Virologia Generale
1/27
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
28 Terms
Struttura e dimensioni dei Virus (VIRIONE)
Il VIRIONE:
è la particella virale che si libera dalla cellula dopo la replicazione
è costituita dal genoma virale racchiuso all’interno del capside (contenitore proteico)
alcuni virus presentano all’esterno del capside un involucro lipoproteico che chiamiamo envelope o peplos, che deriva dalla membrana della cellula ospite modificata con proteine virali
il virione si riveste dell’envelope per gemmazione → quindi si replica nella cellula ospite, poi gemma spingendo parte della membrana della cellula ospite che diventa il suo envelope
le proteine virali si organizzano in peplomeri che si proiettano all’esterno
Lo spazio tra envelope e capside si chiama Tegumento e contiene proteine e enzimi necessari:
all’ancoraggio dell’envelope al capside
per la replicazione del genoma virale
per i meccanismi di immunoevasione
DIMENSIONI:
Dimensioni molto variabili
Per i Deossiribovirus (a dna) → dai 18-26nm (parovirus) a 400nm (poxvirus)
Per i Ribovirus (a rna) → dai 28-30nm (picornavirus) a oltre 300nm (paramyxovirus)
Virioni più grandi → genoma più grande → codifica più proteine → struttura più complessa
Composizione Chimica dei Virus
Il genoma virale può essere a DNA o RNA:
Deossiribovirus (DNA) → DNA bicatenario lineare ad eccetto:
Papovavirus che ha DNA bicatenario circolare
Parvovirus che ha DNA monocatenario
Ribovirus (RNA) → RNA monocatenario lineare, ad eccetto:
Reovirus che ha RNA bicatenario
nei ribovirus più grandi il DNA può essere segmentato
Componente Proteica:
Rappresentata del capside e envelope
Involucri cui funzione è la protezione del genoma e penetrazione nella cellula ospite
Componente Lipidica:
Presente solo nei virus con envelope
Presenta un doppio strato lipidico simile alla membrana cellulare, infatti deriva proprio dalla cellula ospite
Carboidrati:
Presente nei virus con envelope
Questi sono legati a proteine virali formando glicoproteine (chiamate anche spicole o proteine spikes)
Capside
Economia genetica:
i virus hanno pochi geni, quindi producono poche proteine ripetute molte volte, infatti il capside è formato da sub-unità proteiche identiche
Le diverse sub-unità proteiche che formano il capside si chiamano capsomeri e sono uniti da legami non covalenti
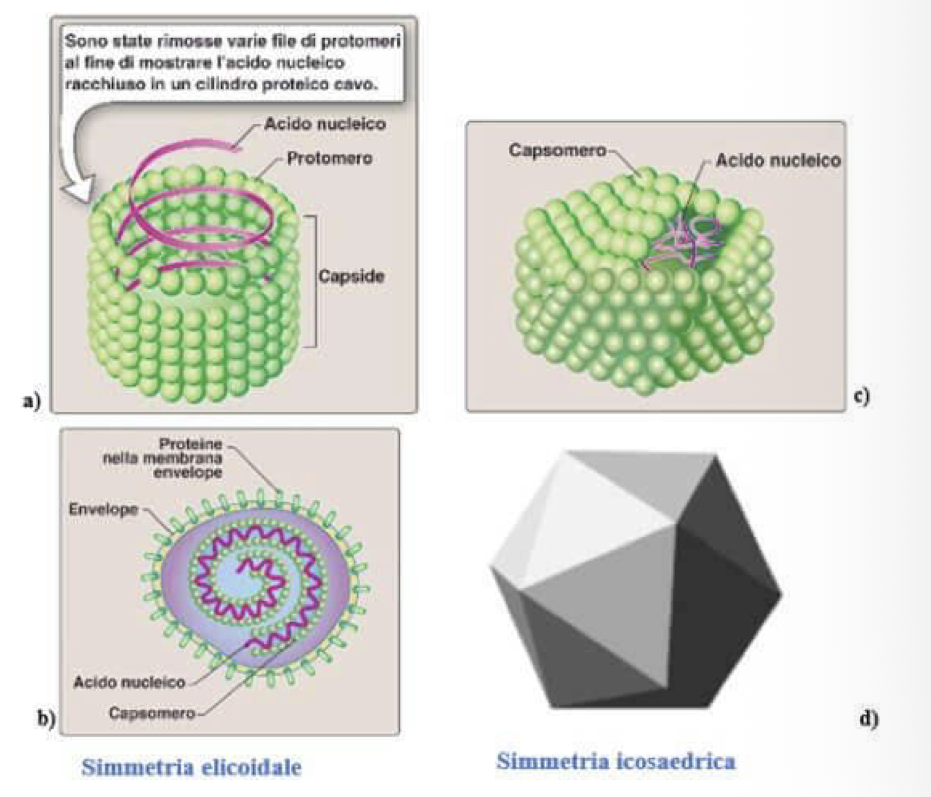
Tipi di simmetria del capside:
Simmetria Elicoidale → proteine disposte a spirale, formando una struttura a bastoncello, al cui interno troviamo l’acido nucleico (in genere RNA) associato ai capsomeri
non hanno l’envelope
Simmetria Icosaedrica → ogni vertice presenta un pentone (capsomero a 5 sub-unità), mentre tutto il resto presenta esoni (a 6 sub-unità)
negli adenovirus dai pentoni partono fibre che servono per l’attacco della cellula
in genere senza envelope
Simmetria Complessa → simmetria non del tutto chiarita
Ad esempio il Poxvirus ha forma ovoidale “a mattone”, dove il genoma è racchiuso all’interno di un nucleoide biconcavo
Strutture interne aggiuntive:
Proteine associate al genoma → ruolo nel processo di replicazione
Caratteri Antigenici
I virus sono molto antigenici perché sono composti per il 90% da proteine, quindi stimolano molto bene il sistema immunitario:
Gli antigeni del capside sono detti antigeni nucleo-proteici (NP), questi:
nei virus senza envelope sono esposti e determinano la specificità del virus
nei virus con envelope sono nascosti e visibili solo se l’envelope si rompe
Gli antigeni dell’envelope si trovano nei peplomeri e derivano dalla membrana della cellula ospite
I Batteriofagi
I batteriologi sono i virus che parassitano i batteri:
Come i virus sono costituiti da proteine e acido nucleico che può essere:
soprattutto DNA bicatenario (per i tipi A, B e C)
talvolta DNA (tipi D, F) o RNA monocatenario (tipo E)
Distinguiamo 6 tipi morfologici:
Tipo A:
E’ il più complesso
Presenta una testa poliedrica
Un collare con sei sottili setole (che unisce testa a coda)
Una coda con un tubo centrale rigido rivestito da guaina proteica contrattile
Una piastra esagonale terminale con spine ad ogni angolo
La pistra si connette a 6 lunghe fibre usate per attaccarsi ai recettori della parete batterica
Tipo B:
Presente una testa poliedrica e una coda lunga, flessibile e senza guaina contrattile
Può presentare appendici terminali
Tipo C:
Presenta una testa poliedrica una coda corta priva di guaina contrattile
Tipo D ed E:
Presenta una testa icosaedrica, privi di appendici
Il tipo D ha capsomeri più voluminosi del tipo E
Tipo F:
Ha un aspetto di un lungo filamento flessibile con nessun altra struttura associata
Classificazione dei Virus
Classificati in base all’ospite in:
Virus dei batteri (batteriofagi)
Virus degli animali → a loro volta suddivisi in virus degli artropodi e dei vertebrati
Virus dei vegetali
All’interno di questi gruppi, i virus vengono classificati in:
Famiglie
Sottofamiglie
Generi
Poi in base all’acido nucleico distinguiamo i Virus:
Deossiribovirus (a DNA)
Genoma formato da una solo molecola di acido nucleico
Le famiglie si classificano in base alla presenza o meno dell’envelope e alla struttura del genoma
I tipi principali di genoma sono il DNA bicatenario lineare (poxviridae, herpesviridae, adenoviridae), DNA bicatenario circolare (papillomaviridae, polyomaviridae), DNA monocatenario lineare (parvoviridae) e DNA parzialmente bicatenario circolare (Hepadnaviridae)
Ribovirus (a RNA)
Anche qui la famiglie si classificano in base alla presenza dell’envelope e struttura del genoma, che presenta una varietà maggiore.
I tipi principali di genoma sono RNA monocatenario non segmentato (coronaviridae, flaviviridae, togaviridae, astroviridae, caliciviridae, picornaviridae, filoviridae, paramyxoviridae, rhabdoviridae), RNA monocatenario segmentato (orthomyxoviridae), RNA bicatenario segmentato (reoviridae), RNA monocatenario diploide (retroviridae), RNA monocatenario segmentato circolare (arenaviridae, bunyaviridae)
Inoltre, nei ribovirus abbiamo un’altra differenziazione in base alla polarità dell’RNA, cioè RNA positivo se funga direttamente come mRNA, o RNA negativo se deve essere prima trascritto
Classificazione di Baltimore
Classifica i virus in base a come producono mRNA:
- Classe I, Virus a dsDNA (es. Poxviridae).
- Classe II, virus a ssDNA (es. Parvoviridae).
- Classe III, virus a dsRNA (es. Reoviridae).
- Classe IV, virus a ssRNA+ (es. Picornaviridae).
- Classe V, virus a ssRNA- (es. Orthomyxoviridae).
- Classe VI, retrovirus a ssRNA (es. Retroviridae).
- Classe VII, retrovirus a dsDNA (es. Hepadnaviridae).
La Moltiplicazione dei Virus
Avviene solo all’interno della cellula ospite, ed è particolare perché:
Il virus perde la sua struttura una volta nella cellula
Acido nucleico e proteine vengono sintetizzate separatamente e poi si assemblano formando nuovi virioni (nei virus a DNA, il dna nel nucleo e le proteine nel citoplasma)
Lo spettro di ospite:
Indica se il virus è in grado di infettare la cellula e in quale può moltiplicarsi
Le cellule sensibili sono quelle che possono essere infettate da un virus, ma affinché la infezione sia produttiva, la cellula deve essere anche permissiva, cioè che permette la replicazione completa del virus
Tipi di infezione virale:
Produttiva:
Quando la cellula è sensibile e permissiva
Si ha un ciclo di replicazione virale completa con progenie infettante
Spesso porta alla morte cellulare
Tipica dei virus patogeni
Restrittiva:
Quando la cellula è sensibile, ma permissiva solo in certe condizioni (come la fase S del ciclo cellulare, dove si ha più disponibilità dei fattori associati alla sintesi del DNA)
Un esempio è il Parvovirus B19, che si replica solo nei precursori eritrocitari nucleati quando sono altamente proliferanti
Latente:
Quando il DNA virale si integra nel genoma cellulare, ma può restare silente o riattivarsi in seguito
Tipica di Herpesviridae e Retroviridae
Abortiva:
Quando la cellula è sensibile, ma non completamente permissiva
Quindi il ciclo di replicazione virale è incompleto e non si formano nuovi virioni
Può comunque causare danno cellulare che conducono e necrosi o trasformazione
Fasi del Cilo di Moltiplicazione Virale
Attacco del virione sulla superficie della cellula (adsorbimento)
Penetrazione del virus all’interno della cellula
Esposizione dell’acido nucleico virale
Sintesi delle macromolecole virali
Assemblaggio dei virioni neo-prodotti e liberazione della progenie virale
Attacco dei Virus alla cellula (assorbimento)
Il primo contatto virus-cellula avviene per collisioni casuali, ma l’attacco sarà efficace solo se avviene il legame tra:
Anti-recettore virale (sul virus) e recettore cellulare (sull’ospite)
Nei virus con envelope l’anti-recettori sono rappresentati da glicoproteine (peplomeri), mentre in quelli senza da proteine del capside
I virus con ampio spettro d’ospite hanno recettori molto diffusi, quindi infettano molte cellule, mentre quelli con ristretto spettro d’ospite hanno recettori molto specifici, quindi infettano poche cellule
As esempio, il Parvovirus B19 usa come recettore l’antigene P dei globuli rossi
Gli individui senza questo recettore (rari) sono insensibili all’infezione
Ruolo degli ioni:
Un ambiente ricco di ioni rende l’attacco più efficace
Essi servono a ridurre la repulsione elettrostatica e facilitare il legame virus-cellula
Anticorpi:
Gli anticorpi più efficaci contro l’attacco sono quelli diretti contro l’anti recettore, infatti impediscono l’infezione
Secondo la teoria del canyon, l’attività degli anticorpi non risiede nel legame anticorpo/anti-recettore, ma nel legame anticorpo-epitopi antigenici in corrispondenza del bordo del canyon
l’ipotesi è che l’anti-recettore virale è nascosto in una depressione (canyon) della molecola proteica virale
Il recettore riesce ad entrare nel canyon, ma gli anticorpi no
Quindi quello che fa l’anticorpo è bloccare fisicamente l’accesso al recettore → risultato neutralizzazione del virus.
Alcuni virus hanno un meccanismo di assorbimento in due step:
Legame iniziale (bassa affinità) dove si lega un primo recettore che serve a portarlo vicino alla cellula
Legame definitivo (alta affinità) dove un secondo recettore (correttore) permette un attacco più stabile per consentire la penetrazione
Penetrazione ed Esposizione (uncoating) dell’Acido Nucleico Virale
La penetrazione:
Mentre l’adsorbimento è un processo passivo (può avvenire fino a 4°C), la penetrazione è un processo attivo che è possibile solo a temperature ottimali per la cellula.
Due modalità in base alla presenza dell’envelope:
Virus senza envelope → entrano nella cellula tramite traslocazione diretta attraverso la membrana o per endocitosi (in vescicole)
Virus con envelope → entrano per fusione dell’envelope con la membrana cellulare esterna o per endocitosi in vescicole e poi l’envelope si fonde con la membrana dell’endosoma
L’esposizione (uncoating):
Consiste nella liberazione del genoma virale nel citosol per renderlo accessibile alla cellula e avviare la replicazione
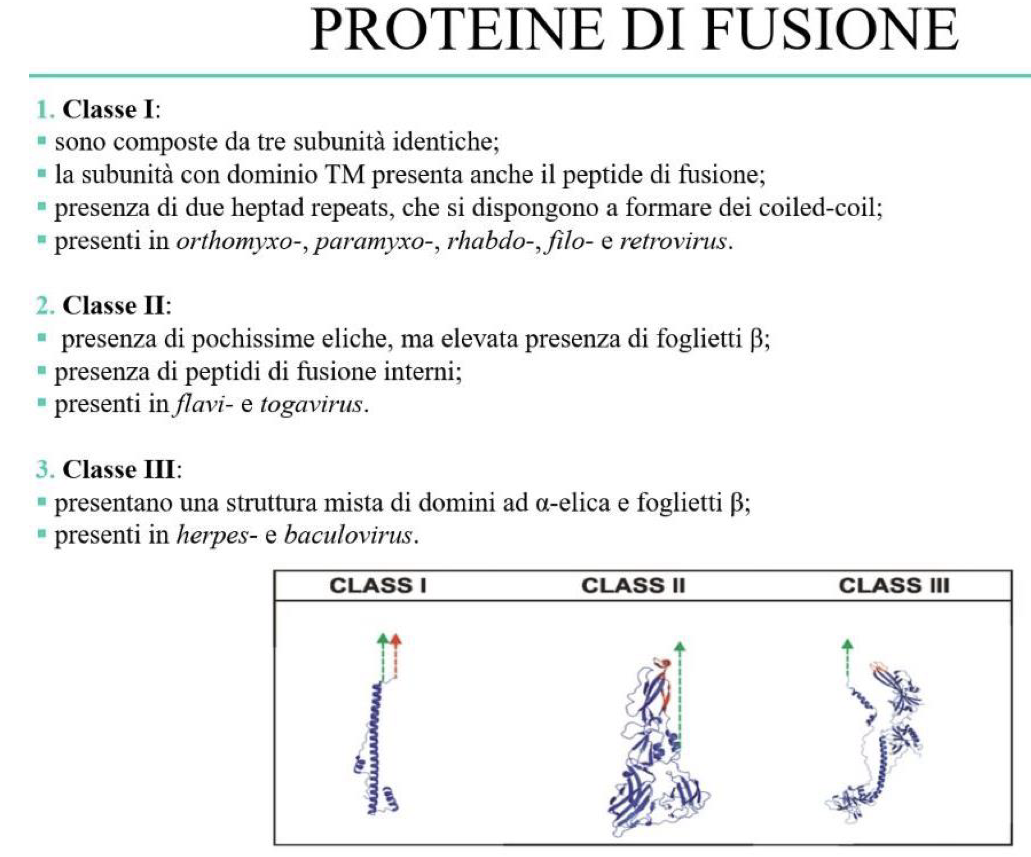
Nei virus con envelope → la fusione richiede proteine virali specializzate (proteine fumogene) che permettono la fusione delle membrane
la loro attivazione avviene a pH neutro e dall’interazione anti-recettore/recettore
Invece la fusione tra envelope e membrana eneolitica avviene a pH acido dovuto all’ambiente endosomale, grazie ad alcuni virus (come quello influenzale) che usa proteine che fungono da canali per H+
Curva di Replicazione Virale
Il tempo che intercorre tra infezione e rilascio della progenie dipende da:
Tipo del ciclo replicativo e fase del ciclo cellulare in cui si trova la cellula quando viene infettata
La replicazione virale non è schizogonica:
Cioè non si divide come i batteri, ma produce molti virioni da una singola cellula
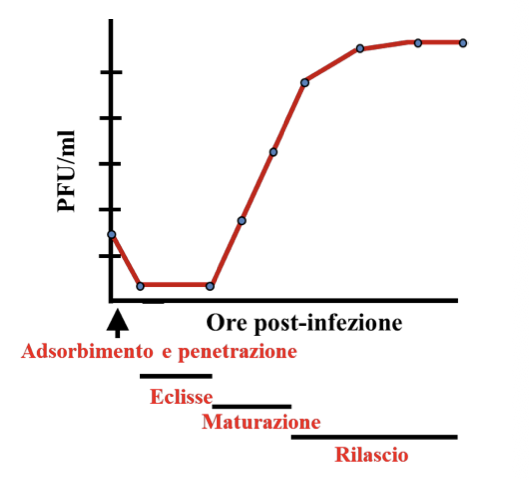
La curva di replicazione virale descrive:
Sull’asse delle X → tempo (ore post-infezione)
Sull’asse delle Y → quantità di virus (PFU/ml)
mostra come aumenta la produzione di virioni nel tempo
La progenie virale è omogenea (stessa struttura e dimensione)
Si distinguono 4 fasi:
Fase di adsorbimento e penetrazione → dove il virus si lega alla cellula ed entra al suo interno
Periodo di eclisse → poca quantità di virus, inizia l’uncoating e la replicazione del genoma virale
Periodo di maturazione → sintesi e assemblaggio dei virioni, le particelle virali si accumulano fuori e dentro la cellula. Se i virus sono citocidi, la cellula muore e la produzione si interrompe, se non lo sono la produzione continua
Fase di rilascio → la progenie virale viene rilasciata per lisi cellulare o gemmazione
Dalle Sbobine - I Vaccini e le presunte preoccupazioni
Ancora oggi si sentono preoccupazioni riguardo i vaccini per possibili effetti collaterali come:
Autismo, infarto, tumori, morte
Roberta Kwok riporta 4 livelli degli effetti collaterali dei vaccini:
Comuni:
dolore, rossore o gonfiore in sede di iniezione
LIEVE febbre → del tutto normale come attivazione del sistema immunitario
Meno comuni:
Febbre alta o rash cutaneo → come iper-risposta del sistema immunitario (come nel vaccino morbillo-parotide-rosolia)
Rari:
Anafilassi → reazione allergica grave e immediata, che richiede un trattamento urgente
Non concludenti:
Alcune persone hanno riferito mal di schiena dopo il vaccino Covid, ma è un dato non concludente perché non ne abbiamo una verifica statistica e clinica
Il legame tra vaccini e autismo:
Venne fatto uno studio su 12 bambini dove si pensò un collegamento tra autismo e vaccino del morbillo, da qui questa paura
Questo studio è stato poi ritirato perché aveva gravi problemi metodologici e quindi non fu considerato una prova scientifica valida
Oggi grandi studi non confermano il legame
Dalle Sbobine - Antigene
Antigene:
Comprende sia sostanze esogene che endogene (nel caso di patologie autoimmuni)
Principalmente sono di natura proteica, polisaccaridica e lipoproteica
Un antigene è in grado di indurre una risposta immunitaria (provocando la produzione di anticorpi o l’instaurarsi di una risposta cellula-mediata)
Differenza tra antigene e immunogeno:
L’immunogeno è qualunque sostanza in grado di suscitare una risposta immune
L’antigene è in grado di legarsi ad uno specifico anticorpo (o linfocita T)
solo alcuni antigeni sono in grado di stimolare la produzione di anticorpi specifici, altri si comportano da APTENI e riescono a farlo solo se legati ad una molecola Carrier che li fa diventare immunogeni
Epitopo o Determinante Antigenico:
Parte dell’antigene che entra in contatto con il sito di legame di un anticorpo o recettore per l’Ag delle cellule T
Dalle Sbobine - Il Vaccino
Il vaccino è un’immunità acquisita attiva artificiale e il suo mediatore è l’anticorpo.
Jenner introdusse il primo vero vaccino dell’epoca moderna contro il vaiolo (nonostante fosse già eradicato, siccome stato conservato per studiarlo), mentre Pasteur pose le basi della vaccinazione moderna sviluppando i principi alla base dell’immunizzazione:
Isolamento dell’agente causale della malattia
Inattivazione dell’agente
Inoculo dell’agente nel soggetto da immunizzare
Tipi di Vaccini:
Batterici
Attenuati: BCG, antitifo orale
Inattivati: antipertosse, colera
Anatossine: antidifterico, antitetanico
Antigeni di superficie: Hib, antipneumo e meningococco
Virali
Attenuati: antipolio orale, antirosolia, antimorbillo, antiparotite, antiamarilico
Inattivati completi: antinfluenzale, antipolio iniettabile, antirabbico
Antigeni di superficie: anti HBV
Dalle Sbobine - Evoluzione e tipi di Vaccini
Evoluzione dei vaccini:
Si è passati da un approccio empirico ai vaccini ricombinanti, ai vaccini coniugati e poi più recentemente alla reverse vaccinology
Oggi possiamo distinguere tra:
Vaccini inattivati
Contengono virus o batteri uccisi, privi di virulenza, patogenicità ed infettività, ma conservano l’antigenicità
Antipolio, Anti-epatite A, anti-influenzali, antirabbico, antitifico, anticolerico, anti-ricketsia provazeki (tifo petecchiale)
Se ho un virus da inattivare uso il calore e la formalina, quindi il virus viene inattivato e “smembrato”, di conseguenza il corpo non lo vede più nel suo assetto naturale e può produrre anticorpi contro tanti pezzi diversi del microrganismo → la risposta quindi è meno efficace e duratura, ma in compenso sono vaccini facili da trasportare e distribuire
Vaccini vivi attenuati
Contengono virus o batteri vivi con virulenza e patogenicità ridotta, capaci di indurre una risposta immunitaria più forte e duratura, ma senza causare la malattia
Antipolio, antimorbillo, antiparotite, antirosolia, antitubercolare, antitifico, anticolerico
Per farlo ci vogliono circa 10 anni
Vaccini a subunità
Contengono solo una parte del patogeno (come una proteina di superficie o una tossina inattiva), quindi non possono causare l’infezione, ma possono indurre la produzione di anticorpi
Per l’epatite B, influenza, rosolia
Per fare il vaccino dell’epatite B si usano tecniche di DNA ricombinante: il gene che codifica l’antigene di superficie viene inserito in cellule produttrici, che sintetizzano grandi quantità di proteina, che viene purificata e poi usata come vaccino
Vaccini a genoma nudo
Sono vaccini in cui viene somministrato direttamente il materiale genetico (DNA o RNA) senza un vero involucro protettivo
Il problema è che così il materiale genetico è fragile e facilmente degradabile da DNasi e Rnasi, quindi per superare questo limite, ad esempio nei vaccini a mRNA del Sars-Cov-2, l’mRNA viene racchiuso in nanoparticelle lipidiche proprio per proteggerlo dalla degradazione, così nella cellula verrà tradotto in proteina virale
Nel vaccino dell’HPV la strategia è diversa, si prendono i geni che formano il capside e lo si inserisce in una cellula di lievito → quest’ultima producono particelle virali vuote (VLPs) che fungeranno da contenitore per il virus.
Dalle Sbobine - Come si fanno oggi i vaccini?
Ad oggi il processo è accelerato grazie alla reverse vaccinology:
In questo approccio si parte dal genoma del patogeno, lo si analizza al computer per identificare le proteine più promettenti, soprattutto quelle esposte al sistema immunitario, conservate tra i ceppi e non simili a proteine umane
Il computer ci dirà quali pezzi di proteina possono essere riconosciuti da anticorpi e test di laboratorio testano i candidati, cioè verifichiamo de le proteine attivano il sistema immunitario
Il primo vaccino sviluppato con questa tecnica è quello contro il meningococco B
Dalle Sbobine - Cosa si intende per Patogenicità?
Con patogenicità si intende la sua capacità di causare un danno all’ospite (malattia), e dipende da caratteristiche del microrganismo e dell’ospite.
La sensibilità dell’ospite al parassita dipende da età, sesso, razza, genetica, nutrizione, risposte immune, malattie in atto, abuso di farmaci.
Un determinato microrganismo può avere patogenicità diverse in base al genere (uomo/donna)
Sia patogenicità che sensibilità dell’ospite variano continuamente:
Ad esempio, i virus epatotropi, soprattuto epatite B e C, hanno effetti diversi se si sviluppano in età pediatrica rispetto l’età adulta
L’epatite B ha più probabilità di cronicizzare in età pediatrica rispetto l’età adulta
il meccanismo di ciò non è ancora chiaro
Dalle Sbobine - Cos’è la Virulenza?
La virulenza è la misura quantitativa della patogenicità:
Abbiamo tre tipologia di determinanti di virulenza, che sono caratteristiche microbiche in grado di causare la malattia:
Strutturali → capsula, parete cellulare, LPS, proteine di membrana esterna
Biochimici → produzione di esotossine, metaboliti, o enzimi ad azione tossica
Genetici → plasmidi trasmissibili che determinano virulenza o antibiotico resistenza
La virulenza di un microrganismo deriva dalla combinazione di:
Invasività → capacità di proliferare nei tessuti raggiungendo concentrazioni che inibiscono le funzioni dell’ospite
Tossigenicità → capacità di produrre tossine, molecole in grado di danneggiare l’ospite
Il livello di virulenza è determinato da:
Numero di microrganismi infettanti (dose)
Presenza di difese dell’ospite (sensibilità dell’ospite)
Fattori di virulenza
Come si misura la virulenza?
Mediante l’impiego di cavie, ad oggi in riduzione grazie all’uso di organoidi
gli organoidi sono elementi cresciuti il laboratorio, una struttura tridimensionale che mima un organo, cioè composto quanto più possibile dalle stesse cellule dell’organo
Mediante l’impiego di cavie, valutiamo:
Infettività → numero minimo di microrganismi necessario per causare l’infezione (dose minima infettante)
Gravità della malattia → misurata in dose letale 50 (minima dose in grado di causare la morte nel 50% degli animali infettati) e dose infettiva 50 (minima dose in grado di causare infezione nel 50% degli animali infettati)
In alcuni casi, coltivare in laboratorio comporta uno svantaggio, perché il microrganismo si adatta e perde virulenza, cioè diventa meno capace di infettare rispetto al ceppo reale, quindi c'è il rischio di misurare un ceppo indebolito e avere una ID50 falsata
Dalle Sbobine - Il processo patogenetico
Il processo patogenetico prevede 4 fasi:
Esposizione → ospite esposto al microrganismo
infezione esogena → dall’ambiente (acqua, aria, terreno)
infezione endogena → secondaria a traslocazione microbica)
infezione zoonotica → trasmessa da animali
L’interazione può avvenire per contatto diretto (persona a persona) o indiretto tramite vettori animati (insetti) e inanimati (fomiti, oggetti)
Adesione → interazione microrganismo e gli epiteli dell’ospite che causa la patogenesi
Invasione → dei tessuti dove il microrganismo prolifera e produce danno mediante enzimi e/o tossine
Induzione di una risposta infiammatoria con reazioni fiprotiche ed allergiche dell’ospite e danno tessutale
L’infezione può risolversi spontaneamente oppure progredire in malattia
Dalle Sbobine - Vie di accesso per la penetrazione del microrganismo
Il microrganismo può entrare nell’ospite attraverso la cute o le mucose, sfruttando le porte d’ingresso.
Quando respiriamo, entriamo in contatto con numerosissimi microrganismi, ma non ci infettiamo grazie ai meccanismi di difesa dell’apparato respiratorio:
Peli e ciglia del naso trattengono molte particelle
Se respiriamo con la bocca abbiamo il lisozima e altre sostanze che degradano i microrganismi
Nei polmoni abbiamo il surfactante che lubrifica e ostacola l’adesione dei microrganismi all’epitelio respiratorio
un alterazione di queste difese favorisce le infezioni
Anche il tratto urogenitale rappresenta una via di ingresso:
Le infezioni urinarie sono più frequenti nelle femmine per la minore lunghezza dell’uretra e maggiore vicinanza alla regione perianale, che facilita il passaggio dei microrganismi intestinali
Altra vie di ingresso si formano:
Attraverso ferite, abrasioni, ustioni o altre soluzioni di continuo (streptococchi, HBV, Leptospira, P.aeruginosa)
Trami accidentali (clostridium tetani)
Puntura di vettore artropode (zanzare, zecche, pulci) (bartonella bacilliformis, rickettsie)
I pazienti ustionati:
Rischio di infezione altissimo, perché viene meno la barriera cutanea
Il lavaggio delle mani:
Rappresenta la principale via di trasmissione delle infezioni
Dalle Sbobine - Meccanismi di difesa aspecifici della Cute
Meccanismi Fisici:
Integrità e spesso della cute
Meccanismi Chimici:
Peptidi ad attività antibatterica (B-defensine)
Acidità gastrica e delle secrezioni sudorifera e sebacea
Attività antibatterica di lisozima/fosfolipasi A in liquidi biologici (lacrime, saliva, sudore)
Meccanismi Meccanici:
Turn-over strato corneo (rimozione meccanica)
durante l’accrescimento della pelle, superficialmente si forma uno strato di cheratina non vitale che ci protegge → si squama, si allontana meccanicamente e viene rimpiazzata in continuo dagli strati inferiori
Meccanismi Biologici:
Competizione con il microbiota residente
Attività pro-infiammatoria e antistreptococcica dei cheratinociti
Dalle Sbobine - Meccanismi di difesa aspecifici della Mucosa
Meccanismi Fisici:
Integrità di mucosa
Secrezioni di mucose (filtro)
Barriera chimica → si lega ai microrganismi per sequestrarli alle cellule sensibili
Meccanismi chimici:
Attività antibatterica del lisozima e fosfolipasi A presente nei liquidi biologici (lacrime, saliva, sudore)
pH acido (gastrico, vaginale, intestinale)
Il batterio Helicobacter Pylori è resistente al pH acido gastrico, quindi attacca la mucosa e causa ulcere intestinali
Meccanismi Fisici (di detersione):
Flusso unidirezionale dei fluidi in organi cavi (urine)
Progressione contenuto del lume (feci e urine)
Sistema mucociliare (mucose respiratorie)
Turn-over di cellule epiteliali
Meccanismi comportamentali e neurologici di difesa come tosse e vomito
Meccanismi Biologici:
Competizione con il microbiota residente
Risposta immunitaria umorale (IgA secretorie) e cellulo-mediata (macrofaci alveolari)
Perché il miglior vaccino è quello che segue la via naturale? Nel corso di un infezione naturale produciamo sia IgM al momento dell’infezione, che IgA dalle mucose, se invece facciamo un vaccino produciamo solo IgG e questo ci dà una protezione minore
Dalle Sbobine - Colonizzazione delle Mucose
Prima di tutto, serve l’attraversamento o distruzione della mucosa per raggiungere l’epitelio, questo avviene attraverso:
motilità flagellare (H.pylori, salmonella, E.coli, V.cholerae)
produzione di macinasi (V.cholerae)
Poi abbiamo l’interazione adesina-recettore:
Adesine (batteriche) → sono proteine (fibrine, fimbrie, pili, proteine di superficie)
Recettori (cellula ospite) → proteine di matrice intercellulare, craboidrati, glicolipidi, glicoproteine
possono avere specificità, cioè il recettore CD4 linfocitario per HIV, che però può infettare anche altre cellule, quindi si ha sia un recettore principale che secondario (backdoor)
il recettore del SARS-COV-2 è il recettore ACE2
Interazione specifica:
E. Coli uropatogeni posseggono pili 1 (che legano D-mannoso uroteliale) e pili P (che legano l’antigene P dei gruppi sanguini
E. Coli enteropatogeni posseggono proteine CFA (fattori antigenici di colonizzazione) leganti l’epitelio dell’intestino tenue
N. gonorrhoeae lega tramite la proteina superficiale OPA, la proteine CB66 dell’epitelio urogenitale
Dalle Sbobine - Penetrazione ed Invasione
Per penetrare ed invadere, bisogna rompere i meccanismi di difesa dell’ospite:
Tossine ciliostatiche causano asincronia e paralisi ciliare respiratoria (P. aeruginosa
Resistenza al killing fagocitario (M. tuberculosis)
Resistenza alla acidità gastrica (H. pylori attraverso ureasi)
Distrutto il meccanismo di difesa, segue la migrazione batterica verso la sottomucosa dove avremo distruzione epiteliale mediata da esoenzimi:
Ialuronidasi (S. aureus) → idrolizza acidi ialuronici, mucopolisaccaridi acidi della matrice extracellulare del tessuto connettivo
Collagenasi (Clostridium) → distrugge il tessuto connettivale
Pneumolisina (S. pneumoniae) → distrugge le cellule epiteliali ciliate del polmone
Mucinasi (H. pylori) → degrada la mucosa gastrica
Può esserci anche penetrazione senza danno fisico:
Neisseria spp. → penetra l’epitelio per endocitosi
Haemophilup spp. → penetra l’epitelio attraverso le giunzioni intercellulari
Shigella spp. (Yersinia spp.) → aderiscono alle cellule M della mucosa intestinale inducendo polimerizzazione dell’actina. Si formano così pseudopodi che internalizzano il batterio che successivamente sfugge al vacuolo fagocitato e proliferando nel citoplasma induce apoptosi della cellula
Salmonella spp. → invadono la mucosa colica grazie a proteine effettrici
Una volta raggiunti i tessuti, inizia la replicazione per raggiungere una concentrazione soglia in grado di manifestare la patologia, ma affinché ciò avvenga, il batterio deve adattarsi alle condizioni trovate nell’ospite che dipendono da:
Condizioni fisiche:
Temperatura
pH
Pressione Osmotica
Condizioni chimiche:
disponibilità di sostanze nutritive
disponibilità di oligoelementi, soprattuto il ferro per le reazioni enzimatiche del metabolismo microbico
riguardo a ciò, la lattoferrina è una sostanza che sequestra ferro, quindi inibisce la replicazione microbica
quindi sia l’ospite che il microrganismo competono per il ferro
Le condizioni più significative per la virulenza dipendono da:
disponibilità di ferro e temperatura (a temperatura ambientale non attivano i geni dei fattori di virulenza 25-30°C)
Dalle Sbobine - Diffusione dell’Infezione
Dopo la moltiplicazione con l’invasione dei tessuti, il passo successivo del processo patogenetico è la diffusione nell’organismo, che avviene attraverso:
Sangue
i patogeni possono circolare come microrganismi liberi (esposti direttamente alle difese dell’ospite) o protetti in cellule circolanti (come EBV, rosolia, brucella in linfociti o plasmodium in eritrociti)
la presenza transitoria di piccole quantità di microrganismi nel sangue lo definiamo batteriemia, normalmente eliminata dai macrofagi del sistema retocolo-endoteliale (milza-fegato)
se poi riescono a sopravvivere a ciò, continuano a moltiplicarsi e raggiungiamo la setticemia, potenzialmente fatale
Nervi (Rhabdovirus, HSV)
Liquor (N. meningitidis, H. influenzae, S. pneumoniae)
Linfa (Sporothrix)
Dalle Sbobine - Tossigenicità
I batteri producono sostanze che causano danno all’ospite, manifestando la malattia:
Tossine → sempre tossiche, responsabili della sintomatologia morbosa dell’infezione
Aggressine → non necessariamente tossiche, promuovono la crescita batterica inibendo le difese dell’ospite
Distinguiamo Patogeni Veri e Opportunistici:
Patogeni VERI (o franchi)
producono sempre una malattia
non richiedono un fattore predisponente
Chlamydia, N. gonorrhoeae, Influenzavirus
Patogeni OPPORTUNISTICI (o occasionali)
Inducono malattia solo in immunodepressi
Richiedono almeno un fattore di rischio
Possono derivare dall’ambiente o dalla flora autoctona
Pseudomonas spp., S. Aureus, S. Maltophilia, funghi, miceti
Dalle Sbobine - Classificazione e sede delle infezioni
Patogeni Extracellulari:
Causano malattia crescendo solo al di fuori delle cellule dell’ospite
Quindi generalmente uccisi dai fagociti
Virulenza determinata dalla capsula
Patogeni Intracellulari Facoltativi:
Patogeni a riproduzione intracellulare preferenziale o occasionale
Possono crescere anche in sede extracellulare
Virulenza multifattoriale
Patogeni Intracellulari Obbligati:
A riproduzione esclusivamente intracellulare
Virulenza multifattoriale
Patogeni Tossigenici:
Causano la malattia principalmente attraverso la produzione di esotossine