02/10, Louise Kauffman (proprioception)
1/20
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced |
|---|
No study sessions yet.
21 Terms
La proprioception correspond à une sensibilité intéroceptive qui nous informe en permanence sur la position et le mouvement de nos membres, ainsi que sur l’état de contraction des muscles et la position des articulations.
Autrement dit, elle permet au système nerveux de connaître à chaque instant la configuration du corps dans l’espace, sans recours à la vision.
Cette sensibilité joue un rôle fondamental dans :
le contrôle de la motricité (planification et ajustement des mouvements),
le maintien de la posture et de l’équilibre lors de la station debout,
et la représentation interne du corps, c’est-à-dire la conscience corporelle.
Bien que cette information ne soit pas toujours perçue de manière consciente, elle est indispensable pour exécuter des mouvements coordonnés et précis. L’altération de la proprioception conduit à une perte du contrôle moteur fin et du sentiment d’incarnation du corps.
Exemple clinique
Le rôle crucial de la proprioception est illustré par certains cas rares de patients ayant perdu cette sensibilité.
Oliver Sacks a notamment décrit le cas célèbre de « la femme désincarnée », qui, à la suite d’une atteinte proprioceptive diffuse, ne percevait plus la position de ses membres.
Elle rapportait la sensation de flotter dans l’air et d’être détachée de son propre corps, ce qui entraînait de graves troubles moteurs, notamment une incapacité à se mouvoir sans repères visuels.
En l’absence de retour sensoriel sur la position des segments corporels, le système moteur ne peut plus corriger ni coordonner efficacement les mouvements.
Récepteurs proprioceptifs
Les récepteurs proprioceptifs sont des mécanorécepteurs profonds, sensibles à des stimulations mécaniques telles que l’étirement, la pression ou la vibration.
Ils sont localisés à l’intérieur du corps, dans les muscles, les tendons et les articulations.
Les fuseaux neuromusculaires
Localisés dans les muscles squelettiques, les fuseaux neuromusculaires sont des structures spécialisées constituées de fibres musculaires intrafusales, autour desquelles s’enroulent les axones des fibres sensorielles de type Ia.
Ces fibres sont mécanosensibles : elles répondent à l’étirement du muscle et renseignent sur sa longueur et la vitesse de son étirement.
Les fibres musculaires intrafusales se distinguent des fibres extra-fusales, qui assurent la contraction musculaire proprement dite.
L’activité des fuseaux neuromusculaires fournit donc une information continue sur l’état d’allongement du muscle, permettant d’ajuster en temps réel la contraction et de maintenir la posture.
Récepteurs proprioceptifs
Les organes tendineux de Golgi
Situés à la jonction entre le muscle et le tendon, ils sont sensibles à la tension exercée lors de la contraction musculaire.
Ils envoient des signaux via des fibres sensorielles Ib, contribuant à la régulation réflexe du tonus musculaire.
Les récepteurs articulaires
Présents dans les capsules et ligaments articulaires, ils renseignent sur la position et les mouvements des articulations.
Leur activité complète les informations issues des fuseaux neuromusculaires et des organes tendineux.
Organisation fonctionnelle
Les informations proprioceptives sont transmises par des fibres sensorielles afférentes vers le système nerveux central, où elles empruntent les voies lemniscales (ou voies de la sensibilité profonde consciente).
Elles se projettent :
vers le cervelet, qui utilise ces signaux pour ajuster la motricité fine et l’équilibre,
et vers le cortex somesthésique primaire (S1), où elles participent à la représentation consciente du schéma corporel.
En résumé
La proprioception est une forme de sensibilité intéroceptive mécanique, assurée par des récepteurs musculaires, tendineux et articulaires.
Elle est essentielle à la motricité volontaire, au contrôle postural et à la conscience du corps.
Sa perturbation provoque une désorganisation motrice majeure et une altération du sentiment d’incarnation, illustrant son rôle central dans le lien entre perception, mouvement et conscience corporelle.
Rôle des fuseaux neuromusculaires dans le réflexe myotatique
Les fuseaux neuromusculaires jouent un rôle central dans la régulation réflexe du tonus musculaire, notamment à travers le réflexe myotatique (ou réflexe d’étirement).
Ce réflexe correspond à la contraction automatique d’un muscle en réponse à son propre étirement. Il s’agit d’un mécanisme involontaire et spinal, qui contribue à maintenir la posture, stabiliser les mouvements et protéger les muscles et tendons d’un étirement excessif.
Principe du réflexe myotatique
Lorsqu’un muscle est étiré, ses fuseaux neuromusculaires — récepteurs mécanosensibles situés au sein du muscle — détectent cette modification de longueur.
Ils transmettent alors des potentiels d’action via des fibres afférentes sensorielles Ia jusqu’à la moelle épinière.
Dans la moelle, ces fibres font synapse directement (réflexe monosynaptique) avec les motoneurones α qui innervent les fibres musculaires extrafusales du même muscle.
L’activation de ces motoneurones α provoque la contraction réflexe du muscle étiré, ce qui s’oppose à l’étirement initial et rétablit la longueur de repos du muscle.
C’est ce mécanisme qu’on observe lors du test du réflexe rotulien : un léger coup de marteau sur le tendon du quadriceps provoque un étirement rapide du muscle, suivi d’une contraction réflexe qui fait se lever la jambe.
Ce réflexe a donc une fonction protectrice : il limite les risques de lésions musculaires ou tendineuses dues à des étirements trop importants.
Rôle des fuseaux neuromusculaires dans le réflexe myotatique
Rôle des motoneurones α et γ
Le contrôle du tonus musculaire implique la coordination entre deux types de motoneurones :
Les motoneurones α innervent les fibres musculaires extrafusales, responsables de la contraction du muscle entier.
Les motoneurones γ innervent les extrémités contractiles des fibres intrafusales, c’est-à-dire les fibres à l’intérieur du fuseau neuromusculaire.
Lorsqu’un muscle s’étire :
Les fuseaux neuromusculaires sont activés → ils déchargent des potentiels d’action (activité afférente Ia).
Cette activité déclenche, dans la moelle épinière, l’activation des motoneurones α → contraction du muscle → diminution de l’étirement.
Lorsque le muscle se contracte, les fuseaux risquent d’être relâchés et donc de perdre leur sensibilité.
Pour éviter cela, les motoneurones γ contractent les extrémités des fibres intrafusales, ce qui retend le fuseau et lui permet de continuer à détecter les variations de longueur.
Ainsi, l’activation conjointe des motoneurones α et γ (appelée coactivation α-γ) garantit que les fuseaux neuromusculaires restent sensibles pendant toute la contraction musculaire, assurant une surveillance continue de la longueur du muscle.
Rôle des fuseaux neuromusculaires dans le réflexe myotatique
Codage de l’étirement et du mouvement musculaire
Les fuseaux neuromusculaires ne se contentent pas de signaler un étirement passif :
ils codent à la fois :
la longueur du muscle (information statique),
et la vitesse d’étirement ou de raccourcissement (information dynamique).
Lors d’un étirement, la fréquence des potentiels d’action des fibres Ia augmente proportionnellement à l’allongement.
Lors de la contraction, la coactivation γ retend les fuseaux, ce qui leur permet de continuer à décharger, reflétant ainsi la dynamique du mouvement musculaire.
Ce double codage permet au système nerveux :
de connaître en permanence l’état mécanique du muscle,
et d’ajuster en temps réel la force et la précision du mouvement.
Rôle des fuseaux neuromusculaires dans le réflexe myotatique
Exemple concret : le réflexe rotulien
Le réflexe rotulien illustre parfaitement ce mécanisme :
Le coup de marteau étire brièvement le tendon du quadriceps → étirement du muscle.
Les fuseaux neuromusculaires détectent cet allongement et envoient un influx afférent vers la moelle.
Activation monosynaptique des motoneurones α → contraction réflexe du quadriceps.
Coactivation γ → maintien de la tension dans les fuseaux, permettant une détection continue.
Ce réflexe assure un retour rapide à la longueur initiale du muscle, stabilisant ainsi la posture.
Les récepteurs proprioceptifs : les organes tendineux de Golgi
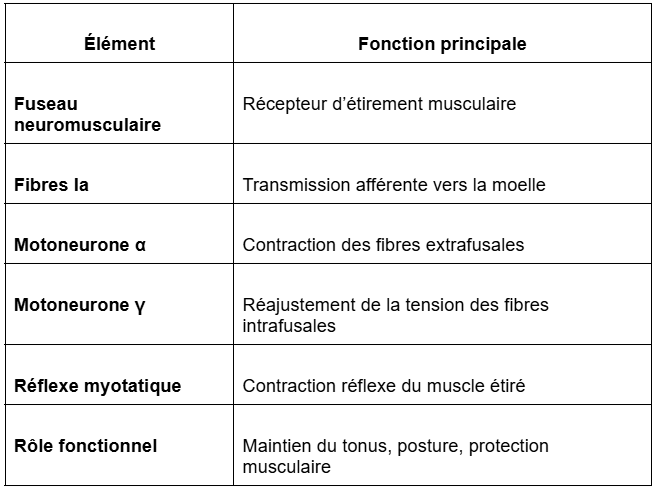
Les organes tendineux de Golgi sont des récepteurs proprioceptifs situés dans les tendons, c’est-à-dire à la jonction entre le muscle et l’os.
Ils sont constitués de fibres nerveuses afférentes de type Ib qui s’entrelacent entre les fibrilles de collagène formant le tendon.
Ces récepteurs sont mécanosensibles : ils réagissent à la déformation mécanique des tissus tendineux provoquée par une contraction importante du muscle.
Lorsque le muscle se contracte sous l’action des motoneurones α, il exerce une tension sur le tendon ; les fibrilles de collagène se tendent et compriment les fibres sensorielles Ib. Cette stimulation génère des potentiels d’action dont la fréquence est proportionnelle à la tension exercée sur le tendon.
Ainsi, les organes tendineux de Golgi codent la force de contraction musculaire à travers la tension tendineuse.
Cette information est complémentaire de celle fournie par les fuseaux neuromusculaires, qui eux, codent la longueur du muscle et ses variations d’étirement.
En d’autres termes :
les fuseaux neuromusculaires renseignent sur la position et la vitesse d’allongement du muscle ;
les organes tendineux de Golgi renseignent sur la force développée par la contraction musculaire.
Fonctionnement comparé avec les fuseaux neuromusculaires
Au repos, lorsque le muscle n’est pas contracté, ni les fuseaux neuromusculaires ni les organes tendineux de Golgi ne sont activés.
Lors d’un étirement, les fuseaux neuromusculaires s’activent et déclenchent le réflexe myotatique, provoquant une contraction réflexe du muscle.
Lors d’une contraction importante, les fuseaux deviennent inactifs (car les fibres intrafusales se détendent), tandis que les organes tendineux de Golgi s’activent fortement.
Ils déchargent alors des potentiels d’action proportionnels à la tension développée par le muscle.
Ce mécanisme permet au système nerveux de mesurer la force musculaire réelle.
Par exemple :
Lorsqu’un muscle se contracte fortement, la tension sur le tendon augmente, activant les récepteurs de Golgi.
De même, lorsqu’on maintient une position fixe sous une charge lourde, le muscle ne change pas de longueur mais reste fortement contracté : la tension tendineuse reste élevée, maintenant ainsi l’activité des organes de Golgi.
Rôle dans le réflexe myotatique inverse
Les organes tendineux de Golgi participent au réflexe myotatique inverse, aussi appelé réflexe d’inhibition autogène.
Ce réflexe est dit inverse car sa réponse est opposée à celle du réflexe myotatique classique :
Dans le réflexe myotatique, l’étirement du muscle entraîne sa contraction réflexe.
Dans le réflexe myotatique inverse, une contraction excessive du muscle entraîne au contraire son relâchement réflexe.
Concrètement, lorsque la tension exercée sur le tendon devient trop importante :
Les organes tendineux de Golgi sont activés et transmettent une influx afférent Ib vers la moelle épinière.
Ces fibres font synapse avec un interneurone inhibiteur, qui agit sur les motoneurones α du muscle concerné.
Cette inhibition provoque le relâchement réflexe du muscle, réduisant ainsi la tension exercée sur le tendon.
Ce réflexe a une fonction protectrice essentielle : il prévient les lésions musculaires ou tendineuses en limitant la force de contraction lorsqu’elle devient excessive.
Les récepteurs proprioceptifs : les récepteurs articulaires
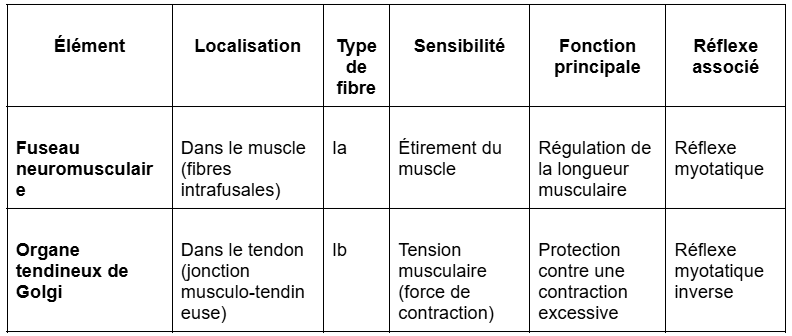
Les récepteurs articulaires constituent le troisième grand type de récepteurs proprioceptifs, après les fuseaux neuromusculaires et les organes tendineux de Golgi.
Ils sont localisés dans les tissus entourant les articulations, notamment dans les capsules articulaires, les ligaments et parfois au niveau des cartilages qui assurent la jonction entre les os.
Ces récepteurs sont des mécanorécepteurs sensibles aux changements de position et de mouvement des articulations.
Ils détectent des paramètres tels que l’angle articulaire, le degré d’ouverture, la direction ou encore la vitesse du mouvement.
Leur rôle est essentiel pour informer en continu le système nerveux central sur la position spatiale des membres et sur la cinématique des mouvements.
On distingue deux grands types de récepteurs selon leur vitesse d’adaptation :
Les récepteurs à adaptation rapide, qui répondent principalement aux changements dynamiques (déplacements, rotations, accélérations des articulations) ;
Les récepteurs à adaptation lente, qui transmettent une information plus statique, relative à la position maintenue de l’articulation lorsque le membre est immobile.
Les récepteurs proprioceptifs : les récepteurs articulaires
Sur le plan histologique et fonctionnel, les récepteurs articulaires incluent différents types de corpuscules sensoriels semblables à ceux de la sensibilité tactile cutanée :
les corpuscules de Golgi, sensibles à la tension exercée sur les ligaments ;
les corpuscules de Ruffini, réagissant à la pression prolongée et à l’étirement articulaire ;
les corpuscules de Pacini, sensibles aux vibrations et aux changements rapides de position ;
des terminaisons nerveuses libres, impliquées dans la nociception articulaire (douleur liée à des mouvements extrêmes ou à une inflammation).
Ces récepteurs fonctionnent selon les mêmes principes de transduction que les récepteurs mécanosensibles cutanés : une déformation mécanique de la membrane du récepteur entraîne l’ouverture de canaux ioniques mécanosensibles, provoquant une dépolarisation et la génération de potentiels d’action.
Les fibres proprioceptives primaires
Les signaux issus des récepteurs proprioceptifs (musculaires, tendineux ou articulaires) sont véhiculés par des fibres afférentes primaires de très grand diamètre.
Ces fibres — de type Ia (fuseaux neuromusculaires) et Ib (organes tendineux de Golgi) — sont fortement myélinisées, ce qui leur confère une vitesse de conduction très élevée, comprise entre 80 et 120 m/s.
Ainsi, un message nerveux proprioceptif peut parcourir environ 1 mètre en 10 millisecondes, assurant une transmission quasi instantanée de l’information sensorielle au système nerveux central.
Ces fibres empruntent les nerfs rachidiens et suivent des voies similaires à celles des fibres de la sensibilité tactile, bien qu’elles appartiennent à des neurones distincts et véhiculent une modalité sensorielle différente (proprioceptive et non tactile).
Rôle fonctionnel global
Les récepteurs articulaires, en interaction avec les fuseaux neuromusculaires et les organes tendineux de Golgi, permettent une intégration fine et permanente des informations relatives :
à la longueur musculaire (fuseaux),
à la tension musculaire (organes de Golgi),
et à la position et au mouvement des articulations (récepteurs articulaires).
Cette intégration fournit au cerveau une représentation continue et cohérente de la position du corps dans l’espace, indispensable à la coordination motrice, au maintien de l’équilibre et à la posture.
Voies et centres nerveux de la proprioception
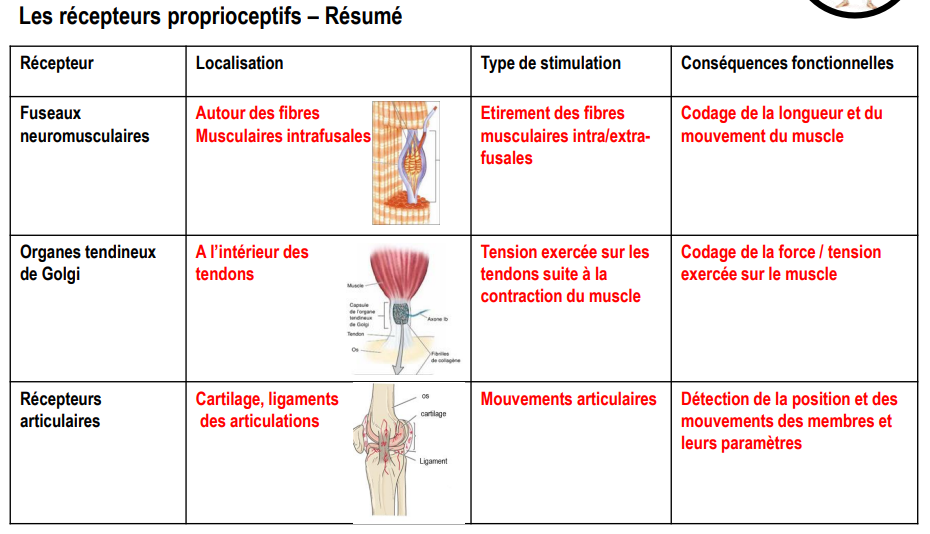
Les axones des récepteurs proprioceptifs (fuseaux neuromusculaires, organes tendineux de Golgi et récepteurs articulaires) se regroupent pour former les fibres sensorielles des nerfs rachidiens.
Ces fibres pénètrent dans la moelle épinière par la racine dorsale et constituent les neurones de premier ordre.
Une fois dans la moelle, ces neurones peuvent emprunter deux voies fonctionnelles principales :
Les boucles réflexes sensori-motrices :
Les neurones proprioceptifs peuvent faire synapse directement (ou via un interneurone) avec un motoneurone α, entraînant une réponse motrice réflexe.
C’est le cas notamment du réflexe myotatique (contraction du muscle en réponse à son étirement) et du réflexe myotatique inverse (relâchement du muscle lors d’une contraction excessive).
Ces circuits assurent des ajustements automatiques et inconscients du tonus musculaire, garantissant la protection du système musculo-squelettique.La transmission vers les centres nerveux supérieurs, pour l’intégration et le contrôle du mouvement.
Cette transmission emprunte deux voies ascendantes distinctes :la voie spinocérébelleuse, qui véhicule la proprioception inconsciente ;
la voie lemniscale, qui assure la proprioception consciente.
Voies et centres nerveux de la proprioception
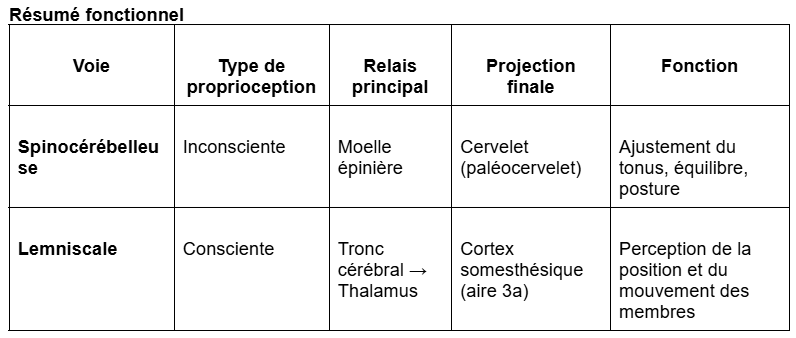
1. La voie spinocérébelleuse : la proprioception inconsciente
La voie spinocérébelleuse transmet au cervelet les informations issues principalement des fuseaux neuromusculaires.
Ces signaux renseignent sur le tonus musculaire et l’état des muscles et des articulations pendant la préparation et l’exécution d’un mouvement.
Selon l’origine corporelle du signal (membres inférieurs ou supérieurs), les fibres proprioceptives pénètrent dans la moelle épinière à des niveaux différents mais toujours par la racine dorsale.
Elles font synapse dans la substance grise de la moelle avec un neurone de second ordre, dont l’axone remonte ensuite dans le tronc cérébral pour atteindre le cervelet, généralement du même côté (ipsilatéralement) que la stimulation d’origine.
Le trajet peut être direct ou indirect (avec croisement puis retour du même côté).
Les informations ainsi transmises se projettent dans le paléocervelet (ou spino-cervelet), situé dans la partie médiale du cervelet.
Le cervelet utilise ces informations pour :
ajuster le tonus musculaire pendant le mouvement,
maintenir la posture et l’équilibre,
anticiper les conséquences motrices et sensorielles d’un mouvement.
Par exemple, avant l’exécution volontaire d’un geste (comme tirer sur une manette), le cervelet peut pré-activer les muscles posturaux (ex. muscles du mollet) afin de prévenir une perte d’équilibre.
Cette anticipation repose sur l’évaluation proprioceptive de l’état musculaire initial, transmise par la voie spinocérébelleuse.
Voies et centres nerveux de la proprioception
2. La voie lemniscale : la proprioception consciente
La voie lemniscale (ou voie des colonnes dorsales–lemnisque médian) est la même que celle empruntée par la sensibilité tactile fine (épicritique).
Elle véhicule les informations de toucher fin, de vibrations, et de proprioception consciente.
Les neurones de premier ordre (dans les ganglions rachidiens) projettent leurs axones dans les colonnes dorsales de la moelle épinière, jusqu’au tronc cérébral, où ils font synapse avec un neurone de second ordre.
Celui-ci croise la ligne médiane (décussation) et remonte vers le thalamus, où il fait relais avec un neurone de troisième ordre.
Ce dernier envoie ses projections vers le cortex somesthésique primaire (S1), situé dans le gyrus postcentral du lobe pariétal.
L’information proprioceptive se projette spécifiquement dans l’aire 3a de S1, distincte de l’aire 3b, dédiée à la sensibilité tactile.
Depuis l’aire 3a, les signaux proprioceptifs sont ensuite relayés :
vers S2 (voie ventrale), pour le traitement perceptif ;
vers le cortex pariétal postérieur (voie dorsale), pour l’intégration spatiale et la planification de l’action ;
et vers le cortex moteur primaire (M1), pour le contrôle précis des mouvements volontaires.
Ainsi, la voie lemniscale permet la conscience de la position des membres, tandis que la voie spinocérébelleuse assure le contrôle automatique et ajusté du mouvement.
Traitement des informations proprioceptives au niveau cortical
Les informations proprioceptives sont rapidement intégrées à d’autres modalités sensorielles, notamment tactiles. Cette intégration multisensorielle permet la réalisation de tâches complexes comme la stéréognosie, c’est-à-dire la reconnaissance d’objets en trois dimensions par le toucher.
Par exemple, lorsqu’on tient un objet dans la main les yeux fermés (une balle ou un cube), la reconnaissance de sa forme ne repose pas uniquement sur les informations tactiles locales (pressions exercées par l’objet sur différentes zones de la peau), mais aussi sur les informations proprioceptives relatives à la position et la configuration des doigts et de la main.
L’intégration de ces deux types d’informations s’effectue dès le cortex somesthésique secondaire (S2), où l’on observe des neurones multimodaux dont l’activité est modulée à la fois par des stimulations tactiles et par la position des doigts. Ces neurones répondent donc à des combinaisons spécifiques d’informations tactiles et proprioceptives.
S2 jouerait également un rôle clé dans la construction du schéma corporel, c’est-à-dire la représentation interne et dynamique du corps — indépendante de la perception consciente — sur laquelle reposent la connaissance de la taille et de la position relatives de nos membres.
Proprioception et action : rôle du cortex pariétal postérieur
Le cortex pariétal postérieur constitue une région d’intégration multisensorielle majeure. Il reçoit :
des informations visuelles issues de la voie dorsale (localisation et mouvement des objets dans l’espace),
des informations somesthésiques (tactiles et proprioceptives) sur la position et l’état du corps,
ainsi que des informations motrices provenant des régions prémotrices et préfrontales impliquées dans la planification du mouvement.
Grâce à cette intégration, le cortex pariétal postérieur élabore un modèle interne du corps dans l’espace et des actions à effectuer, permettant une coordination précise entre la perception et le mouvement.
Les lésions de cette région entraînent divers troubles :
Négligence spatiale (trouble de la représentation de l’espace),
Astéréognosie (incapacité à reconnaître un objet par le toucher),
Apraxie (difficultés d’exécution ou de coordination gestuelle),
Autotopoagnosie (incapacité à reconnaître ou localiser les parties de son propre corps).
Ainsi, l’information proprioceptive n’est jamais traitée isolément : elle est toujours combinée à d’autres informations sensorielles (visuelles, tactiles, motrices) afin de permettre la perception du corps, le maintien du schéma corporel et la planification des actions.
Modification du schéma corporel par la proprioception et les stimulations tactiles
Des expériences ont montré que la stimulation proprioceptive peut profondément modifier la représentation corporelle. Par exemple, l’application de vibrations sur les muscles ou les tendons, en l’absence de contraction musculaire volontaire et sans information visuelle, peut induire une illusion de mouvement.
Dans certaines configurations expérimentales, ces illusions produisent la sensation que certaines parties du corps — comme le nez ou la tête — s’allongent. Ce phénomène illustre la grande flexibilité du schéma corporel : la représentation que le cerveau se fait du corps peut se modifier très rapidement. Le système nerveux cherche toujours à donner une interprétation cohérente aux signaux sensoriels qu’il reçoit, même si cette interprétation est erronée sur le plan physique.
Une étude classique l’a démontré de manière frappante. Des participants, privés d’informations visuelles (yeux bandés), étaient placés dans différentes positions où ils recevaient simultanément :
une stimulation tactile, par exemple en posant leur doigt sur leur nez, et
une stimulation proprioceptive, par application de vibrations sur le tendon du biceps.
Ces vibrations activent artificiellement les récepteurs musculaires (fuseaux neuromusculaires), donnant au cerveau l’impression que le muscle est en train de s’étirer. Dans la configuration où le sujet se touche le nez, le cerveau interprète cette stimulation comme un allongement du bras. Cependant, comme le contact tactile entre le doigt et le nez persiste, la seule explication cohérente pour le cerveau est que le nez s’allonge.
D’autres configurations similaires — par exemple lorsque le sujet se touche la tête — induisent l’illusion d’un allongement du crâne ou d’un déplacement de la tête.
Ces illusions montrent que le cerveau reconstruit la perception du corps à partir des informations sensorielles disponibles, même lorsqu’elles sont ambiguës ou incohérentes. En l’absence de vision, il privilégie l’interprétation la plus probable pour maintenir une cohérence perceptive.
Modification du schéma corporel par la proprioception et les stimulations tactiles
Cette flexibilité contraste avec d’autres situations où le schéma corporel semble au contraire très stable et difficile à modifier.
Par exemple, chez les femmes enceintes, les changements rapides de morphologie ne s’accompagnent pas toujours d’une mise à jour immédiate de la représentation corporelle : certaines continuent à évaluer mal la taille de leur abdomen et se cognent aux portes ou aux objets.
De même, chez les personnes amputées, le membre fantôme illustre la persistance d’une représentation corporelle : malgré l’absence physique du membre, le cerveau continue à générer des sensations et des comportements comme si le membre existait encore (par exemple, éviter un obstacle en “levant” le bras manquant).
Ainsi, le schéma corporel présente une dualité remarquable :
il peut être à la fois rigide — résistant au changement lors de modifications réelles du corps — et extrêmement flexible, capable d’intégrer en quelques secondes des informations sensorielles trompeuses pour reconstruire une image cohérente du corps.
Troubles de la proprioception
Désafférentation: Neuropathie rare affectant les fibres somesthésiques de large calibre (proprioceptives et tactiles) – les fibres motrices et nociceptives restant intactes. Les patients rapportent la sensation de « flotter » ou de « désincarnement », mouvement possible (mais maladroit) avec effort sur la base de l’information visuelle.
Astéréognosie ou Agnosie tactile: Incapacité à identifier la forme d’un objet par le toucher en l’absence d’information visuelle sans trouble élémentaire de la somesthésie (anesthésie). Peut concerner une seule main – la reconnaissance est possible avec la main opposée. Généralement dû à une atteinte du cortex pariétal postérieur