Lecture Exam 2
1/62
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
63 Terms
What are the key characteristics and types of Class Sarcopterygii?
Lobe-finned fishes
Have supportive lobes at the base of paired fins (before the ray-supported portion)
Most are extincy
Two living groups:
Actinistia (Coelacanths)
Dipnoans (Lungfishes)
What are the key skeletal and dental features of Class Sarcopterygii?
Scales and bones have an outer covering of true enamel (derived)
In Actinopterygians, the equivalent is ganoine
A key synapomorphy: enamel on teeth
What is the evolutionary significance of Class Sarcopterygii?
All members are more closely related to mammals and other tetrapods than to other fishes

What are the classification and extant species of Subclass Coelacanthimorpha?
Order: Coelacanthiformes
Family: Latimeriidae
2 living species (genus Latimeria):
Latimeria chalumnae - West Indian Ocean
Latimeria menadoensis - Indonesia


What is the fossil history and rediscovery significance of Subclass Coelacanthimorpha?
Well represented in the fossil record from the Devonian to Cretaceous (~300 million years ago)
Thought to be extinct for about 70 million years
Rediscovered in 1938 by Majorie Courtenay-Latimer

What are the key details about the first modern discovery of coelacanths?
First specimen caught in 1938 near the Comoros Islands (near Madagascar)
Depth: 260-300m
Species: Latimeria chalumnae
Described by J.L B. Smith
Story told in A Fish Caught in Time by Samantha Weinberg
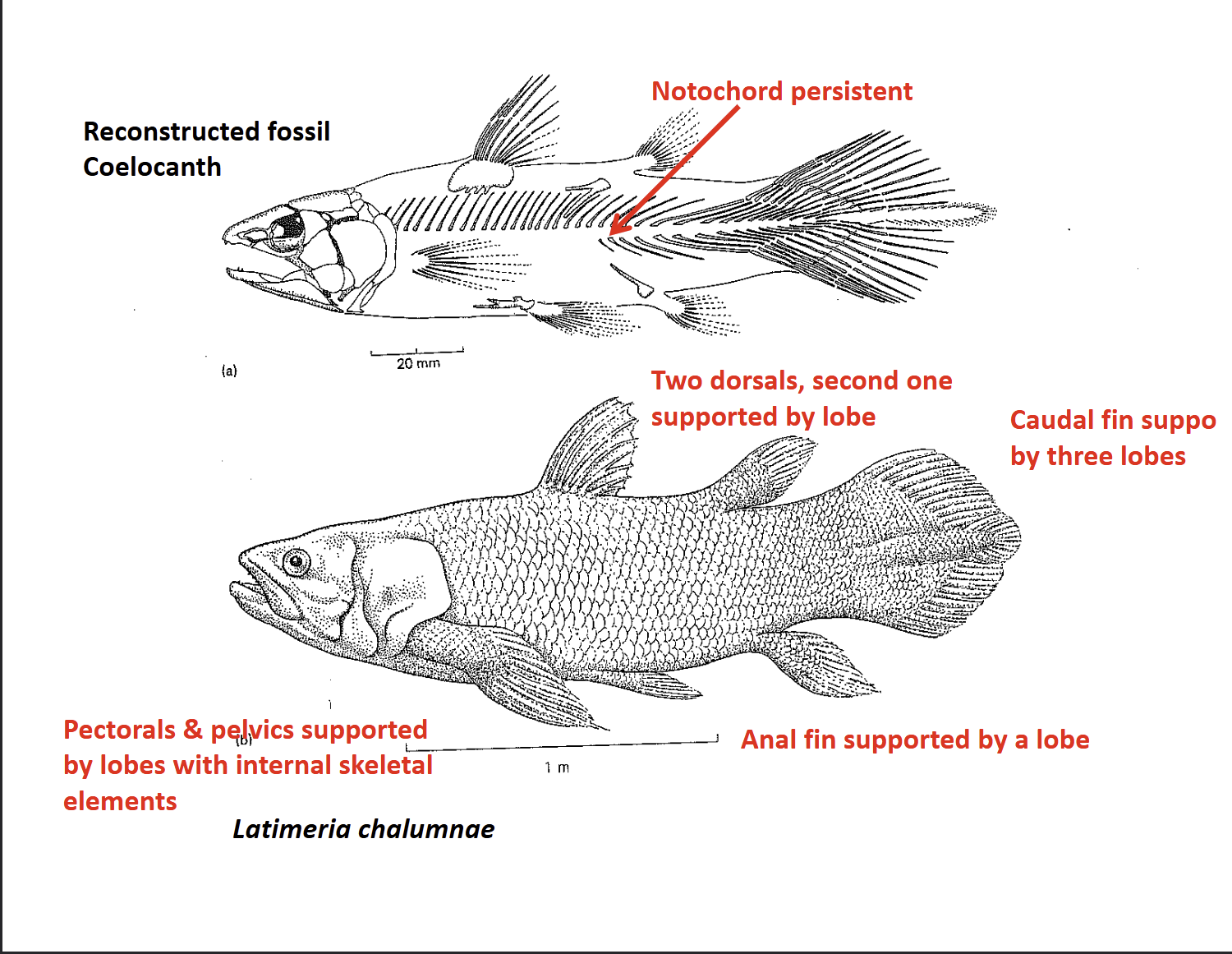
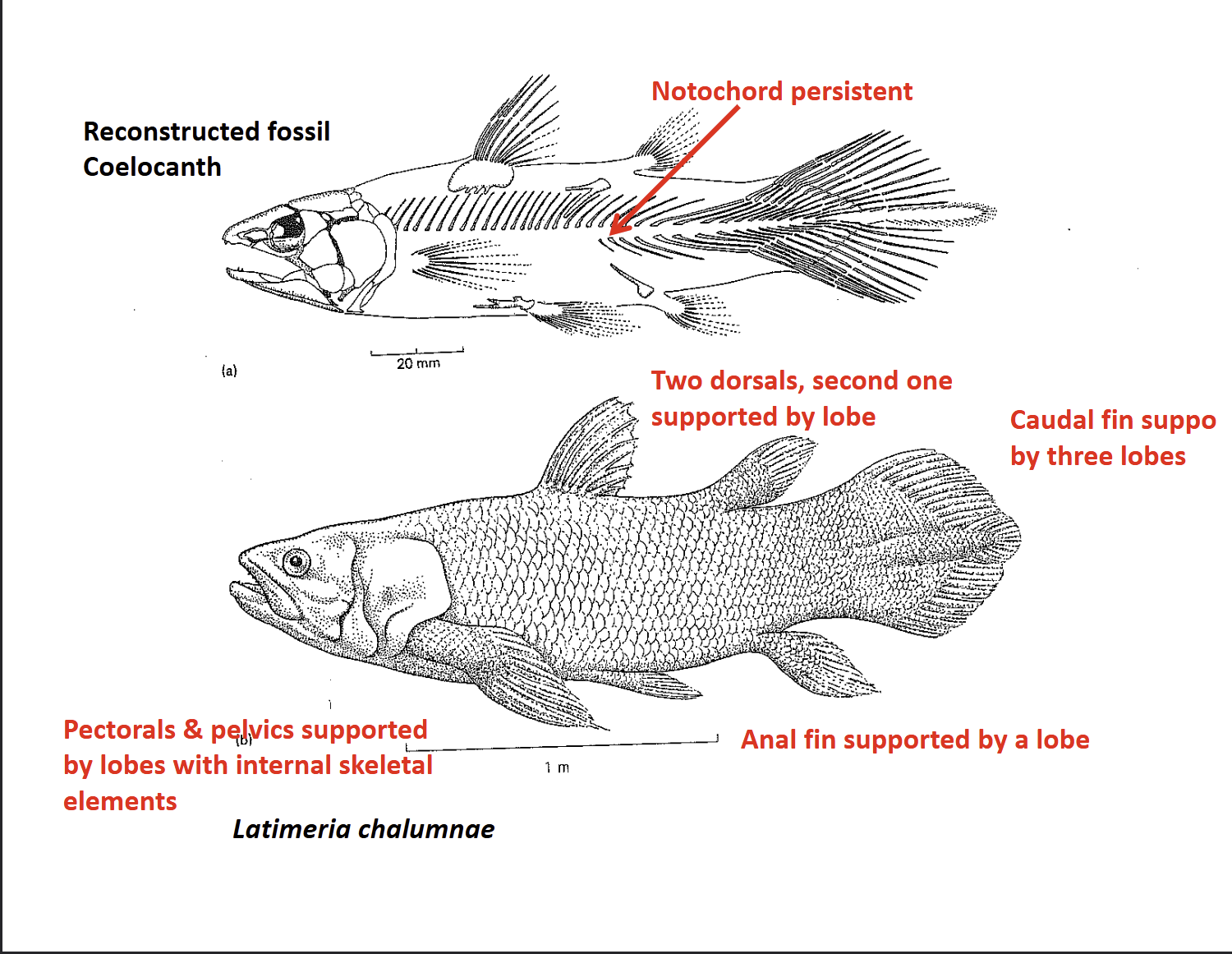
What are the key features of coelacanth fins?
Pelvic & pectoral fins: Strongly developed with internal skeletal elements homologous to tetrapods
Dorsal fins: Two (most bony fishes have one); second dorsal supported by a fleshy lobe
Anal fin: Supported by a lobe
Caual fin: Divided into three lobed sections
What are the swimming behaviors of coelacanths?
Capable of strong bursts using the caudal (tail) fin
Use paired fins to scull slowly and rotate
Can swim upside down
Exhibit “head-stand behavior” (vertical position, head down)
What are the brain and sensory features of coelacanths?
Extremely small brain: about 1/15,00 of body mass (smallest relative size among measured vertebrates)
Eyes adapted to low light: mostly rods in retina
Have a tapetum lucidum but lack melanin sacs, so they can’t adjust well to bright light
What is the rostral organ in coelacanths and its proposed functions?
Rostral organ: Cavity in the snout filled with gelatinous substance, with three openings on each side
Proposed functions:
Detect electrical signals from prey or predators
Sense magnetic fields for orientation at depth
Hans Fricke showed electrical pulses triggered head-stand behavior (rostrum pointed toward source)

What are the main conservation concerns for coelacanths (Latimeria chalumnae)?
Estimated population: ~200-500 individuals (hard to measure)
Threats:
Slow growth & late maturity
Limited habitat and range
Bycatch in fisheries
Ancient lineage (~400 million years old)
Key question: Can it survive the next 50 years?

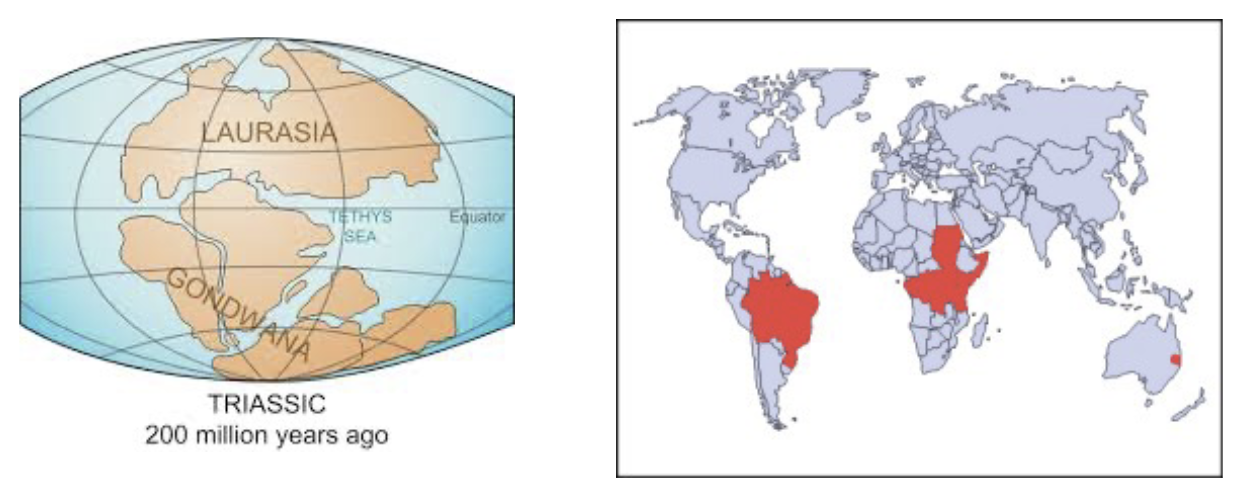
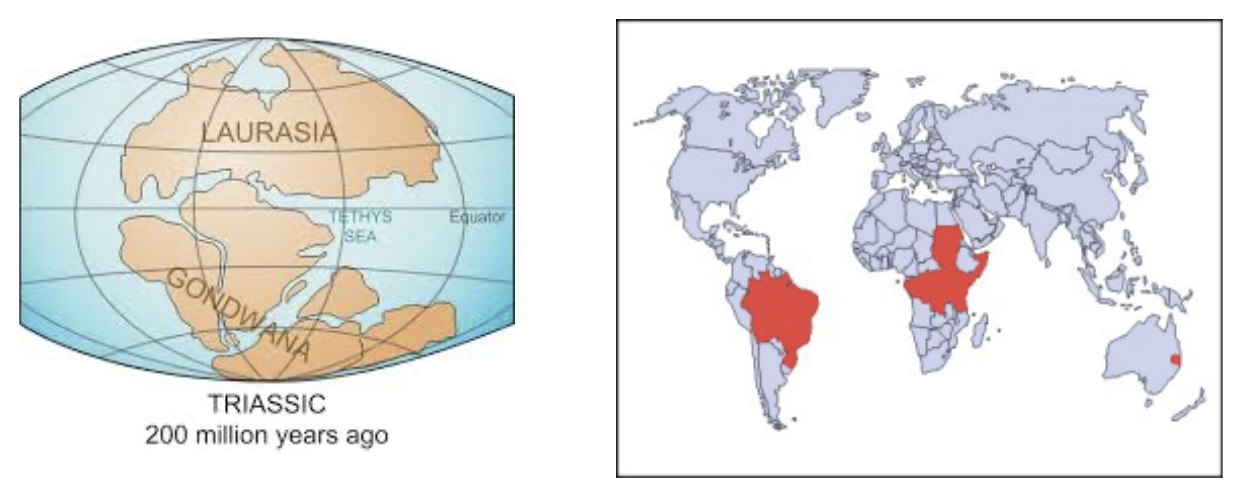
Extant Dipnoans - Lungfishes
Name means “double breath”, refers to presence of functional gills + functional lungs
Not ancestral to the first non-amniote tetrapods
Six species:
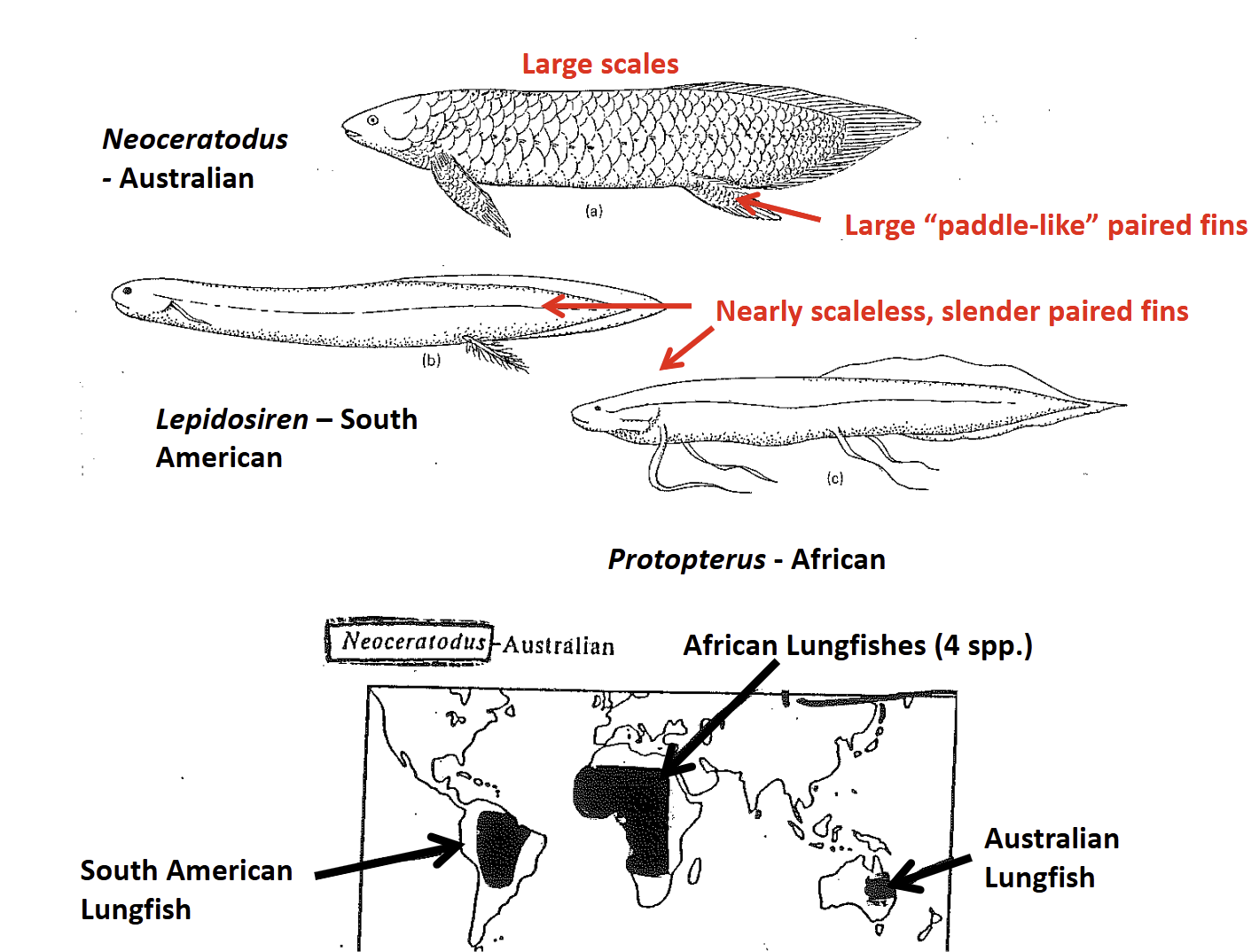
Protopterus spp, - 4 species: Africa
Lepidosiren paradoxa - South America
Neoceratodus foresteri - Australia
Extant Dipnoans have a Gondwanan distrubution

Synapomorphic Character States Extant Dipnoans - Lungfishes
Four characters that are different than Sarcopterygians that were ancestral to non-amniote tetrapods
Autostylic jaw support
fused tooth plates - durophagy
Maxilla & premaxila absent
Endoskeleton of paired fins has a central bony axis with radials on both sides - “biradial”
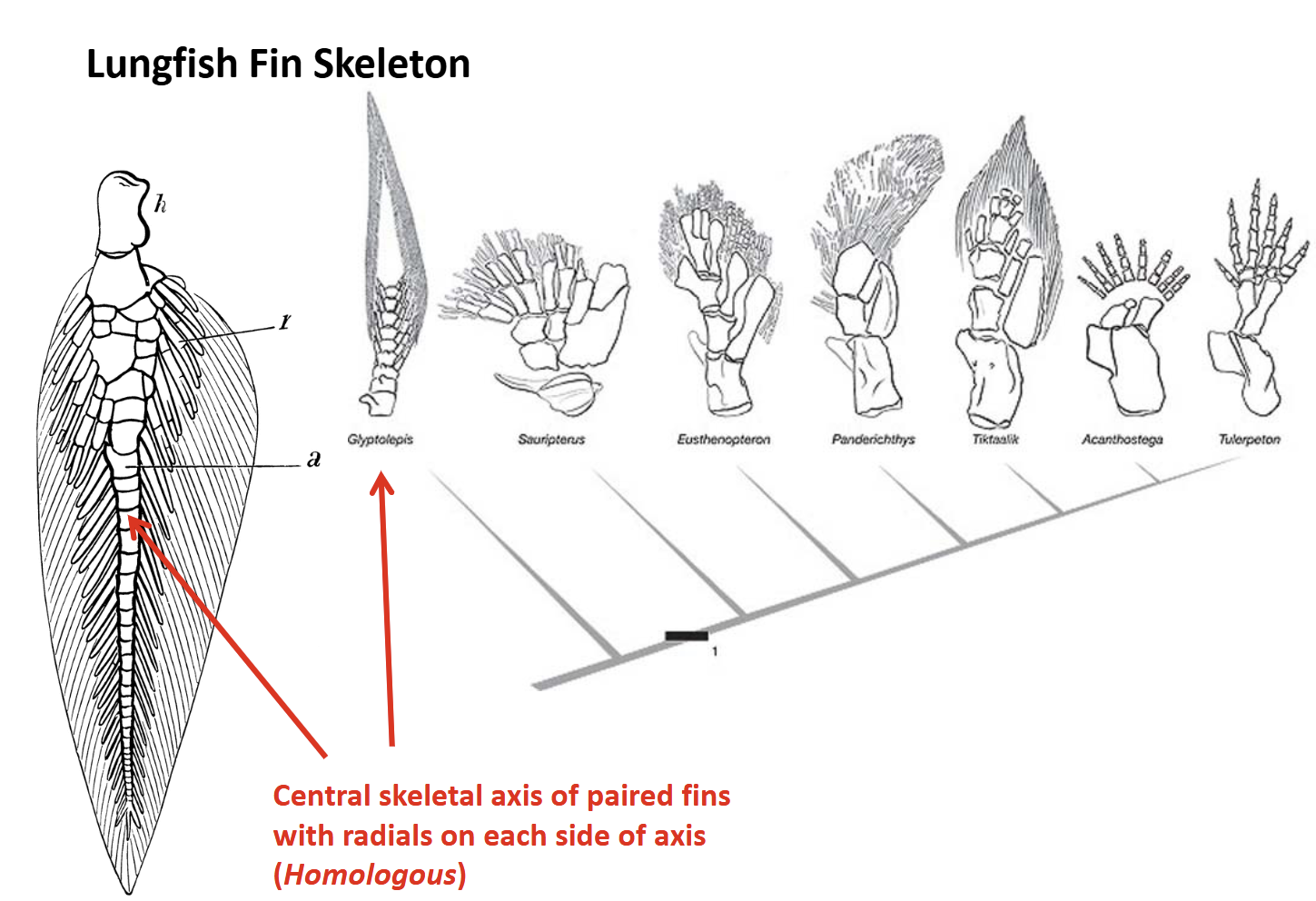
Australian Lungfish - Neoceratodus
Largest species (1.5m, 100 lbs), deep-bodied
Two pairs of large paired fins with large lobed bases
Scales are large & prominent
Diphycercal caudal fin
Dependent upon gills for gas exchange, gulps air into lungs to survive hypoxia (facultative air-breathing)
Stomach absent - spiral valve in intestine, feeds on benthic molluscs
Occurs only in the Burnett & Mary River drainages of Queensland, Australia
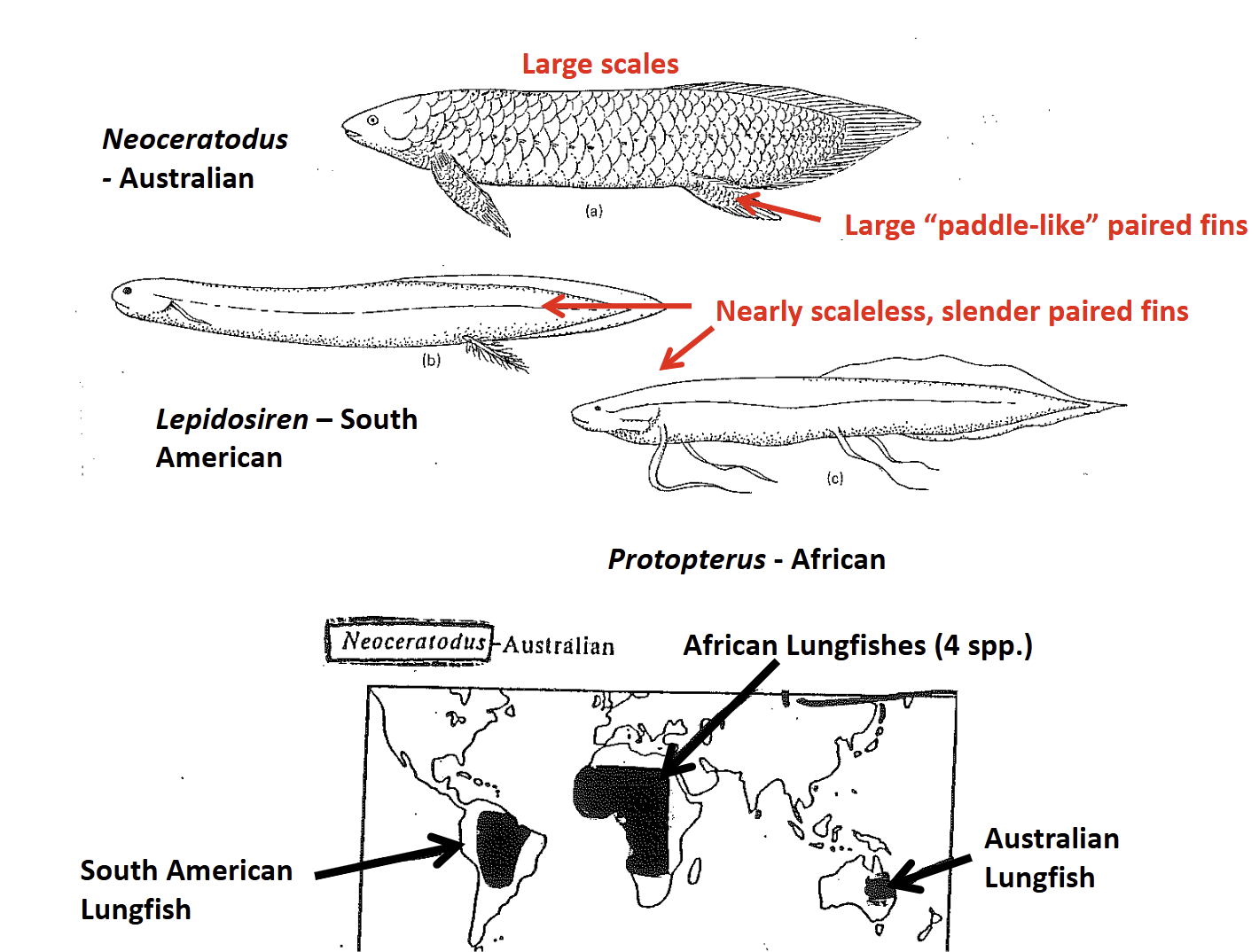
South American (Lepidosiren) & African (Protopterus) Lungfishes
Long (1-2m) slender bodies
Paired fins are very slender, elongate in Protopterus
Nearly scaleless
Skeleton is highly cartilaginous & flexible
Gills are poorly developed, dependent upon aerial gas exchange for 98% of their O2 acquisition (obligate air breathers)
Aestivate to survive drying up of fresh water pond habitats

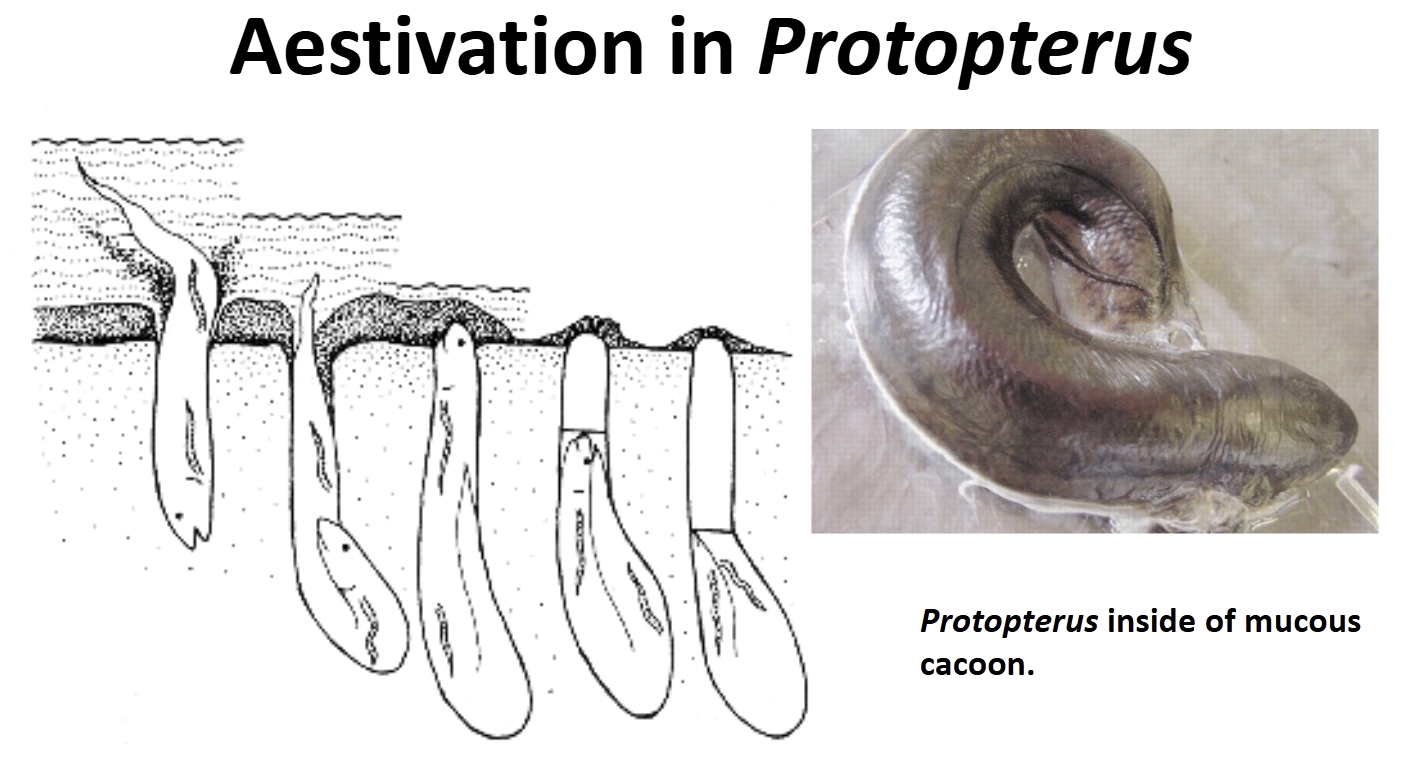
Aestivation in Protopterus
Induced by desiccation during the dry season
First crawls from pool to pool as they dry
Digs a 1m deep burrow with a round chamber at the bottom
As water dries, descends into the chamber, curls up body, secretes a thick mucous which dries into a “cocoon”
Breathes air through a small hole in the cacoon
Metabolism is very low, lives off energy stores
Usually aestivates for 6 mos. Have survived for 4 years in labratory studies
Evolutionary Ecology of the Water-Land Transition in Vertebrates
Transition occured during the Devonian, 400 - 375 mya
First vascular plants colonized terrestial habitats
Arthropods followed
Severe droughts occured, FW bodies dried/shrank
Fish density increased
Competition intensity increased
Predation pressure increased
Water temperature increased
Dissolved O2 content of water decreased
Shallow-water provided ecological release for fishes that meet the challenges posed by moving into these micro-habitats
Challenges Posed by Shallow Aquatic Habitats & the Adaptive Solutions (1)
Decreased dissolved oxygen content - gas exchange using gills alone not sufficient; aerial gas exchange using physostomous G.B./ Lungs
Shallow water - less support of body mass against gravity. Density of abrasive vegetation increasing;
Swimming locomotion less effective
Crawling/ paddling locomotion more effective
Strong selection for change in body shape, limb structure, & endoskeleton
Structural adaptations posed by shallow aquatic habitats
Food organisms different, conditions for prey capture different, inertial feeding less effective, suction feeding less effective
Reduction of fins, changes in the mouth to take advantage of new prey (Arthropods)
Shallow aquatic habitats are more abrasive, air exposure increased, increasing desiccation stress
Evolution of a more desiccation resistant integument

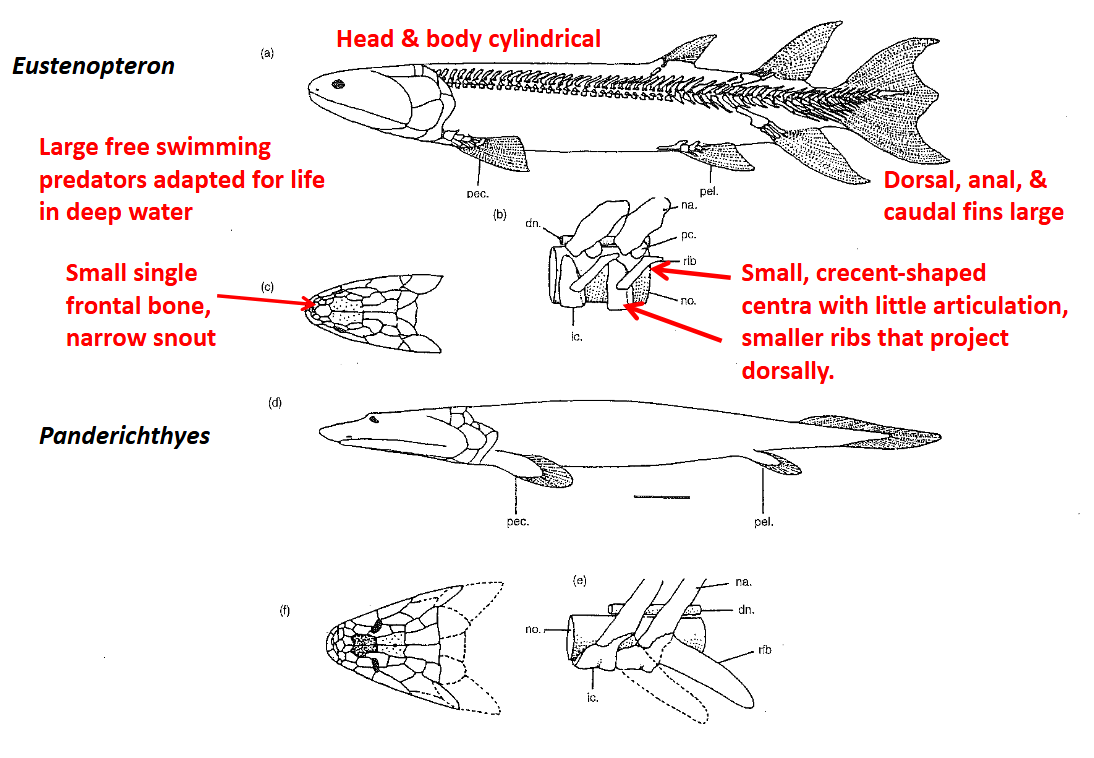
Eusthenopteron (Order Osteolepiformes) Symplesiomorphic traits:
Large, free-swimming predators adapted for life in deep waters
Cylindrical bodies, head not flattened, large (deep) dorsal, anal, & caudal fins
Laterally oriented eyes
Labyrinthine teeth; strong but not flexible
Single frontal bone, snout relatively narrow
Small, paired, crescent-shaped vertebral centra, no articulation
Ribs short, project dorsally (more support from surrounding water)

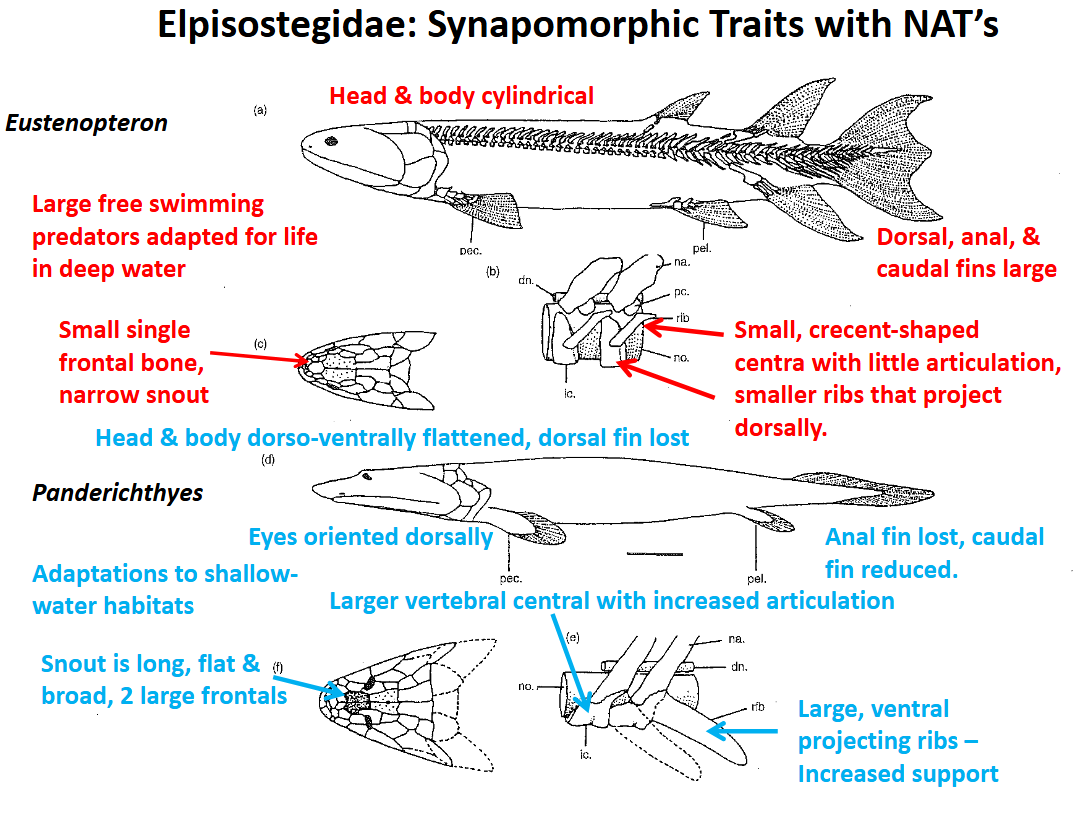
Panderichthyes (Family Elpistostegidae)
Sister group of the non-amniote tetrapoda
Synapomorphic traits with the first non-amniote tetrapoda:
Dorsal & anal fins lost
Caudal fin greately reduced
Body & head dorsoventrally flattened
Snout is long, flat, & broad (2 large frontals)
Eyes are dorsally oriented
Vertebral centra are larger & articulation is increased
Ribs are larger & project laterally & ventrally (increased support against gravity)

Tiktaalik roseae (“fishapod”) Fossil History
The next intermediate for between aquatic & terrestrial vertebrates
Fossils are 375 my old (end of Devonian), from 700 miles above the current Arctic circle
Tiktaalik fossil has high quality preservation of the head & neck region showing important details of braincase, palate,and gill arches


Tiktaalik roseae (“fishapod”) Derived Character States
Braincase was more solid than that of fishes
Cranio-vertebral column articulation was more flexible than in fishes
In ancestral fishes that live in deep waters, selection favors an inflexible cranio-vertebral column articulation for two reasons:
Streamlining
Neutrally buoyant predators can readily change direction by moving the entire body
Proximal portion of pectoral fins were adapted to bear the weight of the anterior body, having distinct wrist bones
Ribs are long & robust for support in very shallow water


Tiktaalik roseae (“fishapod”) Adaptations
When crawling along the bottom in shallow water, cannot easily raise the body to change direction to strike prey. Increased selection pressure of indpendent head movement.
Head mobility was increased because bony articulation between head and gill region was broken. Involed loss of the opercular bones and extreme reduction in the hyomandibula (takes on a new function of sound transmission as an ear ossicle)


Tiktaalik roseae (“fishapod”) Ancestral Characters (symplesiomorphic)
Functional gills present
Dermal scales present
Cranium & mouth large & wide
Pelvic appendages were relatively small and fin-like
Distal portion of pectoral appendages still membranous fin-like structures

First Tetrapoda: Classification
Not correct systematically or ecologically to label first tetrapods “amphibians”
They are not closely related to extant Caudata (salamanders), Anura (frogs), or Apoda (caecilians)
More closely related to extant amniotes
To clarify, extant “amphibia” are correctly labelled Lissamphibia = the extant tetrapods that do not lay shelled (amniote) eggs
NAT’s to describe first tetrapods
First Tetrapoda: Structure
The first tetrapods of the Paleozoic were much different than Lissamphibians in several ways
Much larger (1-2m)
Integument had dermal scales
Did not exchange gases over integument
Similar to extant crocodilians in both appearance & behavior, living in shallow waters aquatic microhabitats, later riparian microhabitat
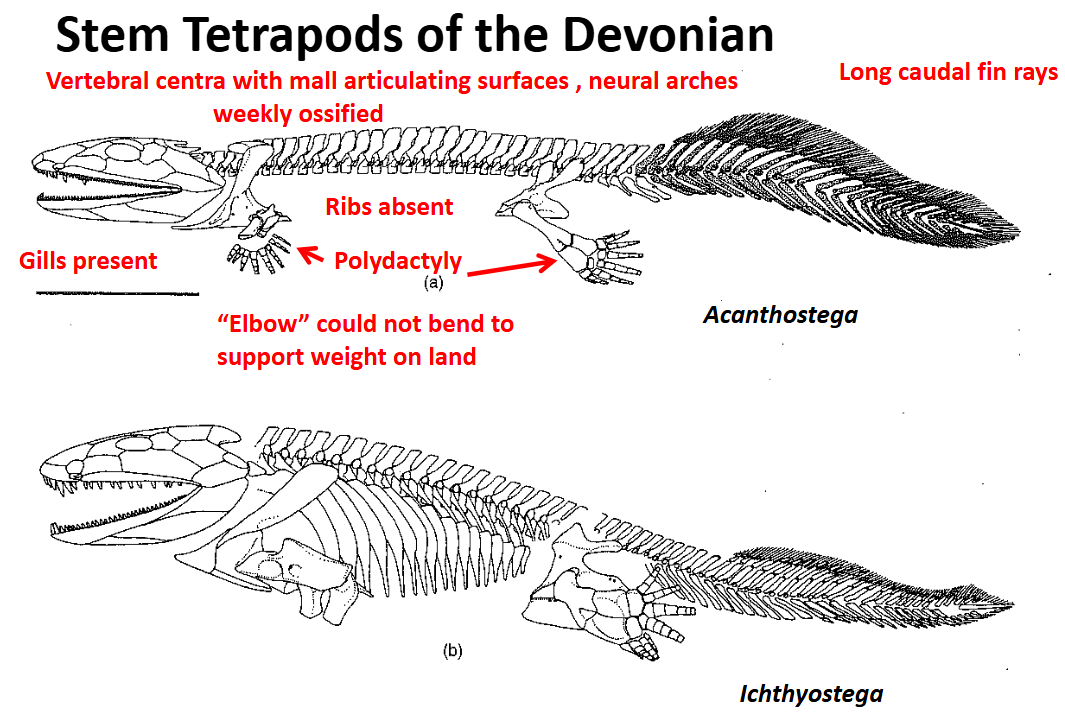
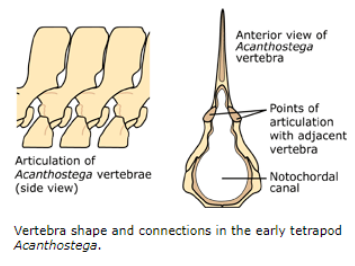
Stem Tetrapods of the Devonian: Acanthostega
Highly aquatic - evidence:
Forelimbs & hindlimbs were polydactylous = more than 5 digits
Forelimbs were very paddle-like, hindlimbs less so
Had a flange on the cleithrum that articulated with the operculum - indicate internal gills that functioned for gas exchange
“Elbow” was not able to bend in a way that would support the body weight on land
Articulating surfaces of the vertebrae were (less supportive)
Neural arches were weakly ossified
Fin rays of the caudal fin were long
Well developed ribs absent

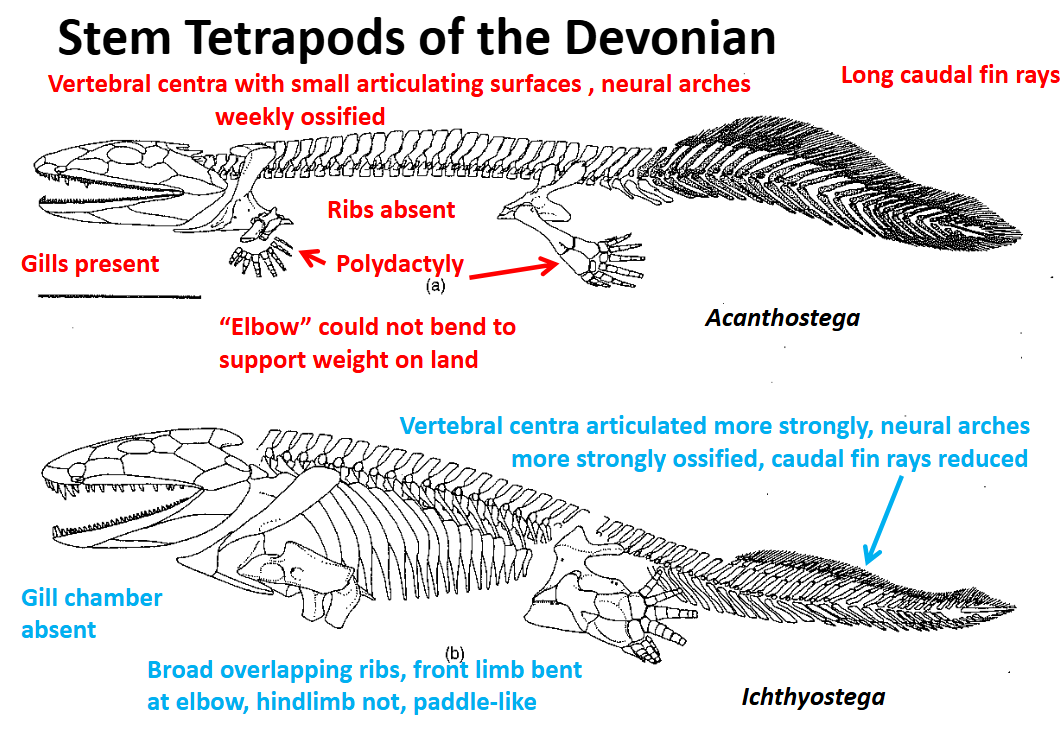
Stem Tetrapods of the Devonian: Ichthyostega
Appears to have been less aquatic - evidence:
Flange on cleithrum is absent suggesting that functional gill chamber was absent
Caudal fin rays were reduced
Had broad, overlapping ribs - support against gravity & muscular attachment, protection of viscera
Hindlimb polydactylous, but less paddle like (forelimb structure unknown)
Proportinos of the humerus & femur are similar to those of some pinnipeds (sea lions)
Forelimb was permantely bent at the “elbow”
Hindlimb was more paddle-like
Suggests Ichthyostega coulid better support weight on land, but still strong swimmer

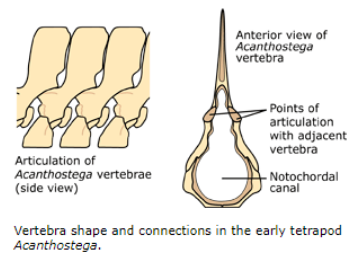
Gravity and Vertebrae
Fishes have simple lateral motion and support from water, so:
Arches are relatively straight and needle-like
So are ribs
On land, a quadruped backbone faces same problem a bridge designer would face - sag!
Evolved a series of interlocking articulations on each vertebra
Helped overcome sag and hold backbone straight and minimal muscle effort
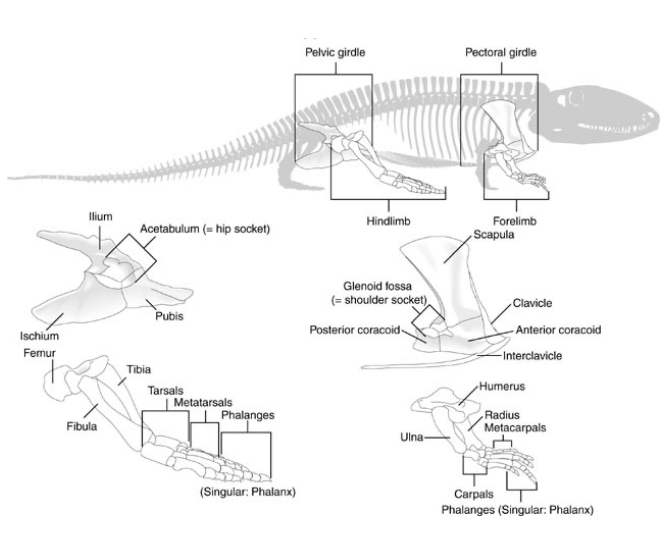
Evolution of the Girdle Skeletons Ancestral Condition (fishes)
Neither the pectoral or pelvic girdles articulate with the vertebral column
Evolution of the Girdle Skeletons Tetrapods
Pectoral Girdle - throughout tetrapod evolution, the pectoral girdle never directly articulates with the vertebral column via skeletal articulation like the pelvic girdle does
Pectoral girdle is held in position by skeletal muscles & connective tissue
Evolution of the Girdle Skeletons Pectoral Girdle
Consists of five paired elements:
Scapula - the dorsal element
Coracoid - one ventral element
Clavicle - a second ventral element when present
Clethrum & interclavicle - are smaller portions of the girdle that are remnants from ancestral fishes. Both elements reduced, and sometimes absent entirely
The articulating surface for the humerus is the glenoid fossa
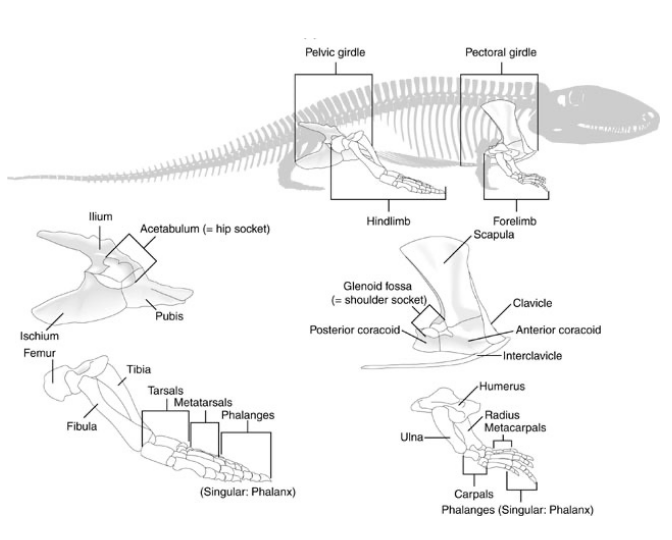
Evolution of the Girdle Skeletons Pelvic Girdle
Beginning with the first non-amniote tetrapoda, the bones of the pelvic girdle become larger and more robust to transmit the weight of the body ventrally through the hind limbs
The pelvic girdle consists of three paired skeletal elements that form a “triangle” when views from the side
Evolutionary Changes of Non-Amniote Tetrapods Observable in Lissamphibians
Endoskeleton
Integument
Circulatory system
Gas exchange system
Mouth/Tongue structure & function
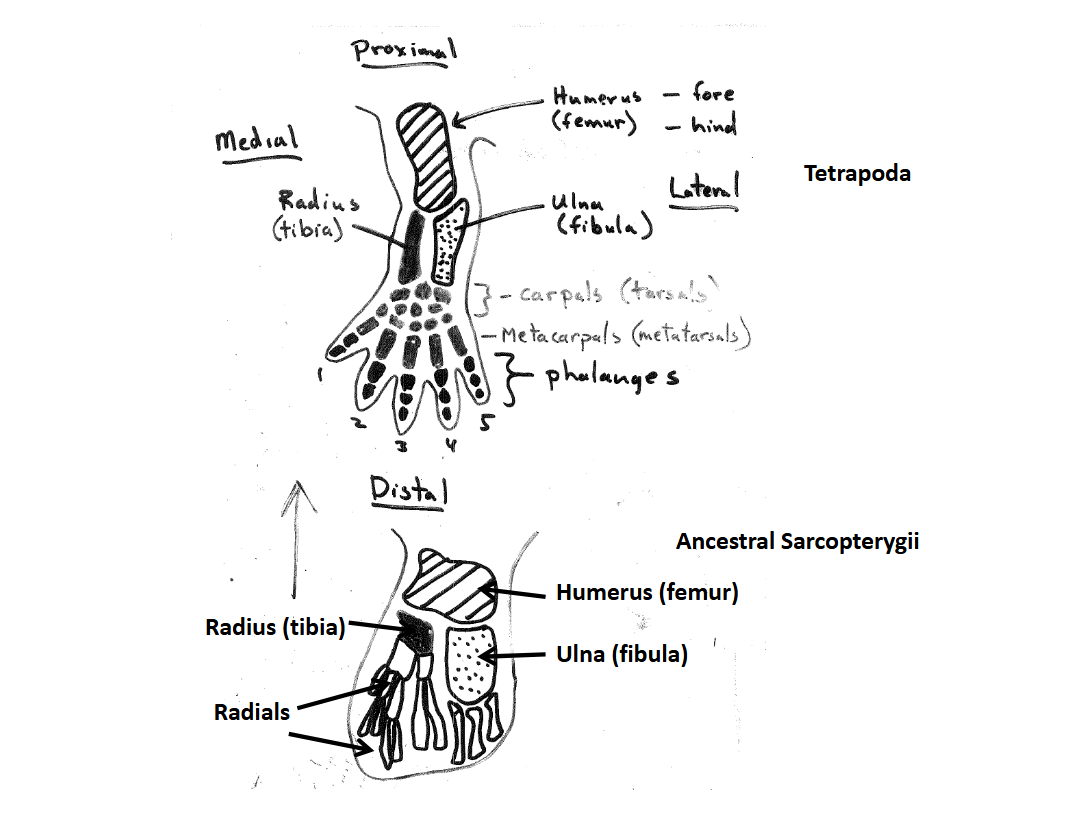
Tetrapod Limb Skeletal Elements
Homologous with those in the pectoral & pelvic fin lobes of the ancestral Sarcopterygians
Forelimb Elements
Humerus - proximal most element that articulates with the pectoral girdle at the glenoid fossa
Ulna - lateral element of the lower forelimb
Radius - medial element of the lower forelimb
Carpals - elements of the “wrist”
Metacarpals - elements in the hand that articulate with the carpals
Phalanges - elements of the digits
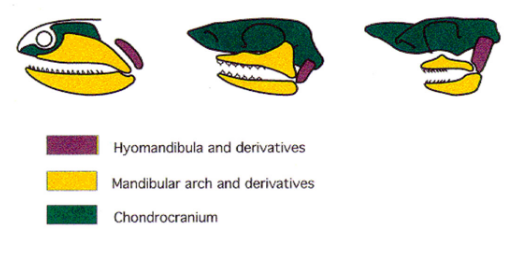
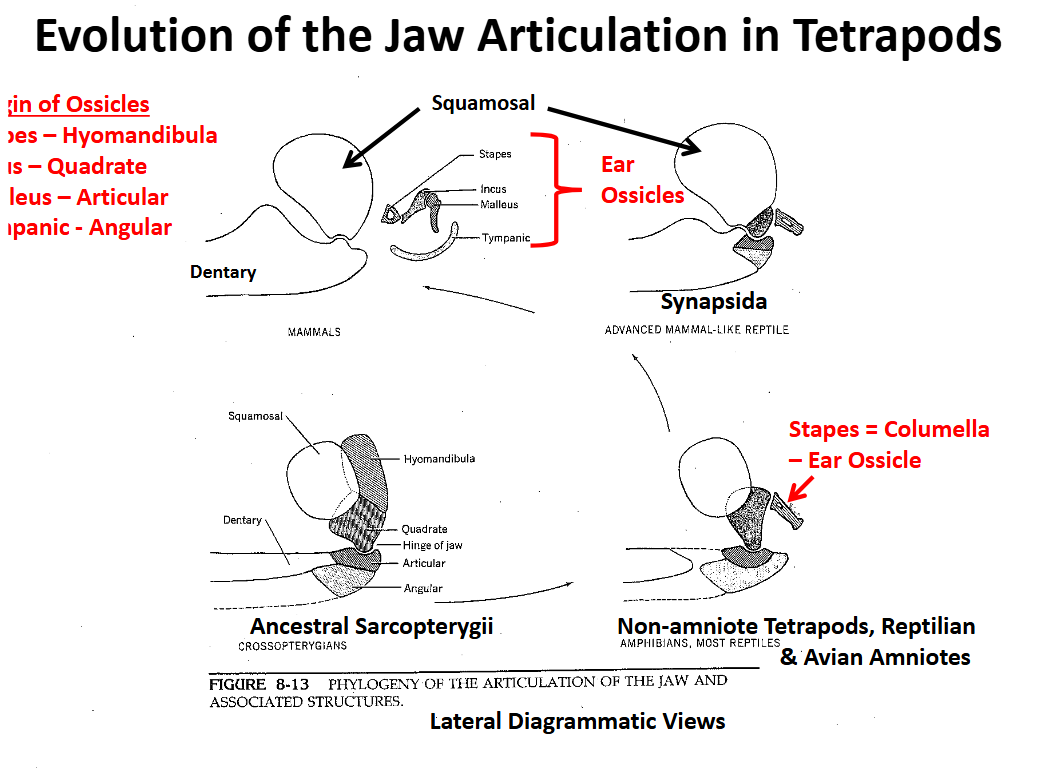
Evolution of the Tetrapod Jaw Articulation Ancestral Condition Sarcopterygii
The quadrate of the upper jaw articulates with the articular of the lower jaw. The hyomandibula supported the articulation, especially the quadrate
The angular supported the articular
The dentary + other elements formed the anterior lower jaw
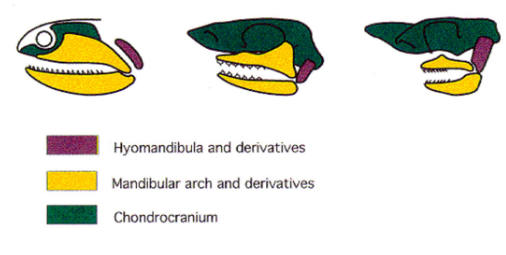
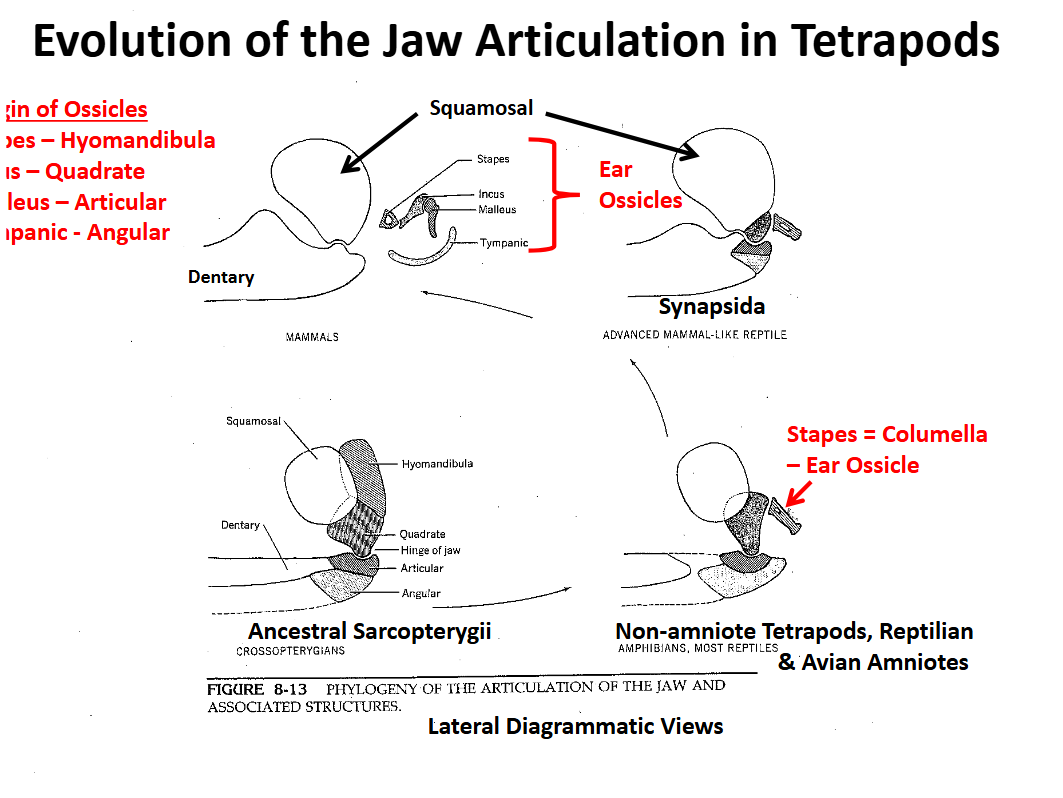
Evolution of the Tetrapod Jaw Articulation Non-Amniote Tetrapods & “Reptilian” & Avian Amniotes
The hyomandibula ceases to support the jaw articulation between the qudrate & articular
The hyomandibula is reduced to a small, slender cylindrical element (stapes = columella) that transmits sound waves from the tympanic membrane to the inner ear
Sound wave transmission is increased in air
Jaw articulation is qudrate-articular
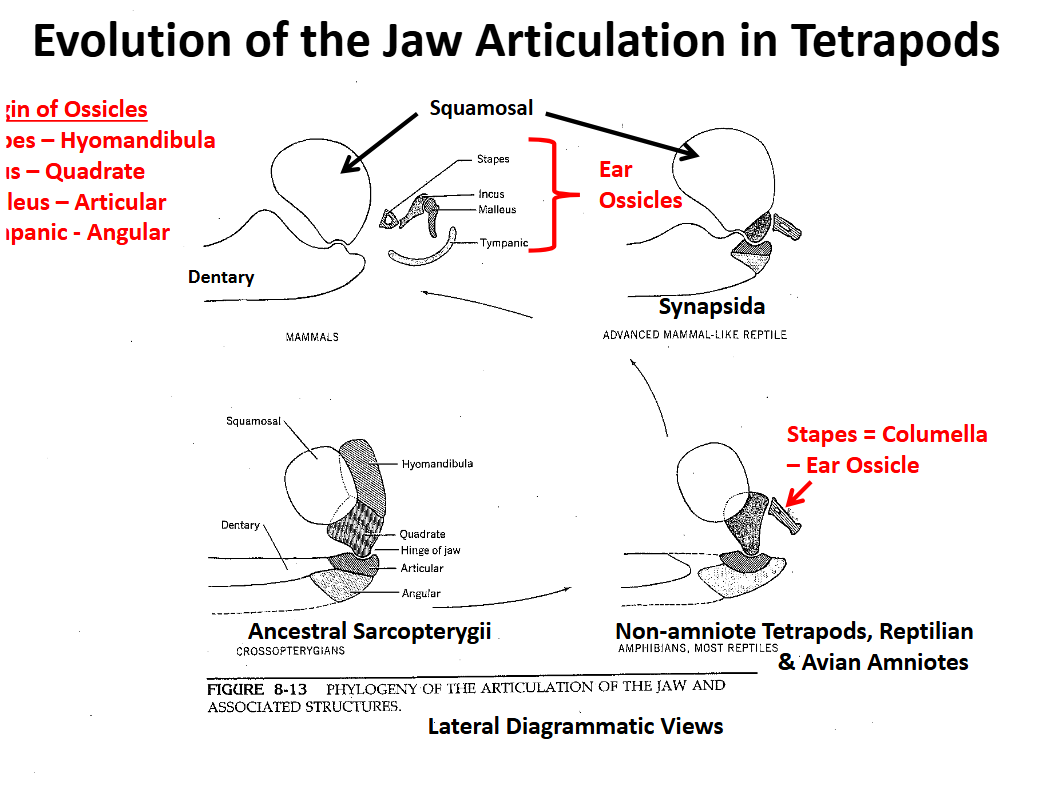
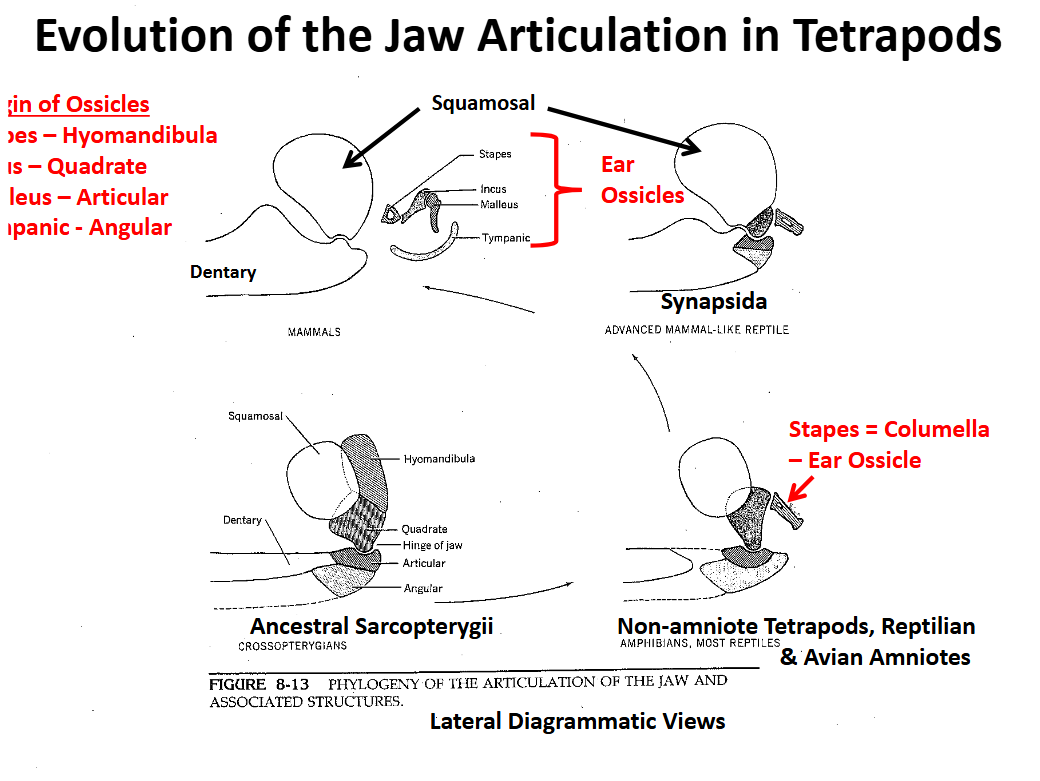
Evolution of the Tetrapod Jaw Articulation Synapsida
Squamosal (upper) & dentary (lower) elements become involved in the jaw articulation
Quadrate, articular & angular are reduced, migrate posteriorly
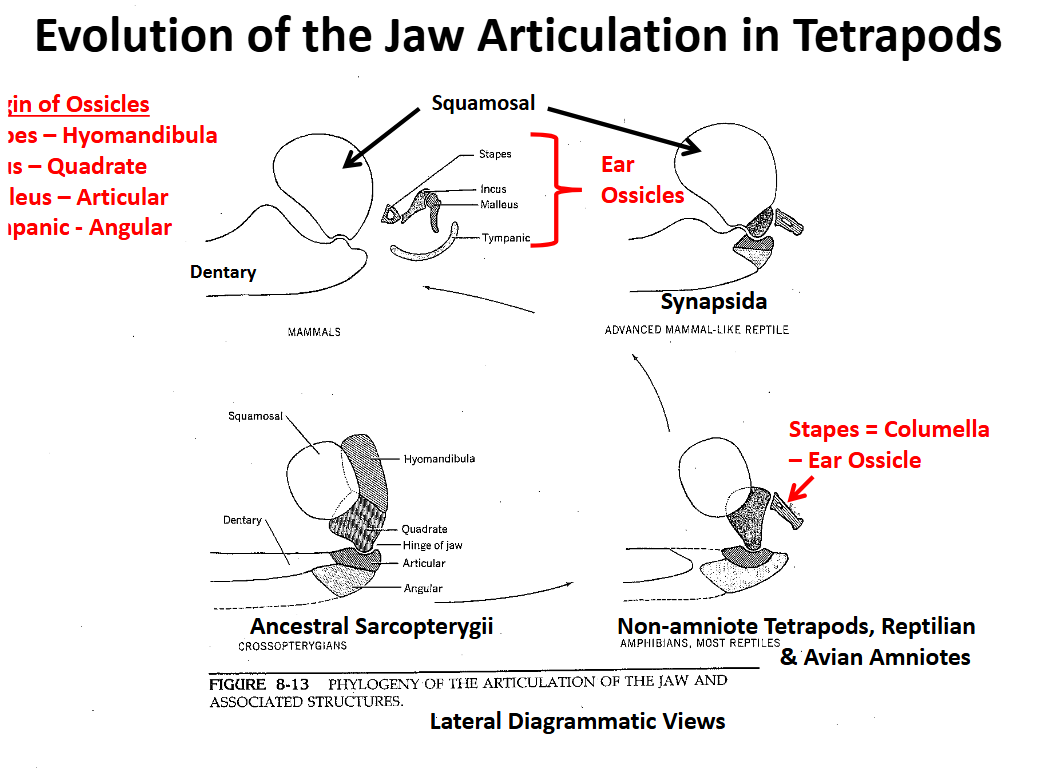
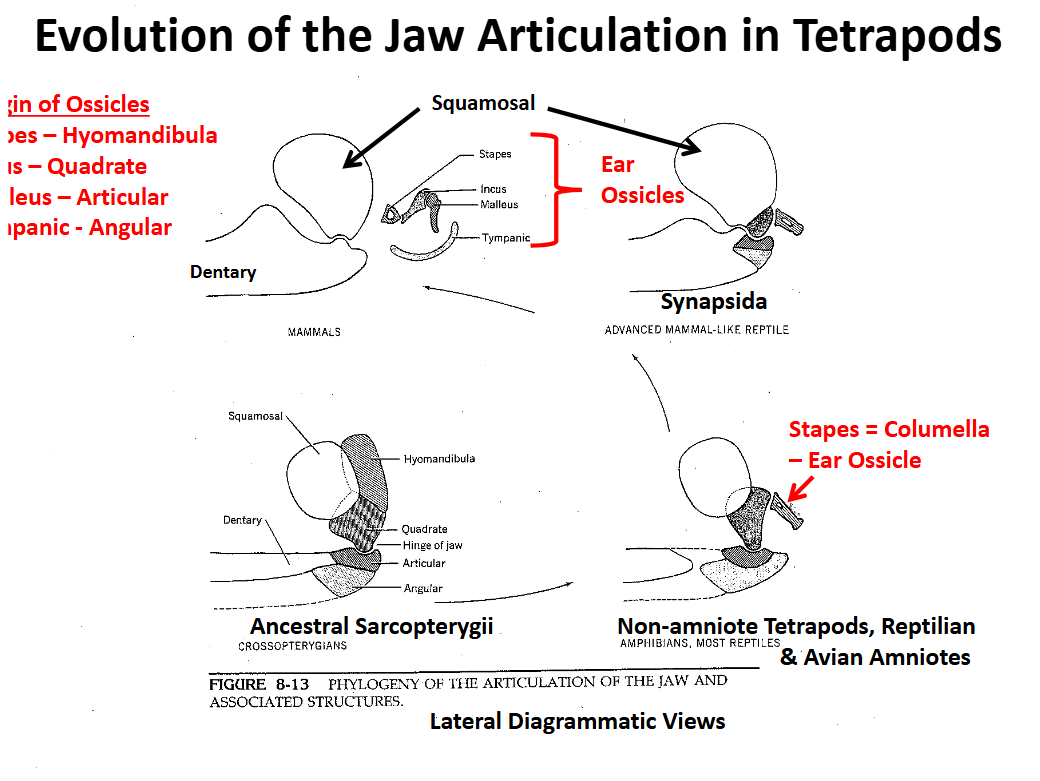
Evolution of the Tetrapod Jaw Articulation Mamalia
Jaw articulation is completely between the squamosal and the dentary
Articulation involving two large bones is stronger
Other bones become involved in sound transmission
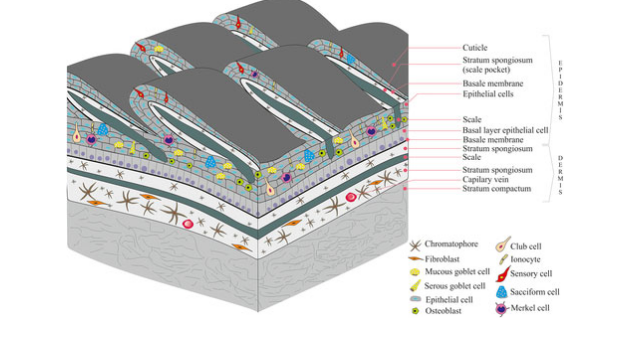
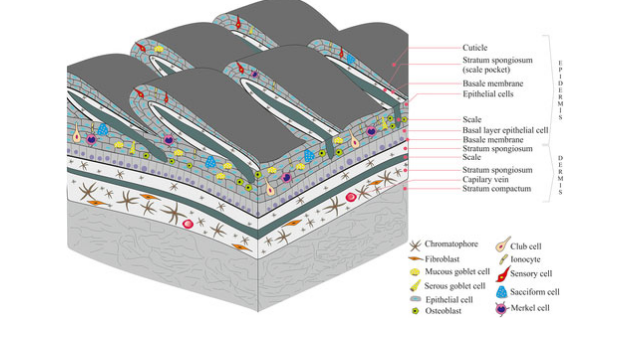
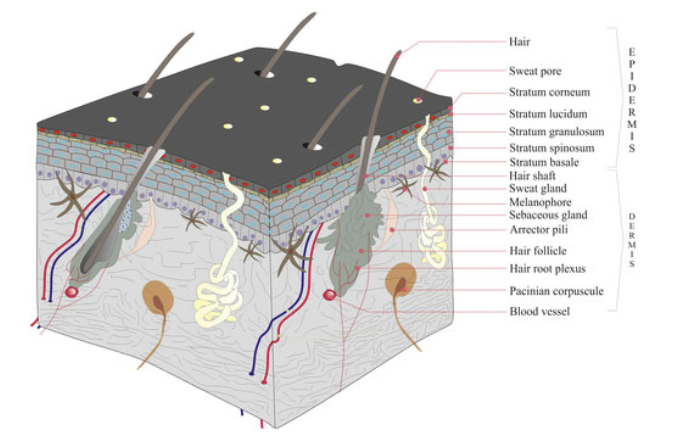
Evolution of the Tetrapod Integument Ancestral Condition Sarcopterygii
Dermis is the “dominant” layer (dermal armor & scales)
Epidermis is a relatively thin outer layer with numerous unicellular mucous glands
In fishes, the mucous functions to reduce resistance while swimming & protect against pathogens
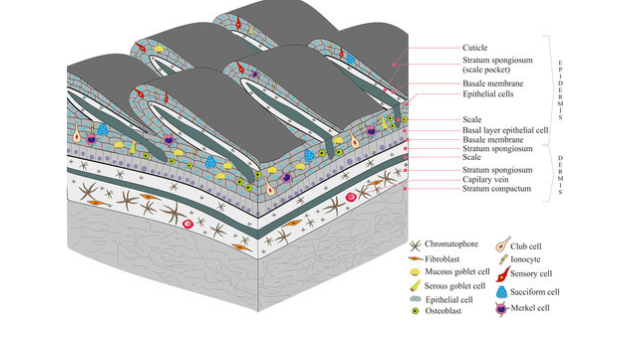
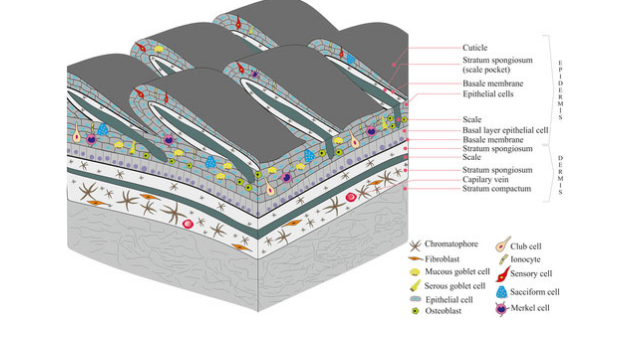
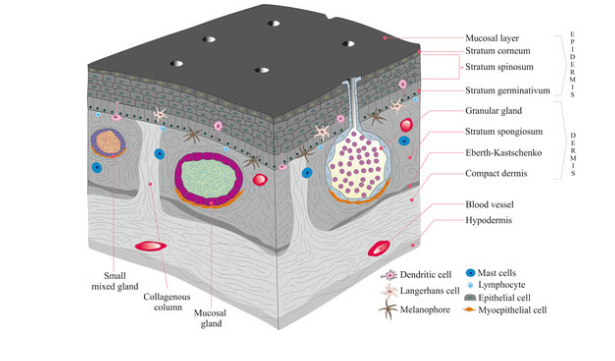
Evolution of the Tetrapod Integument Derived Condition Non-amniote & amniote tetrapods
Epidermis becomes much more prominent
New epidermal cells are produced in mitotically active cells adjacent to the dermis
Cells migrate toward the body surface as they age
Produce keratin & become progressively flattened
Keratin is a tough, evaporation resistant protein
Reduces evaporation of water from deeper cells, and protects against abrasion
Integument in Extant Lissamphibia
Thickest in terrestial forms (e.g. Bufonidae)
Thin in aquatic adults & larvae
Keratin does not stretch. Therefore, eventually limits growth
Ecdysis - to accomodate growth, periodically a new keratinous layer produced beneath the old one which is sloughed off
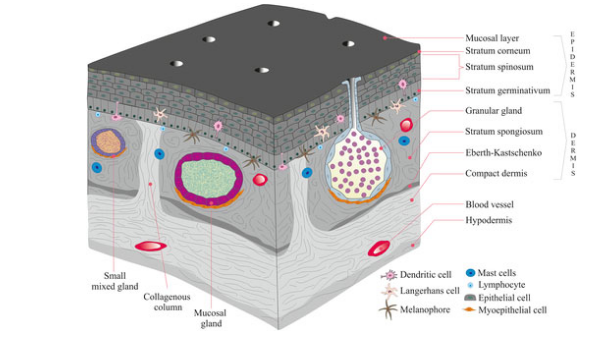
Integument in Extant Lissamphibia Glands & Gas Exchange
Multicellular mucous glands - secrete mucous that protects from pathogens, & deters desiccation, so long as it is kept moist (limits to moist microhabitats)
Specialized mucous glands - in some species, glands are specialized to produce toxins that deter predators (dendrobatids), or secretions that function in social communication
Integumentary Gas Exchange - moist mucous layer allows O2 to go into solution
on the surface of the integument & diffuse into capillaries
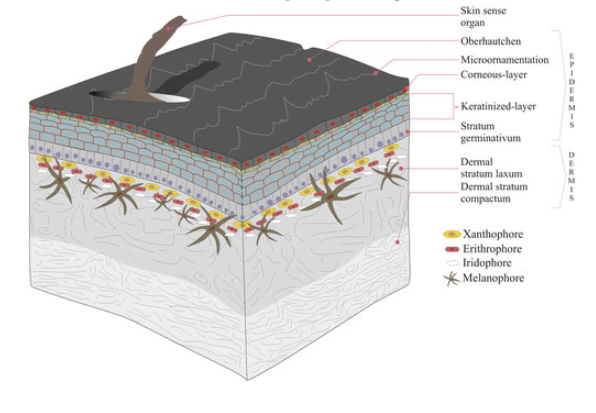
Integument in Amniote Tetrapods “Reptilian” Amniotes
Becomes thicker & more complete (limits O2 exchange)
Provides increased protection from desiccation stress, important for success in terrestial habitats
Forms specialized hard structures: e.g. claws, spines, coverings of horns, rattles, rhamphotheca (beak), egg-teeth (sharp projection on bill when first born)
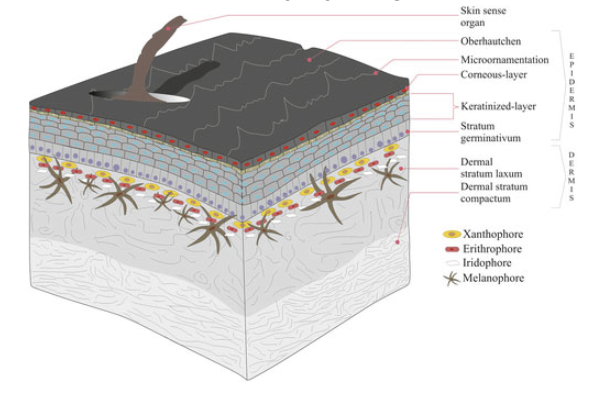
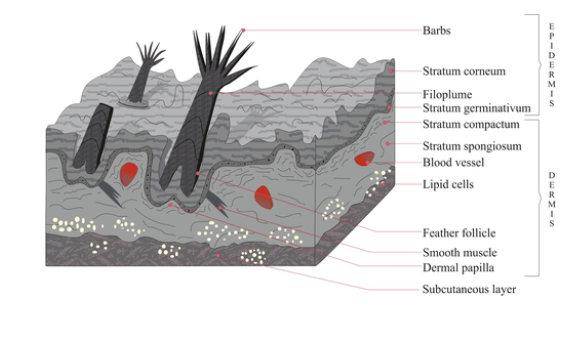
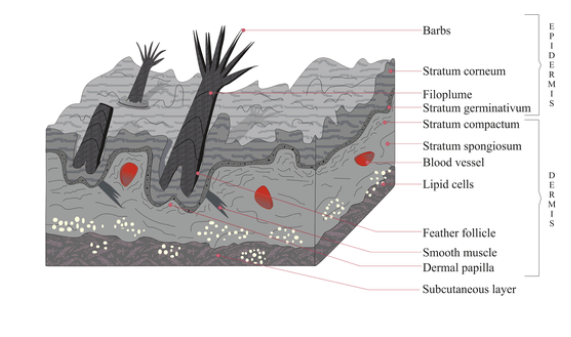
Integument in Amniote Tetrapods Avian Amniotes
Feathers
Beaks (covering), claws, scales
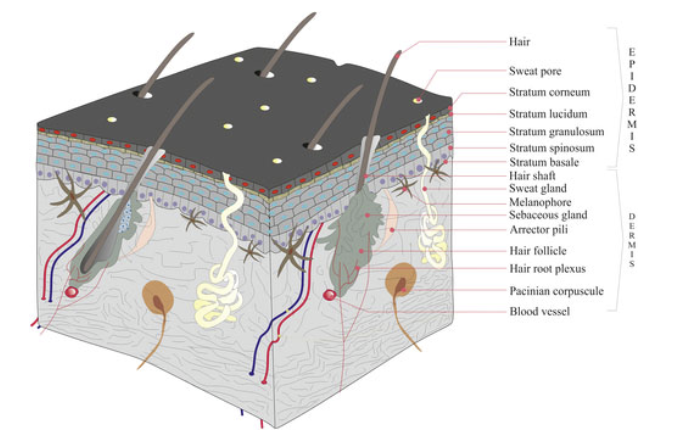
Integument in Amniote Tetrapods Mammalian Amniotes
Fur & hair
Claws, scales, “horns”
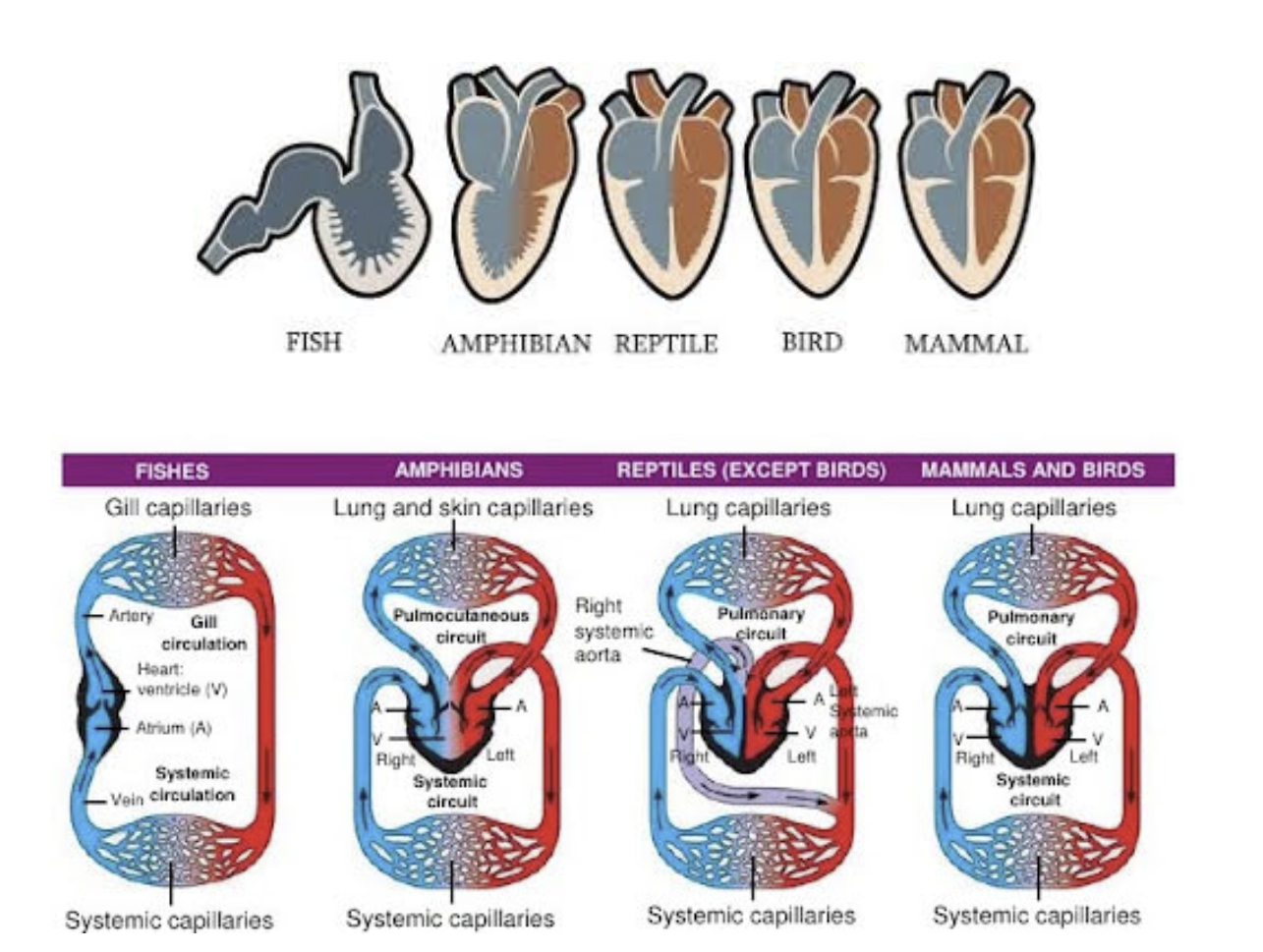
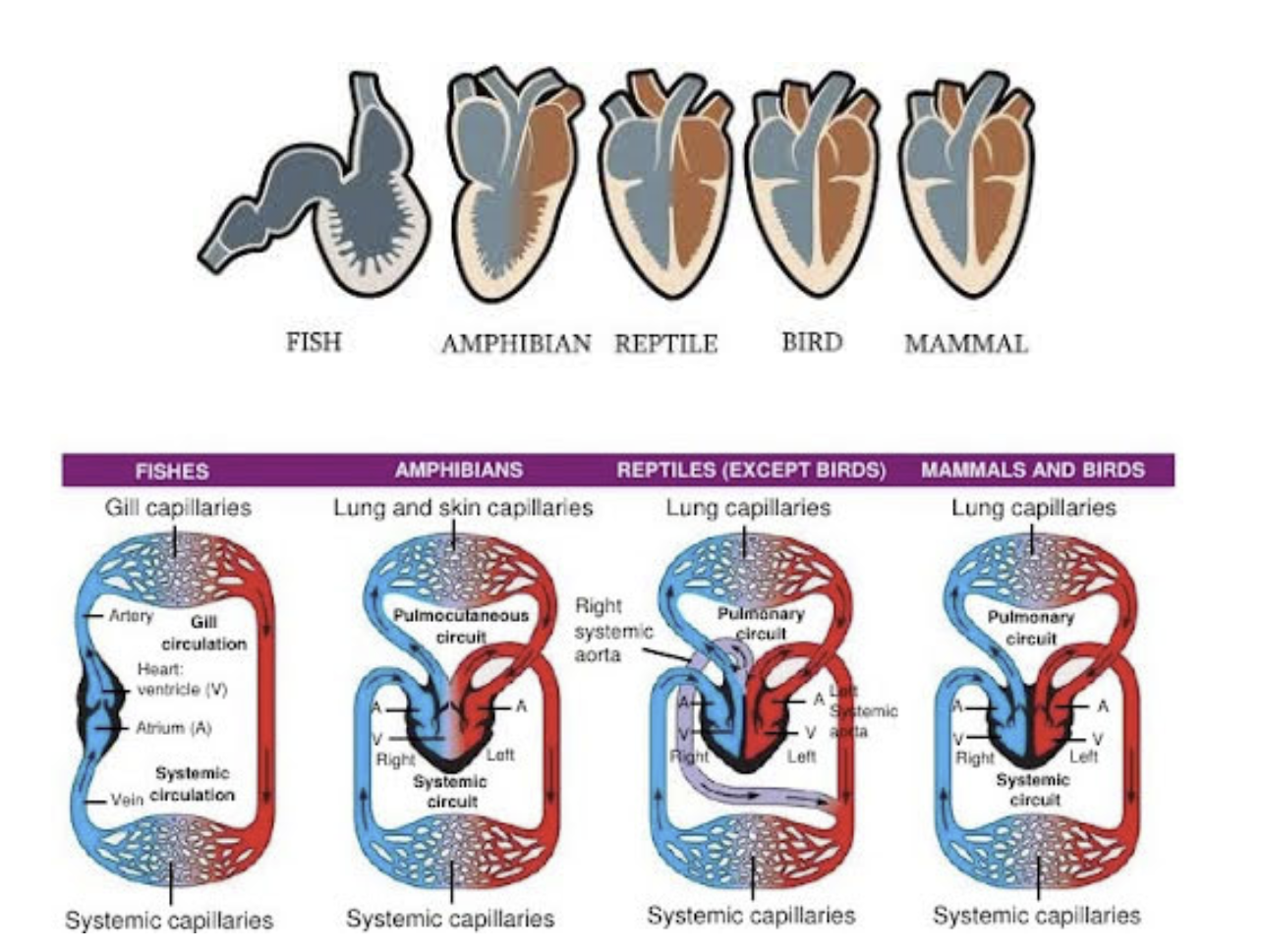
Evolution of the Circulatory System in Non-Amniote Tetrapods Heart
Ancestral condition in fishes - an undivided tubular heart
Increasing reliance on lung ventilation seelcted for a beginning separation of the tubular heart into right & left chambers
Separation is in the atrium, resulting in right & left atria
The ventricle remains undivided
The heart is three chambered
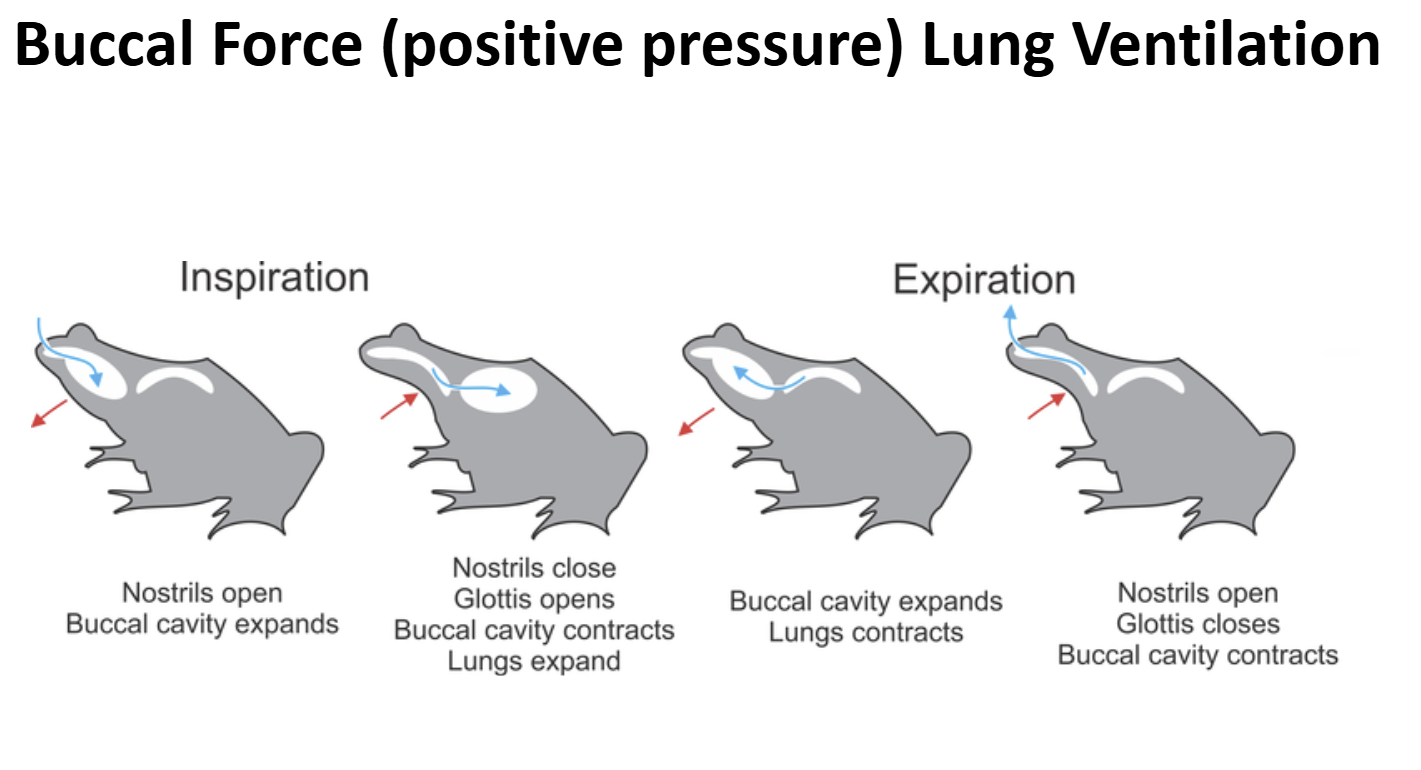
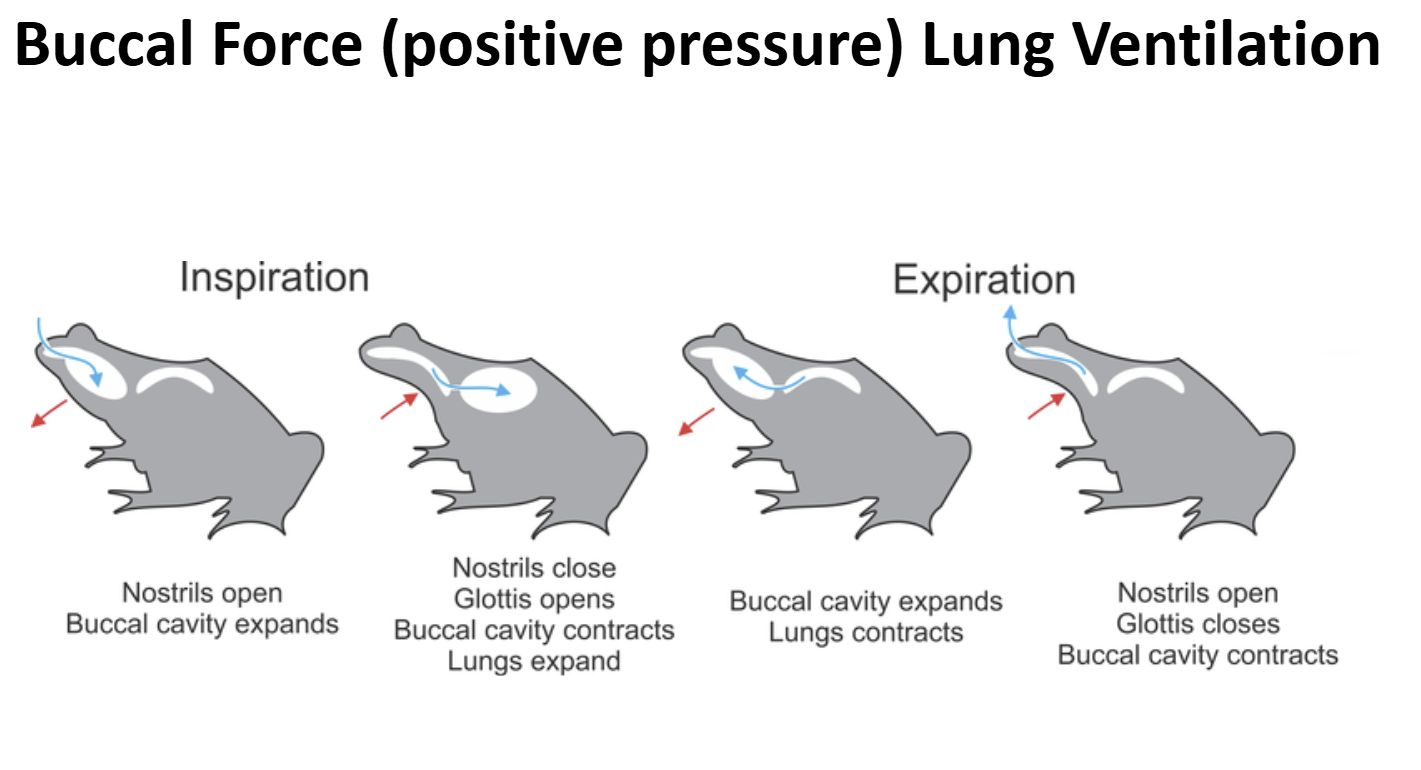
Lung Ventilation in Early Non-Amniote Tetrapods & Extant Lissamphibia Ancestral Condition: Buccal Force (Positive Pressure Ventilation)
Early non-amniote tetrapods lack/lacked a rib cage
The body wall was/ is not muscular & not rigid
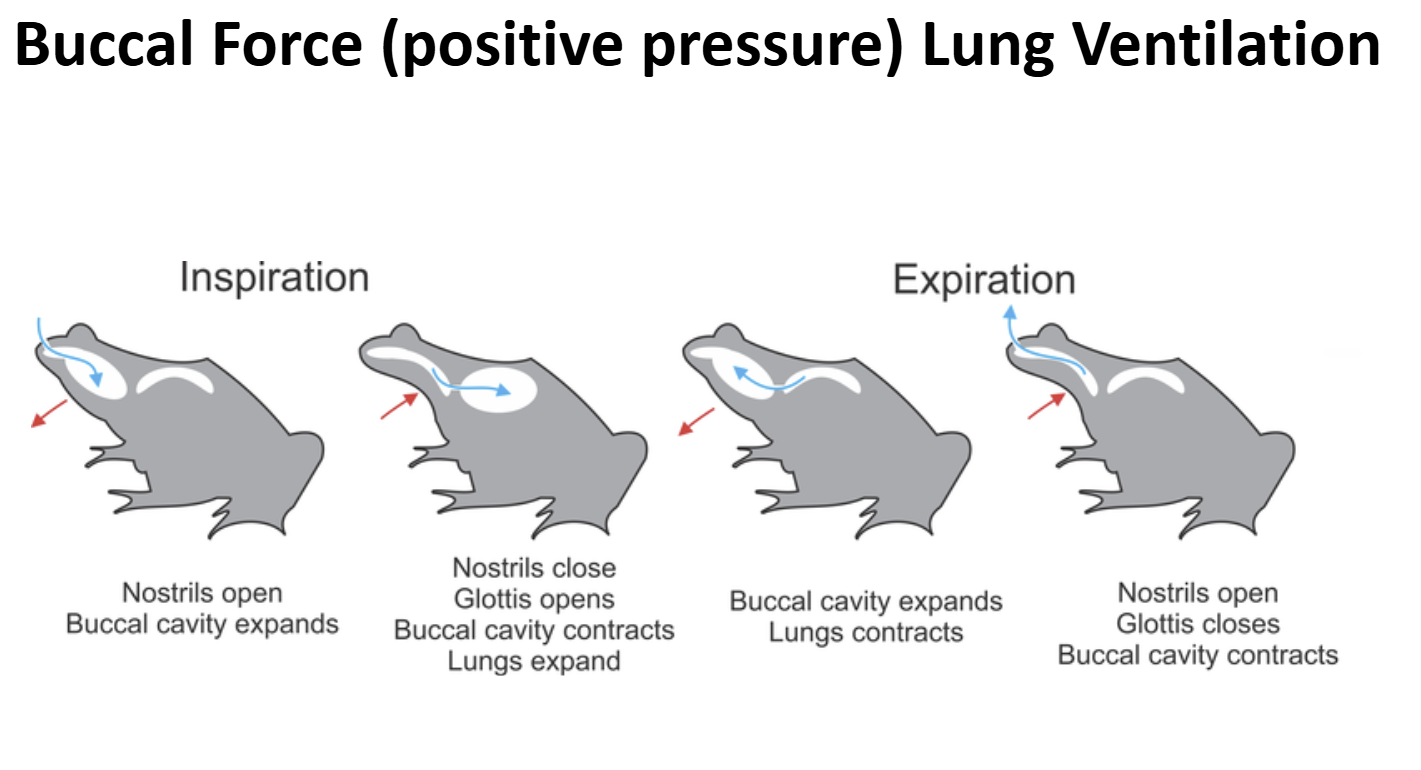
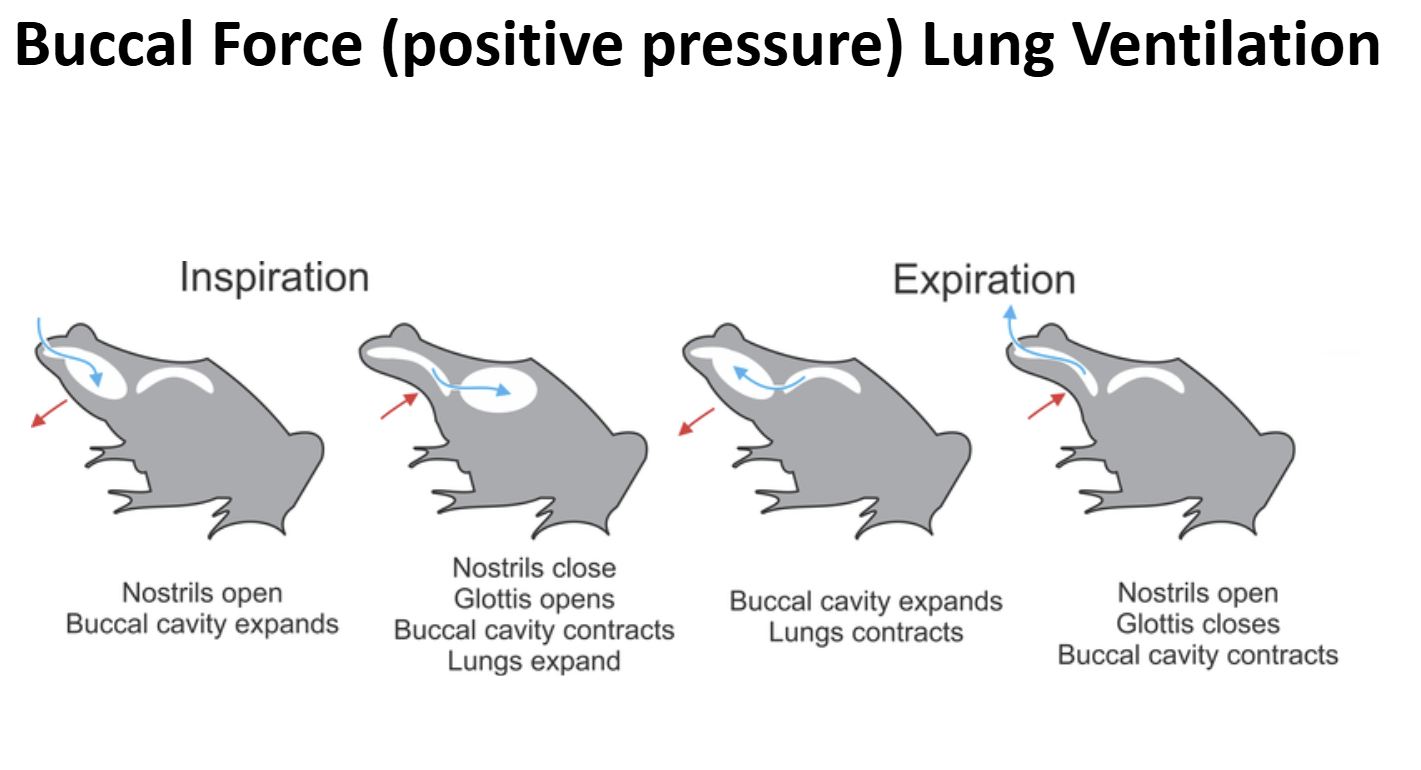
Lung Ventilation in Early Non-Amniote Tetrapods & Extant Lissamphibia Ventilatory Cycle
- A bolus of air is present the lungs & gas exchange is on-going
- Valvular nares are opened, valvular glottis is closed
- Muscular floor of the bucco-pharyngeal cavity is forcefully depressed
- Air is sucked into the bucco-pharyngeal chamberGlottis is opened, external nares opened
- Body wall muscles contract pushing air out of the lungs over the air bolus being held in the bucco-pharyngeal chamber (exhalation)External nares are closed, glottis is opened
- Muscles of the bucco-pharyngeal cavity contract forecully pushing air bolus through the opened glottis into the lungs (inhalation)Glottis is closed, gas exchange occurs
- External nares are opened to allow air to be moved in-and-out for olfaction
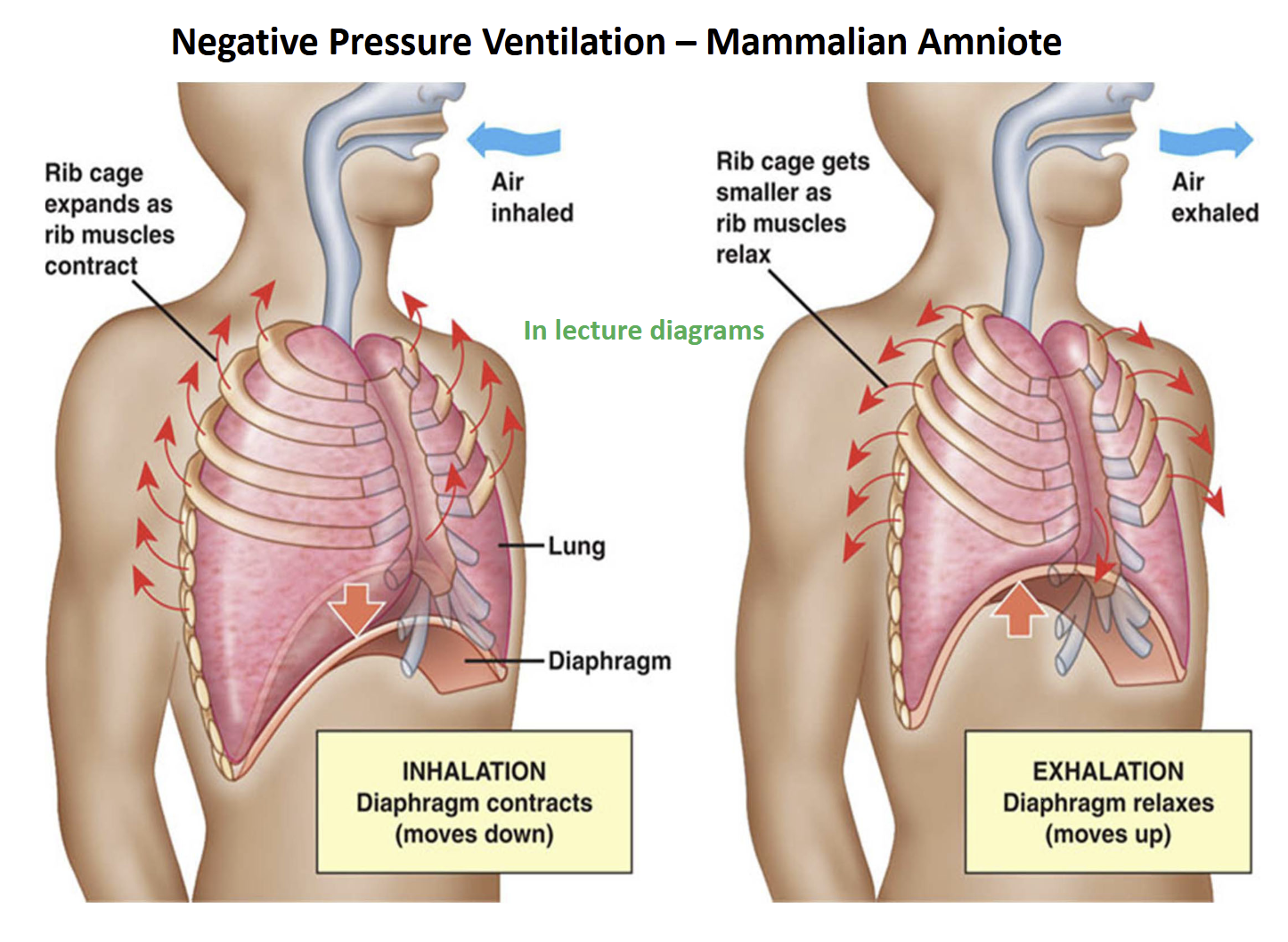
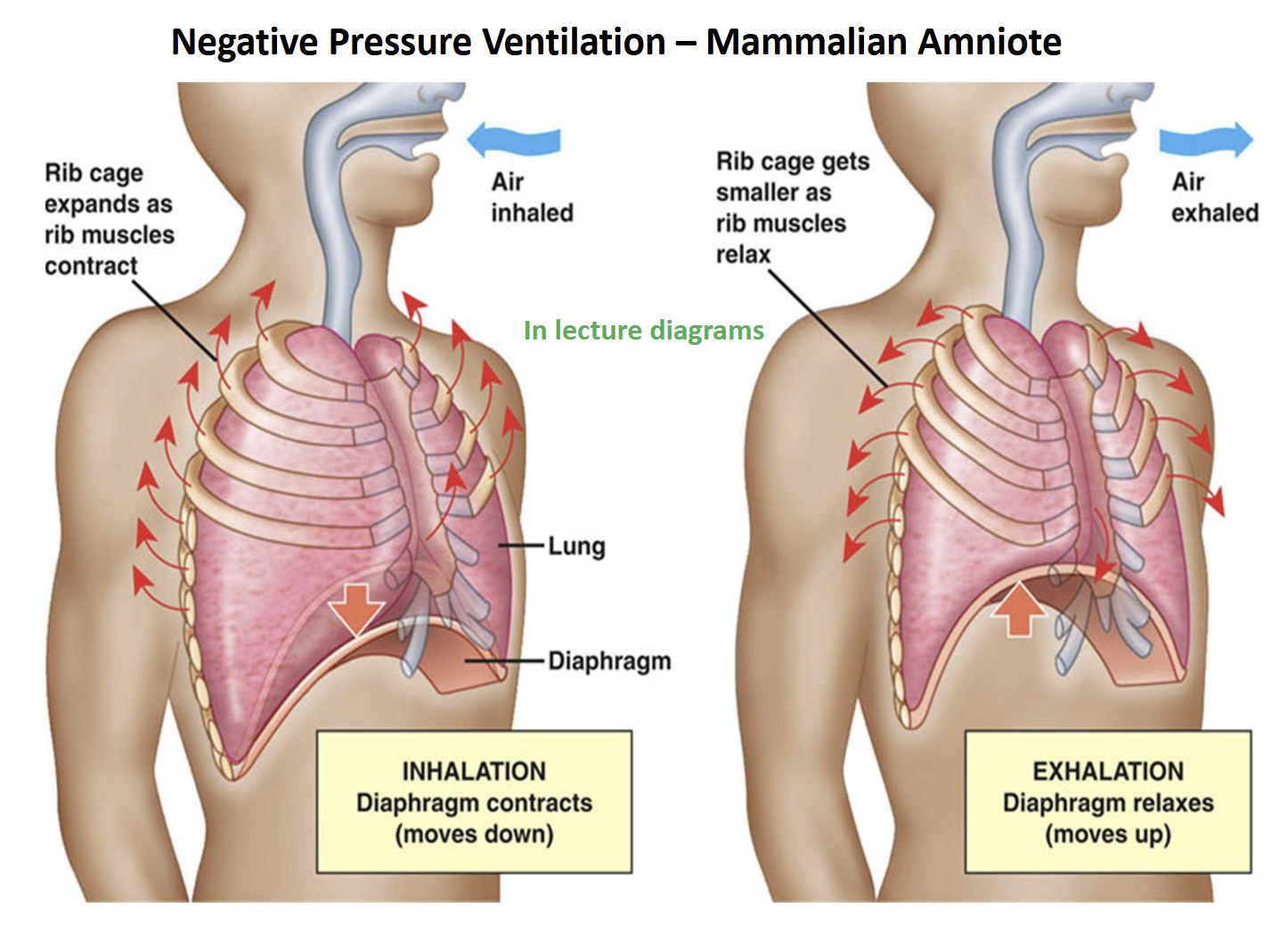
Evolution of Negative-Pressure (suction) Lung Ventilation Derived Condition
Development of longer ribs that projected ventrally eventually culminated in a “rib-cage” in later non-amniote tetrapods, and amniotes
In addition, amniotes developed a muscular partition separating the thoracic & abdominal cavities (= diaphragm)
During the transition from non-amniote to amniote tetrapods, increased rigidity of the body wall of the trunk (ribs + skeletal muscle), promoted ventilation of the lungs by negative pressure (suction)
Air is sucked into lungs with negative pressure
WLT and the Evolution of Mouths in Tetrapods
In water, prey are brought into the mouth using suction + forward intertia. Aquatic prey do not require moistening for swallong, already wet
WLT placed different selection pressures on tetrapod mouths that influenced the tongue, teeth, & oral glands
WLT and the Evolution of Mouths in Tetrapods Ancestral Condition (fishes & aquatic NAT’s)
Primary tongue = a fleshy lobe lacking intrinsic musculature, skeletal support, & is not highly glandular
Few oral glands elsewhere in the mouth
Teeth are homodont, and can be widely distrubuted in the mouth (jaws, palate, tongue), function for grasping prey
WLT and the Evolution of Mouths in Tetrapods Derived Condition
Evolution of the definitive tongue:
Intrinsic musculature present
Skeletal support (hyoid)
Makes mobile; protrusible
Glandular
Tongue functions in prey capture & manipulation of prey inside the mouth
Number of oral glands increases, and they become concentrated in amniotes (= salivary glands)
The total number of teeth decreases & they become confined to the jaws
Teeth are homodont = similar size & shape, continue to function for grasping, crushing, some shearing of prey (no true mastication until mammals)
WLT and the Evolution of Mouths in Tetrapods Derived Condition Differences between Lissamphibia, Amniote, and Turtles
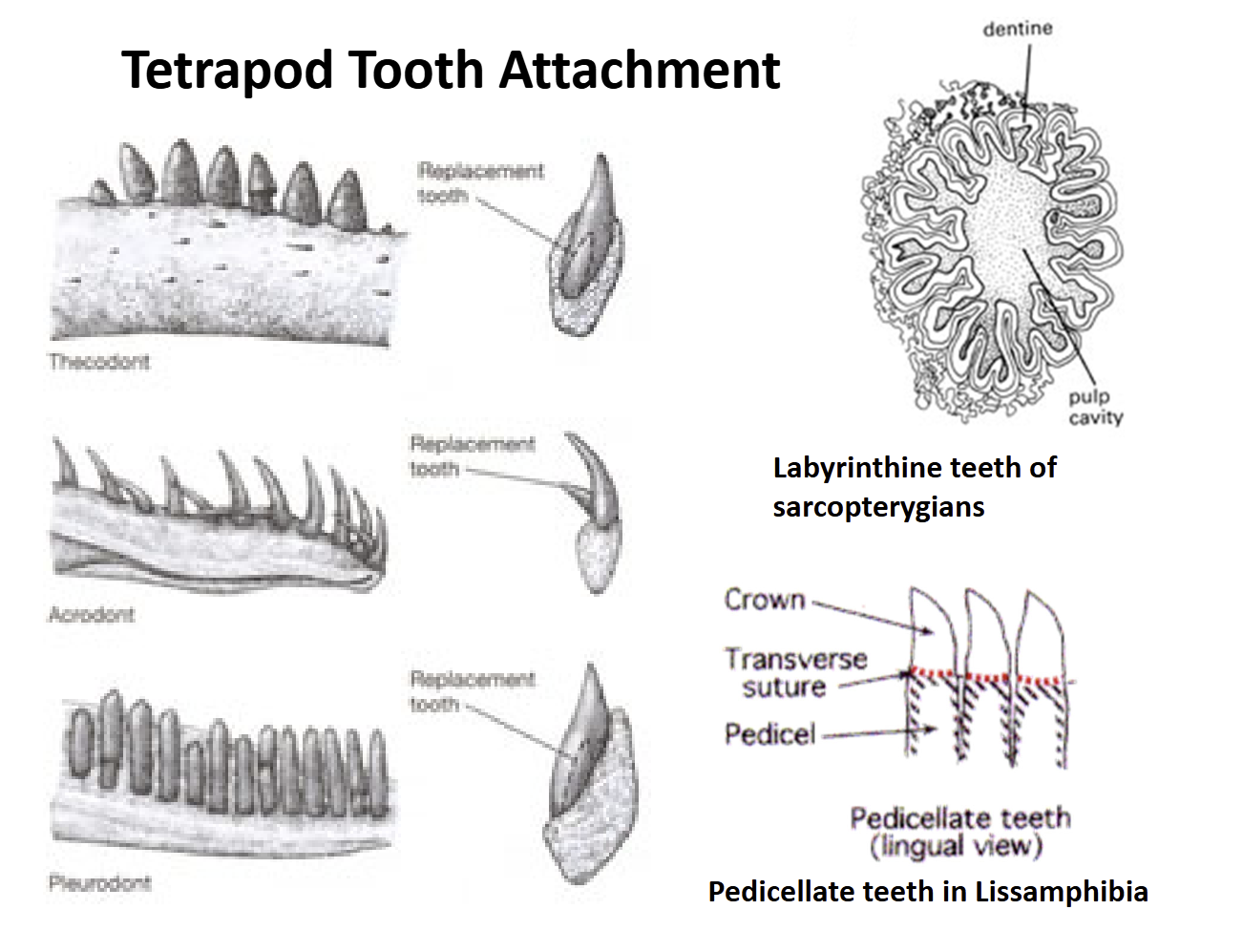
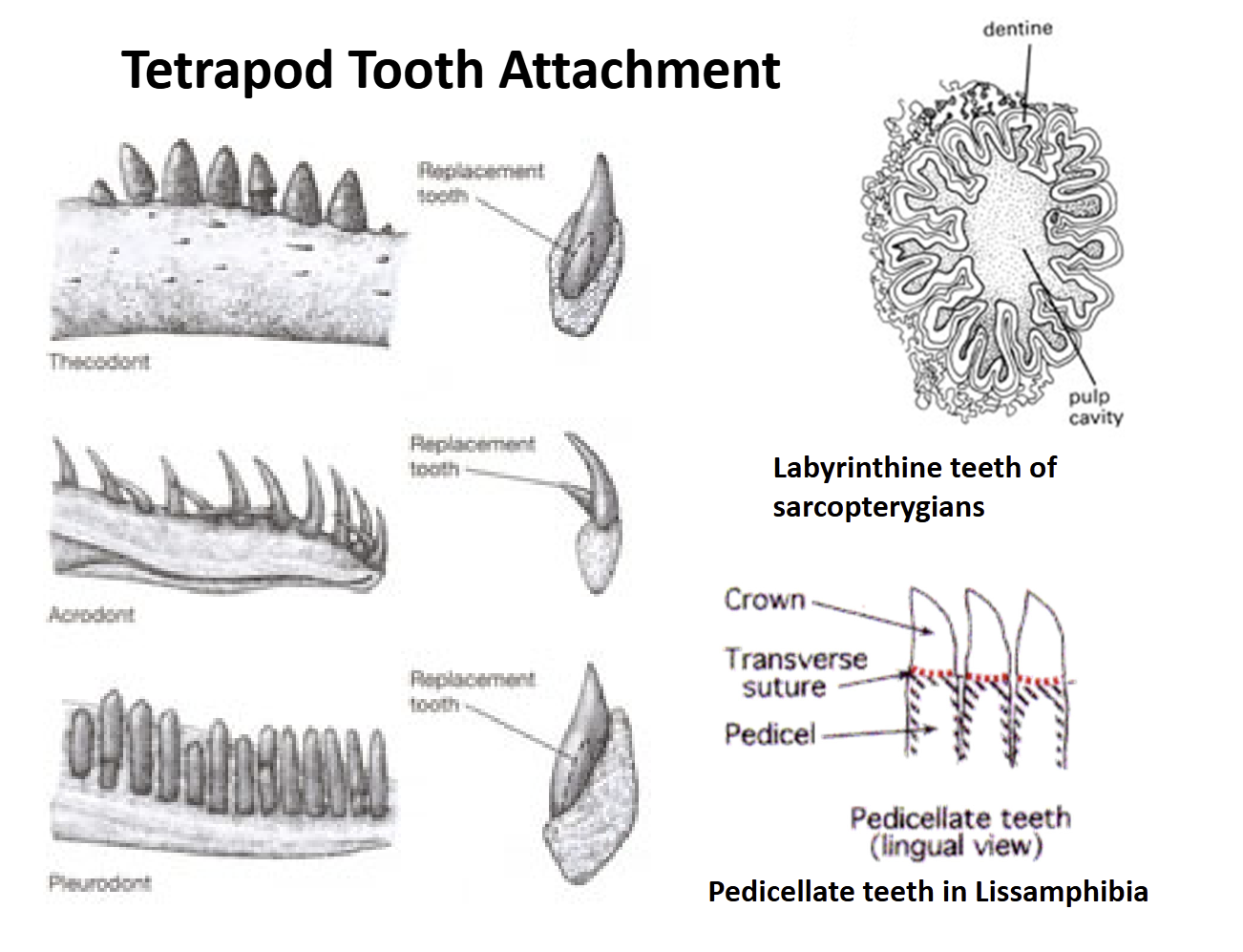
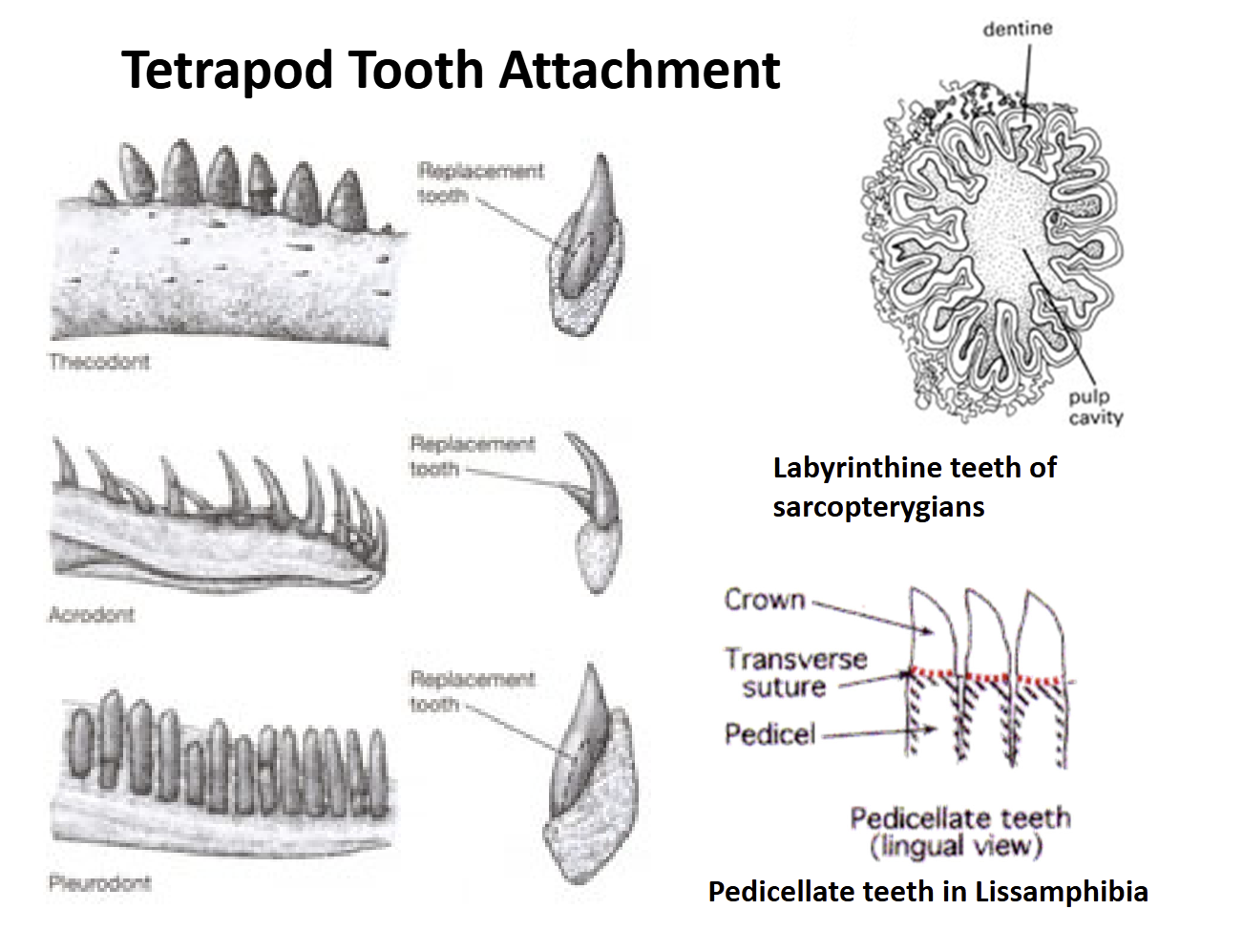
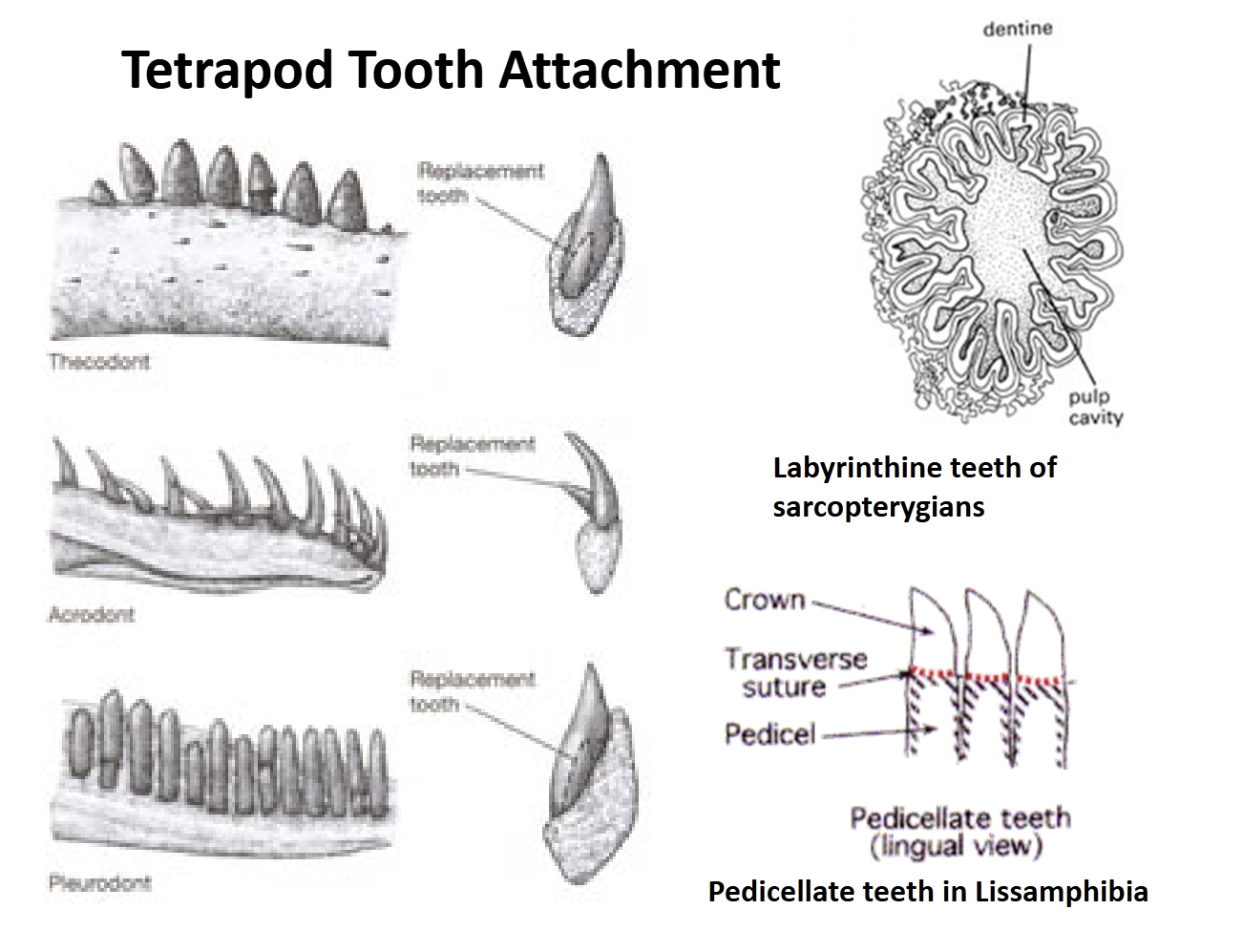
Lissamphibia teeth are pedicellate = have a base (pedicel) that supports a crown. There is a zone of weakness between the crown & pedicel that allows teeth to bend some without breaking
Amniote teeth are not pedicellate. Show three types of attachment to the jaw bones:
Acrodont - attached to the top of the jaw
Pleurodont - attached to the side of the jaw
Thecodont - anchored into sockets in the jawbones
Turtles - teeth are absent, jawbones covered by a thick layer of keratin (derived from epidermis)
Subclass Lissamphibia
“Modern amphibians”
Lissos (greek) → “smooth”, moist and scaleless skin
Thought to be a monophyletic group composed of three distinct lineages
Non-amniote tetrapods, lay eggs in water
Subclass Lissamphibia Shared Derived Characters
Moist, permeable skin with substantial cutaneous gas exchange
Pedicellate, bicuspid teeth
Operculum-columella complex (two bones that transmit sounds to the inner ear)
Green rods ( in addition to red rods common elsewhere) max sensitivity to blue light in low-light conditions
Levitator bulbi muscle - thin sheet in floor of eye socket causes eyes to bulge outward, enlaring buccal cavity
Relatively immobile tongue, not picky carnivores

Distinguishing the Three Lissamphibian Clades
Caudata - salamanders, newts, efts (~ 350 species)
Have well developed tails
Most have 4 limbs
Terrestial locomotion by crawling
Swim with lateral undulation (tail) when in water
Anura - frogs & toads ( at least 22 families, 3,000+ species)
tailless
All have 4 limbs
Terrestial locomotion by hopping (ability variable)
Swimming using webbed hind feet when in water
Apoda - caecilians (many fewer species)
Appendages lost as an adaptation for fossorial (i.e., burrowing) life
Terrestial locmotion by burrowing
Aquatic locomotion by swimming using lateral undulations

Order Caudata A Caudata Curiosity
Most vertebrate genomes are 1-7 picograms, but the mean for salamanders is 35pg (up to 120!)
Goes hand-in-hand with larger cell size than most other vertebrates
Requires morphological and developmental adjustments
Caudata Life History Overview
Locomotion
Paedomorphism
Loss of lungs
Complex life-cycles
Internal fertilizatoin by spermatophore
Complex courtship & mating behavior

Crawling Locomotion in the Caudata
Least specialized skeleton & musculature
Least speciliazed locomotion
In many salamande species, alternate legs on opposite sides of the body move at the same time

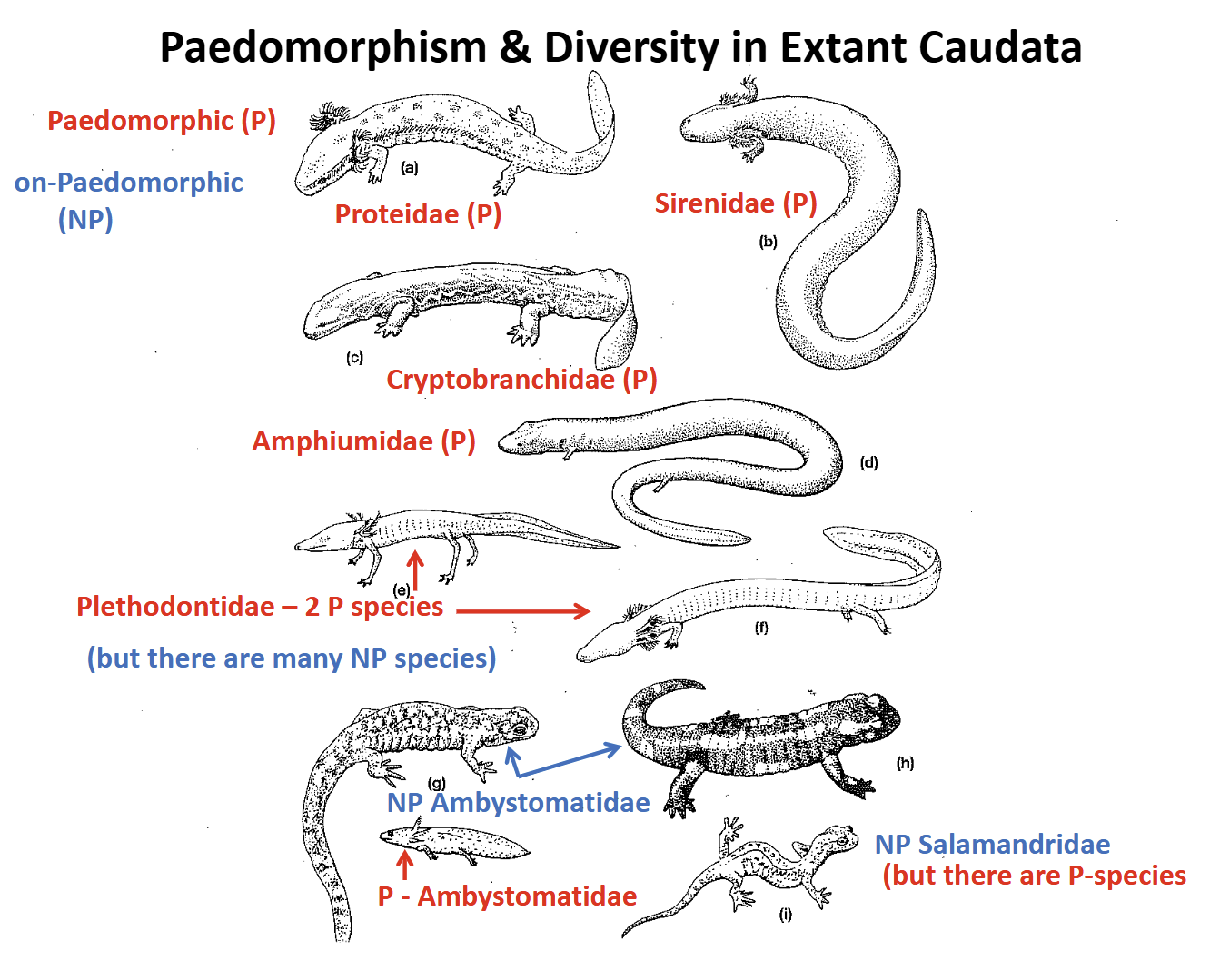
Paedomorphism in Caudata
Retention of the aquatic larval lifestyle in adults & character states that are adaptations for aquatic life:
external aquatic gills
Fin-like tail
Reduced appendages
Lack moveable eyelids
Thin skin covering
Lateral line system