Looks like no one added any tags here yet for you.
Vad används för att visa evolutionära förhållanden mellan organismer, proteiner och gener.
Evolutionträd. Längden på varje gren är proportionell mot deras evolutionära distans. För proteiner och gener baseras detta på hur lika deras sekvenser är. T.ex alla hemoglobiner kommer från samma evolutionära linje (de delar samma stamfader)
Förklara divergent evolution
När ett protein utvecklas till protein X och protein Y. De båda proteinerna delar stamprotein och kallas därmed för homologer.
Varför är homologi inte samma sak som likhet mellan proteiner
Likheter i aminosyrasekvenser mellan proteiner kan tyda på homologi men det kan inte betyda att de två proteinerna är homologer. Två proteiner måste dela en stamprotein för att räknas som homologer.
Förklara ortologer
Ortologer är homologa proteiner som finns i olika organismer. De har oftast samma funktion
Förklara paraloger
Det är homologa proteiner som finns i samma organism. De kan ha olika funktioner i organismen
Förklara konvergent evolution
Det är organismer som inte har samma stamfader men som har samma “lösning” på problem. T.ex flygande insekter och fåglar. De har inte samma stamfader men de har båda vingar vilket gör att de kan flyga. Deras proteiner är därför inte heller homologer
Varför är proteinhomologi viktigt att studera
Homologistudier ger oss en inblick på evolution och proteinutveckling. Det hjälper även till att hitta eventuella egeneskaper, funktioner och strukturer hos okände aminosyrasekvenser. Genom att använda liknande proteiner kan man lära sig mer om proteiner som inte vet mycket om.
Hur kan man se ifall två proteiner är homologer
Genom att jämföra aminosyrasekvenser och 3D-strukturer = bioinformatics
Förklara sekvensanpassning
När två peptider/aminosyrasekvenser jämförs måste de oftast anpassas på olika sätt för att likheter ska kunna hittas i sekvensen. Stora likheter indikerar på homologi
Hur kan man anpassa två aminosyrasekvenser för att hitta matchningar
Genom att flytta sekvenserna så att flest matchningar hittas. Vi vissa regioner av sekvenserna måste även “gaps” introduceras vilket innebär att ena sekvensen hoppas längre fram till där den matchar bättre med resten av den andra sekvensen.
Hur hittar man den bästa (flest matchningar) sekvensapassningen
Genom att använda ett poängsystem som mäter hur väl sekvenserna har blivit anpassade —> den anpassningen med flest poäng är den bästa. Poängsystemet baseras på att för varje matchning fås ett visst poäng och för varje gap som tillsätts dras det av med poäng
Hur vet man ifall likheten mellan två sekvenser är signifikant, alltså inte beror av slumpen
Tumregeln är att för sekvenser med 100 aminosyror är 25% identitet signifikant och en indikation på homologi. Men det kan fortfarande vara fel. Att ha lägre än 25% innebär utesluter inte heller homologi för detta är bara en indikation.
Ett mer säkert sätt att undersöka är också sequence shuffling
Förklara sequence shuffling
En av sekvenserna blandas runt så att nya kombinationer av sekvensen matchas med den andra sekvensen. Poäng för varje möjlig sekvenskombination registreras. Om poänget för den orginala sekvensanpassningen är signifikant högre än resten av de blandade sekvenserna så är anpassningen signifikant —> proteinerna är förmodligen homologa
Förklara Blosum 62 matrisen
Det är ett mer precist poängsystem för sekvensanpassning. Det är en matris som ger/tar olika mycket poäng för varje matchning/icke-matchning. På detta sätt tar den även hänsyn till konservativ substitution i sekvenserna. Detta innebär att vissa aminosyror som har substituerats i ena sekvensen fortfarande kan ge poäng om de är kemisk lika (dessa substitutioner sker oftare). Dessa substitutioner påverkar oftast inte proteinet eftersom de är kemiskt lika och därför kan de fortfarande ge poäng i poängsystemet. Poängen baseras på hur sannolikt substitutionen är och relateras till den evolutionära distansen. Större, radikala substitutioner tar längre tid än konservativa substitutioner. Därför ger substitutioner med liknande kemiska egenskaper mellan aminosyrorna högre poäng.
Vad har gap för innebörd i den evolutionära distansen
Varje gap förknippas med evolutionära distans. Därför vill man ha färre och större gaps än många små gaps i en sekvensanpassning om vill hitta likheter.
Vad ska man titta på förutom sekvensen för att hitta homologi
Proteinstrukturerna. Det finns färre strukturer än sekvenskombinationer (strukturen är stabilare) och därför är strukturjämförelser mer känsliga.
Många enzymer följer Michaelis-Menten-kinetiken. Beskriv ekvationen
E + S k1→←k-1 ES k2→ E + P
V = Vmax * [S]/(KM + [S]) där
KM = (k-1 + k2) / k1
Vid V = Vmax/2 gäller att KM = [S]
Vilka enzym regulatoriska strategier finns det
A) Skapa nya enzymer inklusive isoenzymer (långsam reglering)
B) Reglering av redan existerande enzymer (snabb reglering):
Allosterisk kontroll
Reversibel kovalent kontroll
Proteolytisk aktivitet
Hur fungerar den första regulatoriska strategin (A)
Den är en ganska långsam reglering.
1) Enzymsyntes: alltså transkription/translation
2) Enzymnedbrytning
3) Syntes av en specifik isoenzym
Förklara isoenzymer
Isoenzymer är homologer och har en annan aminosyrasekvens men katalyserar samma reaktion som en annan enzym. De har andra kinetiska parametrar (KM) och finjusterar metabolismen och ser till behoven för sina specifika vävnader. T.ex hexokinas (i de flesta celler) har hög affinitet till glukos och är alltid aktiv. Medan glukokinas (i levern) har lägre affinitet för glukos och är aktiv vid höga glukoshalter → hjälper till att reglera blodsockernivån
Förklara allosterisk kontroll
Allosteriska regulatorer (effektorer) binder till enzymet (allosterisk enzym) som sedan antingen aktiverar eller inhiberar enzymaktiviteten. Effektorn binder reversibelt på ett annat säte än det aktiva sätet med en icke-kovalent bindning
Allosteriska enzymer har vissa egenskaper. Beskriv de
De har en T- (tense; minst aktiv) och R-stadie (relaxed; mest aktiv)
Inbindning av substrat eller allosterisk regulator påverkar skiftet mellan T- och R-stadie
De består av flera subenheter som samarbetar tillsammans
Att subenheterna samarbetar tillsammans ger ett S-kurva aktivitet → substratkoncentrationen är en sigmoid funktion
De följer alltså inte Michaelis-Menten kinetik utan sigmoid funktion
De får stora förändringar i den tertiära strukturen
Allosterisk reglering är ett effektivt sätt att reglera dessa proteiner
Det finns två modeller för allosterisk kontroll: samordnad och sekventiell
Både den samordnade och den sekventiella modellen beskriver hur allosteriska enzymer regleras genom konformationsändringar vid inbindning av ligander. Förklara den samordnade modellen
Alla subenheter i enzymet existerar samtidigt i antingen en T-form (tense, låg affinitet) eller en R-form (relaxed, hög affinitet).
När en ligand binder, skiftar alla subenheter samtidigt till R-formen.
Modellen föreslår alltså en "allt eller inget"-princip, där enzymet snabbt växlar mellan de två tillstånden.
Både den samordnade och den sekventiella modellen beskriver hur allosteriska enzymer regleras genom konformationsändringar vid inbindning av ligander. Förklara den sekventiella modellen
Enzymets subenheter kan ändra konformation en i taget, snarare än alla samtidigt.
När en ligand binder till en subenhet, påverkar det grannsubenheterna, vilket gradvis ökar deras affinitet för ytterligare ligander.
Detta möjliggör mer stegvis och flexibel reglering av enzymaktiviteten.
Förklara feedback-inhibtion mha ATCase (aspartate transcarbamoylase)
Feedback-inhibtion är när slutprodukten i en syntes inhiberar ett proteins aktivitet genom att allosteriskt binda till den.
ATCase katalyserar en reversibel reaktionen mellan carbamoylfosfat och aspartat där N-carbamoylaspartat bildas. Denna reagerar sedan i en serie reaktioner till slutprodukten cytidintrifosfat (CTP). När tillräckligt med CTP har bildats binder den allosteriskt till ATCase och hämmar dess aktivitet.
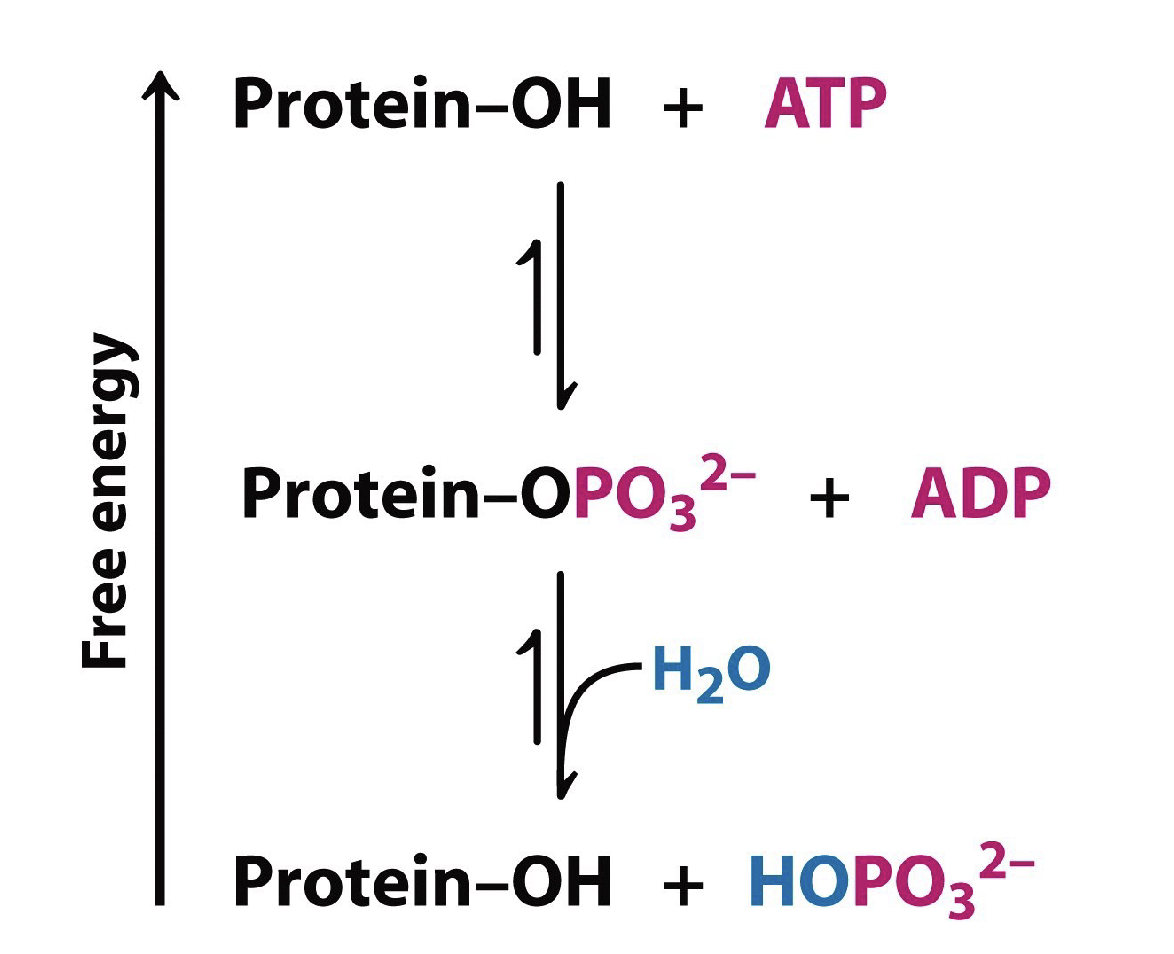
Fosforylering/defosforylering är ett vanligt sätt för reversibel kovalent modifiering. Beskriv vad de är
fosforylering/defosforylering aktiverar/deaktiverar proteiner
fosforylering sker av proteinkinaser
desfosforylering sker av proteinfosfataser
Det leder till en stor förändring i proteinets egenskaper: ±2 negativ laddning och ±2 vätebindningar
Både snabb och långsam reglering beroende på kinas/fosfataset
Kinaser kan antingen vara specifika för vissa proteiner eller påverka många olika proteiner (t.ex proteinkinas A)
Människor har mer än 550 kinaser
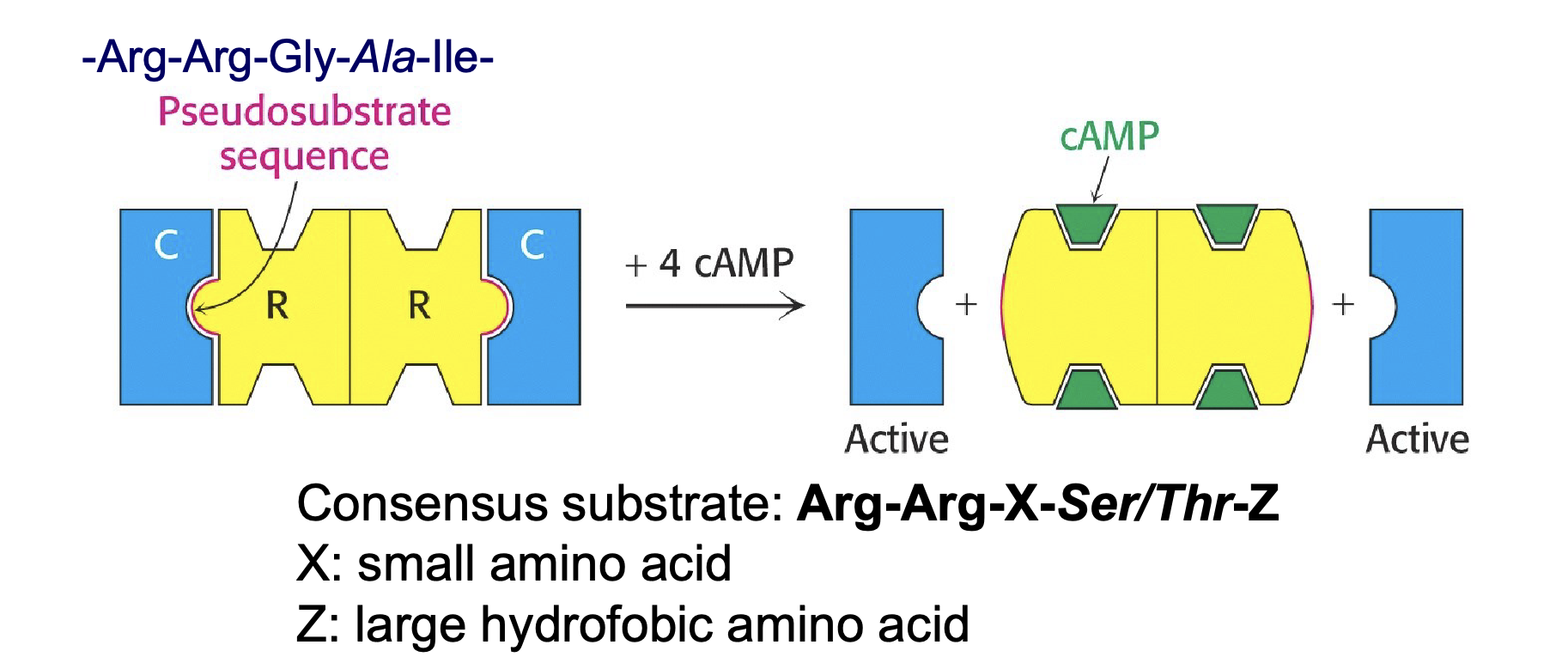
Beskriv proteinkinas A (PKA)
fosforylerar många olika porteiner
PKA aktiveras med cAMP som är en intracellulär messenger (second messenger)
Den aktiveras som respons till en hormonell stimuli: t.ex “fight-or-flight”
Den består av två regulära och två katalytiska subenheter
De regulära och katalytiska subenheterna är bundna genom en pseudosubstrat sekvens (-Arg-Arg-Gly-Ala-Ile-)
När cAMP binder till de regulära subenheterna bryts bindningen mellan regulära och katalytiska subenheterna → katalytiska subenheterna aktiveras
Förklara proteolytisk reglering
Vissa proteiner syntetiseras i inaktiva former så kallade: proproteiner/proenzymer = zymogener
Dessa aktiveras av proteaser när de behövs. Då klyvs zymogener och viks till en aktiv form
Klyvningen kan ske utanför cellen
Proteaser är ofta zymogener
Ge exempel på proteolytisk reglering
Chymotrypsinogen bildar chymotrypsin mha proteaset trypsin (inaktiv form trypsinogen)
Hormoner: insulin kommer från proinsulin
Apoptosis som utförs av kaspaser som formas av prokaspaser
Vad skiljer proteolytisk reglering från de andra regleringsstratergier
Proteolytisk aktivitet är inte reversibel
De aktiverade enzymerna kan bli inaktiva mha inhibitorer