Unnit 4 pt 2
1/33
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
34 Terms
the cytoskeleton mediates cell migration
allows protrusion of the cell membrane to be the leading edge which forms fillip odium and is pushed forward by actin polymerization
the lagging edge is pushed forward by contraction bundles
stress fibers
lagging end of the cell made up of alpha actinin and myosin which contract ot move the cell forward
lamellipodium
made up of arp 2/3 and is the edge of the cell that is pushing forward
filopodium
made up of tight parallel bundles and fibrin
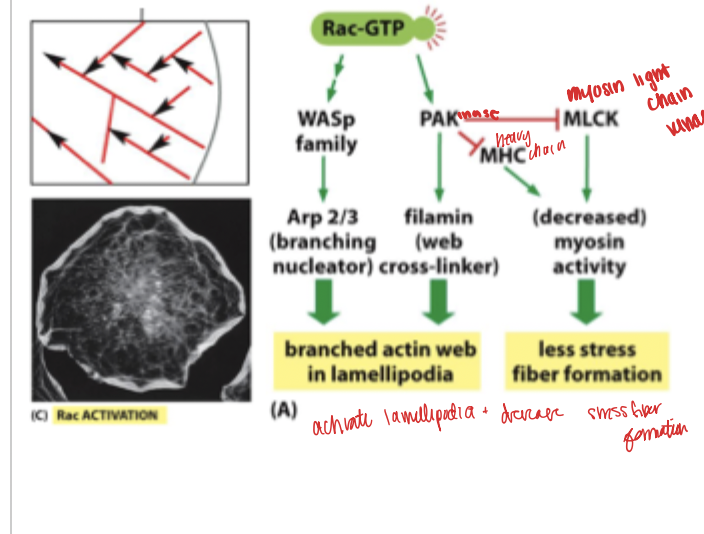
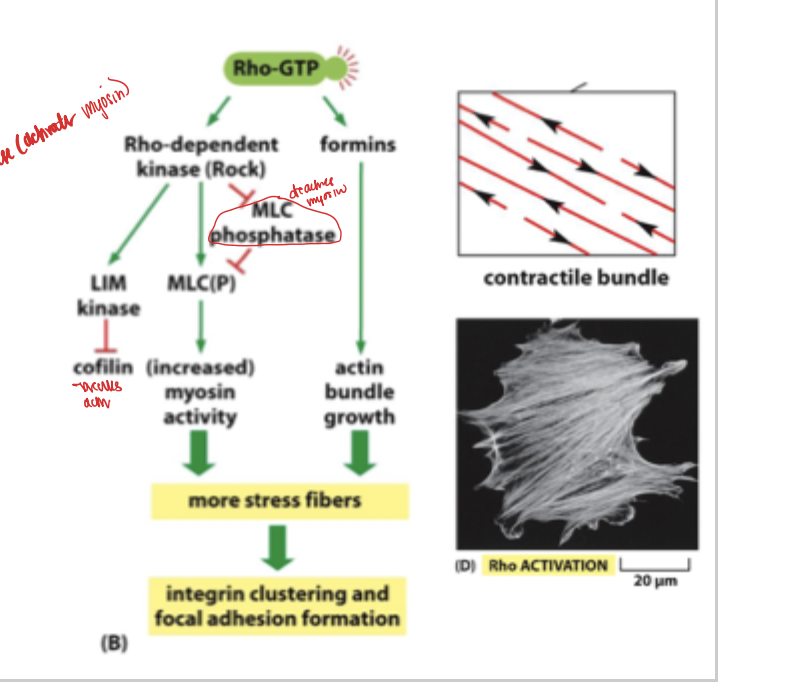
rac and rho and Cdc42
rho activation leads to the activation of contractile bundles- stress fibers
rac activation leads to the activation of dendritic networks and helps ot form lamellipodium from ARP 2/3 activation
Cdc42 activation helps form filopodium
rac pathway
rho pathway
rac and rho, how are we having both of these pathways?
likely active at the same time, want them at different parts.
rac dominates w signal presence and allows actin polymerization and profusion in the presence of a signal (want to activate lamelipodium and filopodium to go towards the signals)
rho dominates with no signal presence and is therefore more present in the absence of close extracellular signals
other ways that rho can be activated
involves contact mediated signals with a EphA4 (type of tyrosine kinase) which uses other protein to activate row on a specific side of a cell.
integrins and motility
integrins are used to anchor the cell the the extracellular matrix to allow the cell to move and stay stable and use actin to protrude forward.
integrin activation
signaling protein activates RAP 1 (in this case, thrombin activates the thrombin receptor which activates RAP1, acting as a GEF). Rap then recruits RIAM, talin and kindlin. uses the RAP1 to bind the talin to the integrin which can then bind to vinicuin to bind to actin and recruit more actin.
response time is crucial
slow response time is because it means that the SM has signaled for altered protein synthesis- either making more or less or a different protein but is the actual synthesis of a protein itself.
fast responses- signal for altered protein function and therefore can occur quickly because you do not have to resynthesizes more proteins
MAP kinase signaling two cascades
map kinase signaling can be attached to scaffold proteins, therefore putting map kinase kinase kinase and map kinase kinase in approximate areas so they can phosphorylate each other. doing this can lead to either a mating response or glycerol synthesis in yeast.
how to prevent both occurring at the same time?
scaffold one binds to each of the different kinases at the same time
scaffold two has a kinase domain as its map kinase kinase and binds to to other kinases for the other two kinases. when salty, scaffold 2 will bind to the high osmolarity sensing receptor
the different activation times for the scaffolding protiens to the GPCR allows for different responses to occur at different times. also, they use different types of MAP kinase so they wouldn’t ve able to have the whole process just to the map kinase kinase
notch signaling pathway
contact dependent signaling pathway typically used in developmental processes or when tissues are changing.
notch signaling
once cell will begin to express the ligand (delta) and when the cell begins ot express delta, it activates notch receptors on the neighboring cells. when activated notches, they are no longer able to express delta. this helps to get the ratio of neuron and skin cells correct when differentiating
notch signaling cascade
notch creation starts in the er, which moves into the Golgi apparatus. while in the golgi lumen.
inside the lumen of the golgi, the notch is cut by a golgi and then reattaches to itself through noncolvalent interactions
this large cut portion inside the golgi then interacts with a delta ligand inside of the golgi lumen which then gets endocytose to make a cell.
the rest of the notch signal that is not bound to delta and is within the extracellular space, proteases cleave the end part of the protein and then another protease cleaves the cytosolic portion which has a NLS and will travel inside of the nucleus and can turn on or off gene expression.
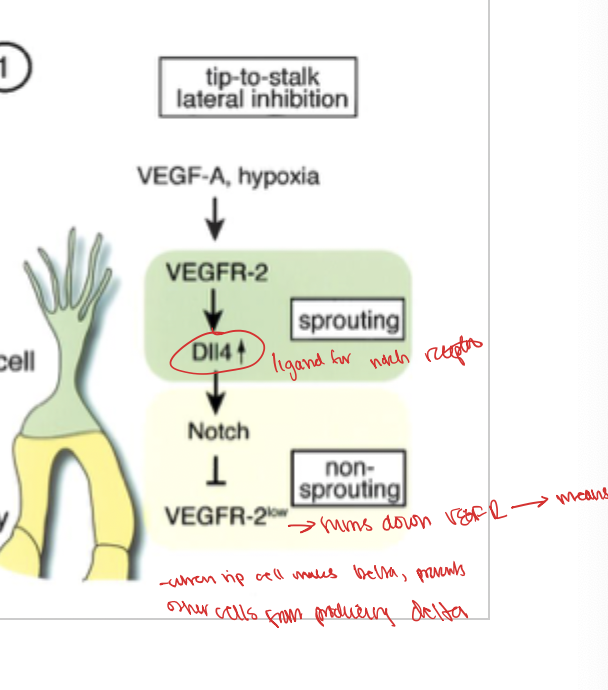
tip to stalk lateral inhibition
the cell that gets the delta ligand out to its cell surface. the presence of delta is then an inhibitor for the creation of the VEGF-2 delta which is why the rest of teh cells cant make it
many signaling pathways result in transcriptional changes in a cell- how do these proteins affect the cell
proteins can change gene expression by binding to an enhancer protein
enhancer protein- distance regulatory region that proteins bind to to either recruit other transcription proteins or block transcription proteins from binding
active versus inactive receptors- how do they switch from one to the other
has a ligand binding domain, DNA binding domain and transcription activating domain
must bind to ligand to get it the right conformation will the inhibitory proteins absent. then the, receptor must bind to the correct portion of the DNA and bind to a coactivator in the proper conformation to successfully activate or deactivate transcription of target gene
Eukaryotic genes have complex regulatory systems
gene control region for a specific gene includes its general transcription factors, coactivators, spacer DNA, transcription regulators, regular sequences, promotor, polymerases and the gene in question
transcription regulators act at _____ sequences
cis-regulatory; they have specific sequences they bind to- short sequences (6 bp)
eukaryotic genes have complex regulatory systems
they are dependent on activators and repressors present in nucleus for if a certain protein will activate or repress the transcription of a particiular gene
one protein can turn a gene off or on but it depends on the other proteins present in the nucleus what it will do
active hedgehog
when active, hedgehog signaling molecule is present which causes patched to be in the cell membrane, this allows smoothened to be present in the cilliary membrane. since smoothened is in the colliery membrane, it allows the Gli2-SuFu sequence to break apart and allows Gli2 to go into the nucelus and turn the hedgehog target genes on
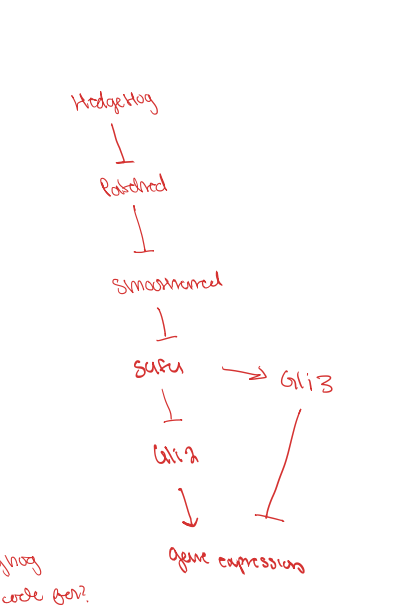
inactive hedgehog
because hedgehog is not bound, smoothened is found in the cell membrane and patched is found in the ciliary membrane. This causes the Gli2 -SuFu complex to not dissociate and instead it turns into SuFu-Gli3 and something else. This then gets proteolytically processed so fufu dissociates and the Gli3 is able to travel into the nucleus to turn the hedgehog target genes off
cilia and flagella have pressure gated channels
calcium channels- these can activate calcium waves and camp-kinase
primary hubs of cilia and flagella are important for what
signaling- defects of cilia and flagella are often because of messed up signaling pathways for primary cilia
beta-catenin contact vs signaling
beta-catenin is related to signaling through its gene regulation but is also a connecting protein within the plasma membrane of cells
beta-catenin is an adaptor protein that forms adherins
it helps to regulate gene expression
more cadherins and surrouding cells, however, means less protein production because it means there is not as much beta cateinin to recruit ot the nucleus
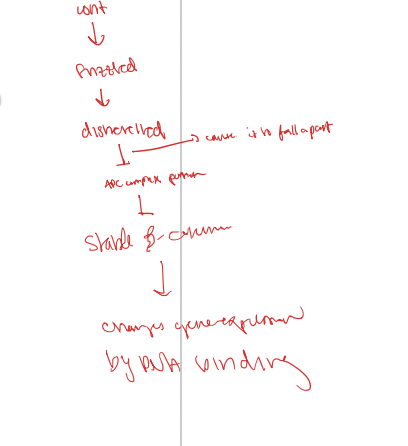
WNT signaling- without
without wnt signal- if too many adherents junctions, no beta catenin will go into the nucelus because there is none
without wnt, there is an apc complex which binds a beta-cateinin to the complex, which includes an axis which holds an active GSK3 and actin CK1 which phosphorylate beta cateinin and make it unusable and degrade
this makes it so the beta catenin is unable ot bind to the LEF1/TCF on DNA so it is unable to turn the target genes on
when wnt is bound
wnt binds to frizzled and activates it (also binds to an LRP) this complex activates a protein called disheveled which then breaks apart the APC bound complex by attracting axin. the kinases phosphouyrlate the active LRP
the stable beta catenin is then able to bind and kicks the Groucho protein off to turn on transcipriton of the target genes