Paleo 419 Midterm 2
1/272
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
273 Terms
When did placentals dominate terrestrial habitats?
Mesozoic
When did eutherians first appear?
Early Cretaceous (Yixian Formation)
Eutherian Synapomorphies
canines with 2 roots
3 molars
lack epipubic bones (except some basal forms)
have malleolus on distal end of tibia
Eutherian skull characters
squamosal expanded to form part of the braincase – contacts alisphenoid
squamosal pierced by blood foramina
two parts of Trigeminal nerve, CN V2 (maxillary) and
CN V3 (mandibular), exit through the alisphenoid
Primitive placentals dental formula
3-1-4-3/3-1-4-3
Eutherian dental characters
four premolars, three molars (some very primitive ones have a fifth premolar)
last premolar tends toward molariform (not seen in marsupials)
upper molars have much smaller stylar ridge (cingulum; on lingual side of protocone) with smaller cusps
Eomaia (Eutheria)
“dawn mother”
Early Cretaceous Eutheria
Barremian part of the Yixian Fm., China ~125 Ma
carbonaceous traces of hair
last premolar shows tendency towards molarization
Asioryctida (Eutheria) characteristics
Late Cretaceous
Incompletely co-ossified atlas ring
Absence of superposition of astragalus and calcaneum
Epipubic bone
Leptictida (Eutheria) Characteristics
Late Cretaceous - Eocene
Unspecialized dentition; probably 5 premolars (primitive)
Asia – Kennalestes
North America – Gypsonictops
Zalambdalestidae (Eutheria)
Asia (Uzbekistan, Mongolia)
3 incisors; first lower incisor procumbent
enamel only on labial side (possibly related to modern Glires)
Palaeoryctida (Eutheria)
Late Cretaceous
Palaeocene
Cimolestes and Paranyctoides in N. America and Asia in Late Cretaceous
specialist carnivores
tendency to develop transversely oriented shearing
edges on molars
Zhelestidae (Eutheria)
Late Cretaceous - Palaeocene
Asia, Europe, North America
basal members of single ungulate radiation
small mammals with incipient ungulate molars (broad, subrectangular in shape)
omnivory (plants & insects) or herbivory
Eutherian survivors of K/Pg extinction
Leptictida – abundant in North America and also in Europe through Eocene and to Oligocene; China in Palaeocene
Palaeoryctida – common North America through Eocene
Zhelestidae – a few teeth still in early Eocene North America
New Palaeocene Placental Groups
Anagalida
Mixodonta
Pantolestida
Apatemyida
Plesiadapiformes
‘Condylarthra’ = condylarths (carnivorous and herbivorous forms
Taeniodonta
Pantodonta
Tillodonta
Dinocerata
ArctostylopidaMeridiungulata (5 groups)
Anagalida (Placentals)
early Palaeocene, China
herbivores and omnivores
procumbent lower incisors
relationships not clear; maybe with Asian Cretaceous zalambdalestids
Mixodonta (Placentals)
Palaeocene
Asia
small, omnivorous
gliriform incisors
Mimotona – dental formula like lagomorphs
Heomys – dentition like rodents
Pantolestidae (Placentals)
Palaeocene - Oligocene
otter-like
North America and Europe
well-developed canines
Eocene Messel Buxolestes with fish in stomach
Apatemyida (Placentals)
North America and Europe
early Palaeocene to late Oligocene
single large, procumbent lower incisor
large first upper incisor
bunodont
Plesiadapiformes (Placentals)
Palaeocene
North America, Europe
most small but some up to 5 kg
some at least arboreal
related to Euarchontoglires, but maybe not Primates
Condylarths (Placentals)
Palaeocene - Eocene
North America
variety of small primitive members
larger herbivores and secondarily carnivorous forms
ancestors of various modern ungulate groups
paraphyletic; ‘archaic ungulates’
crushing molars rather than shearing
bunodont
incipiently ‘hoof-like’ terminal phalanges
Carnivorous condylarths (Placentals)
Palaeocene
North America, Asia; into Eocene
tendency to evolve higher sharper molar cusps; shearing edges
first radiation of relatively large, predaceous placentals
Cete: Triisodontidae, Hapalodectidae, Mesonychidae
Phenacodontids (herbivorous condylarths)
Palaeocene - Eocene
premolars enlarged, more molar-like (molars broader)
digitigrade
well-developed hooves on each digit
China & North America
Taeniodonta (Placentals)
Palaeocene - Eocene
North America
omnivorous
early forms - generalized, non- cursorial, good climber, opossum- like life style
later forms - gliriform incisors, more powerful build, flattened claws, digger
Schowalteria clemensi (Taeniodonta)
Scollard Formation Trochu, Alberta
Late Cretaceous (Maastrichtian)
Fox and Naylor
largest Mesozoic mammals
robust canine with restricted enamel on crown, enlarged incisor I2, structure of zygomatic arch
Pantodonta (Placentals)
early Palaeocene China; mid-Palaeocene to mid- Eocene NA
first large herbivorous placentals
dentition complete (canines present, no diastema)
V or W shaped lophs on postcanines
Tillodonta (Placentals)
early Palaeocene – late Eocene China; late Palaeocene – middle Miocene North America; early Eocene Europe
powerful jaws
enlarged chisel-like second incisors with enamel only anteriorly
lost other incisors, canines
Trogosus
Dinocerata (Placentals)
Palaeocene - mid-Eocene
China and North America
Cope and Marsh
mid-Eocene, China and North America
bony protuberances on skull
reduce or lose upper incisors, and enlarge and protrude upper canines
browsers, possibly with mobile proboscis
v-shaped crest on molars
Digitigrade
Meridiungulata (Placentals)
Palaeocene - Plio-Pleistocene
South American native ungulates
large grinding lophodont premolars and molars
hoofed feet
Darwin
Notoungulata (Meridiungulata)
Palaeocene
South American; Mixotoxodon made it to Central America in Pleistocene
broad, flat skull; strongly lophodont and full hypsodonty
4 groups (notioprogonians, toxodonts, typotheres and hegetotheres)
Astrapotheria (Meridiungulata)
Palaeocene
no upper incisors; upper canines large tusks; last two molars huge
nostrils on top of skull
very large; Astrapotherium
Xenarthra (armadillos, sloths, anteaters) Synapomorphies
South America
unique accessory articulations between vertebrae (hence name which means “ancient joint”)
absence of enamel on teeth
Glyptodonta (Cingulata, Xenarthra, armadillos)
trilobed, unrooted cheek teeth for grinding
middle Eocene to Pleistocene
largest Glyptotherium (Pleistocene) which was more than 3m long
Argentina
Phyllophaga (Pilosa, Xenarthra)
sloths
modern sloths = arboreal; most of Cenozoic ones were ground sloths
late Eocene; abundant in Miocene S. America, extinct ~5000 ybp
Megatherium
Plio-Pleistocene Great American Biotic Interchange
North and South America were disconnected until about 3 mya, when volcanic Isthmus of Panama formed
Sparassodont marsupials disappeared for unclear reasons
didelphimorphs invaded Central America
many northern forms came south
Flow of water cut-off
Rodents replaced small marsupials
Animals going North during the Plio-Pleistocene Great American Biotic Interchange
xenarthrans: ground sloths, armadillos, glyptodont (Glyptotherium), giant anteater (Myremcophaga); notoungulates (Mixotoxodon); hystricomorph rodents (porcupine Erethizon into Canada); didelphid marsupials (as far as Canada)
Animals going south during the Plio-Pleistocene Great American Biotic Interchange
mustelids (skunk); Tayassuid peccary; Horse (Hippidion and others); Carnivora (dogs and cats); bears; gomphothere elephants; tapirs; camels; deer; shrews; several rodents
Animal in Pre-Pliocene South America
carnivores = borhyaenid (marsupials)
meridiungulates = dominant herbivores
xenarthrans with specialist rolls (anteaters and leaf browsers)
marsupial carnivores declining prior to Pliocene
meridiungulates in decline (some survive till late Pleistocene)
Afrosoricida (Afrotheria)
Chrysochlorida – golden mole
Tenrecida – tenrecs and otter shrews
very poor fossil record, restricted to Africa (Chrysochlorida) and Madagascar (tenrecs), Miocene; possible Fayum specimen (late Eocene)
zalambodont molars
Macroscelidea (Afrotheria)
elephant shrews
only fossils are in Africa
middle Eocene teeth of Chambius from Tunisia
late Eocene Herodotius from Fayum
four-cusped bunodont molars lacking lophs
Tubulidentata (Afrotheria)
Oligocene Europe and Miocene Africa
dentine prisms
aardvark
single living species (Orycteropus afer)
Preserved with elephant birds
Sirenia
early Eocene in Europe; world-wide by late Eocene
bilophodont molars like proboscideans
Prorastomus (primitive Sirenia)
skull only
from Jamaica
lack ventral deflection of rostrum found in others
Pezosiren (Sirenia)
uncompressed neck unlike more derived forms
probably capable of terrestrial locomotion
tall anterior neural spines
strong connection between ilium and sacral vertebrae
relatively short limbs
aquatic: dorsal nostrils, dense ribs
Order Desmostylia
early Oligocene to the late Miocene
exclusively northern Pacific Rim
marine mammals; semi-aquatic, hippo-sized
6 genera from
ate seaweed, and probably related to Sirenia
Diastema
Broad hands and feet
Palaeoparadoxia (Order Desmostylia)
well-developed limbs
paddle-like hands and feet
incisors and canines well developed
cheek teeth resemble those of proboscideans
order Embrithopoda
primarily African but first appear in Europe
jaws and teeth
late Palaeocene to late Eocene
most basal Paenungulata
Arsinoitherium
Hyracoidea (Afrotheria)
earliest = late Eocene/early Oligocene of Fayum
dispersed into much of Asia
browsers (teeth bunodont, lophodont,
selenodont)
Titanohyrax bigger than modern hyrax
Megalohyrax – skull 40 cm long
Proboscidea (Afrotheria)
earliest Eocene, Morocco
true lophodont molars
transverse crests uninterupted by conules
bilophodonty (primitive proboscidean condition)
Phosphatherium, Palaeomastodon
Moeritherium (Proboscidea, Afrotheria)
enlarged second incisors
bilophodont molars
reduced tail
short stout limbs semiaquatic
maybe Sirenia or Desmostylia rather than Proboscidea?
African proboscidean radiation
Dinotheres (late Oligocene Ethiopia) seem to have evolved directly from a Moeritherium-like ancestor
deinotheres represent a distinct line evolution that probably diverged very early in the history of proboscideans
Deinotheres (Proboscidea, Afrotheria)
first in late Oligocene, Ethiopia
only 2 or 3 simple lophs on molars
tusks
Came into Asia through ‘Gomphothere Land Bridge’ in middle Miocene (Arabian Peninsula)
Palaeomastodon and Proboscidea evolution
believed to have given rise to subsequent radiation of proboscideans other than deinotheres
Mastodons (Mammutidae, Afrotheria)
Miocene until start of Holocene
name derives from Greek: nipple tooth
e.g., Zygolophodon, one of largest terrestrial mammals ever
Thomas Jefferson
Gomphotheriidae (Proboscidea)
molars differ from elephants (but like mastodons) - bunodont
earlier species with four tusks
retracted facial and nasal bones - trunks
widespread in North America during the Miocene and Pliocene (12–1.6 mya)
lived in parts of Eurasia, Beringia, and with the Great American Interchange into South America
starting about 5mya - replaced by elephants
Platybelodon grangeri (Gomphotheriidae, Proboscidea)
tusks like incisors
Scooped up water and plants
Lived in wet areas
Stegadontidae (Elephantoidea)
Miocene to Pleistocene
distinct ridges on molars
Beresovka Mammoth
Euarchontoglires
arisen in Early Cretaceous
Plesiadapiformes, Primates, Dermoptera (colugos = flying lemurs), and Scandentia (tree shrews)
Scandentia (Euarchontoglires, tree shrews)
Eocene Eodendrogale, Asia
1-2 Miocene forms (plus extant genus Tupaia)
Auditory bullae
Complete zygomatic arch
Dilambdodont
Dermoptera (Euarchontoglires, flying lemurs)
two extant species: Cynocephalus volans = Philippine flying lemur (colugo); Galeopterus variegatus = Sunda flying lemur
two extinct species Dermotherium major and D. chimaera from Eocene, Thailand are fully evolved dermopterans
2 pairs of lower incisors comb-like – cheek teeth with wrinkled enamel
postcranial adapted for gliding membrane stretched between all four limbs
Nearly completely postorbital bar
Boreoeutheria/Boreotheria
Euarchontoglires + Laurasiatheria
sister groups probably split about 85 to 95 mya;
early Palaeocene
males share characteristic of external testicles (keep sperm cool); except in rhinos, hippos, cetaceans, etc.
Palaeontologists that discovered Purgatorius
holotypes of two species with a single tooth (molars) each from Montana
one species (Purgatorius ceratops) was from the Hell Creek Formation
Leigh van Valen and Robert Sloan
Purgatorius
the oldest and most primitive plesiadapiform
long known only from isolated teeth and jaw fragments since it was first discovered 50 years ago
Ankle bones diagnostic characteristics for mobility
Plesiadapiformes
most primitive primates
Cretaceous specimens from Alberta and Montana:
Immediate post-Cretaceous origin for Primates followed by Palaeocene radiation
Arboreal
mobile shoulder and elbow joints
mobile hip joint capable of a great range of abduction and lateral rotation
lower ankle joint mobility for inversion of the foot
terminal phalanges suggest “claw-clinging”
Primate characters lacking in Plesiadapiformes
no postorbital bar
no shortened snout
supposedly no opposable hallux and pollex
Plesiadapis vs Carpolestes (Plesiadapiformes)
Plesiadapis - short fingers, extremely long hook- like claws, lessened ability to grasp small-diameter supports
Carpolestes - shorter claws, longer fingers and toes, relatively shorter metacarpals and metatarsals, foot with a divergent, opposable hallux with a nails, better adapted for grasping small diameter supports in a powerful and precise manner
Carpolestes simpsoni (Plesiadapiformes)
almost complete specimen found in U. Paleocene, Wyoming
has opposable first digits with nails
Ignacius dawsonae (Plesiadapiformes)
Ellesmere Island
Early Eocene
Showed Arctic was warm and diverse
Order Primates
found in wide diversity of habitats on all continents except Antarctica
Auditory bulla derived from petrosal portion of the temporal bone
Adapiformes (Primates)
thrived in Eocene; disappeared from most of Northern Hemisphere with cooling climate; gone by end Miocene (~7 mya)
some are lemur-like
may/may not be link between Strepsirrhini and Haplorrhini
unspecialized bunodont dentition – frugivores
Notharctus
‘Ida’ Darwinius masillae (Primates)
oldest and most complete primate fossil in the world. Even the stomach contents have been preserved
Messel, Germany
‘prosimians’ (Primates, lemurs, galagos, lorises, tarsiers)
retain primitive features:
small brain, long snout, postorbital bar not expanded to enclose back of orbit
incisors rounded not spatulate
mandibular symphysis unfused
earliest Eocene, then radiate through northern continents
Strepsirrhini
lemurs (Madagascar), galagos (bushbabies, Africa), lorises (India and SE Asia and pottos of Africa), †Adapiformes (Eocene, N. America, Europe, Asia)
wet nose/rhinarium
vomeronasal organ
bicornuate uterus with epitheliochorial placenta
tapetum ludicum
have bony eye ring but lack thin wall of bone behind
produce vitamin C
tooth comb (not in adapiforms)
grooming claw on second toe (?adapiforms)
Lemuriformes & Lorisiformes (Strepsirrhini)
very poor fossil record
first lemuriform = Miocene East Africa; Pleistocene,
Madagascar
first lorisiform = mid Eocene, Fayum (teeth)
Omomyoidea (Haplorrhini, Primates)
Eocene
insectivorous,
arboreal
tarsier-like (large orbits, no PM1)
Shosonius
Tarsiiformes (Haplorrhini)
fossil = Europe, north Africa, Asia, North America
extant = SE Asia, Tarsiidae
may include Omomyidae
Semi-bunodont
Simiiformes
Haplorrhini excluding Tarsiiformes
migration to South America via rafting or land bridge in Oligocene
Platyrrhini (Simiiformes, Haplorrhini)
New World monkeys; five extant families
only spider monkeys and relatives (Atelidae) have prehensile tails
‘flat nose’ and sideways nostrils compared to Old World monkeys and apes
earliest fossils; 25 mya Branisella; 20 mya Chilecebus
Catarrhini (Simiiformes, Haplorrhini)
Old World monkeys and apes
Africa and Asia
Aegyptopithec
Cercopithecidae (monkeys) + Hominoidea (apes = Hylobatidae + Hominidae)
arboreal to fully terrestrial
Aegyptopithecus (Catarrhini, Simiiformes, Haplorrhini)
Catarrhini
Jbel Qatrani Fm, Fayum, ~30 mya
Hylobatidae (Catarrhini, Simiiformes, Haplorrhini)
small or lesser apes = Gibbons
primary mode of locomotion = brachiation
walk bipedally with arms raised for balance
Yuanmoupithecus - Yunnan, China Miocene
Macroscelidea (Elephant Shrews)
Africa, Eocene to present
insectivorous
saltorial
When did Rodents and lagomorphs diverge and radiate?
diverged from other placentals by a few million years after K-Pg boundary
radiated in Cenozoic
Standard rodent dentition
only 2 gliriform lower incisors
double layer of enamel on front surface of rodents (single in lagomorphs)
fully molariform fourth premolars
described by Hayden, Hatcher, Leidy, Osborn, Marsh
White River Badlands
Rodentia radiation
majority of families appear in late Eocene (explosive radiation)
took over niches from multituberculates that were lost by end-Cretaceous extinction
Spread all over, crossing oceans from Africa to Madagascar and South America; only terrestrial placentals to reach Australia by themselves
Ischyromyids (Euarchontoglires)
late Palaeocene of North America
standard rodent dentition
primitive jaw- closing musculature
no specialization in rostrum for masseteric musculature
basal rodents
Two gliriform incisors (enamel on the front)
Fully molariform fourth premolars
Paramys (Ischyromyid)
Eocene
sister to squirrels and mice rather than as a
basal sciuromorph
North America, Eurasia, Africa
may have given rise to Sciuromorpha and Myomorpha
Messel, Germany
Mixodonta (Euarchontoglires)
early Palaeocene Asia
Heomys
single pair of gliriform incisors, postcanines similar to basal rodents
Groups in Rodentia
Castorimorpha–beavers, pocket gophers, kangaroo rats, kangaroo mice
Hystricomorpha–(includes caviomorphs) gundis, porcupines, pacas, pacaranas, agoutis, cane rats, capybaras, nutria, chinchillas
Myomorpha–mice, rats, gerbils, hamsters, lemmings, voles
Sciuromorpha–mountain beaver, squirrels, chipmunks, marmots, flying squirrels, dormice
Hystricomorpha (Rodentia) dispersal
Originate in Asia about 40 mya, invade Africa in late Eocene, then into South America (across ocean) by early Oligocene
Rodentia dispersals during the Great American Biotic Interchange
New World porcupines head north
sigmodontines (New World mice/rats) head south and explosively radiate in South America, preventing other rodents from doing the same
When did Rodentia arrive in Australia
arrive in Australia from Indonesia at the end of the early Pliocene about 5 mya
Rodentia zygomasseteric system
propalinal (fore-aft motion while teeth in occlusion) motion of jaw enabled by an extension of the zygomatic arch and the division of the masseter muscle into three distinct parts
Rodentia 3 masseter muscles
superficial masseter
lateral masseter
medial masseter
moving origin of muscles anteriorly - additional range of movement for lower jaw and stronger force
main force for closing rodent jaw
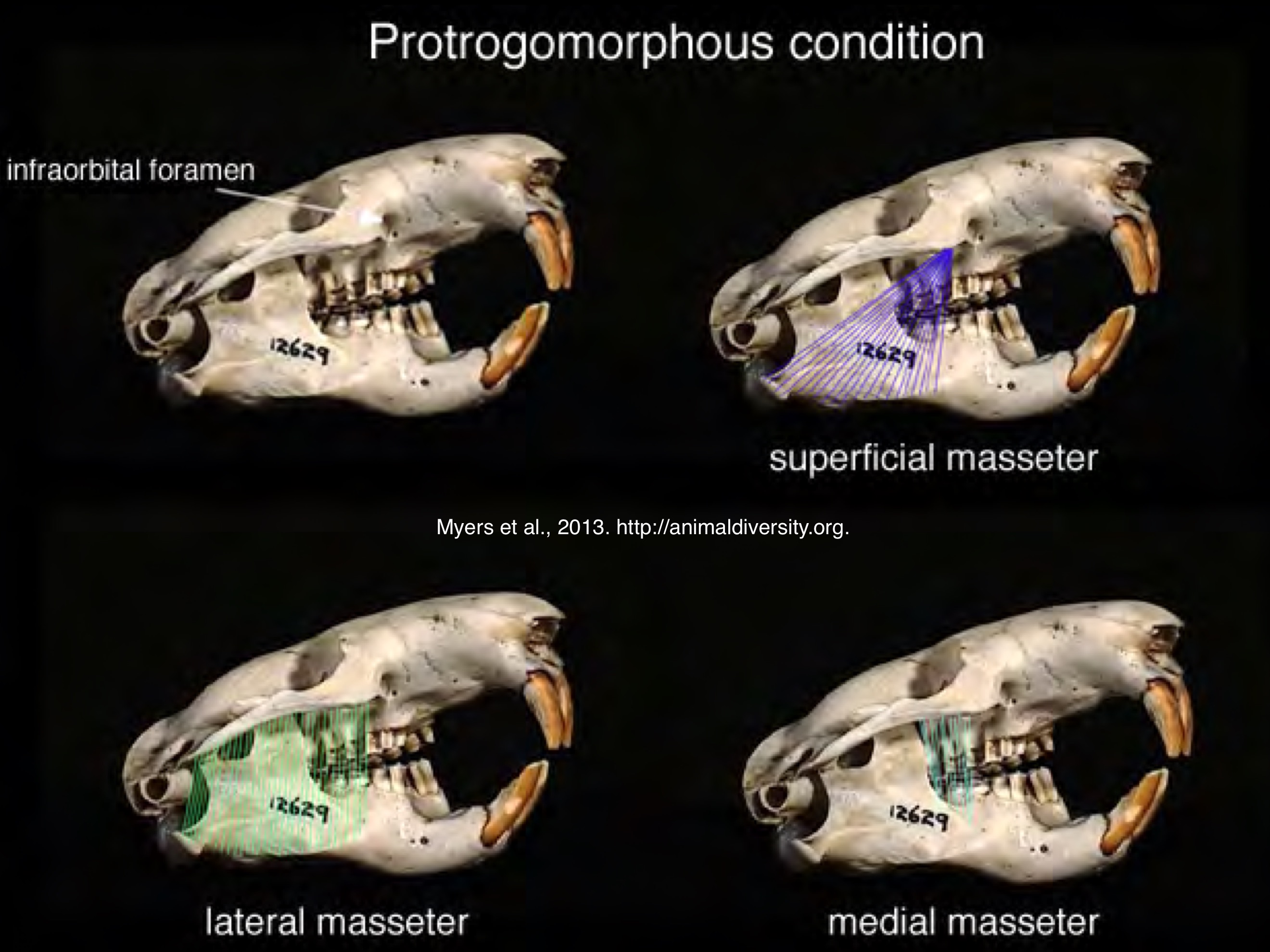
Rodentia protrogomorphy zygomasseteric system
snout unmodified; masseter originates on ventral surface of zygomatic arch
Only seen in aplodontiidae (mountain beaver) and some fossil groups
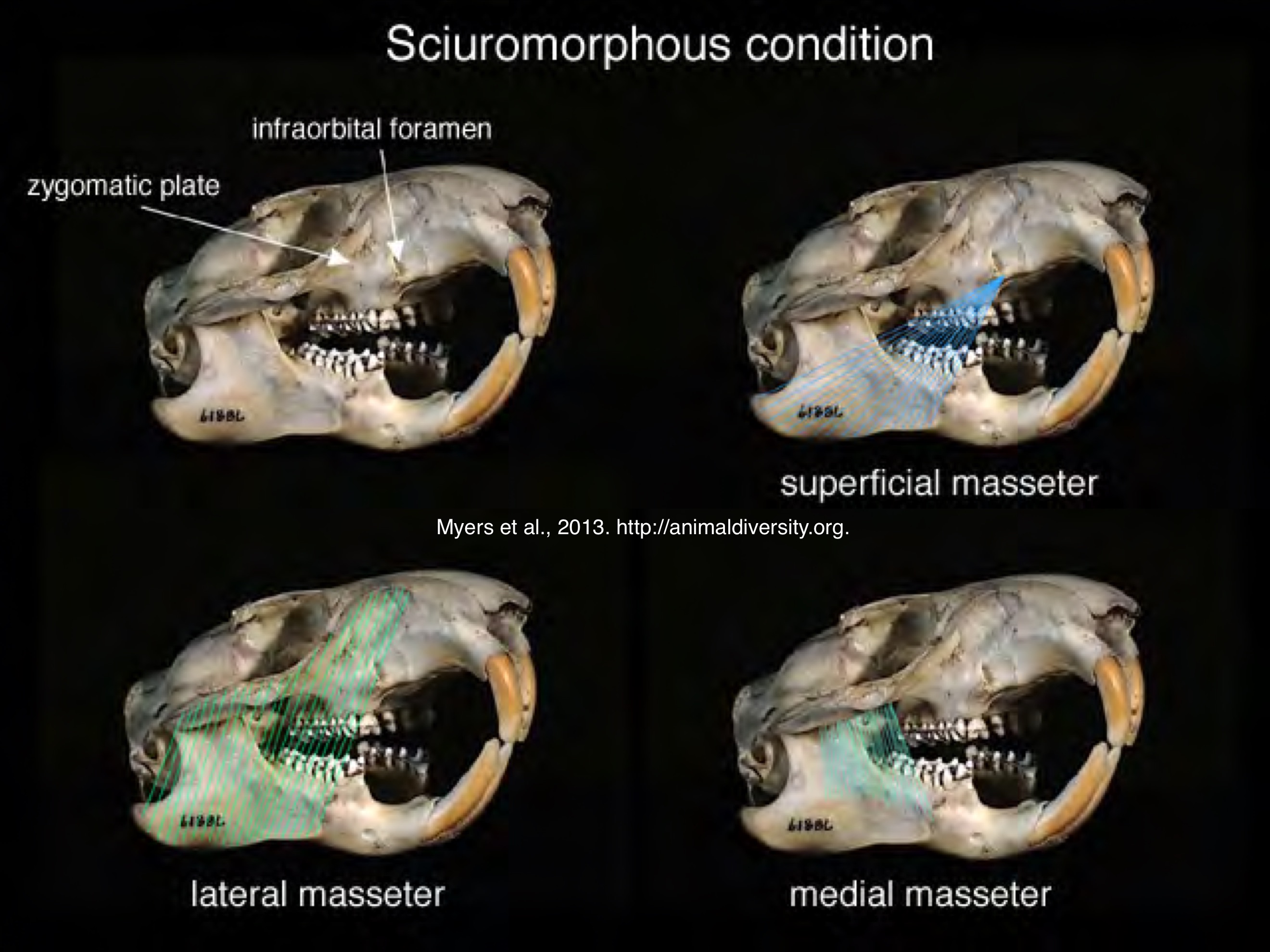
Rodentia sciuromorphy zygomasseteric system
ventral surface of zygoma (cheek) tilts and broadens into zygomatic plate; lateral masseter extends forwards onto snout, superficial masseter extends forwards along zygoma
in Sciuromorpha
Trogontherium cuvieri von Waldheim (Rodentia)
Europe, Siberia, China
Pliocene, Pleistocene
What group are Casteroides ohioensis, Trogontherium cuvieri von Waldheim, Palaeocastor in?
Rodentia
Palaeocastor - Miocene beaver
Hystricomorph rodents radiation
South American radiation originated with dispersal from west Africa during Oligocene
includes capybaras
What group are Erethizon dorsatum (porcupine), Josephoartigasia monesi, Phoberomys and the Patagonian hare in?
Hystricomorph rodentia
Phoberomys - Late Miocene Venezuela, hippo-sized
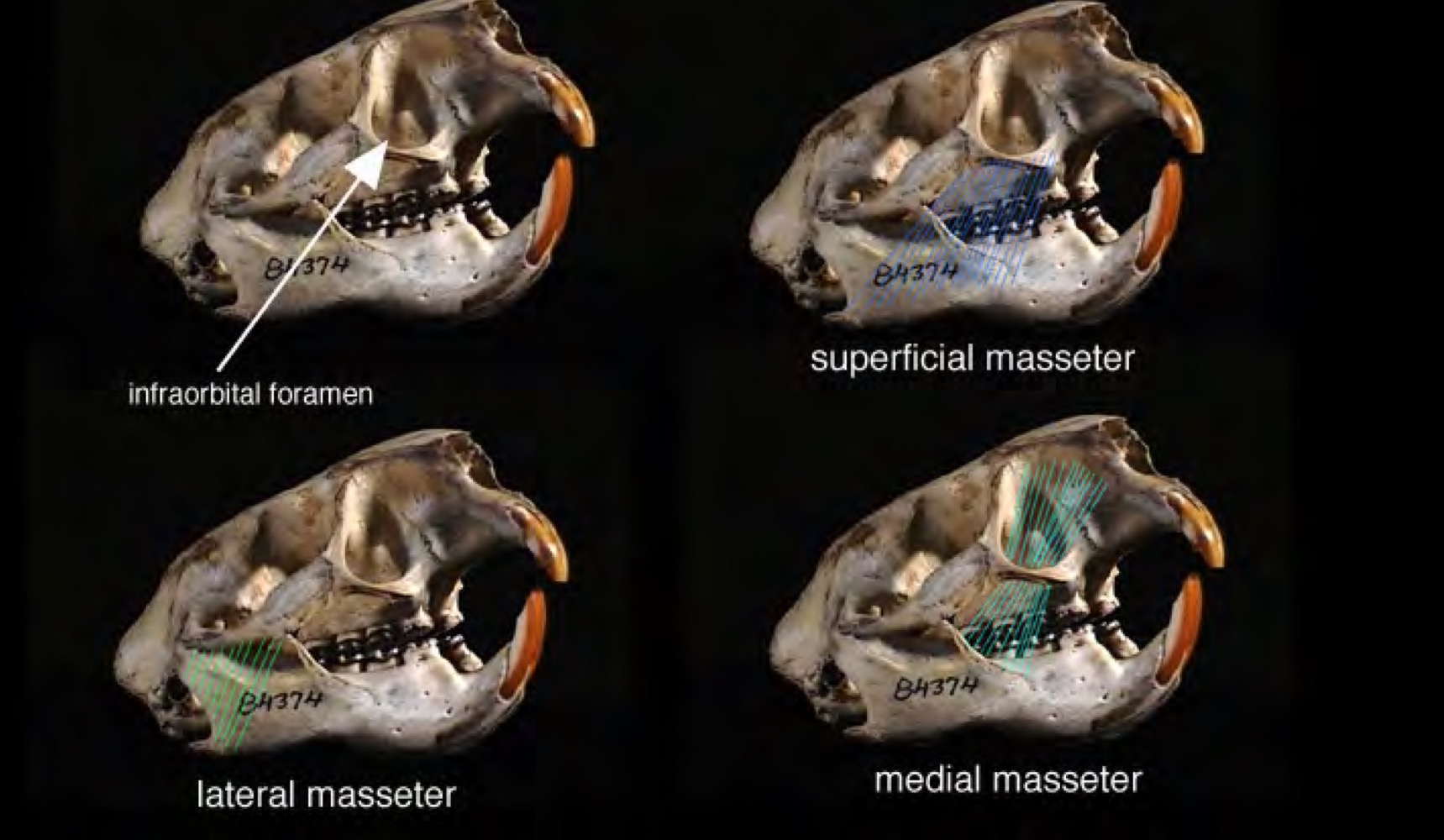
Rodentia Hystricomorpha zygomasseteric system
medial masseter enlarged, passes through enlarged infraorbital foramen; superficial masseter originates on front edge of zygoma; lateral masseter extends along zygoma
Hystricomorpha, Anomaluromorpha, some Sciuromorpha (Gliridae, dormice)
ateral masseter attaches to the back of the zygomatic arch
origin of the superficial masseter is on anterior part of zygomatic arch
angular process lies lateral to the vertical plane of the tooth row.