CHAPTER 7 – ELECTRICAL EXCITABILITY AND ACTION POTENTIALS
1/54
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced |
|---|
No study sessions yet.
55 Terms
inhoud
1 — potentialen
2 — ionkanalen
1 — POTENTIALEN
soorten potentiaal
graded potentials = elektrotonische potentialen
action potentials = AP
graded potentials
lineair verband tss amplitude en stroom
gevoelig aan;
demping
storingen
action potentials
niet-lineair verband tss amplitude en stroom
niet gevoelig aan;
demping
storingen
Vx
Vx = V0 * e(-x/λ)
V0 = startwaarde
λ = lengteconstante
lengteconstante (λ)
λ = sqrt(rm/ri) = sqrt(aRm/2Ri)
rm = Rm/2rπ ; weerstand membraan
ri = Ri/r²π ; interne weerstand
a = straal
afstand waarop je potentiaal gedaald is tot 37% (= 1/e%) van zijn oorspronkelijke waarde
exponentiële daling
Ri is meest bepalend voor λ (want r²)
grote λ
nodig;
grote rm
kleine ri
grote straal neuron
leidt tot verdere verspreiding signaal
probleem axonen
hebben kleine λ, maar we willen grote λ
opl; myelinisatie (tot 300x) + knopen Ranvier
Rm ↑ ; signaal raakt verder
Cm ↓ ; signaal wordt sneller doorgegeven
tijdsconstante (τ)
τ = Ri * Cm
tijd waarop je potentiaal gedaald is tot 37% (= 1/e%) van zijn oorspronkelijke waarde
graded potentials vs action potentials — tabel
graded | action | |
type |
|
|
amplitude | lineair verband | niet-lineair; alles-of-niets |
optellen | ja | nee |
drempel | nee | ja (~ -60mV) |
propagatie-snelheid | neemt af naarmate je verder gaat | constant |
graded potentials vs action potentials — afbeelding
merk op; boogvormig ipv vierkant vanwege capacitaire eigenschappen membraan
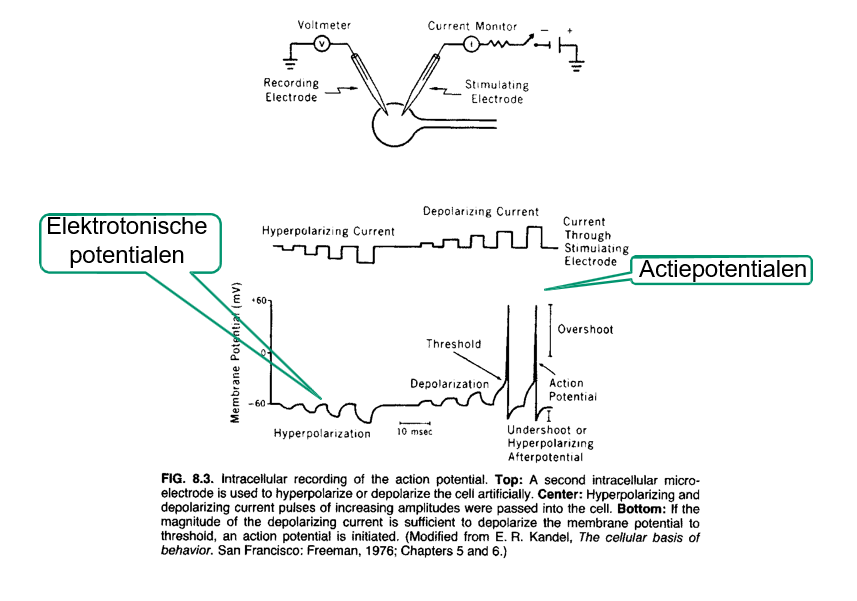
graded potentials vs action potentials — afbeelding
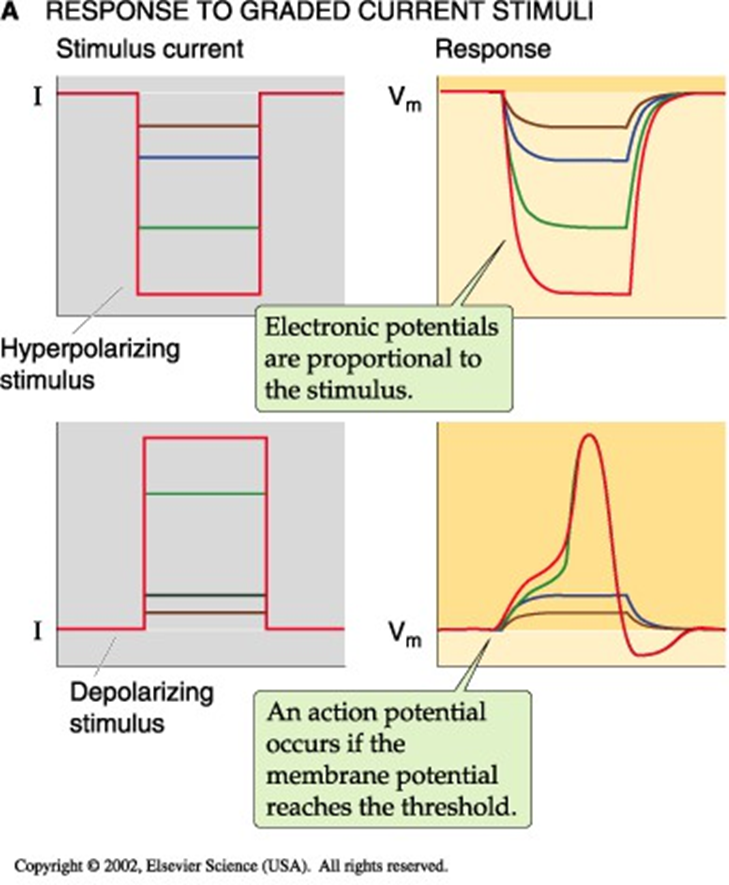
actiepotentiaal
tijdsafhankelijke verandering in Vm
alles-of-niets
amplitude = cte; maar verchilt wel van weefsel tot weefsel
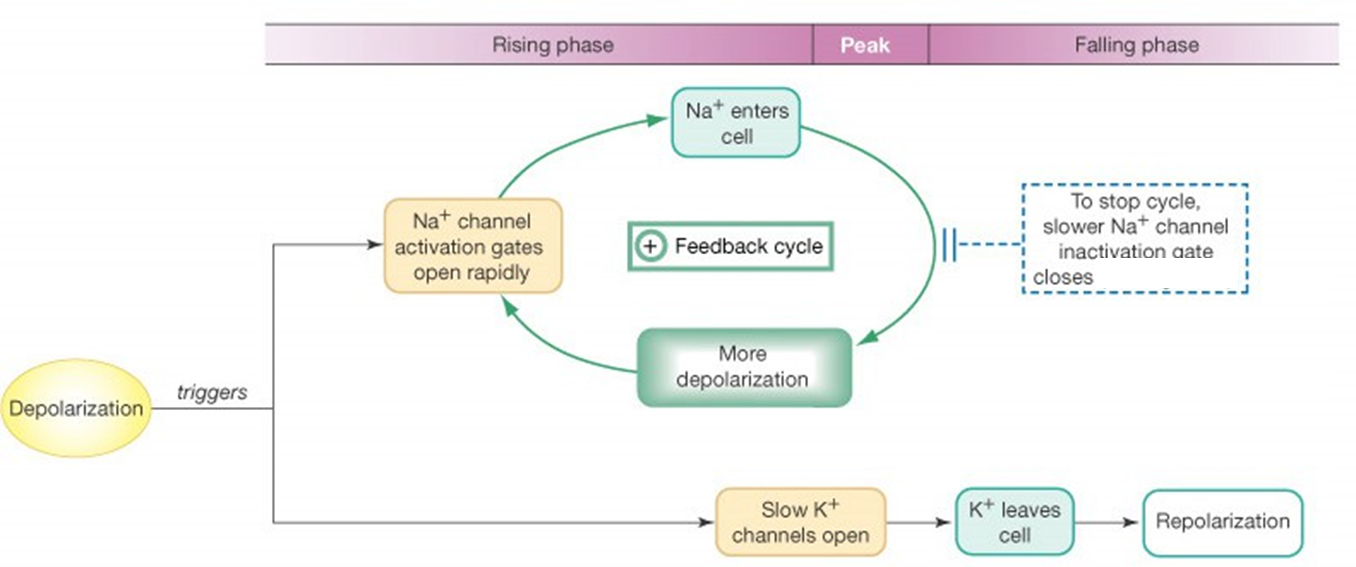
principe actiepotentiaal
membraan depolariseert door bepaald signaal
INa inwaarts → depolarisatie wordt versterkt → AP
IK uitwaarts → repolarisatie → AP stopt
opm; K werkt trager dan Na = delayed rectifier
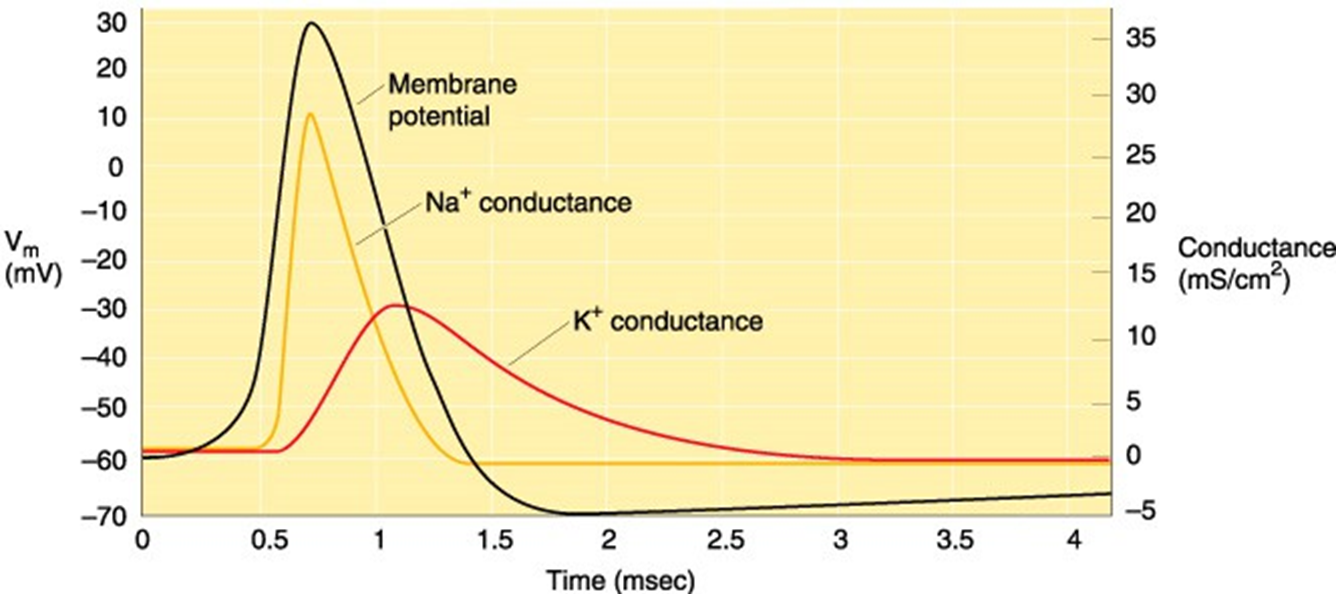
INa en IK
INa = m3h * GNa * (V-Erev)
IK = n4 * GK * (V-Erev)
opm; n / m / h verwijzen naar de
(in)activatiepoortjes (zie verder)
principe actiepotentiaal
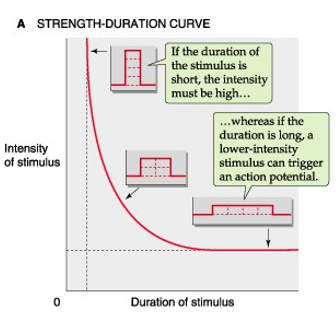
strength-duration curve
kort, maar krachtig
lang, minder krachtig
→ sterkte x tijd
chronaxie = min. tijdsduur
rheobase = min. sterkte
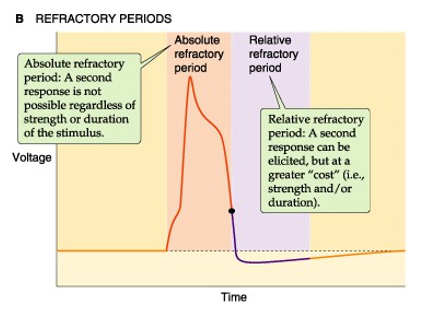
refractaire periode
absoluut
geen nieuw AP mogelijk
te veel h-poortjes zijn dicht
relatief
nieuw AP mogelijk, maar drempel-waarde ligt hoger
nog voldoende h-poortjes open
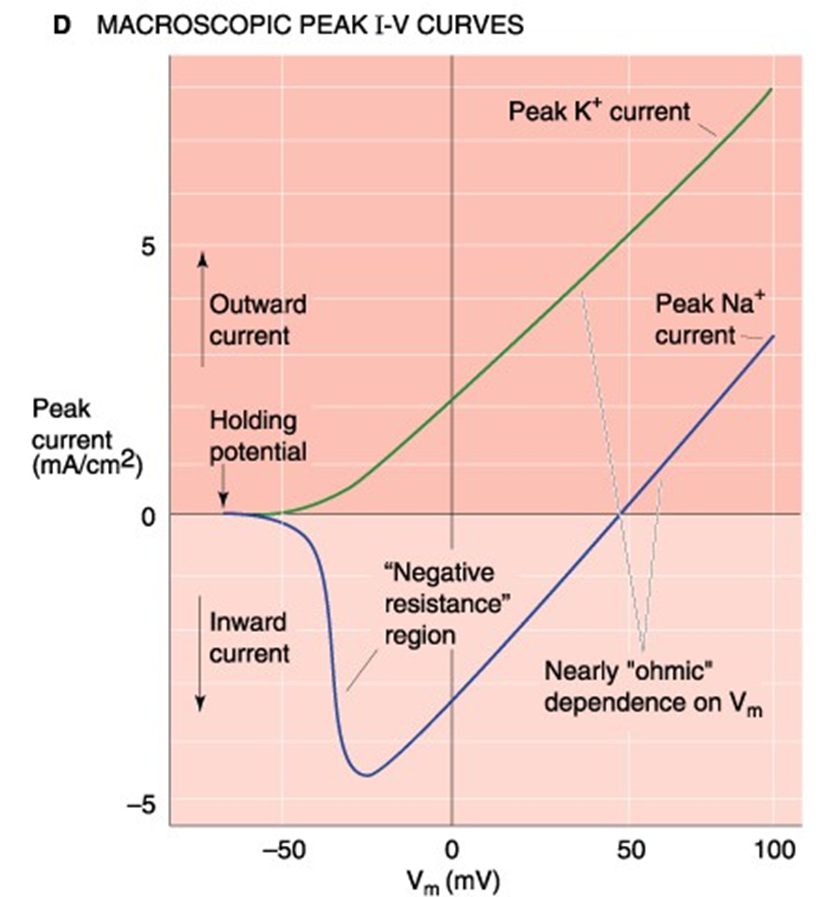
I-V-curve
regio negatieve weerstand
positief terugkoppelsysteem
Na+ - instroom depolariseert membraan nog meer, wat leidt tot nog meer Na+ - instroom
neurotoxines (3)
TTX & STX
afkomst;
TTX; bacteriën → kogelvis, octopus …
STX; dinoflagellaten → schelpdieren
blokkeren Na+ - kanalen
voorkomen depolarisatie, en dus ook AP
TEA
synthetisch
blokkeert K+ - kanalen
voorkomt repolarisatie; verlengt AP
drempelwaarde
houdt 2 zaken in;
drempelspanning voor activatie van Na+ - kanalen
opening van een voldoende aantal Na+ - kanalen om een kritische Na+ stroom te bereiken die andere repolariserende stromen (open K+ kanalen, lekstroom) kan overwinnen
2 — IONKANALEN
Na+ - kanalen & K+ - kanalen — poortjes
Na+ - kanalen
m-poortje; activatie
h-poortje; inactivatie
K+ - kanalen
n-poortje; activatie
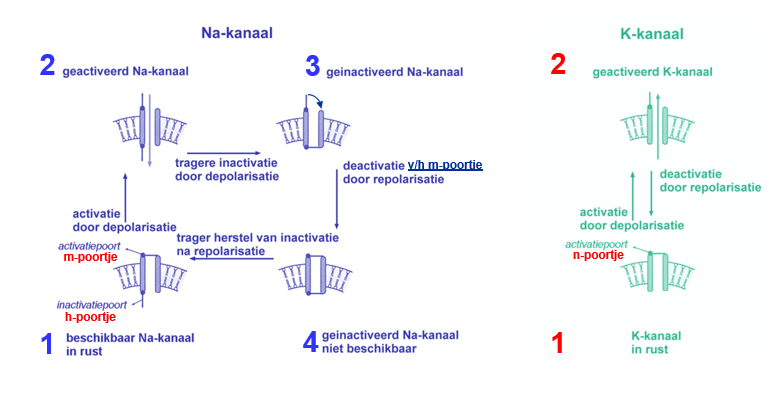
Na+ - kanalen & K+ - kanalen — poortjes
in rust; n en m dicht
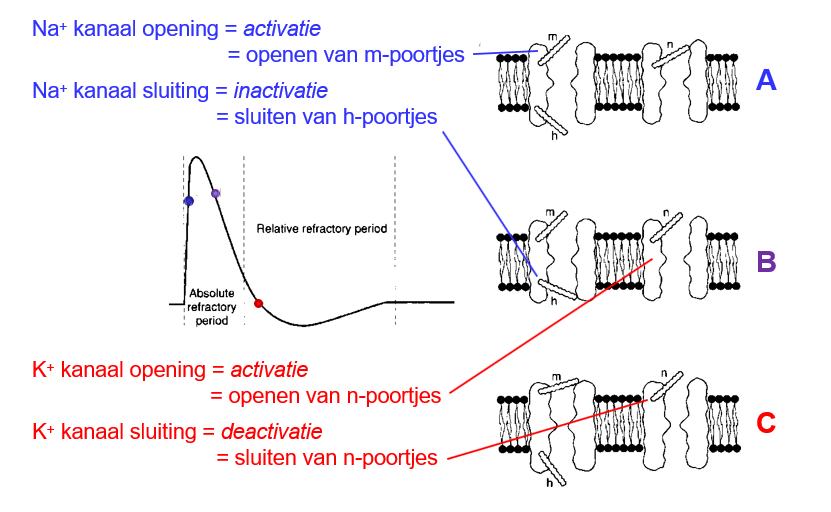
n∞
steady-state activatie
K+ - kanalen
kans dat n-poortje (activatie) open is bij een bepaald membraanpotentiaal
n∞ = 0 → n-poort dicht (kanaal is inactief)
n∞ = 1 → n-poort open (kanaal is actief)
m∞
steady-state activatie
Na+ - kanalen
kans dat m-poortje (activatie) open is bij een bepaald membraanpotentiaal
m∞ = 0 → m-poort dicht (kanaal is inactief)
m∞ = 1 → m-poort open (kanaal is actief)
h∞
steady-state inactivatie
Na+ - kanalen
kans dat h-poortje (inactivatie) dicht is bij een bepaald membraanpotentiaal
h∞ = 1 → h-poort dicht (kanaal is inactief)
h∞ = 0 → h-poort open (kanaal is actief)
n∞ , m∞ en h∞ — grafiek
merk op;
m∞ en h∞ zijn niet samen 100%
het zijn verschillende en onafhankelijke processen
m∞ = m-poortje open → iondoorstroming
h∞ = h-poortje dicht → voorkomt openen kanaal na depolarisatie
kanaal kan geopend zijn, maar toch inactief door de werking van de inactivatiepoort
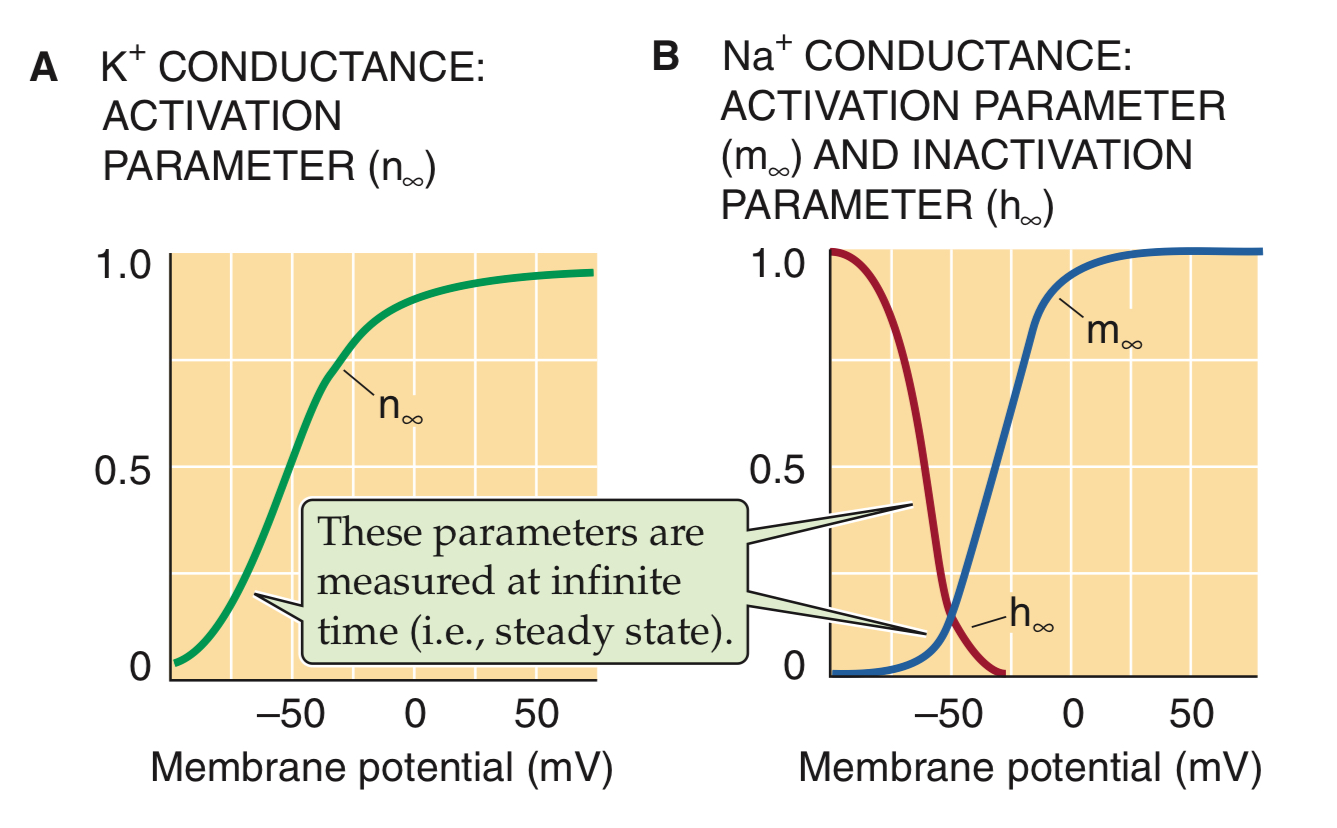
belangrijke factoren bij AP (3)
conductantie voltage (G-V)
h∞
RFI = recovery from inactivation
conductantie voltage (G-V)
G = 1/R = hoe gemakkelijk kan een ion door een kanaal bewegen
beschrijft hoe de geleidbaarheid van een kanaal verandert bij verschillende voltages
h∞
steady-state inactivatie
= hoe snel wordt het kanaal inactief na opening bij een bepaald voltage?
bepaalt dus de herstelcapaciteit van het kanaal voor de volgende actiepotentiaal
RFI = recovery from inactivation
tijd die het kanaal nodig heeft om uit de inactieve toestand te komen en weer klaar te zijn om te openen bij een nieuwe depolarisatie
accomodatie
trage depolarisatie boven de drempel
openen activatiepoorten (m-poorten)
sluiten inactivatiepoorten (h-poorten)
exciteerbaarheid daalt
trage depolarisatie onder de drempel
exciteerbaarheid stijgt
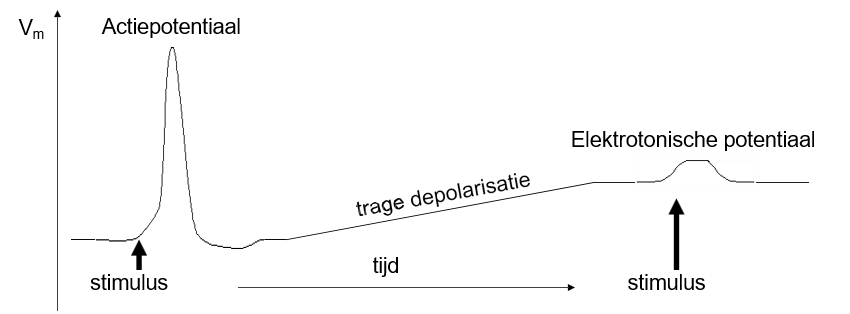
directionaliteit AP
meestal unidirectioneel
gaat weg van de plaats waar undershoot plaatsvindt; AP kan die nooit overwinnen
voortgeleidingssnelheid (v)
v = Δx / Δt
v ~ λ/τ = 1/Cm * sqrt(a/2rmri)
a = straal
afhankelijk van;
diameter axon (grote a → grote λ)
myeline (λ wordt groter)
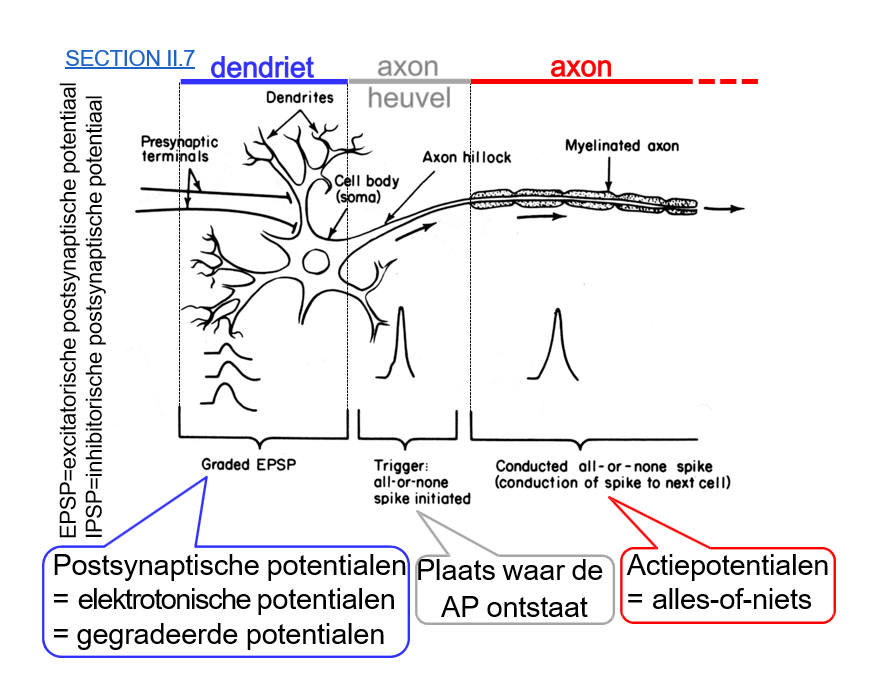
voortgeleiding signaal
dendriet
elektrotone signalen komen toe
axonheuvel
ion-kanalen
ontstaan AP; alles-of-niets
axon
saltatoire geleiding over knopen Ranvier
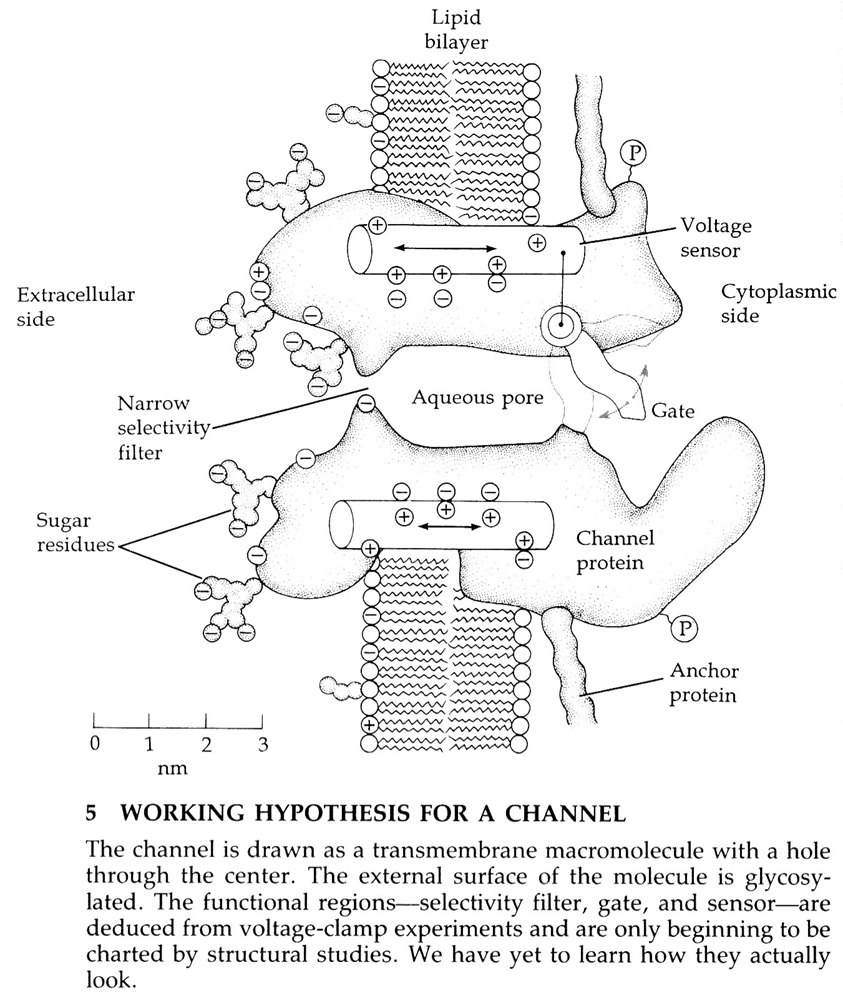
ion-kanalen — algemeen
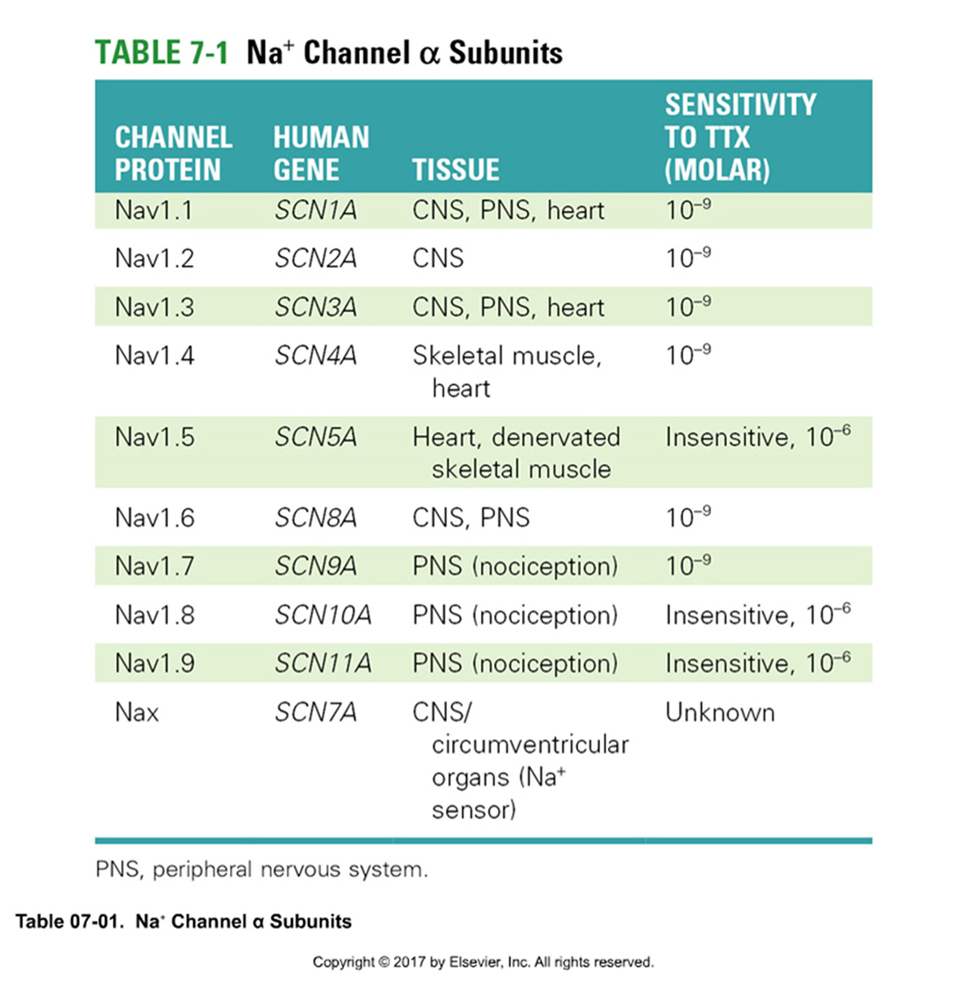
Nav - kanaal — opbouw
pseudotetrameer
4 homologe subunits
6 transmembranaire segmenten
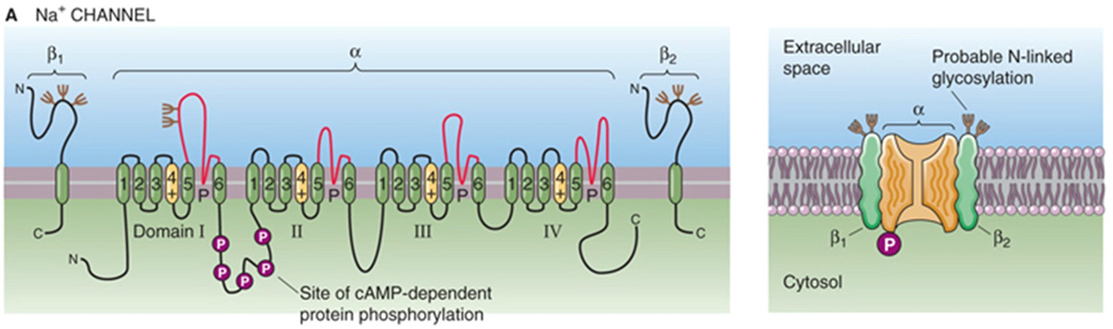
Nav - kanaal — opbouw
4 β-subunits beïnvloeden;
functie
modulatie
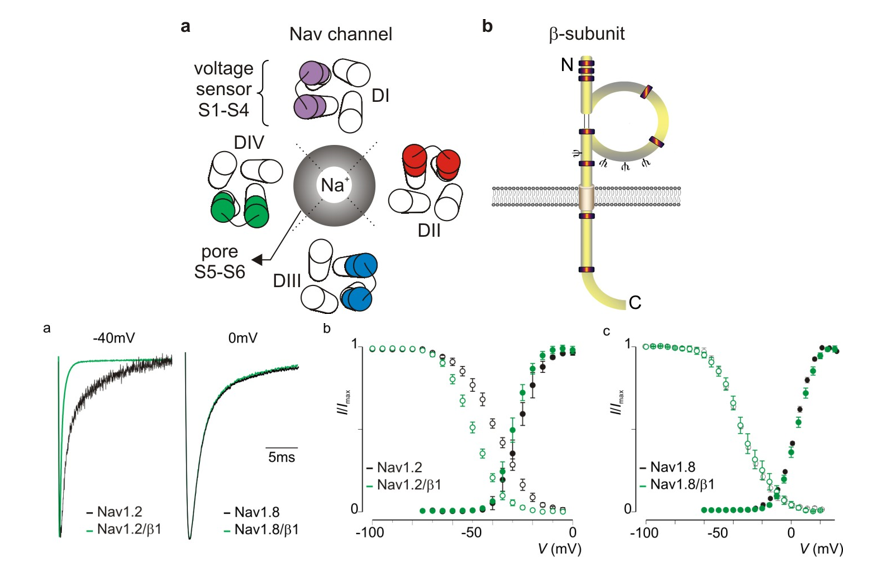
Nav - kanaal — types
CNS = centraal zenuwstelsel
PNS = perifeer zenuwstelsel
Nax
functie onbekend
lijkt op Na-kanaal, maar kan geen Na doorlaten
geen voltage-geactiveerd kanaal
Cav - kanaal — opbouw
monomeer
6 transmembranaire segmenten
Cav - kanaal — kinetische eigenschappen
membraan depolarizatie; vnl plateau-fase AP
excitatie-contractie koppeling (spiercontractie)
excitatie-secretie koppeling (neurotransmitter vrijzetting)
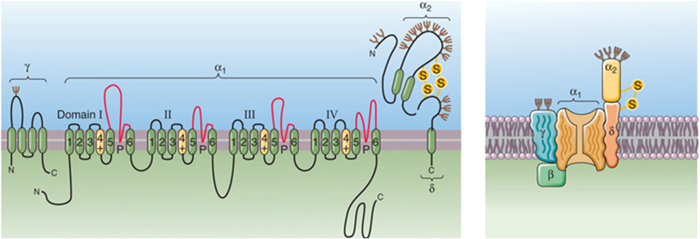
Cav - kanaal — types
Cav1 — L
spieren; DHP-receptoren
DHP blokkeert kanaal
Cav2 — N, P/Q, R
Cav3 — T
low thershold (<30mV)
repetitive firing
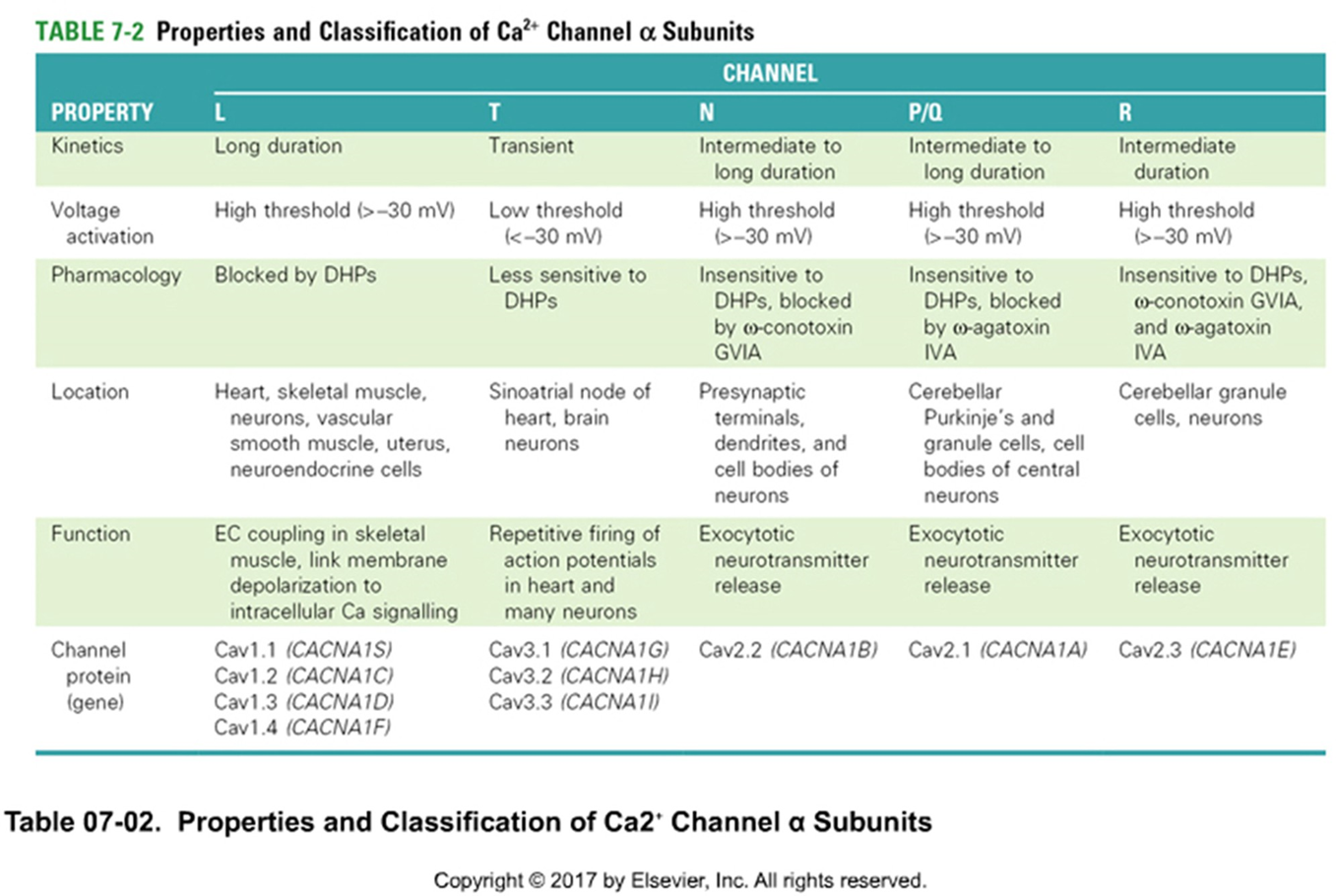
K+ - kanaal — opbouw
tetrameer
6 transmembranaire segmenten
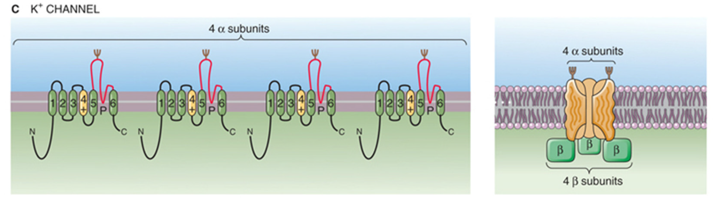
K+ - kanaal — types
op basis van eigenschappen stromen;
delayed en transient outward rectifiers
inward rectifiers
Ca2+- activated K+ currents
op basis van verschillende genfamilies;
Kv kanalen (voltage)
K2P kanalen (two-pore)
Kir kanalen (inward-rectifier)
small, intermediate en large conductantie kanalen (SKCa, KCa, BKCa)
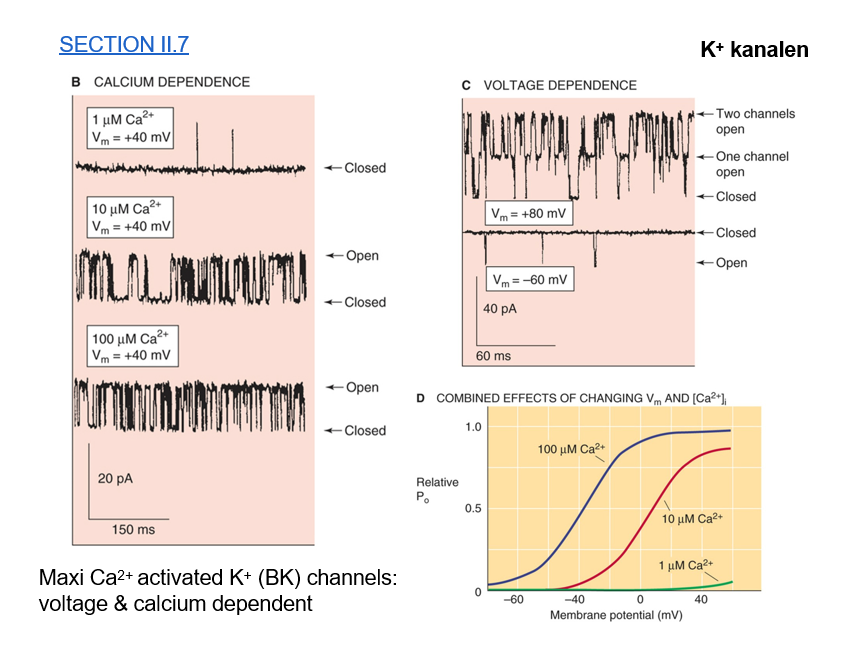
K+ - kanaal — large conductantie KCa kanalen (BKCa)
RCK = regulator conductance of K+
calcium bowl
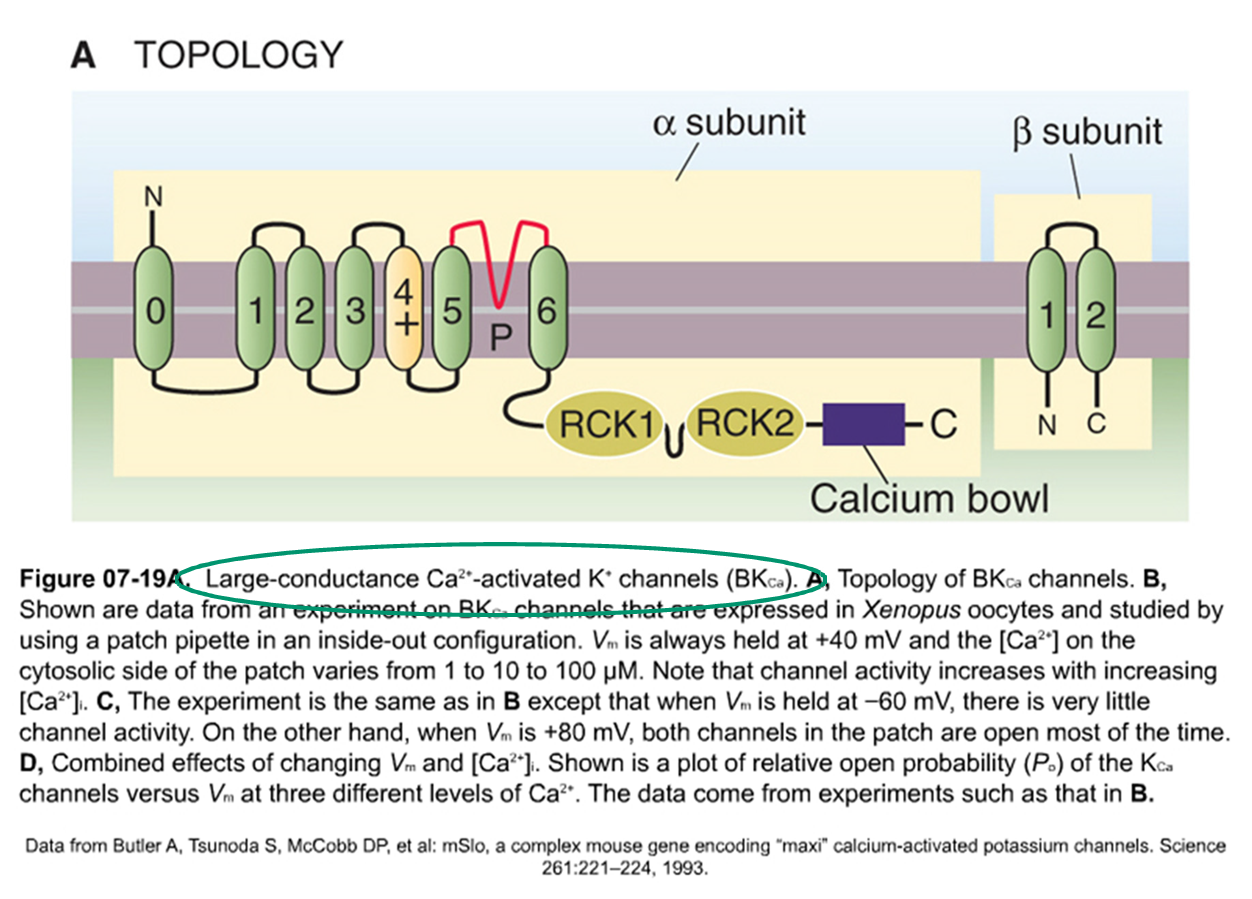
K+ - kanaal — large conductantie KCa kanalen (BKCa)
activatie door Ca
modulatie door voltage
K+ - kanaal — hyperpolarized-cyclic-nucleotide gated (HCN) kation kanalen
lagere PK/PNa ratio; reversal potential = ~ -30mV
= inwaartse stroom
omgekeerde voltage-afhankelijkheid: open bij hyperpolarizatie!!
voltage- en cAMP-gevoelig
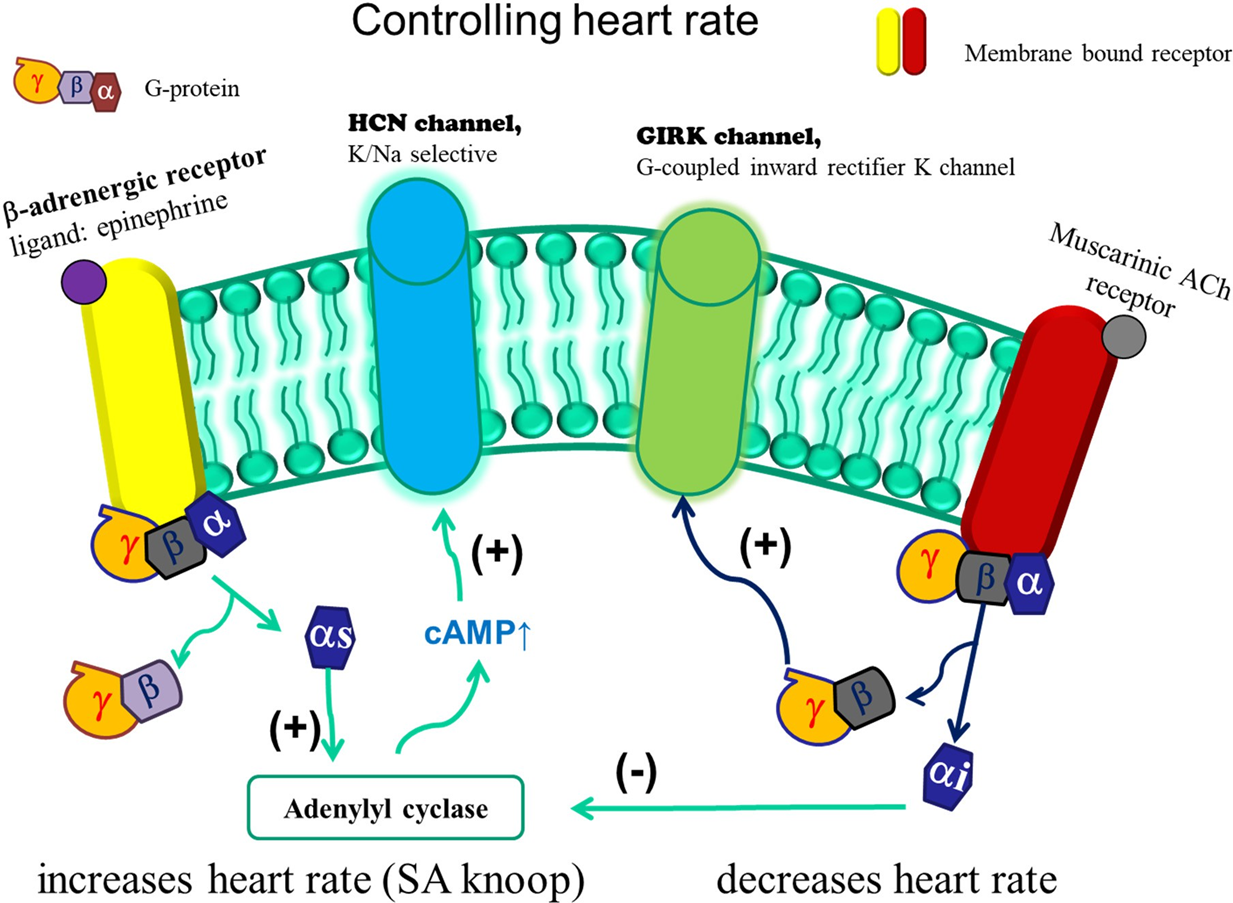
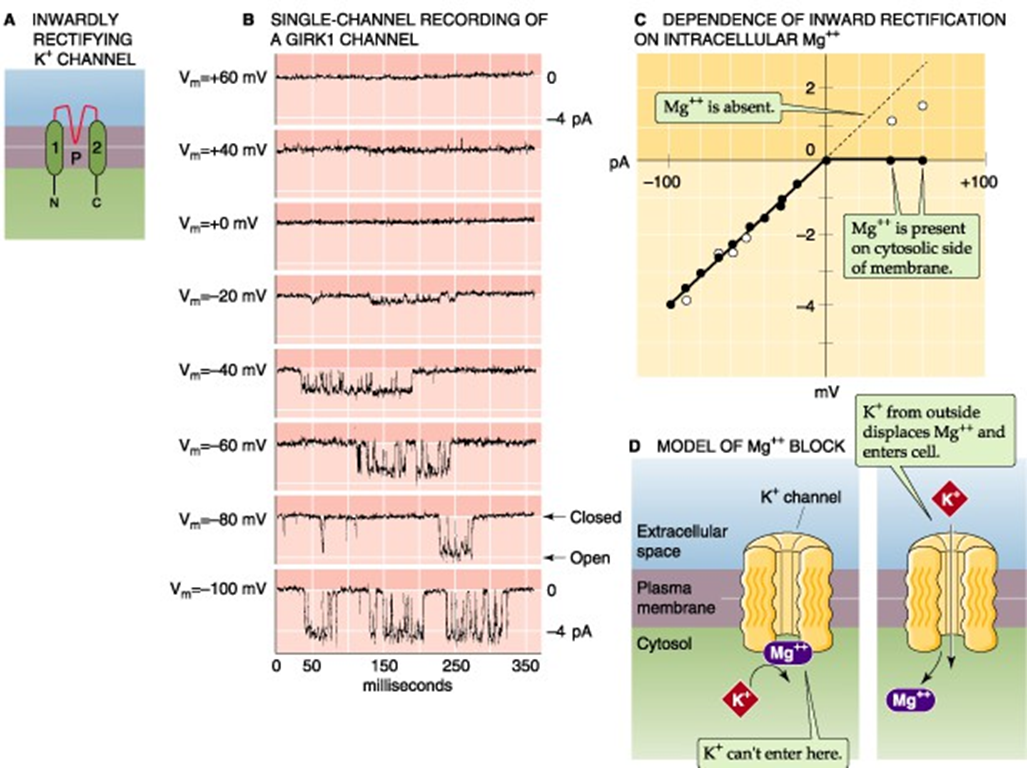
K+ - kanaal — inward rectifiers
vb. GIRK-kanaal
GPCR + inwardly rectifying + K+
2 Tm domeinen = “lekkanalen”
actief na mAChR stimulatie;
βγ-subunit + PIP₂
werking
theoretisch; enkel inwaartse stroom door Mg2+ blokkade
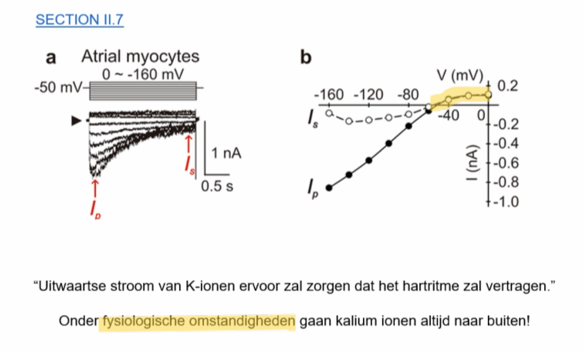
praktijk; altijd uitwaartse stroom (onder fysiologische omstandigheden) → verlaging hartslag
GIRK-kanaal — theorie
GIRK-kanaal — praktijk
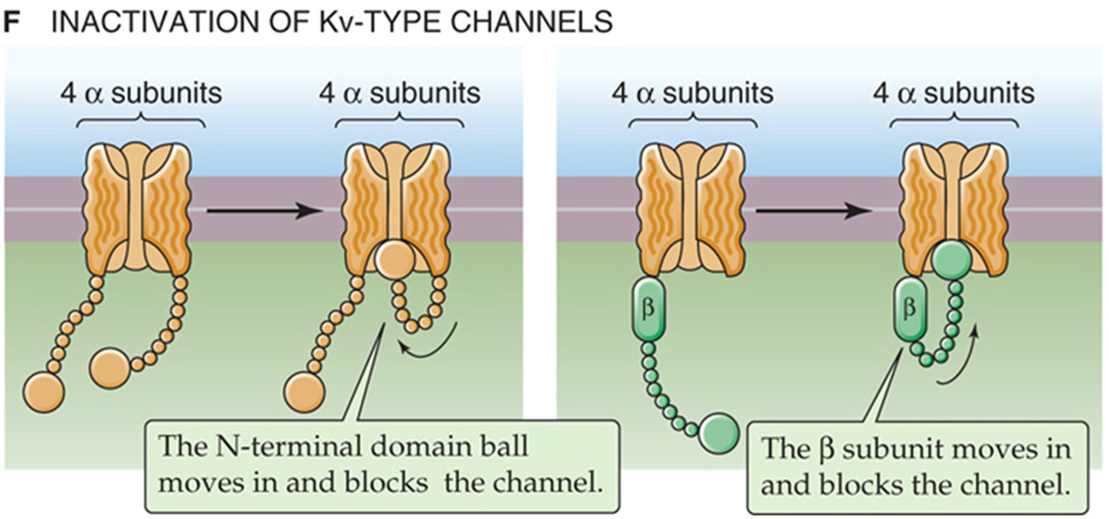
Kv - kanaal — types inactivatie
N-type inactivatie
β-subunit inactivatie
C-type inactivatie (niet op afb)
opm; dus ook bij sommige K-kanalen is inactivatie mogelijk, maar werkt anders dan bij Na-kanalen (h-poortjes)
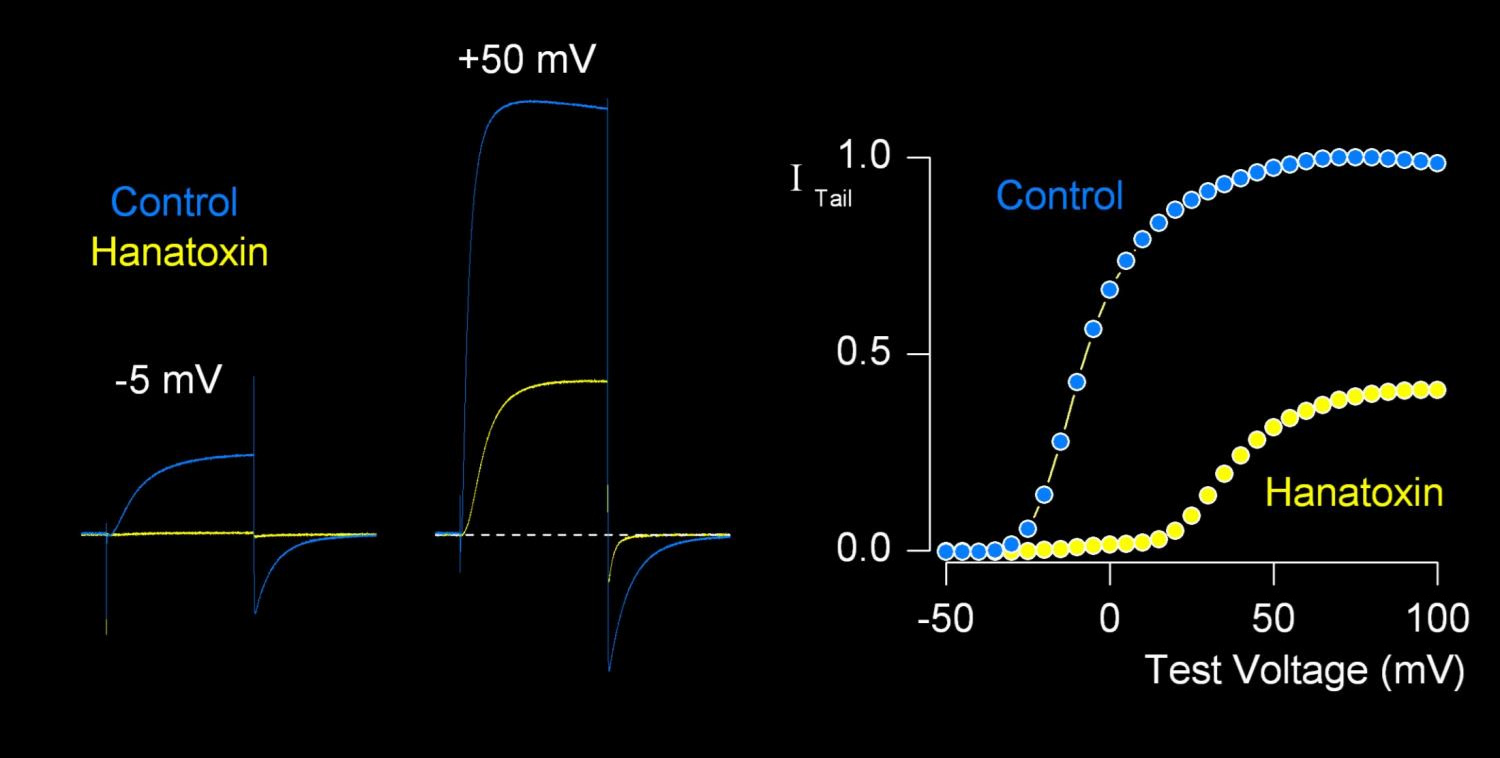
K+ - kanaal — toepassing Hanatoxine
inhibitie K-kanalen; duwen voltagesensoren naar beneden
minder effectief bij hoge voltages
K+ - kanaal — inward rectifiers
— toepassing hartslag
αi = inhiberend
αs = stimulerend