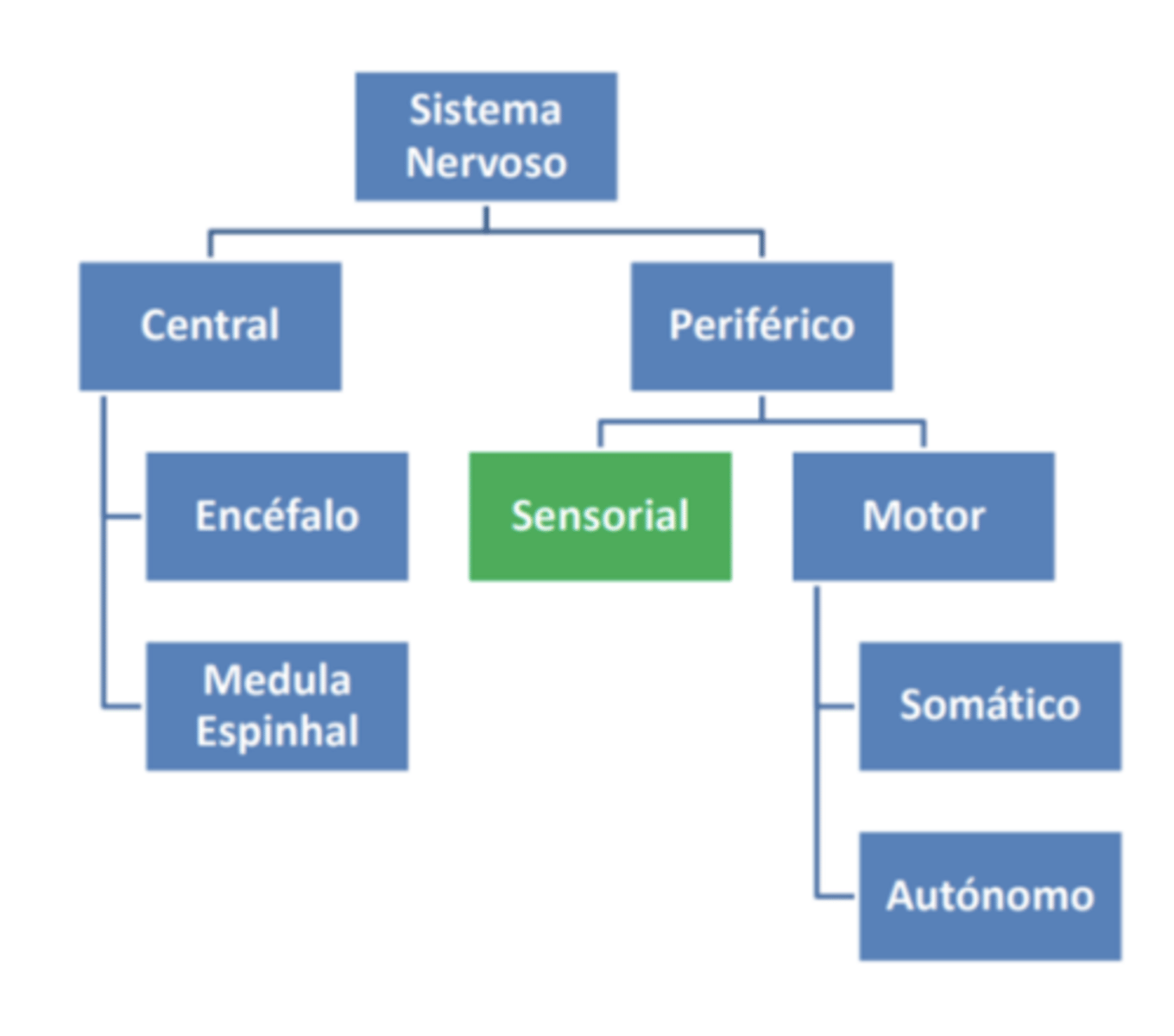
Classificação
Este sistema pode ser subdivido em sistema somatossensorial e em sentidos especiais.
O sistema somatossensorial subdivide-se no sistema exteroceptivo (tato), no sistema proprioceptivo (nos músculos e articulações, dando informação sobre o grau de contração do músculo, bem como o grau de distensão dos tendões), e no sistema enteroceptivo (cada órgão interno tem o seu; nas paredes de órgãos).
Nos sentidos especiais encontramos os sentidos químicos (olfato e paladar), a visão, a audição e o equilíbrio (ambos criados no ouvido interno).
Sistema Sensorial funcionamento
Neste sistema, começamos por detetar um estímulo, que tem de atuar sobre uma célula recetora específica. Este estímulo tem de ser convertido, na célula recetora, numa informação de natureza um pouco diferente. Tem de haver transdução sensorial dentro da célula recetora para a informação ser transformada em informação elétrica.
De seguida, essa célula recetora gera potenciais de ação que são propagados ao longo de vias aferentes (fibras sensitivas), e a informação é levada ao SNC. No SNC ocorrem dois fenómenos: por um lado, ocorre a criação da sensação, e por outro a capacidade do indivíduo conseguir pensar sobre essa sensação, aperceber-se das circunstâncias que motivaram essa sensação - a perceção.

Transdução Sensorial
Em função dos diferentes tipos de estímulos que as nossas células podem receber, nomeamos de forma diferente os recetores.
• Na visão detetamos, ao nível da retina, fotões; os recetores são fotorrecetores.
• Dentro dos órgãos viscerais no sistema músculo esquelético e na pele, detetamos variações na tensão e torção mecânica de uma região através dos mecanorrecetores.
• Variações de temperatura são sentidas por termorrecetores.
• Qualquer região que receba uma substância química tem quimiorrecetores.
Vários destes recetores conseguem originar uma informação diferente se a quantidade de estímulo for excessiva; por exemplo, se a deformação mecânica for excessiva ao
ponto de irritar ou criar outro tipo de lesão numa célula recetora, ela deixa de emitir a sua informação original e passa a emitir a informação de dor. Portanto, mecano, termo
e quimio recetores podem funcionar como nociceptores, recetores da dor.
Quimiorrecetores
Encontramos na cavidade nasal e oral, no SNC e no corpo carotídeo.
O corpo aórtico e os corpos carotídeos detetam diminuições de pO2, aumentos de pCO2 e diminuições de pH.
Os quimiorretores centrais encontram-se em vasos sanguíneos que profundem o encéfalo, detetando também diminuições de pO2, aumentos de pCO2 e
diminuições de pH.
Barorrecetores/Mecanorrecetores
Detetam uma variação mecânica associada a uma pressão.
Encontramos na pele, sistema músculo esquelético, dentro do músculo e nos tendões ao nível da inserção dos ossos, nas células-cabelo do nosso ouvido interno (células que detetam o som e variação da posição da cabeça, sendo ativadas por variações na posição dos seus cílios), nos órgãos de Golgi o nos vasos carotídeos e aórticos, onde detetam variações de pressão arterial.
Termorrecetores
Não detetam temperaturas absolutas mas sim variações de temperatura.
Encontram-se em mucosas como a cavidade oral, a pele, e mesmo vasos sanguíneos.
Em função das variações da temperatura do sangue, a informação será enviada para o hipotálamo, que poderá arquitetar um conjunto de respostas de termorregulação,
aumentando ou diminuindo a temperatura.
Fotorrecetores 1
Encontram-se apenas na retina.
Recetores - Adaptação
Estas células recetoras têm capacidade de adaptação ao próprio estímulo. Em função da rapidez com que uma célula recetora é capaz de se adaptar, classificamos estes recetores em recetores fásicos e em recetores tónicos.
Nos recetores fásicos (adaptação rápida), em virtude da aplicação de um estímulo, surgem muitos potenciais de ação; ao fim de algum tempo, a frequência de potenciais de ação começa a diminuir - significa que a célula deixou de responder perante a presença do estímulo, ou começou a responder mais lentamente.
Estes recetores respondem rapidamente ao estímulo mas também deixam rapidamente de responder ao mesmo estímulo.
Nos recetores tónicos (adaptação lenta) geram-se também potenciais de ação, mas a uma frequência menor do que a mencionada acima. Essa frequência nunca diminui entre o ponto de aplicação e o ponto de remoção do estímulo - o recetor responde sempre da mesma maneira ao estímulo.
Quando um recetor entra em modo de funcionamento como um nocicetor (recetor da dor), ele transmite esta transformação de uma forma lenta e persistente.
Potencial Recetor
Nome especial que damos a potenciais gradativos que ocorrem nestas células recetoras.
Gráfico potencial membrana vs tempo, em que estamos a aplicar um estímulo com 4 intensidades diferentes em 4 experiências diferentes, para se atingir um limiar de excitabilidade; ou se dá um estímulo muito forte, ou em alternativa têm de ser vários estímulos fracos que, por somação temporal e espacial, atingem o limiar.
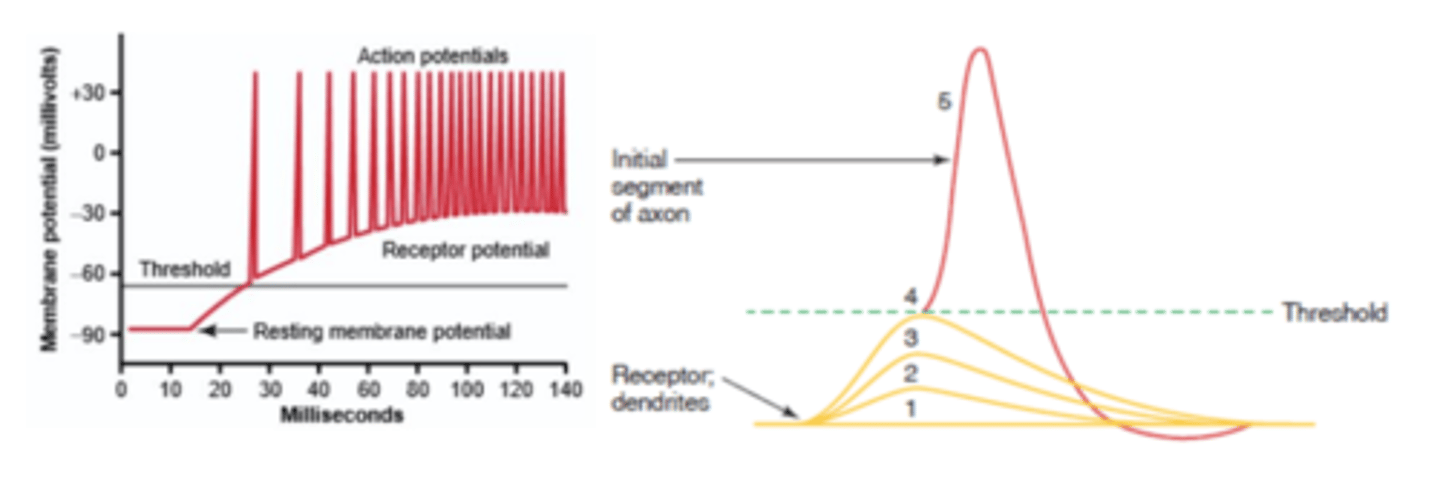
Como é que uma célula recetora codifica a intensidade do estímulo e vai portanto codificar a intensidade da sensação propriamente dita?
Quando o estímulo é muito intenso, não vai ter dificuldade em atingir o limiar de excitabilidade; o que vai acontecer é que a frequência de potencias de ação vai ser muito grande.
Quando o estímulo é pouco intenso, a sensação vai ser pouco intensa, e por isso o número de potenciais de ação vai ser menor por unidade de tempo.
Um estímulo mais intenso gera uma frequência de potencias de ação maior e, portanto, uma sensação mais intensa; um estímulo menos intenso gera uma menor frequência de potenciais de ação e, portanto, uma sensação menos intensa.
Células Recetoras
Podem ser simples, se forem neurónios propriamente ditos, ou células especializadas, que geralmente são células epiteliais ou derivadas delas, que estão em relação sináptica química com neurónio aferente propriamente dito.
Paladar
Consiste na capacidade de o organismo identificar e consumir alimentos com valor nutritivo e evitar toxinas e materiais indigeríveis. É a capacidade do organismo detetar substâncias potencialmente úteis e substâncias potencialmente tóxicas.
Há substâncias tóxicas que têm um sabor doce: chumbo; já foi usado para conservar alimentos, tendo havido muitos casos por intoxicação por chumbo.
O paladar é dado ao nível da língua, faringe, uma porção da laringe e esófago. Consiste na deteção dos gostos, sendo os cinco gostos base o doce, salgado, amargo, azedo e umami (sabor agradável, saboroso ou com sabor a carne). Os gostos acessórios são o caráter gorduroso, metálico ou sabor da própria água.
Índice de Sabor
Para classificar uma determinada substância quanto ao gosto base que ela cria, escolhemos substâncias que vão funcionar como referência. Todas as substâncias com índice superior a um são mais intensas, e inferior a um são menos intensas.
Os edulcorantes são usados porque são mais doces que as substâncias doces da natureza, que são calóricas.
Limiar de Excitabilidade
As substâncias azedas e amargas são aquelas que numa concentração mais pequena são capazes de ativar as células recetoras. O organismo humano está preparado para
reconhecer muito mais cedo substâncias azedas e amargas, porque estas substâncias, na sua maior parte, são potencialmente tóxicas, devendo ser evitadas e reconhecidas
mais cedo.
Língua
A maior parte das células recetoras estão concentradas na língua, na superfície ou face superior da língua. Neste órgão, temos três tipos de pupilas: fungiformes, foliadas e circunvaladas. Todas as regiões da língua conseguem detetar os cinco gostos base.
Botões Gustativos e Recetores
Cada botão consiste num aglomerado de células neuroepiteliais que estão em
comunicação sináptica com fibras aferentes. Estes botões são enervados por fibras de
vários pares cranianos.
Estas células designam-se de I a IV: I, II e III são células recetoras, as células IV são
basais, indiferenciadas, encontrando-se no polo inferior de cada botão.
• As células do tipo I detetam substâncias salgadas, deixam entrar catiões e geram
potencias de ação.
• As células II e III estão verdadeiramente em sinapse química com os neurónios
aferentes.
• As células do tipo II detetam os gostos doce, amargo e umami, e o
neurotransmissor usado por estas células é o ATP; libertam ATP sobre recetores
purinégicos destas fibras e criam o potencial de ação.
• As células do tipo III detetam o sabor azedo, e o transmissor usado é a
serotonina.
• As células do tipo IV estão na base do botão gustativo e são capazes de regenerar
todas as outras quando forem lesadas.
Os recetores para azedo e salgado são ionotrópicos. Os recetores para as moléculas
doces, amargas e umami são metabotrópicos.
Salgado
As moléculas que dão um sabor salgado são catiões metálicos (sódio, potássio, lítio...).
Nas células do tipo I existem recetores ionotrópicos que deixam entrar estes catiões, criam uma despolarização, atinge-se o limiar de excitabilidade e ocorre o potencial de
ação.
Azedo
O azedo consiste na deteção de ácido, muitas vezes proveniente de alimentos deteriorados/contaminados por bactérias.
Os protões podem entrar diretamente nas células tipo III e despolarizar a célula.
Se a substância ácida ainda estiver neutra quando entrar na célula, se for suficientemente pequena, pode sofrer difusão simples para dentro da célula e ionizar-se em bases conjugadas e protões, despolarizando diretamente a célula e fechando canais de
potássio.
Doce
As moléculas doce, amargo e umami têm a mesma via de transdução de sinal que ocorre via recetor metabotrópico.
Via comum para doce, amargo e umami:
a molécula atua sobre um recetor metabotrópico e ativa-o; quando o recetor fica ativo, sofre uma alteração de conformação que motiva uma interação com uma proteína G. Estas proteínas G heterodiméricas estão ancoradas ao folheto interno da membrana citoplasmática por cadeias acíclicas.
Amargo
Os ligandos que conferem informação sobre o amargo têm uma estrutura muito variada.
Há indivíduos que têm maior sensibilidade e menor sensibilidade para o amargo; os que tiverem uma sensibilidade muito elevada têm um limiar de excitabilidade muito baixo
(supertaster); já os indivíduos com um limiar elevado são pouco sensíveis (nontaster).
A sensibilidade ao amargo vai influenciar as escolhas alimentares dos indivíduos, sendo também hereditária.
Umami
As moléculas que garantem o caráter de umami são aminoácidos, principalmente o L-glutamato e 5'-ribonucleótidos, que são formados aquando do processo de cura de um alimento. Estes ligandos umami são adicionados frequentemente aos alimentos, quer em receitas caseiras quer a nível industrial, para melhorar o sabor dos alimentos.
Sabor
Não só os gostos base influenciam sabor do alimento: também a textura, a temperatura e o cheiro influenciam.
Efeito da temperatura
Os refrigerantes que contêm muito açúcar têm o intuito de tornar as pessoas mais adictas aos refrigerantes, pois o açúcar promove a libertação de substâncias que fazem as pessoas sentirem-se bem (endorfinas). No entanto, para ingerir elevadas quantidades de açúcar, é necessário baixar a temperatura da bebida ("deve ser consumido fresco").
Olfato
O olfato é o sentido filogeneticamente mais antigo dos seres vivos. Os estímulos são moléculas facilmente efusivas pelo ar. A cavidade nasal está equipada por um epitélio olfativo que contém quimiorrecetores.
O centro integrador do olfato é o córtex olfativo, que está distribuído em várias regiões encefálicas.
Alterações na capacidade olfativa podem conduzir a alterações de apetite, intoxicações alimentares, ou incapacidade de detetar moléculas tóxicas na atmosfera.
Aumenta ou reduz assimilação dos aliementos
Classificação de Odores
As moléculas odoríferas têm morfologias muito diferentes e, portanto, moléculas diferentes podem originar o mesmo odor ou odores diferentes. Também a mesma molécula em diferentes concentrações pode criar odores diferentes. A classificação de odores é empírica.
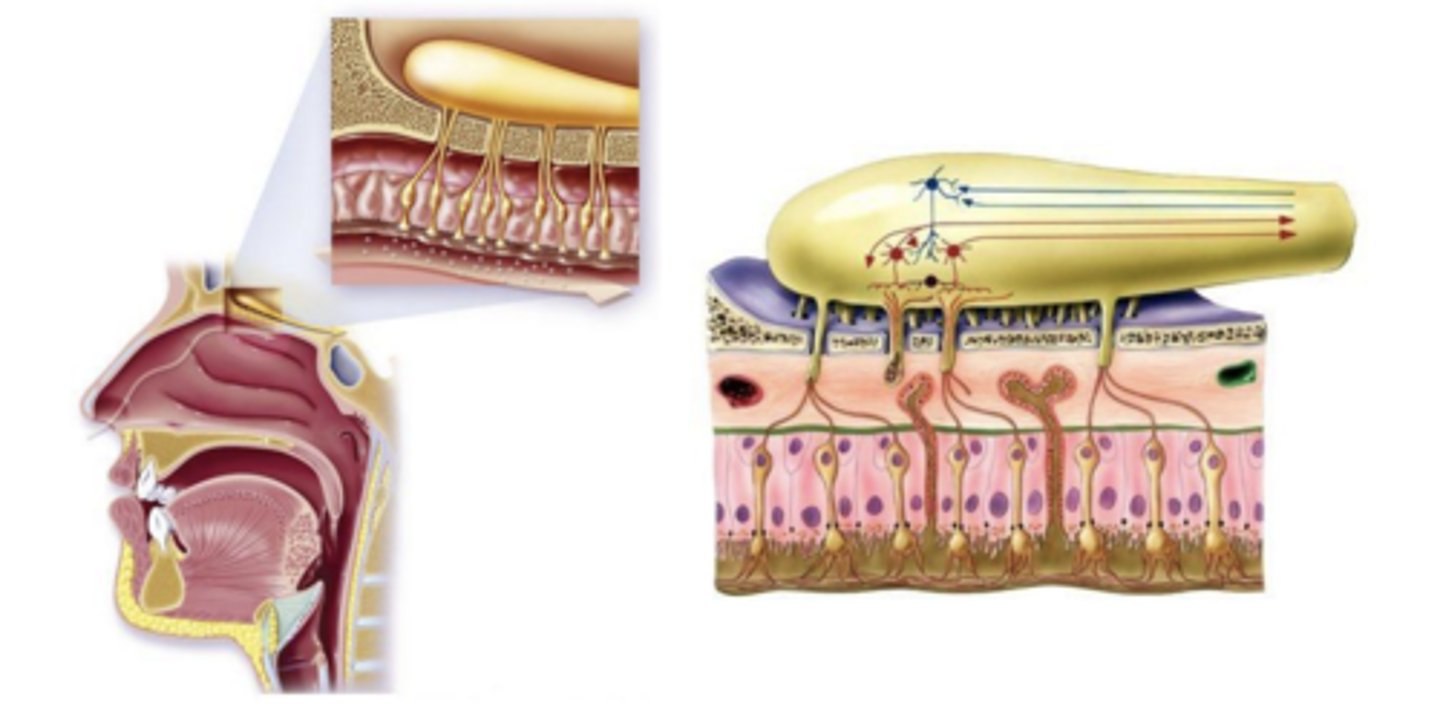
Cavidade Nasal
As moléculas são inaladas e sofrem aspiração, que pode ser ortonasal, se as moléculas entrarem pelas narinas, ou retronasal, se as moléculas entrarem primeiro na cavidade oral e só depois ascenderem até à cavidade nasal.
Este epitélio olfativo estende-se desde a placa crivosa até à concha ou corneto nasal inferior. e vai identificar as moléculas odoríferas. A localização é variável entre indivíduos, o que justifica a diferente sensibilidade olfativa entre indivíduos. Quando este epitélio olfativo é lesado, é convertido em epitélio respiratório, e o indivíduo perde a capacidade olfativa.
teto é formado por lamina, placa crivosa, em cujas preforações deixam passar neuronios até ao SNC.
microsmático, epitelio olfativo menor
moleculas entram por narinas ou por nasofaringe
Epitélio Olfativo
O organismo pode ser classificado em organismo macrosmático e miscrosmático.
A maior parte dos mamíferos são macrosmáticos(contêm uma área de epitélio bastante grande); os humanos são microsmáticos, porque contêm uma área de epitélio bastante
pequena. A sensibilidade olfativa é diretamente proporcional à área do epitélio olfativo.
As principais células deste epitélio são as células olfativas recetoras neuronais(células bipolares).
Existem ainda as células de suporte. No topo do epitélio olfativo temos as células basais.
É através dos cílios que estão virados para a mucosa que as moléculas odoríferas vão ser reconhecidas.
Para além destas células, existem glândulas de Bowman que produzem muco viscoso rico em proteínas, que garante que as moléculas odoríferas ficam retidas.
celulas glandulares - produzem muco que agarra moleculas odoriferas.
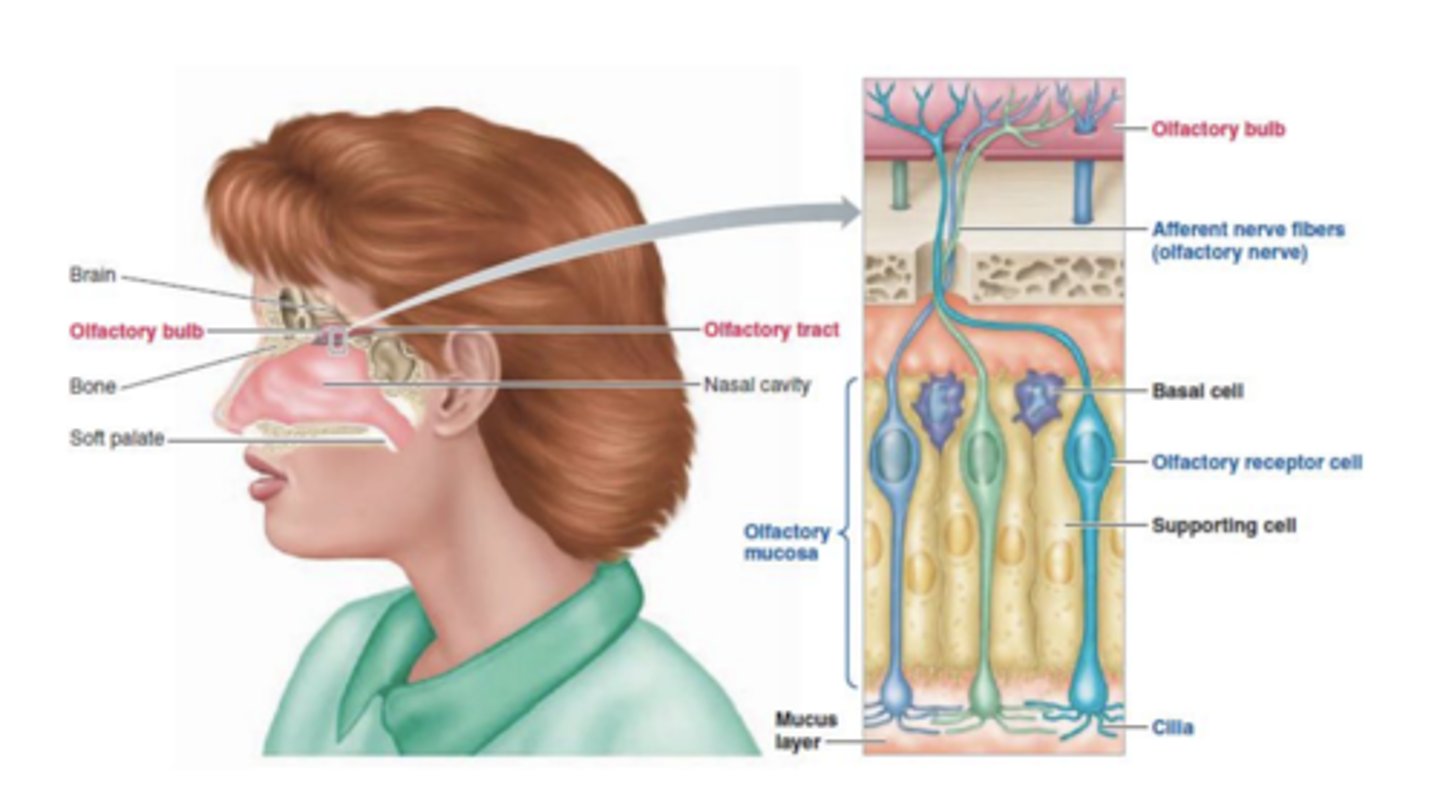
Célula Recetora
Cada célula bipolar emite uma terminação inferior, que termina em cílios, e um polo superior, que vai para o bulbo olfativo. Esses axónios não são mielinizados.
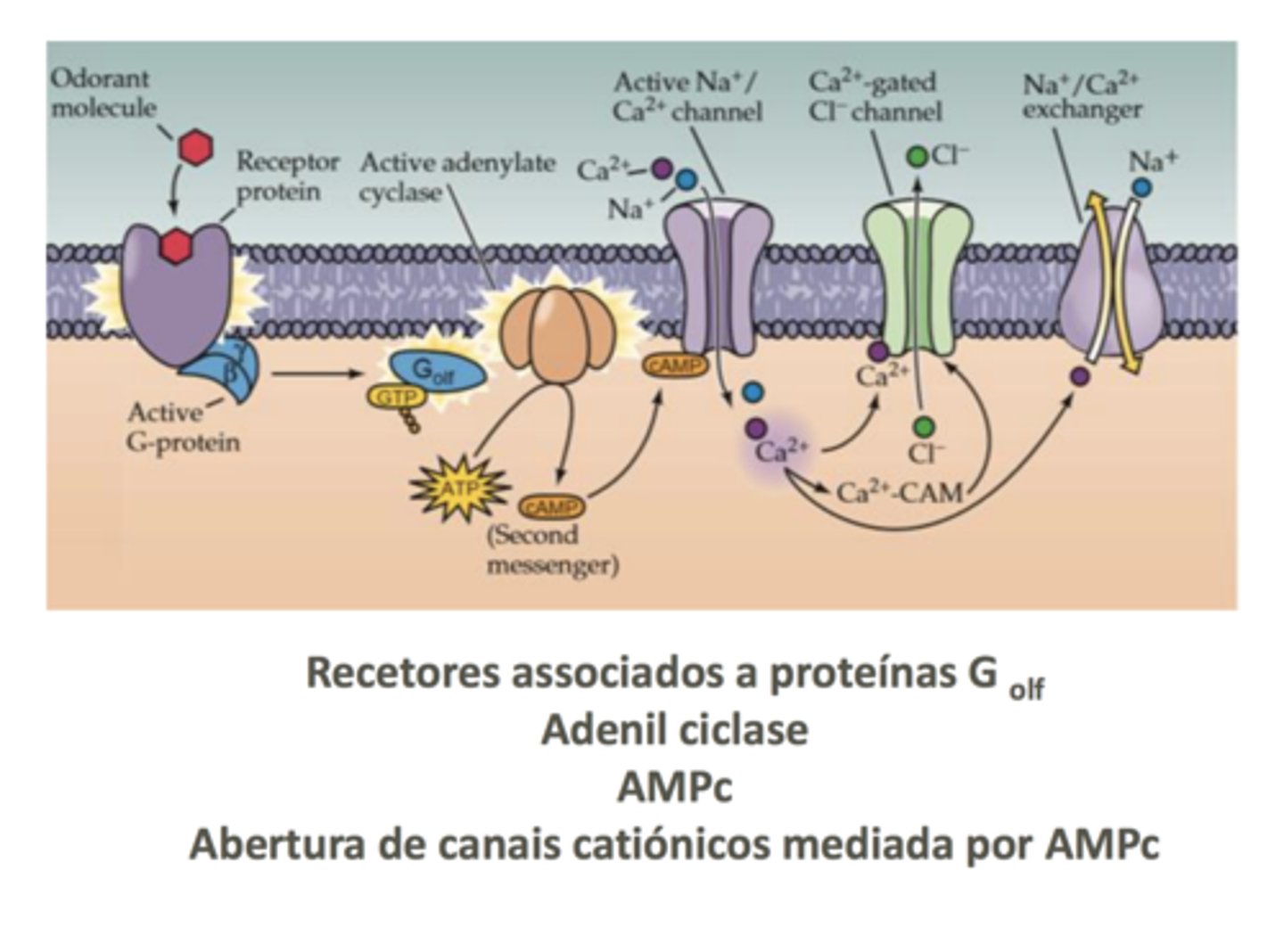
Transdução de Sinal
Começa num recetor membranar metabotrópico; a molécula odorífera liga-se ao recetor, ativa-o, e ele por uma alteração de conformação interage com a proteína G. A subunidade alfa é ativada (subunidade olfativa propriamente dita), e interage com a proteína alfa (adenilciclase).
O cálcio e sódio vão entrar e abrir canais de cloreto (nestas células olfativas e a título de exceção, o cloreto vai estar mais concentrado no meio intracelular); o cálcio interage com o canal de cloreto, abrindo-o e fazendo sair cloreto. Este fenómeno despolariza a célula, levando-a ao limiar de excitabilidade e a um potencial de ação.
Bulbo Olfativo
É uma estrutura par, é constituída por várias camadas de neurónios, sendo que a camada mais inferior está em relação sináptica com as células bipolares do epitélio olfativo.
Estes bulbos têm uma dimensão muito grande porque contêm muitos conjuntos de células. As várias células que existem no bulbo são aglomeradas na forma de glomérulos. Os glomérulos contêm células periglomerulares, células tufosas, células mitrais e células granulares. Cada célula bipolar apenas vai emitir os seus axónios para um único tipo de glomérulo.
• As células tufosas e mitrais são excitatórias e vão conduzir o potencial de ação que geram até às estruturas olfativas do SNC.
• As células granulares são mais pequenas e são inibitórias, inibindo as células mitrais de enviarem informação para o SNC.
• As células periglomerulares fazem a comunicação entre células mitrais de glomérulos diferentes, fazendo com que apenas um determinado glomérulo esteja ativo num determinado momento.
têm elevada convergência
Os feixes nervosos não mielinizados - Fila olfactoria - passam por foramens da placa cribriforme do etmóide penetrando no bulbo olfativo
2 adaptações de insensibilidade
recetores saturados e n enviam tantos potenciais de ação ou
celulas glandulares ativam-se e inibem tufosas e mitrais
Adaptação a Odores
O indivíduo é capaz de se adaptar à presença de odores. Existem dois tipos de adaptação: uma mais rápida e outra mais lenta.
A primeira adaptação leva segundos a ocorrer, ocorre no epitélio olfativo e leva a uma diminuição da frequência dos potenciais de ação ao nível das células bipolares, uma vez
que os recetores das células ficaram saturados; isto ocorre ao fim de segundos.
A segunda adaptação é a mais efetiva. Ocorre no bulbo olfativo, através da ação inibitória criada pelas células granulares, em virtude da ativação das células granulares
por células mitrais.
As células granulares, ao ficarem ativas, vão libertar transmissores inibitórios sobre as próprias mitrais, para diminuir a frequência de potenciais de ação que elas propagam.
Assim, as células mitrais não permanecem ativas durante tanto
tempo, e, portanto, ao fim de alguns minutos, esta inibição cria uma diminuição da sensação que promove finalmente uma adaptação do indivíduo a esse ambiente.
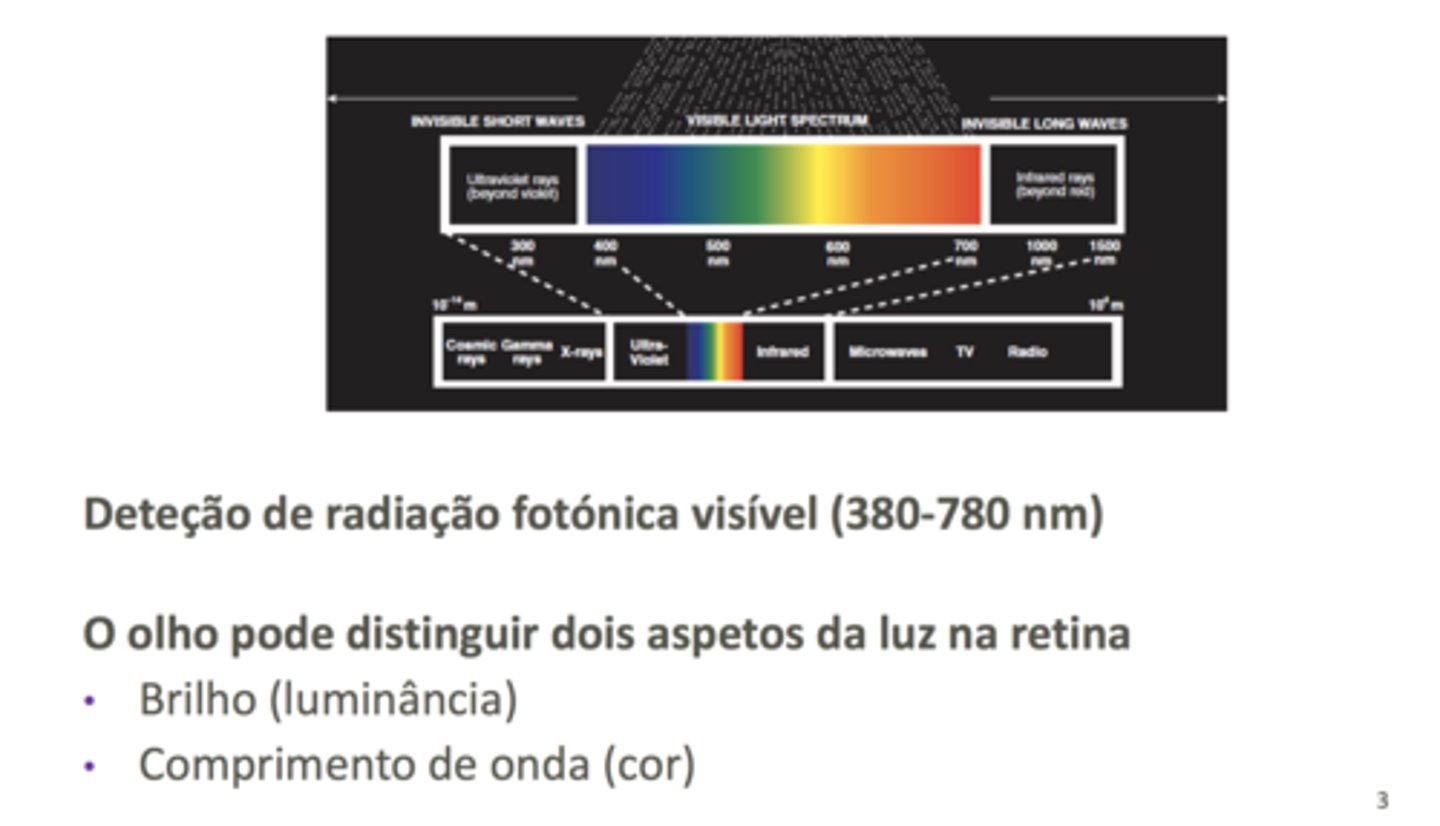
Visão
Consiste na deteção de radiação eletromagnética, na região do visível. O olho humano é capaz de distinguir dois aspetos da radiação eletromagnética visível: a intensidade da radiação e a cor.
A radiação eletromagnética é detetada nas células da retina especializadas para o efeito - fotorrecetores. Há indivíduos com mutações raras que detetam radiação UV, mas isso
é muito raro.
bstonetes detetam brilho
cones detetam varções de cor
Estrutura do Olho
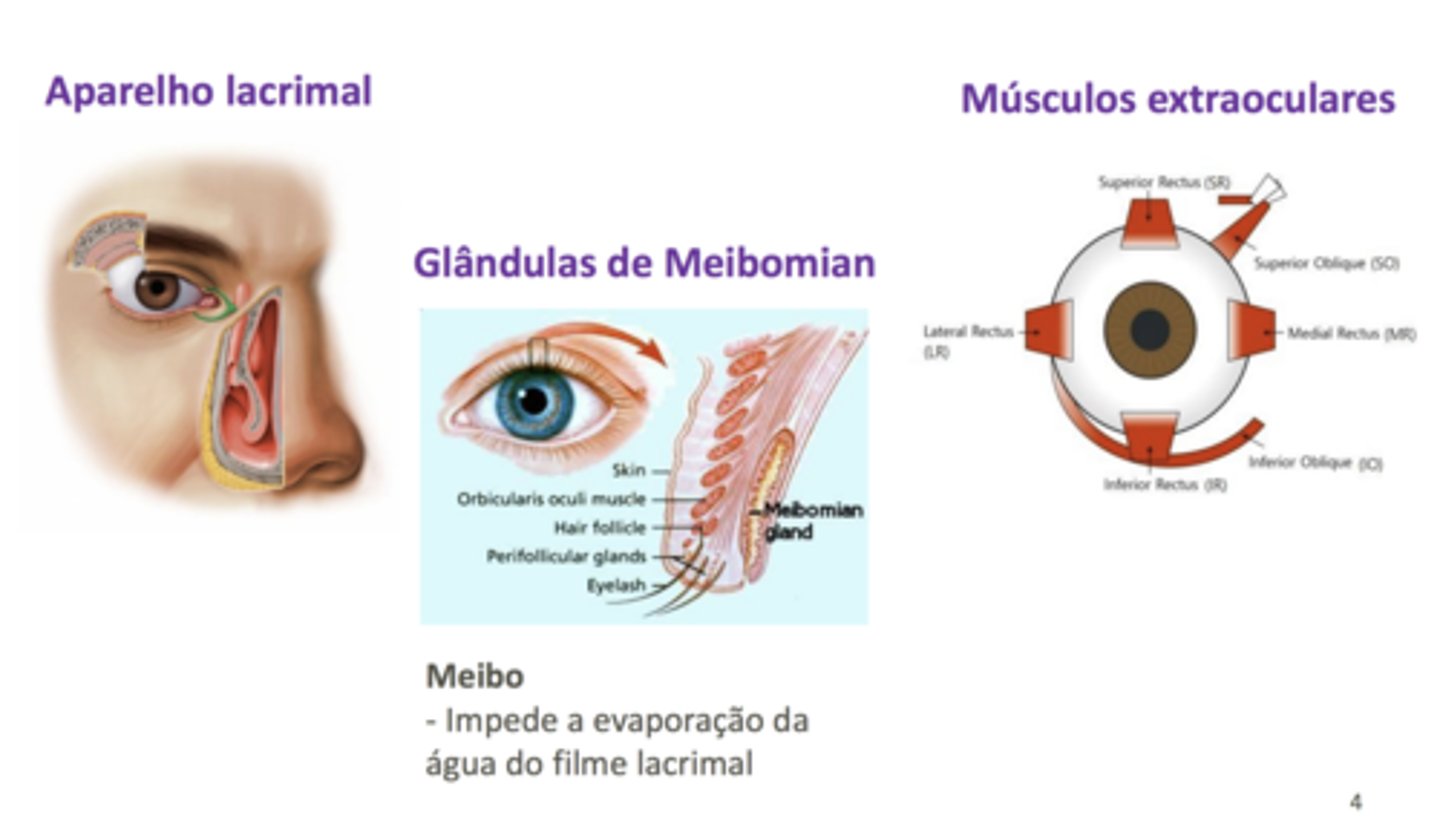
As glândulas lacrimais dão estrutura ao olho; produzem a lágrima, que serve para hidratar a superfície interior do olho e prevenir a dissecação do olho e amortecer arito entre pálpebra e olho, impedindo que ele seja facilmente agredido por poeiras ou outras partículas que existem no ar, e que sofra de irritação pela própria secura. Esta lágrima está constantemente a ser produzida e vai ser drenada ao longo do ducto lacrimal para a cavidade nasal depois escorrendo posteriormente.
Outra estrutura acessória importante é a glândula de Meibomian, que se encontra na face interna das pálpebras. Esta glândula produz uma substância serosa, o chamado
meibo (rico em lípidos), adicionado à lágrima. O meibo tem como função dificultar a evaporação de água da lágrima; se for colocada uma substância lipídica no meio, diminui-se a probabilidade desta água evaporar e o olho fica seco mais dificilmente.
Os músculos extraoculares comanda posição do olhos na órbita
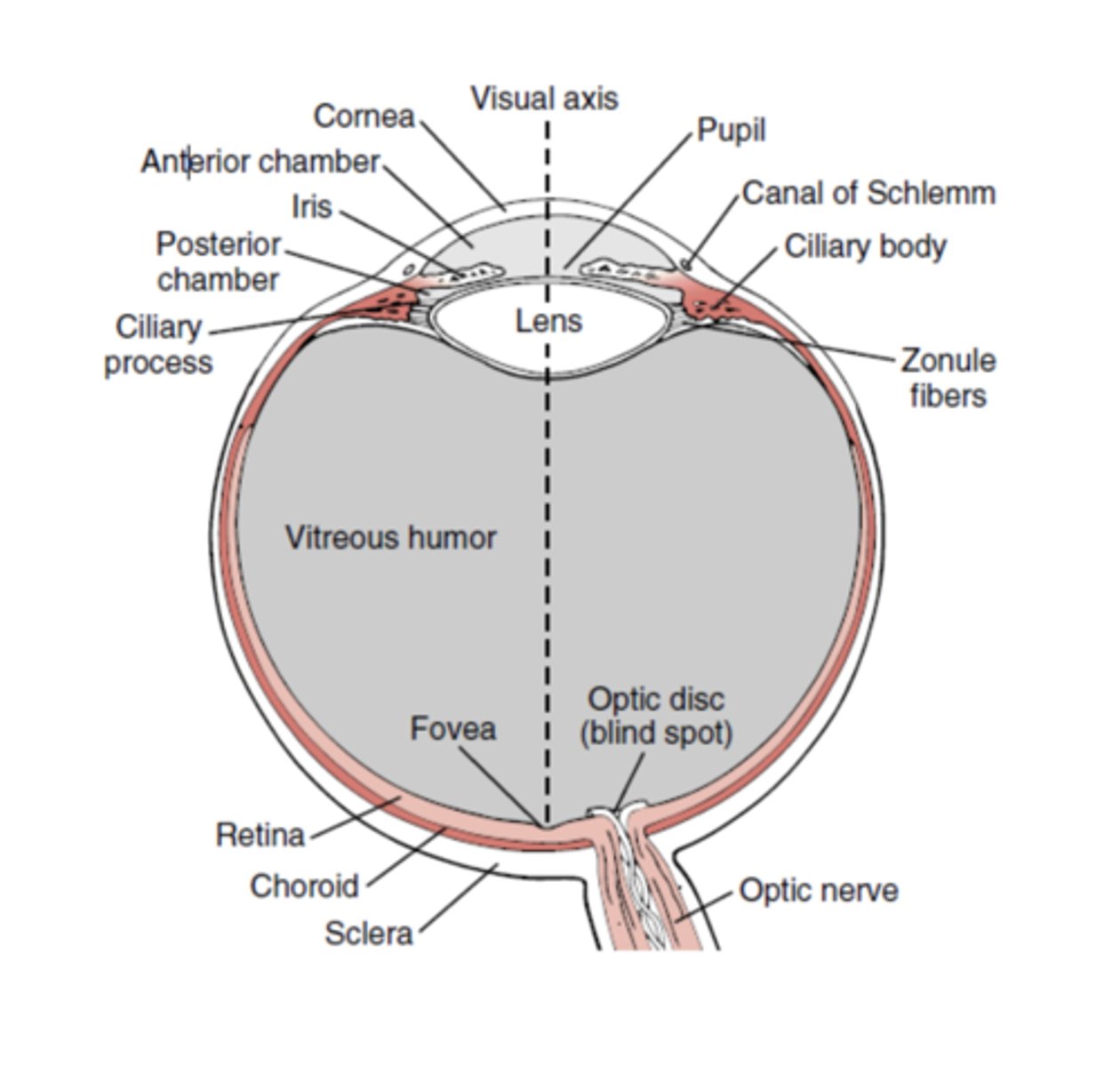
Estrutura do olho:
• Camada Externa, fibrosa: córnea - meio transparente e avascular, semiesférica - e esclerótica - tecido conjuntivo, branco e opaco;
• Camada Média, vascular: íris, 2 muscukos musculo radial e o contratil (simpático) corpo ciliar e coróide;
• Camada Interna, neural: retina - tec nervoso;
• Camada Anterior: humor aquoso, constituído pela câmara anterior e pela câmara posterior; cristalino é biconvexo para a luz ser canalizada np ponto onde há maior densidade de neurónios ancorado pelas fibras pelo corpo ciliar que tem musculo (qd contraido arredonda cristalino); corpo ciliar - epitelio produz liquido; Íris está perfurada no centro pela pupila
• Camada Posterior: humor vítreo.
estrutura do olho
A córnea é a porção transparente do olho (aquela sobre a qual assenta a lente de contacto). Posteriormente, temos uma cavidade anterior subdividida em duas câmaras (câmara anterior e câmara posterior); estas duas são distinguidas pela íris, estrutura muscular que contém músculo liso e é pigmentada.
A íris muda de dimensão, tal como a pupila (orifício escavado no centro da íris).
Posteriormente à íris temos a câmara posterior, que está em contacto direto com o cristalino. O cristalino é a nossa lente; é uma estrutura biconvexa que recebe luz e a encaminha para um único ponto da retina. Como o processo de focagem não é perfeito, a luz não se consegue focar num único ponto e acaba por se projetar numa área focal relativamente grande.
O corpo ciliar está a ancorar o cristalino através de fibras de tecido conjuntivo chamadas fibras zonolares. Posteriormente temos a retina, camada neural do olho que recobre internamente o olho.
O ponto de maior acuidade visual chama-se fóvea, e é o ponto para o qual a maior parte da radiação converge, sendo a região que tem a maior concentração de fotorrecetores
por unidade de área. Onde existe nervo ótico na retina não existem fotorrecetores.
A região de emergência do nervo ótica chama-se disco ótico, e como não contém fotorrecetores é o ponto cego do olho.
Entre o cristalino e a retina temos a maior parte do volume do olho, preenchida pelo humor vítreo, que preenche esta cavidade posterior.
Humor Aquoso
Produzido por uma mucosa que reveste o corpo ciliar. Este corpo ciliar está revestido por uma mucosa que produz este humor aquoso, e o humor circula entre as câmaras
posterior e anterior.
A pressão que este humor exerce à frente do nosso cristalino chama-se pressão intraocular.
Um excesso de pressão de intraocular pode levar a um glaucoma; este glaucoma pode deformar o corpo ciliar e ameaçar a inserção do cristalino, podendo também comprometer o processo de focagem.
Existem dois tipos de glaucoma: glaucoma de ângulo aberto e glaucoma de ângulo fechado. O tipo de glaucoma depende da existência, ou não, de uma drenagem para o humor aquoso.
O humor aquoso é drenado pelos canais de Schlemm, que levam o humor para veias. A existência de um excesso de humor aquoso poderá ser devida a um excesso de produção, a um efeito de drenagem ou a ambos. O glaucoma será tão mais perigoso quanto menor for a possibilidade de drenagem.
Quando o ângulo que a íris faz com a córnea for grande, tratar-se-á de um glaucoma de ângulo aberto; quando o ângulo for mais apertado, tratar-se-á de um glaucoma de
ângulo fechado. A diferença entre os dois é que o glaucoma de ângulo fechado é uma emergência médica, enquanto que podemos nem saber que temos glaucoma, no caso
do glaucoma de ângulo aberto.
Humor Vítreo
Corpo viscoelástico transparente e gelatinoso; tem grande viscosidade porque contém muitas proteínas. Dá estrutura ao olho, dando suporte e protegendo contra alterações bruscas de morfologia do cristalino, funciona como uma reserva metabólica, facilita a acomodação, suporta e protege a lente, auxilia no processo de focagem e participa no
estabelecimento da barreira hemato-ocular.
Focagem
Consiste no encaminhamento da luz que incide na córnea para um único ponto da retina. O papel quer da córnea quer do cristalino é convergir esses múltiplos feixes num único ponto. O ponto de convergência da radiação chama-se ponto focal, e no caso do olho humano é a fóvea.
Em função da morfologia do cristalino, ele será capaz de fazer uma maior ou menor convergência, ou seja, uma maior ou menor refração. Quanto mais alongado o
cristalino, maior a distância focal; quanto mais arredondada o cristalino, menor a distância focal.
A distância focal é expressa em metros, e é definida como a distância do cristalino ao ponto focal na retina. O inverso da distância focal designa-se dioptrias. Quanto maior a distância focal, menor o número de dioptrias, e quanto menor a distância focal, maior o número de dioptrias. Quanto maior a curvatura de uma lente, maior o seu poder de refração (no dioptrias).
O processo de focagem de refração convergente promove a inversão da imagem formada na retina face ao objeto.
Refração
Mudança da direção de um feixe de radiação quando este muda de meio de propagação num ângulo oblíquo.
Lentes côncavas provocam a divergência de radiação.
Temos duas estruturas que fazem refração convergente no nosso olho: primeiro a córnea, e depois o cristalino.
A córnea tem o maior poder refratário; no entanto, não pode mudar a sua morfologia, não podendo ser responsável pela regulação da refração.
O cristalino tem aproximadamente metade do poder refratário da própria córnea, pode mudar a sua morfologia, e, portanto, determinar uma maior ou menor convergência da radiação.
A luz vai chegar ao nosso olho propagando-se por meio aéreo; quando a luz passa do meio aéreo exterior para o meio aquoso interno, sofre uma primeira refração na região fronteira - na córnea. A radiação vai depois passar do humor aquoso para dentro do cristalino; como estas duas estruturas têm uma constituição diferente, teremos uma segunda refração, na margem anterior do cristalino.
Quando a luz passa do cristalino para o humor aquoso, temos uma terceira refração.
Depois de todas estas refrações, a luz converge para o ponto de maior acuidade, a fóvea da retina.

Acomodação
A alteração da curvatura do cristalino é levada a cabo por uma alteração do tónus do músculo ciliar (controlado pelo SN parassimpático), sendo caracterizada como a capacidade de alterar a focagem de objetos a diferentes distâncias através da alteração da curvatura da lente.
Focar luz com diferentes origens (par III) - refração - pela córnea (maior parte da refração) devido à sua forma.
E no cristalino, menos potente, este consgue alterar a sua estrutura para corrigir a refração a luz. Quando o parassimpático estimula o músculo ciliar, ele contrai e promove o arredondamento do cristalino; já quando o parassimpático não o estimula tanto, ele relaxa e o cristalino fica mais alongado/estreitado. À capacidade de modificação da estrutura do cristalino damos o nome de acomodação.
Feixes de luz de objetos que estejam próximos do olho necessitam de uma maior convergência do que objetos que estejam distantes.
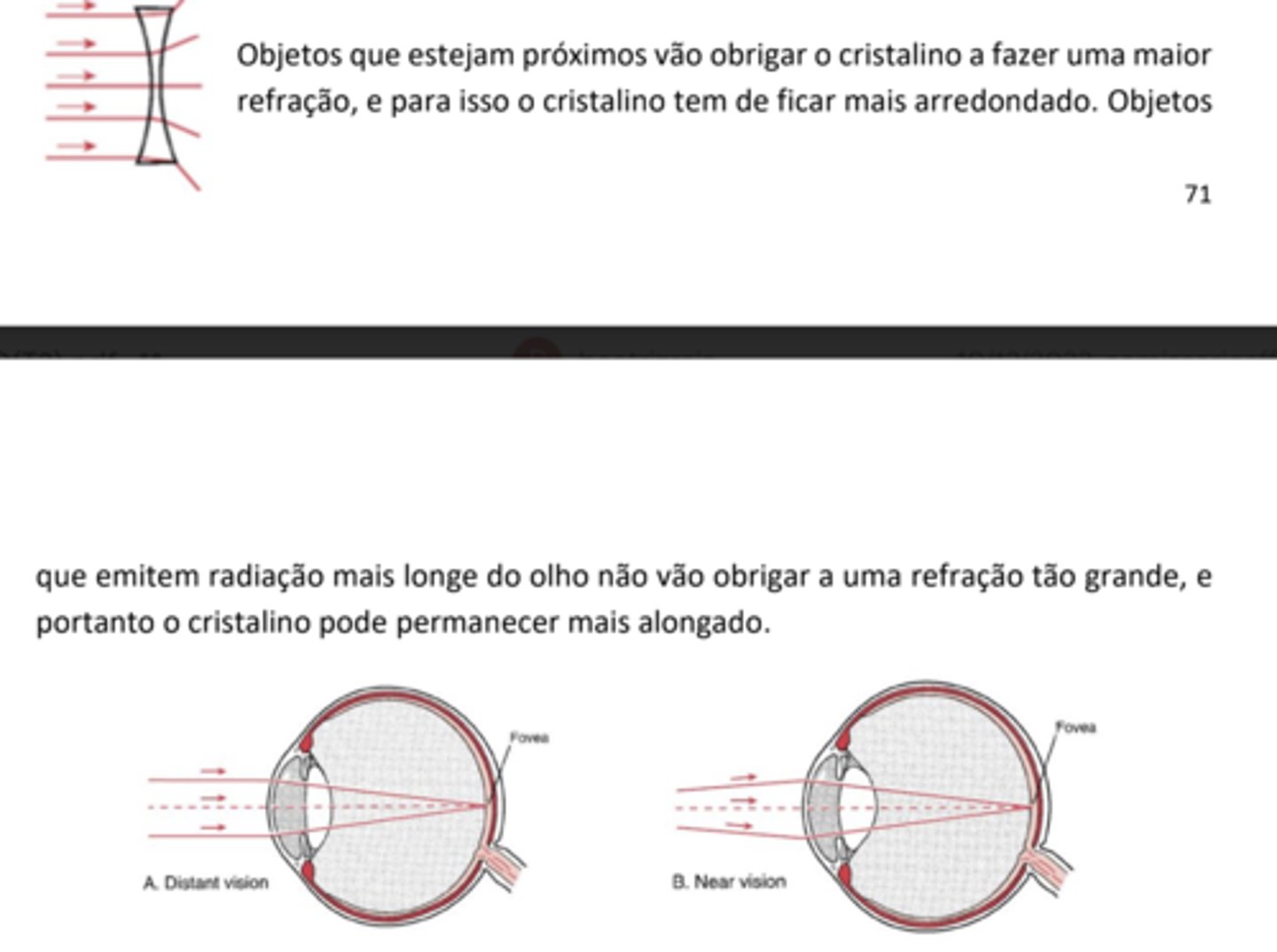
Retina
A luz vai inicialmente propagar-se da camada mais interna para a retina, depois até à camada mais externa da retina, e depois para trás; apenas quando a luz volta para trás é que ocorre a via de transdução sensorial no olho.
A luz vai atingindo camadas cada vez mais externas, e parte desta radiação vai ser absorvida pela camada mais externa: camada pigmentar rica em melanina. A parte que não é absorvida é refletida, voltando para trás, chocando com os bastonetes e com os cones (células fotorrecetoras).
Os bastonetes são responsáveis pela deteção de radiação e informam o SNC da quantidade de luz (brilho). Os cones detetam o comprimento de onda (cor). Os bastonetes e os cones estão em relação sináptica com outras células.
As células bipolares são inibitórias, e as fotorrecetoras são excitatórias.
As células bipolares estão em comunicação com as células ganglionares - células semelhantes a neurónios, cujos corpos estão na retina e cujos axónios se vão agrupar e formar os
nervos óticos.
Se luz bate na fóvea há foco
Reagião onde sai nervo ótico nao tem neurónios nem fotorrecetores
nervo ótico é suprimido por vasos e coberto por meninges
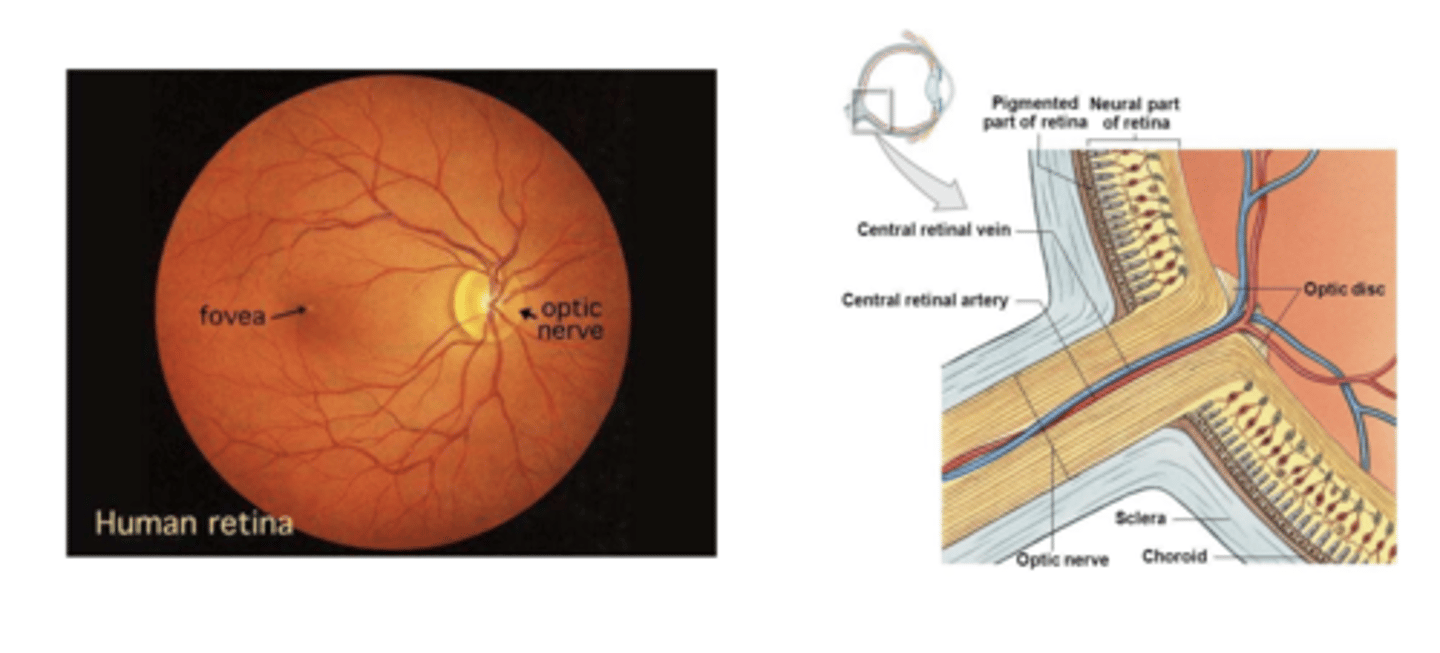
Retina 2
parte da luz, melanina, é absorvida e refletida de volta para o humor vitrio excitandp os neuronios cones e bastonete
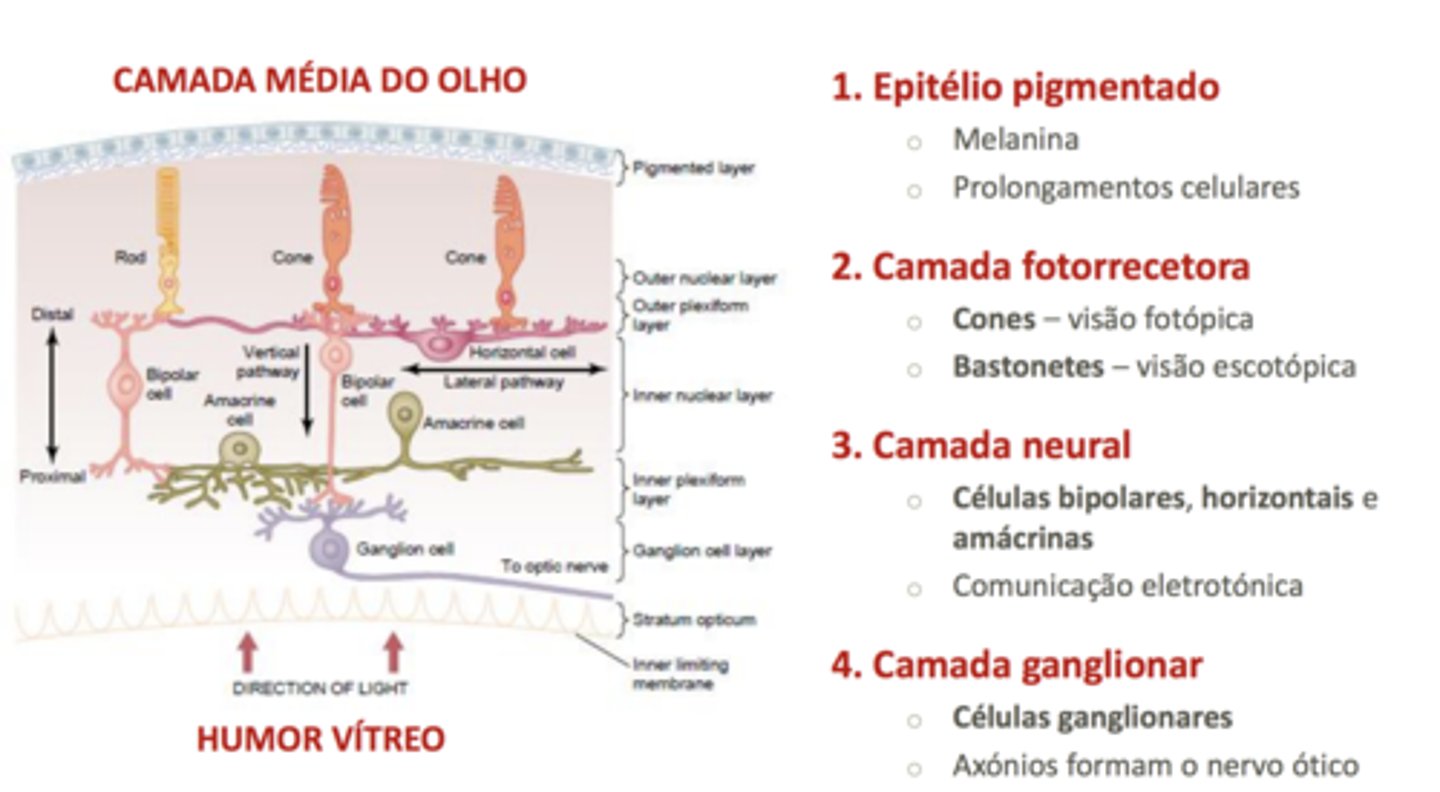
Cones
Têm 3 tipos de pigmentos (opsinas): verdes, azuis e vermelhos. Todas as cores que vemos são o resultado da mistura destas 3 cores visíveis.
Teoria tricromática: para termos um campo visual de cores, temos de ter, no mínimo, dois tipos de pigmentos.
Fotorrecetores 2
Os fotorrecetores têm, no seu segmento mais externo, um citoplasma organizado por
uma membrana plasmática dividida em vários discos altamente concentrados em
proteínas ligadas a grupos pigmentados.
Em função da receção de radiação, ocorre uma alteração de conformação dessas
moléculas orgânicas que constituem os pigmentos desses discos, e em função dessa
alteração, a própria proteína que está em ligação com o pigmento também muda de
conformação, dando início a uma via de transdução sensorial.
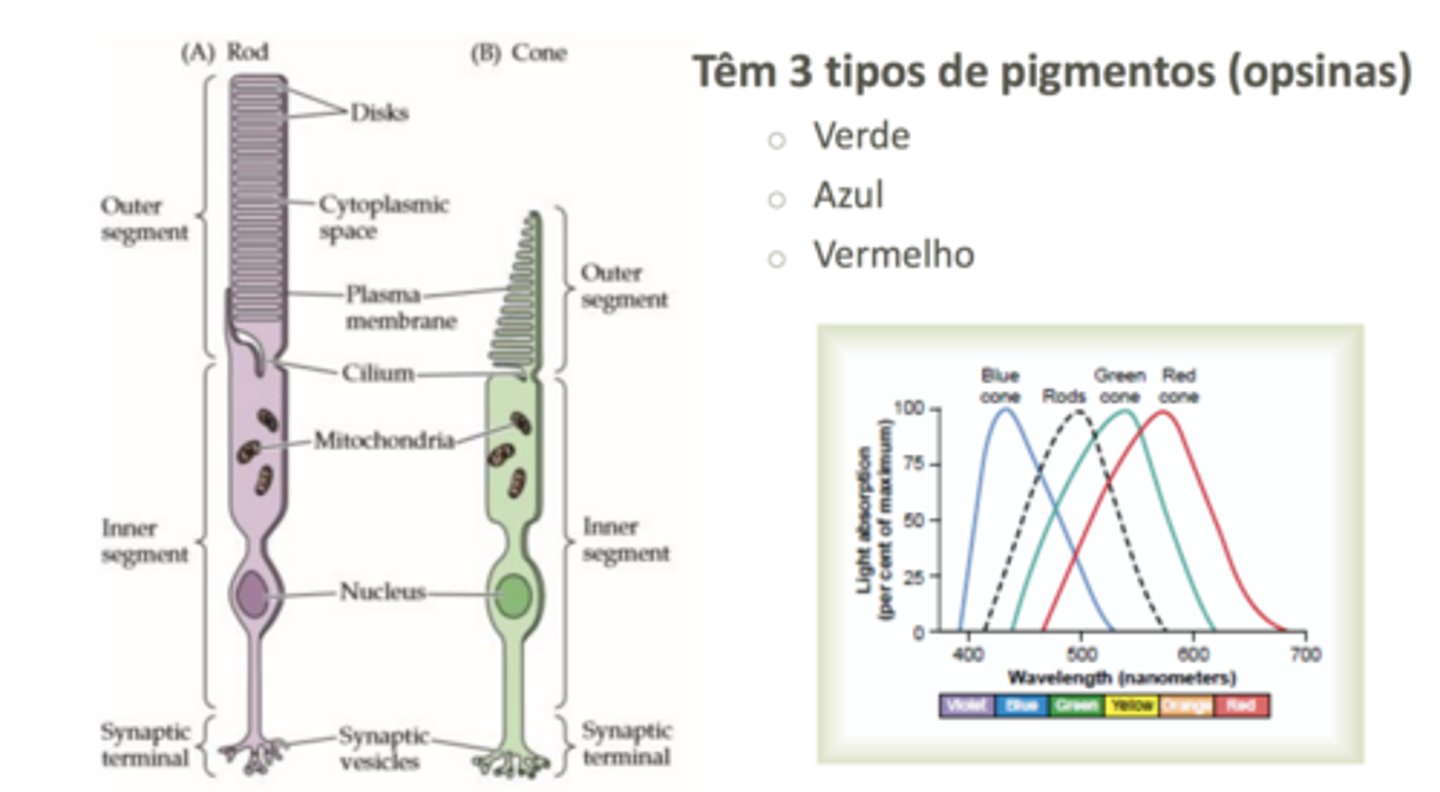
discos fotorrecetores
emos proteínas que estão acopladas a pigmentos. O nome geral do recetor para a proteína é rodopsina; a componente proteica da rodopsina é a escotopsina e o pimento à qual ela está associada é o 11-cis-retinal, que passa de cis para trans mudando comformação do próprio recetor. Este recetor é um recetor acoplado a um proteína G chamada transducina.
Fototransdução
A luz vai incidir sobre o pigmento do recetor, num disco fotorrecetor; em virtude desse choque da radiação no 11-cis-retinal, ele transforma-se em 11-trans-retinal. O recetor
ativo sofre uma alteração de conformação e ativa a proteína G, trocando GDP por GTP e ficando ativa. Ao ficar ativo, desloca as suas subunidades beta e gama e interage com a sua proteína efetora (é uma PDE: fosfodiasterase - enzima que destrói os segundos mensageiros AMPc e GMPc).
No caso de ser uma célula fotorrecetora, o segundo mensageiro que ela está constantemente a produzir em virtude de uma ativação tónica da guanilciclase é o
GMPc, e, portanto, um fotorrecetor que não receba luz terá grandes quantidades de GMPc. Quando chega a luz, a PDE é ativada e destrói o GMPc que existe na célula.
O GMPc que existia estava constantemente a abrir canais de sódio; com o canal aberto, o sódio entrava e despolarizava a célula, formava o limiar de excitabilidade, ocorria o potencial de ação e esta célula fotorrecetora libertava o seu transmissor sob a célula vizinha.
Quando chega luz, a fosfodiasterase destrói o GMPc e ele deixa de poder manter aberto o canal de sódio; o sódio deixa então de entrar, a célula não é capaz de despolarizar, e, pelo contrário, hiperpolariza e não liberta transmissores.
sinapse fotorecetore e bipolares
bipolares e ganglionares
Ausência de Luz
Arco reflexo simpático - midríase
A célula está despolarizada, e o transmissor que ela liberta sobre a célula bipolar é o glutamato. O glutamato é um transmissor excitatório: cria um potencial de ação na célula bipolar, obrigando-a a libertar os seus transmissores sobre as células ganglionares.
Quando não há luz, os fotorrecetores estão despolarizados - estão a ativar células bipolares para que estas possam inibir células ganglionares. Como as células ganglionares estão inibidas, e como são elas que levam informação ao SNC, não há informação a ser levada ao SNC → a ideia que o SNC cria é de um campo visual escuro.
escuridão excita fotorrecetores, geram potencias de ação e libertam glutamato na célula seguinte bipolar bipolar é inibitório ppoois secreta gaba sobre o terceiro neurónio que é ganglionar
Presença de Luz
Arco reflexo parassimpático - miosee
Quando há luz, os fotorrecetores estão hiperpolarizados, e, portanto, não libertam tanto glutamato sobre as células bipolares.
Como não libertam tanto glutamato, as células bipolares não estão tão ativas, e portanto não podem libertar tantos transmissores inibitórios sobre as células ganglionares - as células ganglionares estão mais ativas.
As células ganglionares ativas enviam informação na forma de potencias de ação para o SNC, e em virtude do local onde essa célula ganglionar está ativa, aí se cria a existência de um campo visual com brilho e cor.
Vias Aferentes
Metade dos axónios destas células ganglionares sofrem decussação para o hemisfério cerebral contralateral (decussam ao nível do quiasma ótico), enquanto que a outra
metade segue para o hemisfério ipsilateral. No quiasma ótico, 50% das fibras de um nervo ótico são projetadas para o hemisfério cerebral contrário.
Metade das fibras do olho esquerdo seguem um trajeto homolateral, e a outra metade segue um trajeto contralateral, acontecendo a mesma coisa para o olho direito.
Sempre que ocorrer uma lesão do nervo ótico, em qualquer porção do seu trajeto, o campo visual ficará alterado de forma aferente.
Audição
Capacidade do individuo detetar e interpretar ondas sonoras., que são zonas de concentração e rarefação de moléculas. O ser humano tem uma gama de frequências audíveis limitada, mas tem uma maior sensibilidade entre os 1000 e 4000 Hz.
O ouvido divide-se numa porção externa, média e interna.
ondas harmonicas sobrepostas na mesma frequencia é o timbre
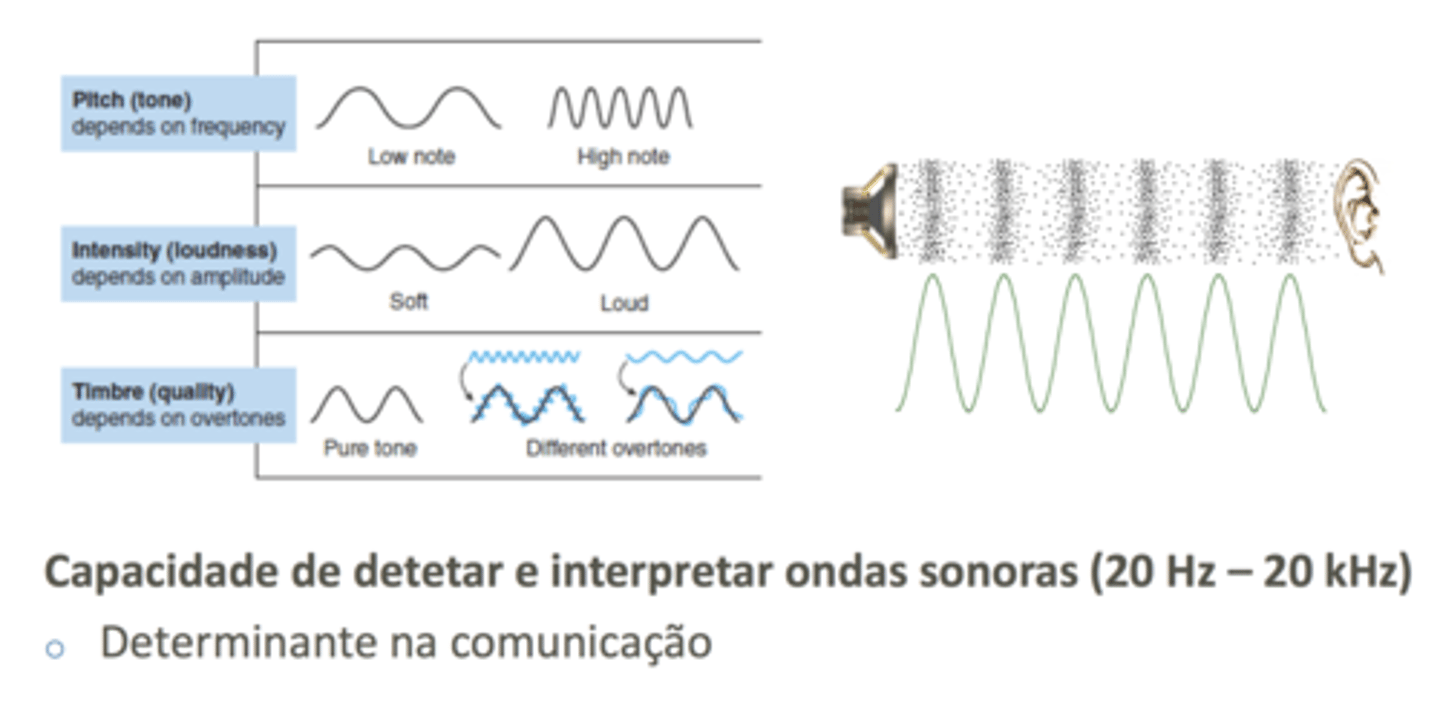
Ondas Sonoras
São zonas de alternância de compressão e rarefação das moléculas do ar; em qualquer zona onde estejam mais concentradas, produz-se um som mais intenso.
Sempre que houver menos moléculas por unidade de volume, temos uma rarefação, e aí a intensidade do som diminui.
Em termos de ondas sonoras, conseguimos detetar a nota musical associada ao som (som grave ou agudo), dependente da frequência; a intensidade, dada pela amplitude das ondas sonoras; e conseguimos distinguir diferentes timbres, qualidades entre ondas sonoras.
Sobretons são ondas sonoras que se sobrepõem a uma onda sonora original. Um tom puro gera ondas sonoras de forma sinusoidal.
Condução Sonora
O ouvido está dividido em 3 porções.
O ouvido externo está dividido em pavilhão auricular e canal auditivo externo.
O som chega ao canal auditivo externo, faz vibrar as estruturas que se encontram no ouvido médio, e o ouvido médio faz então a transmissão desse som para o ouvido interno.
O ouvido interno está dividido em duas porções, a cóclea e o vestíbulo (labirinto).
A forma como o som atinge o ouvido interno pode ser diferente. Há duas formas de condução sonora: a condução aérea e a condução óssea.
condução óssea
consiste na vibração dos ossos, inclusive dos dentes, aquando do processo de produção de som pelo próprio organismo - quando o individuo fala ou mastiga - sendo uma condução rápida. Este tipo de condução explica o porquê de ser mais difícil ouvir enquanto mastigamos alimentos duros e crocantes.
condução sonora
ocorre quando o som sai do corpo, propaga-se pelo ar e entra pelo canal auditivo externo, e só depois atinge o ouvido interno.
Quando ouvimos a nossa voz falada e a nossa voz gravada, entendemos que são duas vozes completamente diferentes; quando falamos e nos ouvimos a falar, estamos a receber som por duas vias - via óssea e via aérea. Quando ouvimos a nossa voz gravada, detetamos esse som apenas por condução aérea.
Ouvido Externo
Constituído pelo pavilhão auricular e pelo canal auditivo externo. A arquitetura do pavilhão auricular serve para amplificar e concentrar as ondas sonoras no canal auditivo
externo.
A separar o ouvido externo do ouvido médio temos a membrana timpânica, ricamente enervada, e para lá da membrana timpânica temos o ouvido médio.
Ouvido Médio
Cavidade no osso temporal cheia de ar; este ar tem uma pressão que se equilibra geralmente com a pressão atmosférica (pressão medida na cavidade oral). Comunica
com a nasofaringe pela trompa de Eustáquio, que está fechada. Vários movimentos relacionados com a musculatura mastigatória podem fazer abrir esta trompa e equilibrar
as duas pressões: espirro, bocejo, deglutição e manobra de Valsalva.
Temos os 3 ossos mais pequenos do corpo humano, chamados ossículos do ouvido: estribo, martelo e bigorna, que vão funcionar como um pistão e transmitir a vibração do tímpano até à janela oval da cóclea/caracol, amplificando também o som.
Ouvido Interno
Constituído pela cóclea, é a cavidade espiral do osso
temporal cheia de fluído.
Esse fluido está dividido em 3 compartimentos longitudinais, e a cada um damos o nome de escala: escala vestibular, escala timpânica e escala média/ducto coclear.
Os nossos recetores para o som vão estar albergados na
escala média.
O tímpano, ao vibrar, faz vibrar a cadeia ossicular, e esta faz vibrar, pelo estribo, a janela oval. O som vai viajar a partir do canal superior até à extremidade do caracol, e depois vai voltar para trás pelo canal inferior - escala timpânica. Entre estes dois canais, temos a escala média, onde temos as nossas células recetoras.
É quando o som volta para trás que faz vibrar o som do túnel que está no meio, e quando o chão desta escala média vibra, as células que lá estão montadas chocam com o teto e ficam ativadas.
As escalas vestibulares e timpânicas são ricas em sódio. Já a escala média é um fluido atípico porque é rico em potássio. Só há doistipos de células no organismo humano que despolarizam pela entrada de potássio, sendo que as células do ouvido interno são um desse tipo.
Órgão de Corti
A membrana tectorial é uma estrutura rígida que funciona como um teto para as nossas células sensoriais, células compridas que terminam a sua extremidade apical em cílios
que não se movem.
Essas células estão apoiadas numa base, e essa base está apoiada num chão - membrana basilar; este chão vibra, enquanto que o teto não.
A vibração da membrana basilar provoca o deslocamento do órgão de Corti em relação à placa tectorial, que é estática.
Sempre que as células sensoriais chocam com a membrana tectorial devido à vibração, os cílios sofrem uma alteração de posição. À superfície dos cílios, temos canais de potássio que respondem a essa deformação e que deixam o potássio entrar e assim despolarizar a célula.
Cóclea
As diferentes regiões da cóclea respondem a diferentes frequências. Regiões mais próximas do ouvido médio são mais sensíveis a sons de elevada frequência, e regiões
mais distantes do ouvido médio são mais sensíveis a sons mais graves.
A morfologia peculiar da cóclea serve para assegurar a decomposição e fracionamento em termos de frequência.
Células-Cabelo
Quando estas células chocam com a placa tectorial, os cílios sofrem deslocamento, do eixo mais curto para o mais longo; no topo dos cílios existem canais de potássio, abertos com este deslocamento, e o potássio entra.
As células cabelo sofrem defleção positiva pela vibração sonora.
Na escada média, a concentração de potássio é muito maior que a de sódio.
Quando ocorre lesão auditiva interna, muito provavelmente por exposição a som de elevada intensidade, estes cílios dobram-se no sentido contrário, levando a perda auditiva de origem cerebral.
Labirinto
O equilíbrio é assegurado em grande parte pelo labirinto, constituído por três canais semicirculares - horizontal, posterior e anterior - que convergem em três regiões de
dilatação - máculas, somando-se ainda dois órgãos otolíticos - utrículo (sensível à aceleração horizontal) e sáculo (mais sensível à aceleração vertical).
Dentro destas estruturas circula fluido; dentro das máculas e dos órgãos otolíticos vamos encontrar as nossas células ciliadas/células-cabelo, que vão responder à forma como o fluido se desloca sobre as células.
Canais Semicirculares
São canais sensíveis a rotações na cabeça. Ao nível das máculas, estas células sensoriais estão montadas em elevações chamadas cristas; no topo destas cristas existe um
conjunto de filamentos proteicos gelatinosos a que damos o nome de cúpula. A ampola é a dilatação do canal semicircular.
Esses filamentos têm uma posição relativamente fixa, mas como são gelatinosas não são rígidas e podem deslocar-se e chocar com as nossas células sensoriais à mesma velocidade e ritmo que a própria cabeça.
Utrículo
Os dois órgãos otolíticos não têm cúpula, mas sim cristais de carbonato de cálcio que recobrem as células sensoriais. Quando estamos de pé, as células-cabelo do utrículo encontram-se na vertical, a apontar para cima.
O utrículo e o sáculo são sensíveis às acelerações horizontais e verticais da cabeça, respetivamente.
Membrana Otolítica
Os cristais são imóveis. Quando a cabeça se movimenta, são os próprios cílios que se movimentam; no entanto, como os cristais são fixos e não se movimentam, cria-se uma inércia entre os próprios cílios e os cristais, provocando-se o deslocamento dos cílios, a entrada de potássio e o potencial de ação.
Sistema Somatosensorial
Classificação de Fibras Nervosas
As fibras (axónios) têm diferentes velocidades de condução, diferentes diâmetros, sendo algumas são mielinizadas e outras não. Em virtude destas características, foi criada uma classificação de fibras, em A (α, β, γ, δ), B e C. As fibras quando se encaminham de A para C perdem diâmetro e, portanto, perdem velocidade.
As A e B são mielinizadas, enquanto as C não são mielinizadas (muito lentas, finas). As fibras B são encontradas nos neurónios pré-ganglionares autonómicos, e as C são encontradas nos pós-ganglionares autonómicos, em neurónios sensitivos.
Sistema Exterocetivo
O sistema somatosensorial divide-se em exterocetivo, enterocetivo e propriocetivo.
O sistema exterocetivo tem como objetivo proteger o organismo de lesões, promover a interação social, promover a reprodução e promover o desenvolvimento ontogénico.
o Evitar lesões
o Interação social
o Reprodução
o Desenvolvimento ontogénico (crescimento do corpo, só c/ a deteção de peso é que o esqueleto se encontra mineralizado; calibre das celulas musculares é 100% recuperavel mas mineralização do esqueleto não é)
Homúnculo Cerebral
Exibe a sensibilidade, e não o tamanho de uma zona do corpo.
É através da deteção de pressão aplicada sobre os sistemas músculo-esqueléticos, quer sobre os ossos, quer sobre as articulações, quer sobre os músculos, que todas estas estruturas ganham uma determinada rigidez: os ossos ganham mineralização, os músculos ganham volume, e é assim que o sistema músculo-esquelético se mantém com dimensões e funções adequadas. A gravidade zero, os ossos tendem a desmineralizar e os músculos tendem a atrofiar.
O homúnculo cerebral é uma figura que representa, através de progressões exageradas, a sensibilidade relativa de cada região.
Recetores Cutâneos
São mecanorrecetores, termorrecetores e nocicetores.
Estão essencialmente dispostos na pele, quer na epiderme quer na derme, mas menos presentes na hipoderme.
Podemos ter:
• Terminações livres: sensíveis à pressão, variação de temperatura, e dor. São fibras Aδ e C-polimodais - recebem vários tipos de estimulação.
• Corpúsculos de Meissner (Aβ): encontram-se nas zonas glabras da pele (zonas que não têm pelo, como as pontas dos dedos e lábios). Detetam movimento sobre a superfície da pele e localizam bem o estímulo na pele; muito sensíveis e
rápidos, informam sobre a textura do estímulo.
• Corpúsculos de Merkel (Aβ): bastante superficiais, encontram-se em regiões pilosas da pele, sendo de adaptação mais lenta; conferem informação sobre o
contacto constante de um objeto junto à pele. mielinizados e espessos
• Terminações pilosas (Aβ): junto dos folículos pilosos, detetam o movimento de objetos sobre a pele e informam sobre o contacto inicial de objetos com a pele.
• Terminações de Ruffini (Aβ): são as terminações mais profundas da pele (em articulações), de adaptação mais lenta, e dão informação sobre uma deformação contínua.
• Corpúsculos de Paccini (Aβ): estruturas discóides, encontrados em zonas profundas da pele, respondem à compressão local rápida e frequente, ou seja, à vibração da pele.
Teste de discriminação entre dois pontos: teste para avaliar a sensibilidade da pele.
RECETORES FÁSICOS
ADAPTAÇÃO RÁPIDA
ex: corpusculos de meissner
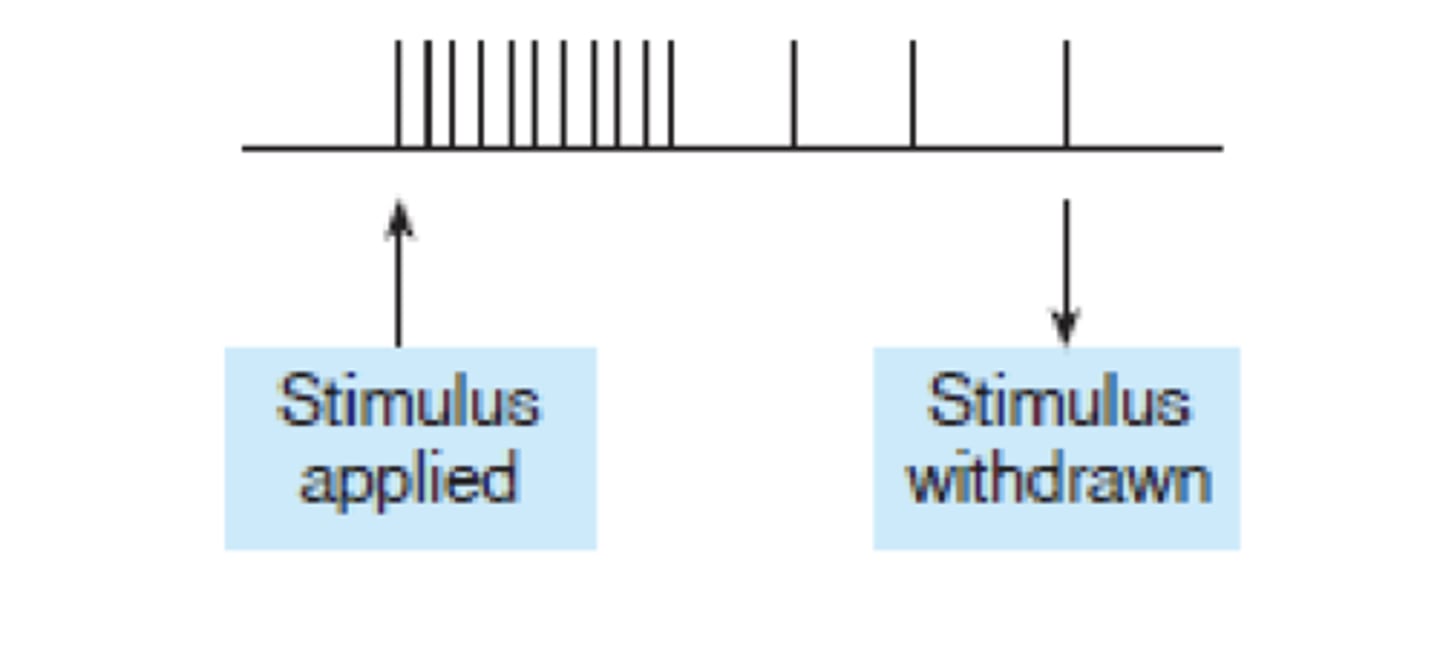
RECETORES TÓNICOS
ADAPTAÇÃO LENTA
ex: terminações de ruffini, merkle, A e beta
Termorrecetores Cutâneos
Não detetam temperaturas absolutas, mas sim variações. Há duas subpopulações:
• Frio: mais numerosos e mielinizados.
• Calor: não são mielinizados e são menos numerosos.
Sempre que as temperaturas não forem nem muito quentes, nem muito frias, ambas as subpopulações despolarizam, criando a noção de uma temperatura mais amena.
quando superam limites sao percetidos como informação dolorosa
Nocicetores
Respondem a estímulos que, em virtude da sua intensidade ou frequência, ameaçam ou lesam o organismo ao criar uma sensação de dor.
• Fibras Aδ: primeiras a responder, criam a chamada primeira dor (localizada, curta e intensa), dor epicrítica, e são preferencialmente estímulos mecânicos.
• Fibras C-polimodais: podem responder a várias modalidades de estímulos, e produzem a segunda dor, mais difusa e prolongada, chamada de protopática.
Por que motivo a sensibilidade olfativa diminui durante um estado de congestão nasal?
muco cobre cilios e as moleculas olfativas nao chegam
Ainda aprendendo (11)
Você começou a estudar os termos a seguir. Continue assim!