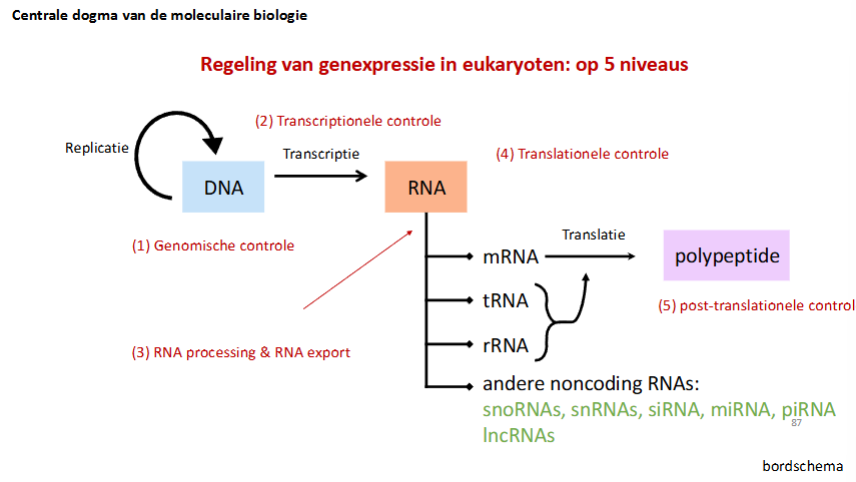
20.4: Regeling van genexpressie in eukaryoten: post-transcriptionele controle (3de - 4de - 5de niveau)
1/101
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
102 Terms
slide niveaus van controle
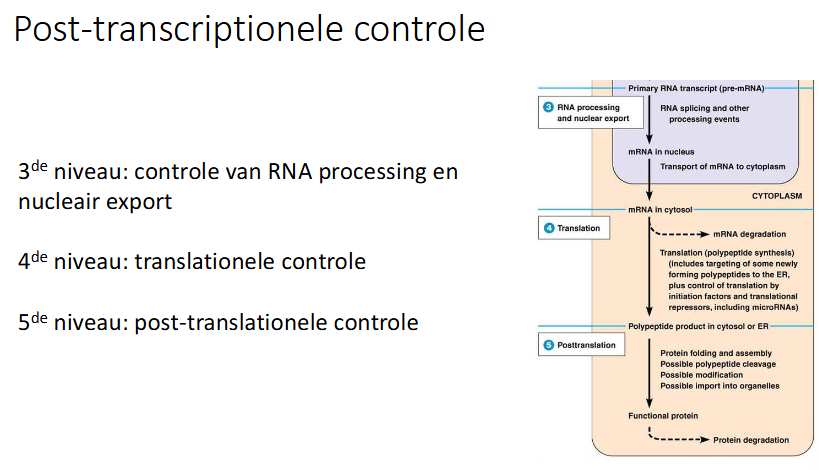
slide schema
wat is 3e niveau van regeling van de genexpressie bij eukaryoten
regeling van RNA processing en nucleaire export
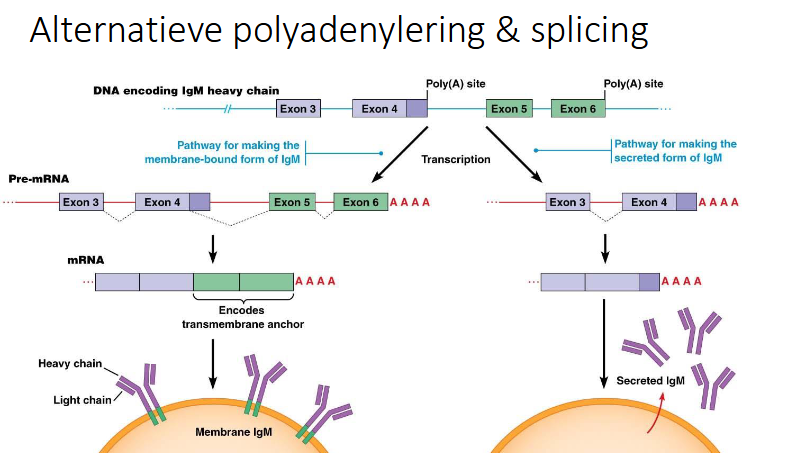
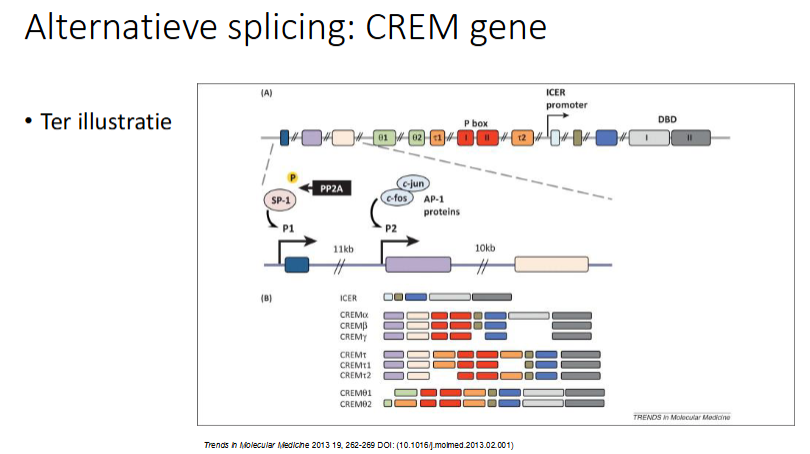
wat gebeurt er bij alternatieve splicing en polyadenylering
Regulatorische proteïnen en snoRNAs binden op splicing enhancer en silencer sequenties van het pre-mRNA (kunnen zoxel in exon als intron gelegen zijn)

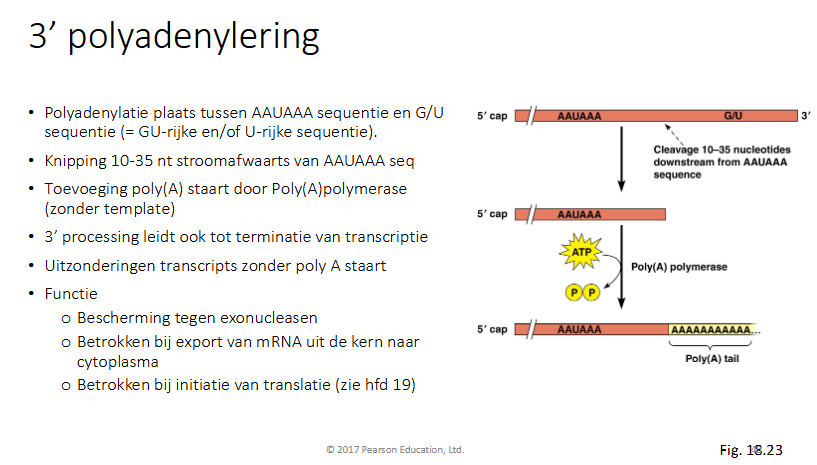
Hoe gebeurt 3’ polyadenylering
Polyadenylatieplaats tussen AAUAAA seq en G/U seq (: GU rijke en of U-rijke seq)
Knipping 10-35nt stroomafwaarts van AAUAAA seq
toevoeging van polyAstaart door polyApolymerase (zonder template)
3’ processing leidt ook tot terminatie van transcriptie
uitzondering transcripts zonder polyAstaart
functie
bescherming tegen exonucleasen
betrokken bij export van mRNA uit de kern naar cytoplsma
betrokken bij initiatie van translatie
slide alternatieve polyadenylering en splicing
slide ter illustratie
wat gebeurt er met mRNAs die geen EJC gebonden hebben
worden afgebroken in de kern (zoals alle defecte mRNAs)
welke soort mRNAs worden wel geëxporteerd uit de kern en gebeurt dit altijd spontaan
Enkel mature mRNAs (met EJC gebonden) worden geëxporteerd als mRNP doorheen de kernporieën (Ran onafhankelijk)
Soms stimulus-gergelde export
bv HUV viru synthtiseerd Rev proteine dat een NES (nucleair export signaal) hheeft
het Rev bindt HIV in de kern en brengt dit RNA naar het cytoplasma door de nucleaire poriën zodat het afgeschreven kan worden

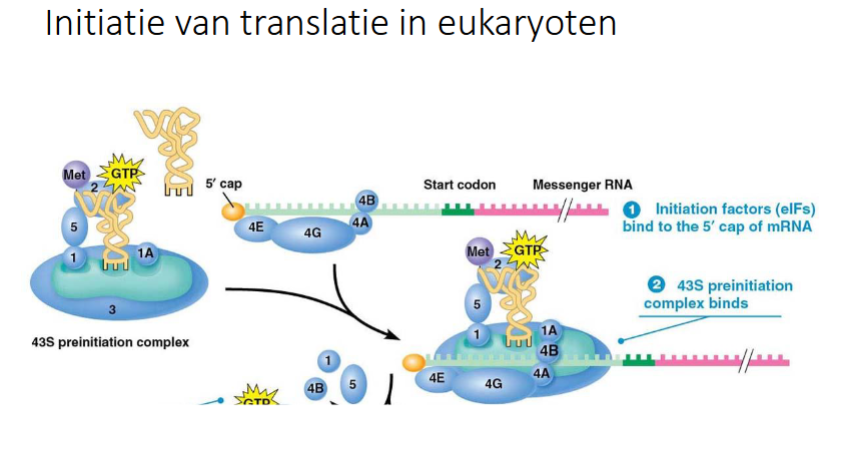
slide initiatie translatie bij eukaryoten
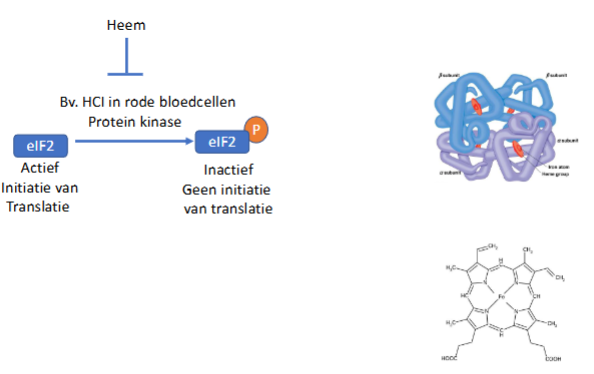
slide inhibite eIF2
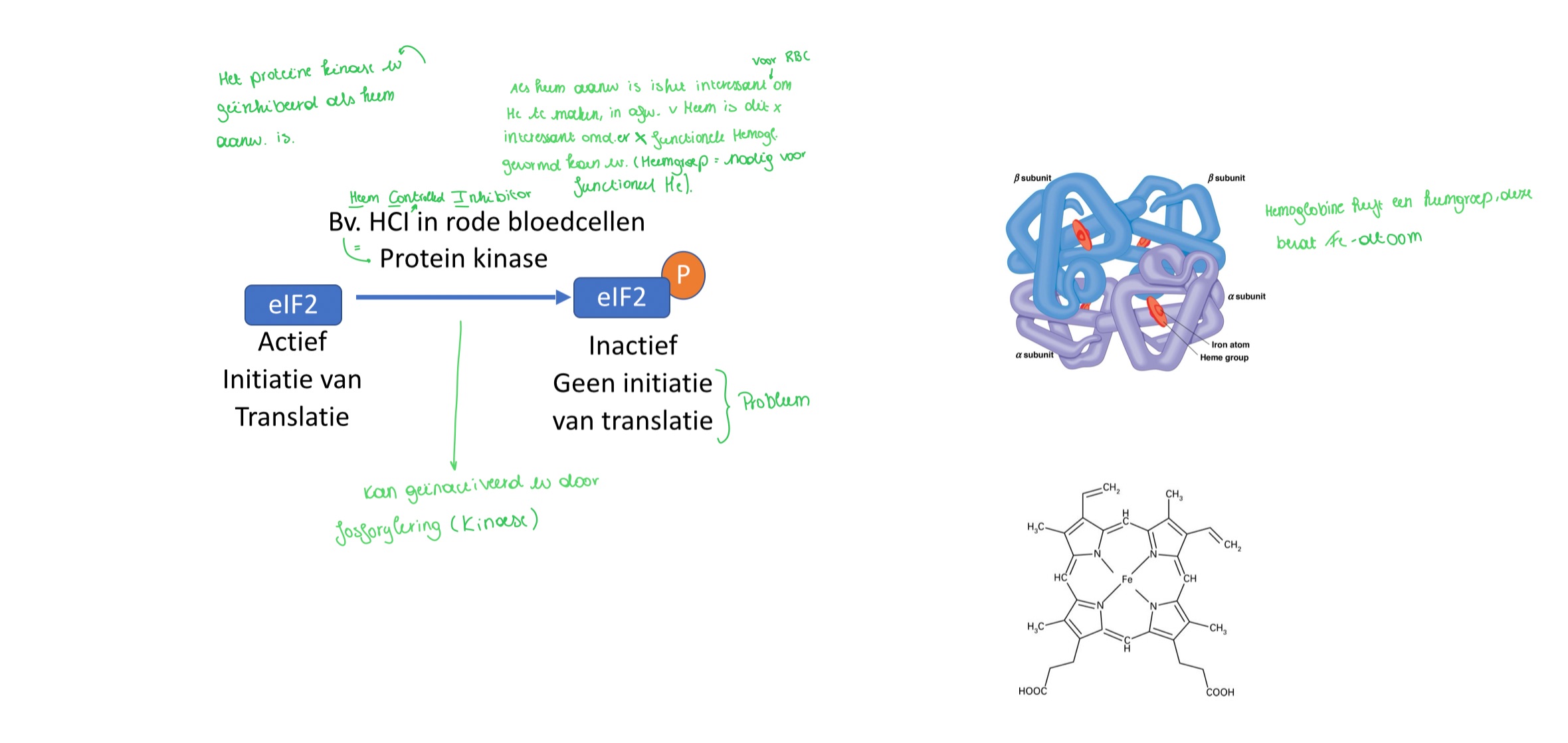
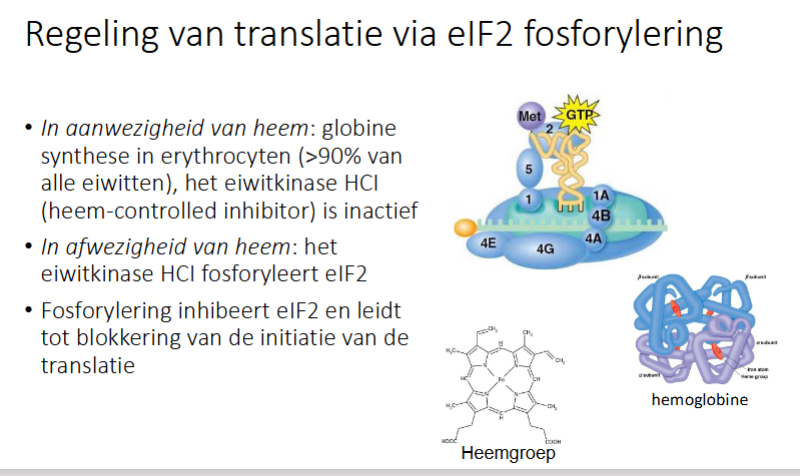
hoe gebeurt de regeling an translatie via eIF2 fosforylering?
in aanwezigheid van heem: globine synthese in erythrocyten (>90% van alle eiwitten), het eiwitkinase HCI (heem controlled inhibitor) is inactief → translatie kan optreden
In afwezigheid van heem: het eiwitkinase HCI fosforyleert eIF2 → inhibeert eIF2 en leidt tot blokkering van initiatie van translatie
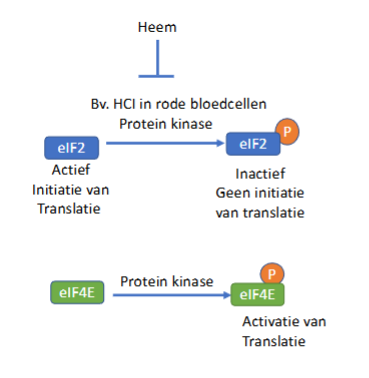
wordt eIF4E ook inactief door fosforylering
nee, omgekeerde van eIF2
eIF4E wordt net geactiveerd door de fosforylering
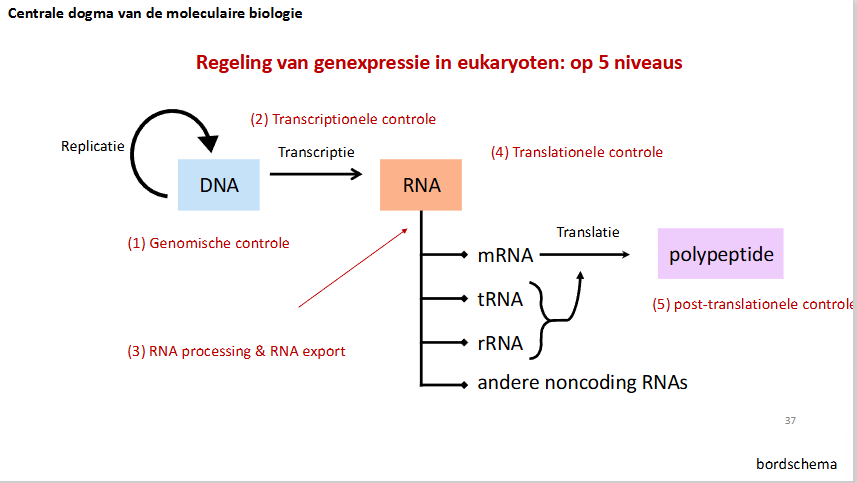
slide bordschema
slide initiatie translatie eukaryoten
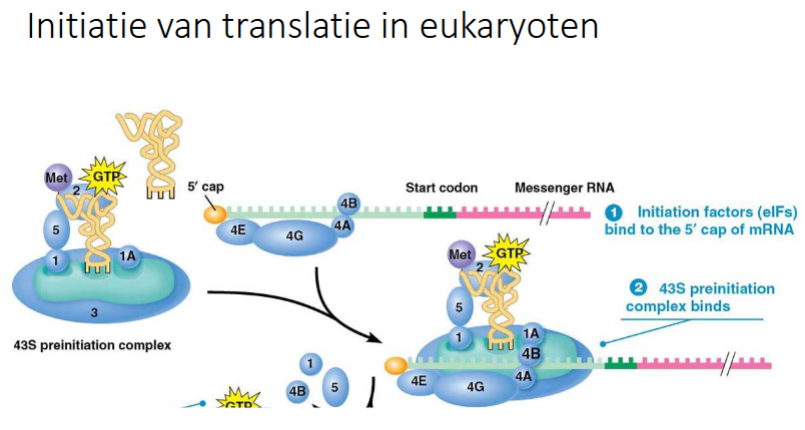
wat is eIF4F
complex van eIF4E, eIF4G, eIF4A
waar bindt eIF4E
aan de 5’ cap van mRNA
is eIF4E actief of inactief bij fosforylering
actief
hoe en waarom blokkeren sommige virussen de fosforylering van eIF4E
Sommige virussen produceren een protease dat eIF4G proteolyseert in 2 stukken (hierdoor geen translatie van cellulaire mRNAs en wel van virale mRNAs)
1 fragment bindt aan 5’ cap via binding op eIF4E
ander fragment vindt op IRES (international ribosome entry sequence) van viraal mRNA, zodat er wel translatie is van virale mRNA en cellulaire

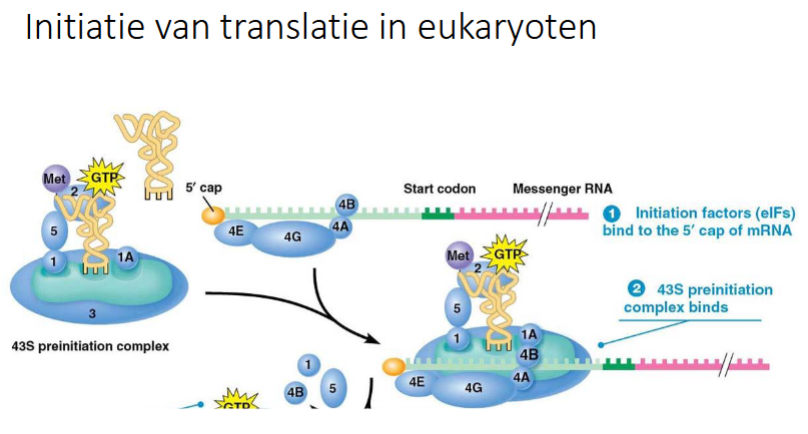
slide initiatie van translatie
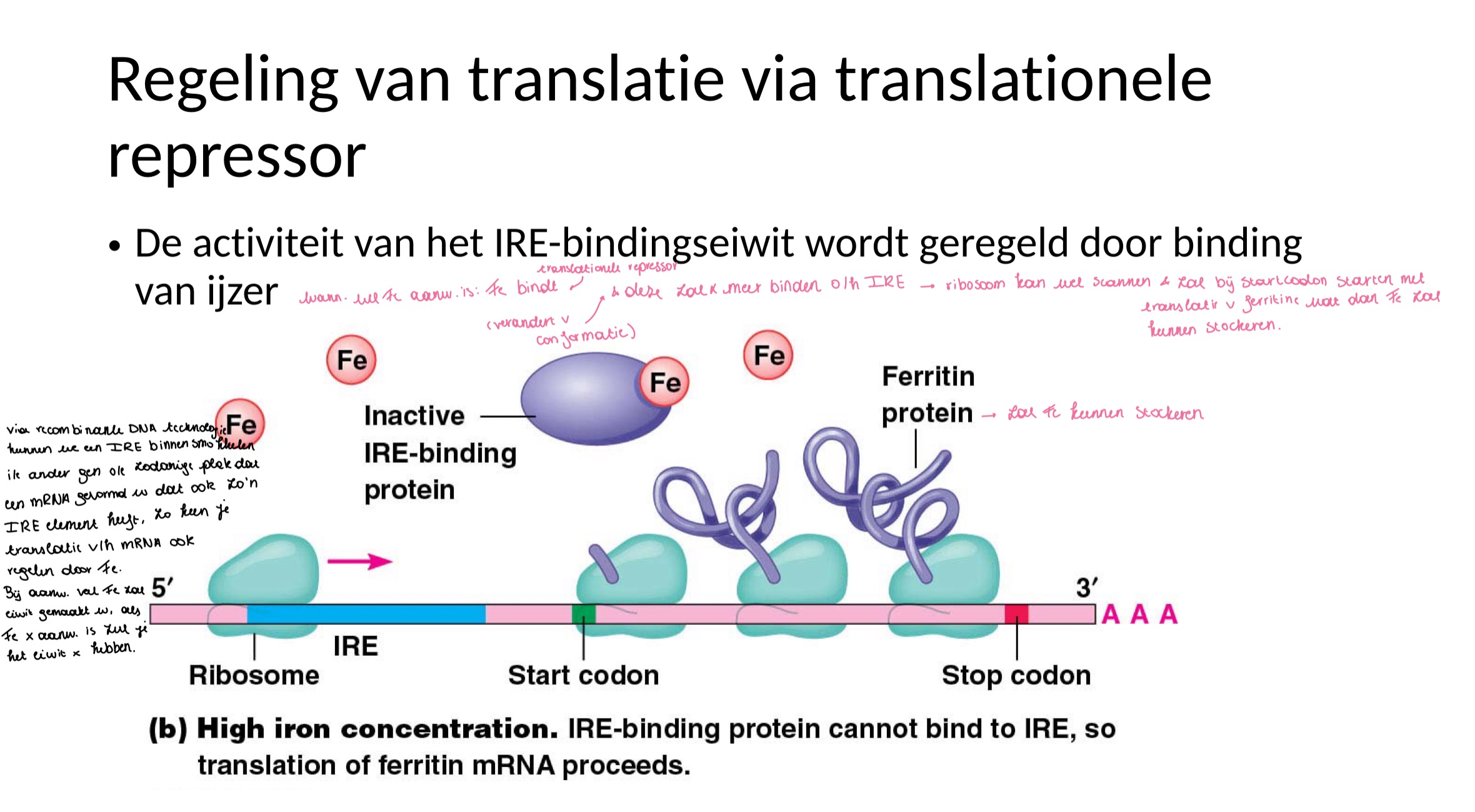
hoe gebeurt regeling van translatie via translationele repressor
ferritine = intracellulaire stockage van Fe
het zal dus gesynthetiseerd moeten worden wanneer er veel Fe aanwezig is
veel ferritine nodig bij hoge ijzerconcentratie → synthese selectief gestimuleerd door Fe
IRE = ijzer-respons element in 5’ UTR
IRE-bindingseiwit = translationele repressor en allosterisch eiwit (heeft dus 2 conformaties en kan een effectormolecule binden, nl Fe kan zorgen voor verandering van de 3D structuur)
Bij lage concentratie aan ijzer bindt de translationele repressor (IRE bindins eiwit) zodat er geen translatie is van ferritine mRNA.
want 43S pre-initiatie complex van het ribosoom wordt eerst gevormd aan de cap → dan scant het (beweegt over het mRNA tot het AUG tegenkomt)
normaal zou hier initiatie gebeuren, dat zal hier niet gebeuren omdat de scanning wordt geblokkeerd door translationele repressor).

functie TRANSLATIONELE repressor
controleert translatie van een specifiek mRNA door binding op een respons element in mRNA
slide Fe aanwezig
met welke experimenten kun je de halfwaardetijd van mRNA nagaan
Pulse-chase experimenten (radioactief nucleotiden toevoegen en kijken wanneer ze afgebroken zijn)
wat bepaalt de levensduur van een mRNA
lengte polyAstaart
aantal AU-rijke elementen in 3’ UTR (hoe meer, hoe minder stabiel)

Hoe gebeurt de rgeling van mRNA afbraak via ijzer
Dit gebeurt bv bij mRNA voor transferrine (een transmembranair eiwit dat verantwoordelijk is voor transport van Fe naar de cel, nodig wanneer [Fe] laag is).
IRE bindend eiwit (translationele repressor) bindt in IRE in 3’ UTR en zorgt er zo voor dat het mRNA niet wordt afgebroken wanneer [Fe] laag is
Bij een hoge Fe concentratie moet transferrine niet aangemaakt worden → Fe bindt op traslationele repressor en deze zal niet meer op het IRE binden → IRE is vrij en kan worden afgebroken
![<p>Dit gebeurt bv bij mRNA voor transferrine (een transmembranair eiwit dat verantwoordelijk is voor transport van Fe naar de cel, nodig wanneer [Fe] laag is).</p><p>IRE bindend eiwit (translationele repressor) bindt in IRE in 3’ UTR en zorgt er zo voor dat het mRNA niet wordt afgebroken wanneer [Fe] laag is</p><p>Bij een hoge Fe concentratie moet transferrine niet aangemaakt worden → Fe bindt op traslationele repressor en deze zal niet meer op het IRE binden → IRE is vrij en kan worden afgebroken</p><p></p>](https://knowt-user-attachments.s3.amazonaws.com/85b36764-a398-47dc-8c6c-aafa3ddab2b0.png)
slide afbraak mRNA

hoe gebeurt mRNA afbraak via exosoom of P-bodies
3’ → 5’ pathway: uitgevoerd door cytoplasmatisch exosoom
inkorten polyAstaart (3’ uiteinde)
verwijderen van polyAbindende proteïnen
degradatie van mRNA in de 3’ → 5’ richting door exonucleasen van cytoplasmatisch exosoom
afbraak 5’ cap door Cap-afbrekende enzymen
5’ → 3’ pathway
inkorten polyAstaart
verwijderen 5’ cap
afbraak mRNA in 5’ → 3’ richting door exonucleasen gelocaliseerd in een cytoplasmatische structuur gekend als P-bodies (mRNA processing bodies)

slide bordschema
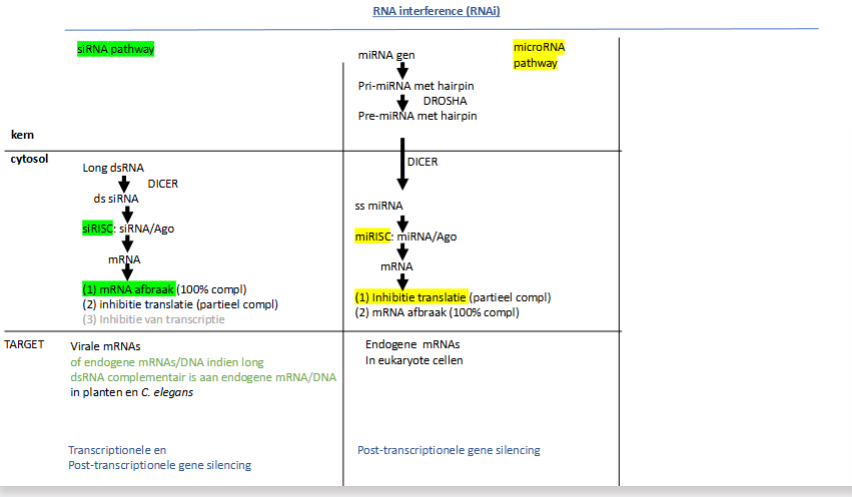
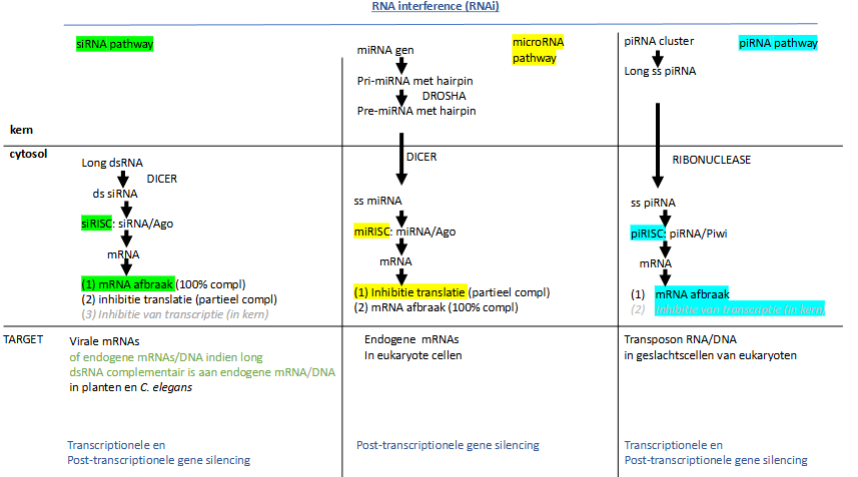
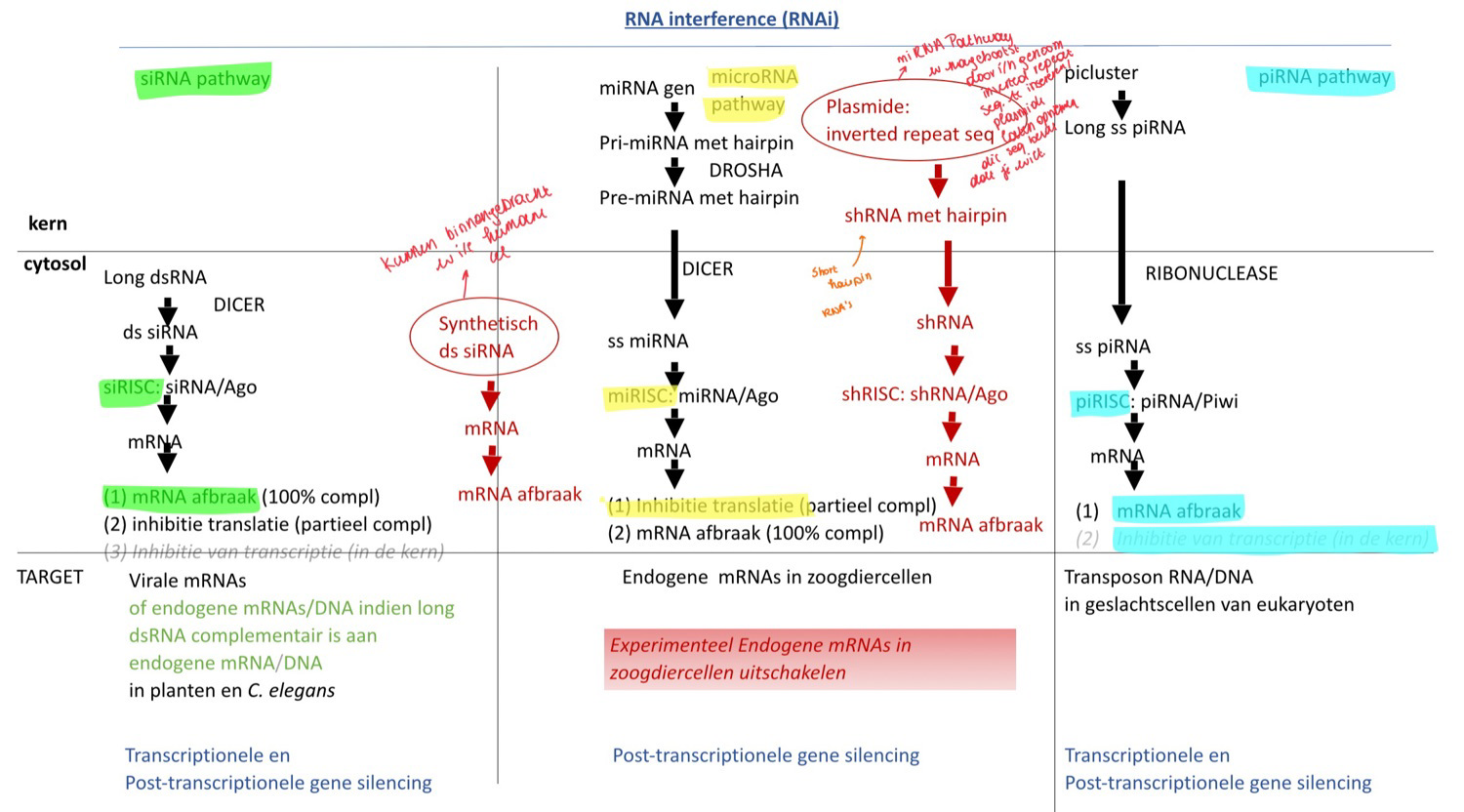
wat is RNA interference
(RNAi)
RNA-gemedieerde inhibitie van genexpressie
slide kleine RNA’s

waardoor wordt RNAi respons vesterkt
RNA afhankelijk RNA pol
waarvoor staat siRNA
small interfering RNA
waarvoor staat siRISC
RNA induced silencing complex
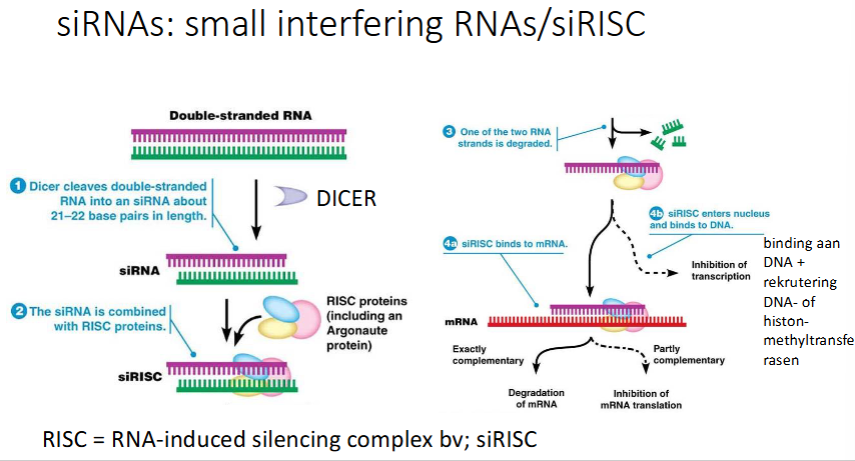
Hoe werken siRNAs
Plantencel / C;elegans wordt geïnfecteerd met een virus, het injecteert zijn genoom (dsRNA)
In het cytoplasma zit een ribonuclease dat dsRNA in stukken zal knippen. Zo krijg je siRNAs (= kleine stukken dsRNA) → DICER
RISC eiwitten herkennen siRNA’s
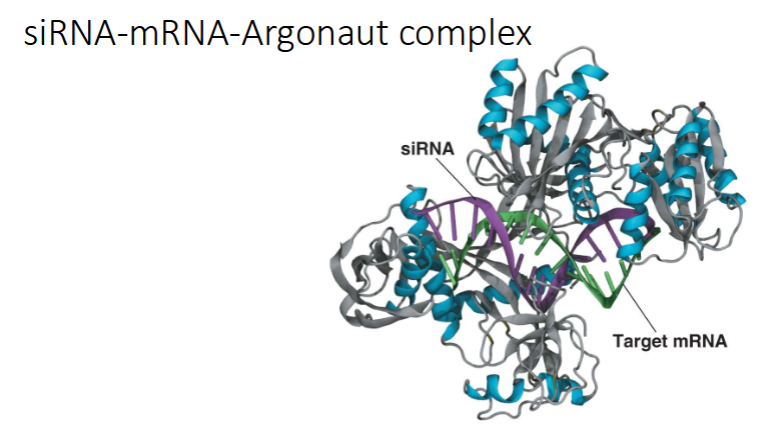
siRISC complex wordt gevormd → 1 van de eiwitten is argonaut (ook een ribonuclease → breekt RNA af)
1 van beide strengen wordt afgebroken en we krijgen ss siRNA met eiwitten (waaronder argonaut)
2 opties:
Hoofdweg:
complementaire siRISC bindt op viraal mRNA
Als het 100% complementair is, zal het mRNA worden afgebroken.
Als het niet 100% complementair is, zal de translatie geïnhibeerd worden, maar blijft het mRNA
Niet-hoofdweg:
siRISC gaat naar nucleus → binding aan DNA → rekrutering DNA- of histon methyltransferasen
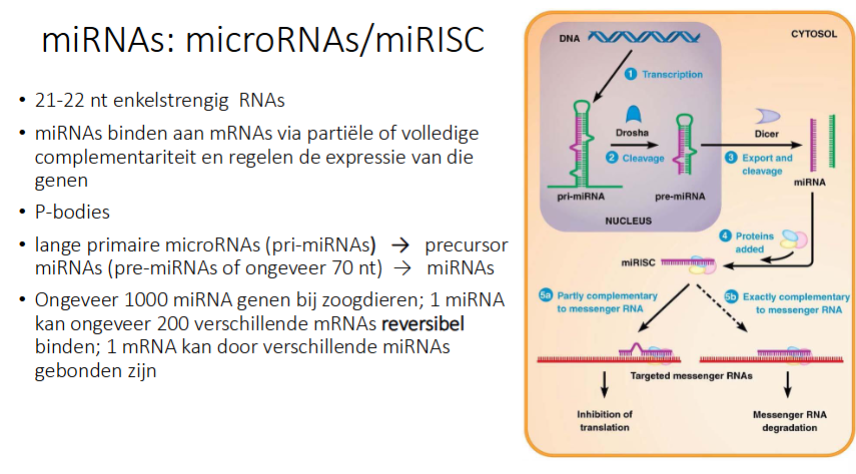
waarvoor staat miRNA
micro RNA
codeert miRNA voor een eiwit
nee, er zijn miRNA coderende genen in het lichaam
hoe werke miRNAs
Transcriptie → pri-miRNA
Drosha (= nucleair ribonuclease) knipt pri-miRNA en je krijgt pre-miRNA
pre-miRNA gaat naar cytosol
Dicer in cytosol knipt pre-miRNA in kleine ss miRNA
1 van de 2 strengen wordt afgebroken, de andere wordt bezet met eiwitten
2 opties
Hoofdweg:
het miRNA hybridiseert op mRNA obv partiële complementariteit (dus niet 100%), dus 1 miRNA kan op verschillende mRNAs binden
We krijgen inhibitie van translatie, maar mRNA blijft
het mRNA:miRNA complexen worden gestockeerd in P-bodies
Als de cel het dan nodig heeft, kan het uit de P-bodies komen → miRNA afbreken → kan weer translatie gebeuren
Niet hoofdweg:
miRNA is 100% complementair aan mRNA → mRNA wordt afgebroken
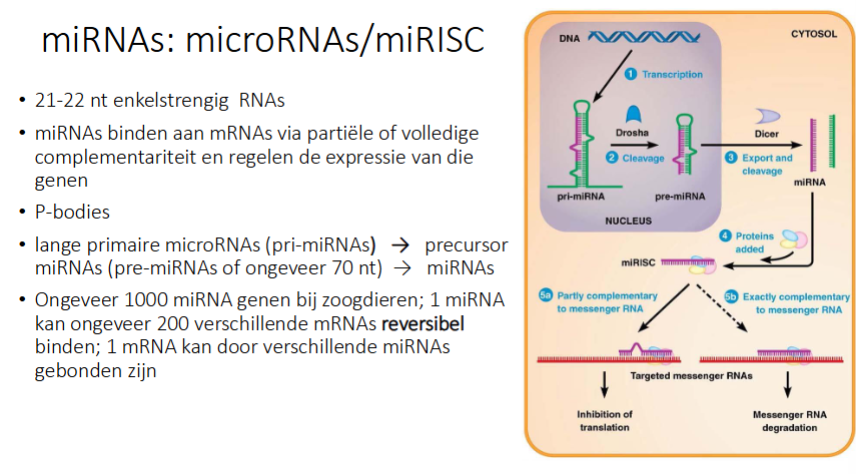
hoeveel mRNAs kan een miRNA binden, is dit reversibel of niet
1 miRNA kan ongeveer 200 verschillende mRNAs reversibel binden
kan 1 mRNA door verschillende miRNAs gebonden zijn
ja
waarvoor staat piRNA
Piwi-interacting RNAs
waarvan zijn piRNAs afkomtig
van lange ssRNA
Hoe worden piRNAs afgeschreven
piRNAs worden afgeschreven van een piRNA cluster (is genomische en zit in elke cel, maar komt niet in elke cel tot expressie) die bestaat uit stukjes van transposon DNA
wat is de piwi familie
piwi familie (behoren tot de argonaut proteïnen en hebben dus ribonucleaseactiviteit) binden de piRNAs in een RISC complex (piRISC)
waar binden piRNAs op en wat is het gevolg
wat doen ze nog
RNA dat afgeschreven is van een transposon met RNA afbraak tot gevolg
op transposons DNA en inhibeert de transcriptie (moet dus naar kern) (2e niveau van regulatie)
→ beschermen ook tegen transposon activiteit
waar komen piRNAs hoofdzakelijk tot expressie en waarom
in geslachtscellen → moeten beschermd worden tegen transposon activiteit

slide siRNA

slide miRNA
slide piRNA
ribonuclease is NIET dicer
slide siRNA-mRNA-Argonaut complex
RNAi: hoe kun je ds RNA binnenbrengen
Via injectie: blijkt wel een RNAi respons te geven
via ds RNA virussen: geen RNAi → werkt enkel bij planten en C-elegans en dus niet bij de mens
via een plasmide dat een inverted repeat tot expressie brengt dat een haarspeld structuur zal aannemen (want inverted repeat geeft aanleiding tot interne complementariteit)
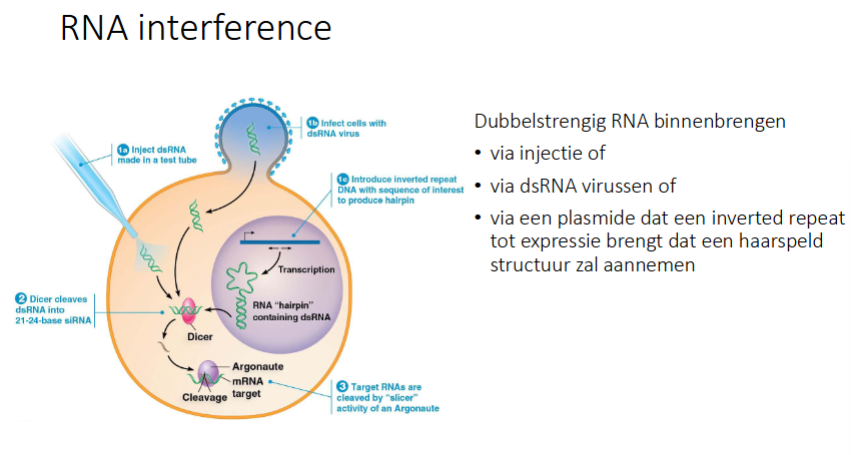
als we geen siRNAs hebben, hoe kunnen we dan onzelf beschermen tegen virale infecties
interferon respons element
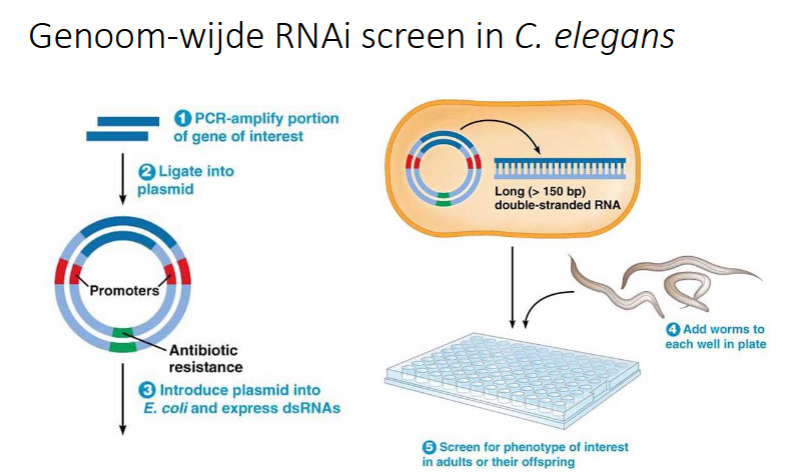
hoe doe je een genoom-wijde RNAi screen in C. Elegans
voor elk gen van C. Elegans wordt een bacterie genetische gemodificeerd
met PCR gen van ons interesse amplificeren
gen insereren in een plasmide
Plasmide in E. Coli doen
Plaat met welletjes: in elk welletje zijn bacteriën met een plasmide (elk stukje een andere plamide corresponderen dmet het gen)
in elke welletje worden C. Elegans gestoken
de wormen zullen de bacteriën opeten en zo komt het dsRNA vrij.
het gen wordt uigeschakeld in alle cellen
sommige wormen zullen afsterven en dan weet men welke de essentiële genen zijn bij uitschakeling → lethaal
bordschema
slide schema
op welke manier is therapeutisch gebruik van RNAi mogelijk
bv als een gen te actief is in kanker een miRNA toevoegen om het uitschakelen
miRNA coderende genen bevatten soms mutaties, als een miRNA niet functioneel is in de cel kan het tot een fenotype leiden afhankelijk van hoe drastisch de functie beïnvloed wordt.
Door 1 miRNA te muteren verander je de translatie van meerdere genen.

Een onderzoeker voegt een siRNA toe aan een humane cel om de genexpressie van gen A uit te schakelen.
Het siRNA zal een sequentie hebben dat identiek is aan de
a) intron van gen A
b) exon van gen A
c) promotor van gen A
d) enhancer van gen A
b
mRNA heeft geen promotor
waarvoor staat lncRNA
long noncoding RNA
coderen lncRNA voor een eiwit
nee
wat brengen lncRNA tot expressie
RNA
waarmee overlappen LncRNA genen
40% overlap met proteïne coderende genen
heeft een LncRNA een promotor
ja
doet LncRNA trans of cis inhibitie
cis
wat is cis inhibite door LncRNA
inhibitie van transcriptie van dichtbij gelegen genen en / of chromosomen
vb van cis inhibitie door lncRNA
Xist RNA
een RNA pol zal het eiwitcoderend gen afschrijven, een RNA pol zal ook het LncRNA afschrijven.
De RNA pol kunnen botsen tegen elkaar, wat kan het effect hiervan zijn
beïnvloedt transcriptie
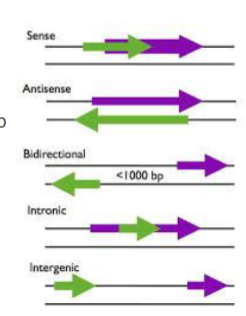
wat is een sense lncRNA
een lncRNA dat overlapt met een eiwitcoderend gen in dezelfde richting
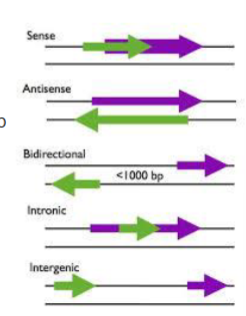
wat is een antisense lncRNA
een lncRNA dat overlapt met een eiwitcoderend RNA in de andere richting
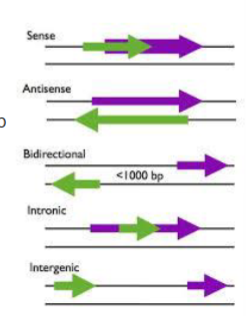
wat is een bidirectionele lncRNA
een lncRNA met een bidirectionele promotor (verschillende richtingen) op minder dan 1000 bp van eiwitcoderend gen
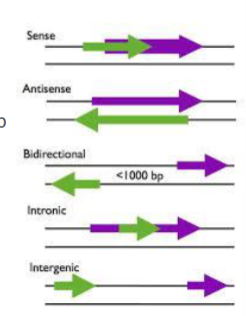
wat is een intronisch lncRNA
een lncRNA dat ligt in een intron van een eiwitcoderend gen
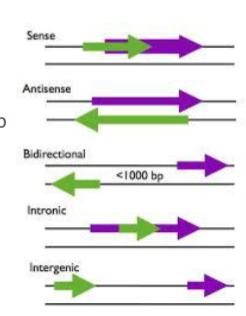
wat is een intergenisch lncRNA
een lncRNA dat ligt tussen 2 eiwitcoderende genen
wat ka lncRNA nog doen naast transcriptie inhiberen van dichtbij gelegen genen en / of heel chromosoom
lncRNA kan associëren met het chromatine modificerende/regulerende eiwitten en deze ribonucleocomplexen targeten naar een andere locus
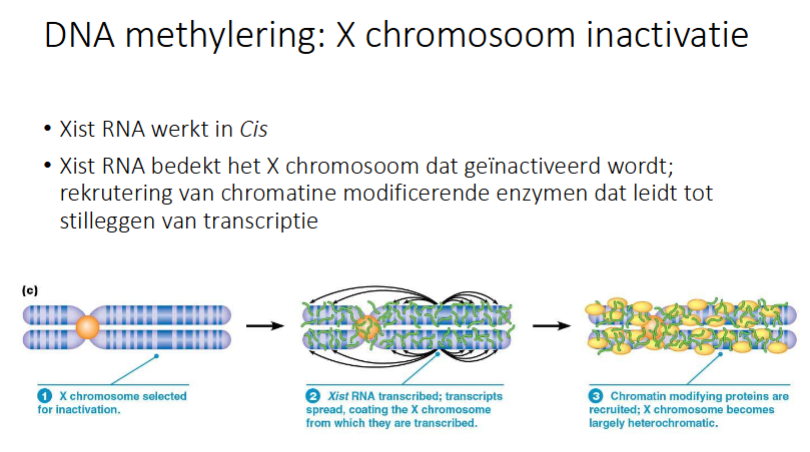
Hoe werkt Xist RNA
Het werkt in Cis
het X-chromosoom dat geïnactiveers zal worden zal transcriptie doen van Xist RNA gen doen waardoor Xist RNA tot expressie komt
Xist RNA verspreid zich over het gehele X-chromosoom dat geïnactiveerd wordt
chromatine modificerende enzymen worden gerekruterd en dat leidt tot stilleggen transcriptie
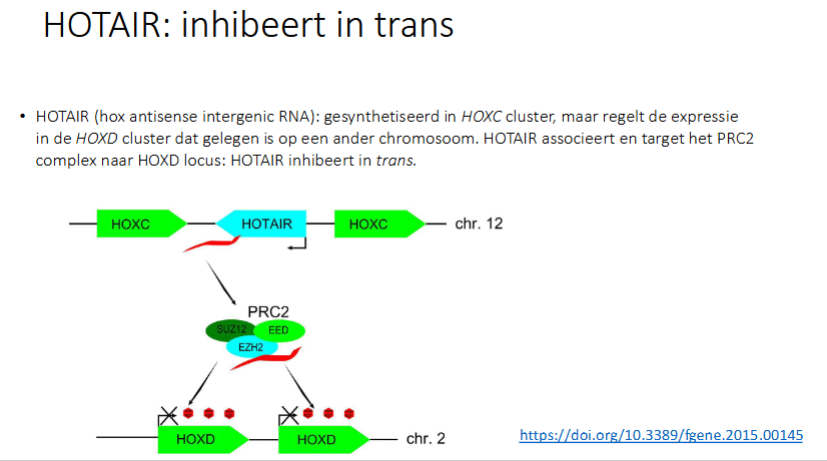
Xist RNA is een vb van hoe lncRNA genen die in de buurt liggen / chromosomen in cis kunnen inhiberen, wat is een vb van hoe lncRNA ribonucleocomplexen kan targeten naar een andere locus
HOTAIR
inhibeert HOTAIR in cis of trans
trans
waarvoor staat HOTAIR
Hox antisense intergenic RNA
wat doet HOTAIR
het inhibeert in trans: ht inhibeert een gen op een ander chromosoom
het is gesynthetiseerd in de HOXC cluster, maar regelt de expressie in de HOXD cluster dat gelegen in op een ander chrosmosoom.
HOTAIR associeert en target het PRC2 naar HOXD locus
slide bordschema
wat zijn omkeerbare post-translationele modificaties
(de)fosforylering
(de)deacetylering
(de)methylering
voorbeeld van permanente modificatie
proteolytische knipping (bv processing insuline)
welke eiwitten regelen eiwitvouwing
moleculaire chaperones
wat is het doel van eiwittargeting
eiwitten naar intra- en intercellulaire locaties
vb van eiwitten naar intercellulaire locaties
secretie buiten de cel
welke molculen kunnen eiwitactiviteit reguleren
regulatorische moleculen of ionen bv cAMP Ca2+
wat bepaalt de concentratie van een eiwit in een cel
snelheid van synthese en afbraak
wat is de formule voor eiwitconcentratie
P = ksyn/kdeg
snelheid eiwitsynthese / snelheid van eiwitafbraak
waartussen kan de halfwaardetijd van eiwitten variëren
minuten tot weken
wat gebeurt er als synthese en afbraak van eiwitten in balans zijn
eiwitconcentratie blijft stabiel

Hoe gebeurt eiwitdegradatie door proteasomen
Ubiquitine wordt aan E1 (ubiquitine activerend enzyme) gebonden
ubiquitine wordt doorgegeven van E1 naar E2 (ubiquitine-conjugerend enzym).
Ook E3 (ubiquitine ligase) bindt eraan, het markeert eiwitten die afgebroken moeten worden en zorgt dat ubiquitine op E2 getransereerd wordt naar Lys in het target eiwit dat moet afgbroken worden (dit is poly-ubiquitine → meerdere ubiquitines komen op een eiwit, meerder Lys kunnen ge-ubiquitineerd worden en op 1 Lys kunnen meerdere ubiquitines zitten.
Wanneer de ubiquitines gebonden zijn op de Lys van het af te breken eiwit, zullen E2 en E3 dissociëren
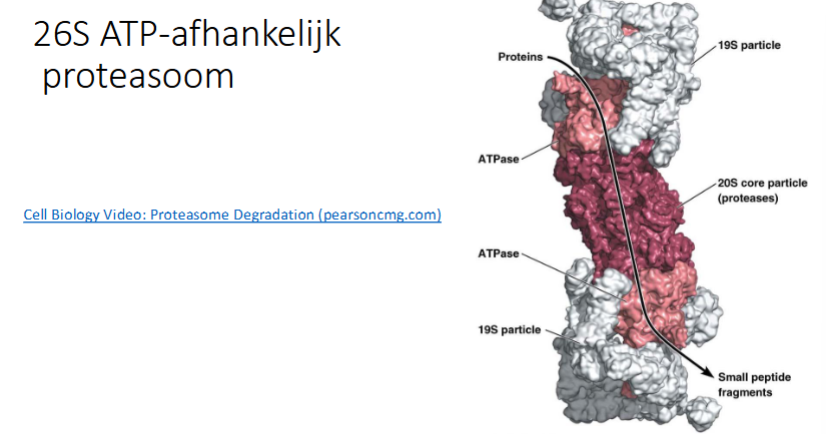
Het ge-ubiquitineerde eiwit wordt herkend door proteasoom
het proteasoom hydrolyseert de eiwitten tot kleine peptiden
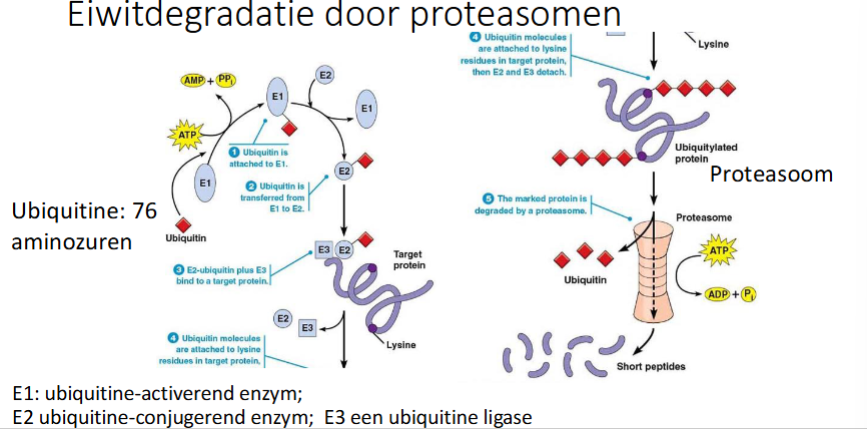
wat is ubiquitine
een klein peptide van 76 AZ’en
heeft een proteasoom ATP nodig
ja
slide Angelman syndroom en Prader-Willi syndroom

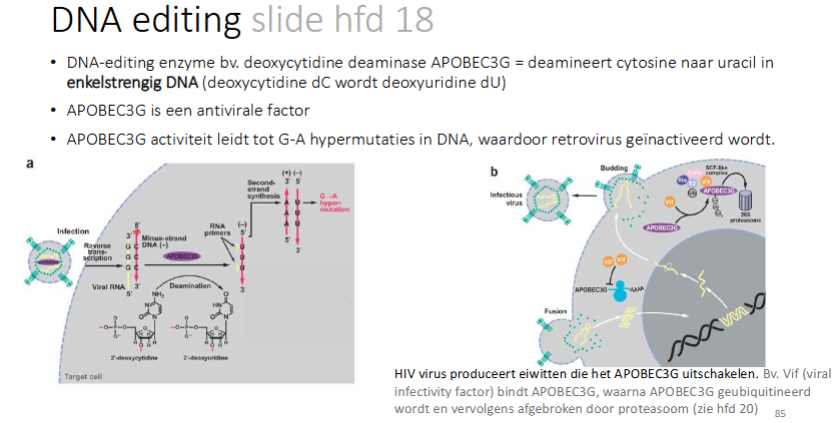
Hoe werkt APOBEC3G en hoe wordt het geïnhibeerd
DNA-editing enzyme bv. deoxycytisine deaminase APUBEC3G → deamineert cytosine naar uracil in ss DNA
APOBEC3G is een antivirale factor
APOBEC3G activatie leidt tot G-A hypermutaties in DNA, waardoor retrovirus geïnactiveerd wordt
Het HIV virus produceert eiwitten die het APOBEC3G uitschakelen (bv Vif (viral infectivitey factor) bindt op APOBEC3G waarna het ge-ubiquitineerd wordt en vervolgens afgebroken door proteasomen).
op welke manier kunnen we aan eiwitafbraak doen
ubiquitinering gergelde eiwitafbraak (ook defecte eiwitten worden geUb en dan afgebroken door proteasoom)
Sumoylering (Small ubiquitin-related modifiers, SUMOs, peptiden) regelt eiwitstabiliteit, nucleoplasmtisch transport en functie van transcriptiefactoren.
Eiwitafbraak via lysosomen (microautofagie); niet altijd selectief

welk ligase bepaalt welk residu in welk eiwirt wordt geUb (substraat specificiteit)
E3 ubiquitine ligase
E3 ubiquitine ligasen herkennen bepaalde sequenties, welke
N-terminale AZ’en
Interne AZ seq, gekend als DEGRONS
slide overzicht bordschema
slide ter illustratie

slide ter illustratie
