Module 7: Genetic Mechanisms of Axis Determination
1/69
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
70 Terms
Tagmas
The three main regions of the Drosophila body plan
Head
Thorax (T1-T3)
Abdomen (A1-A8)
The Drosophila body plan is organized into segments or metameres, and the anterior-posterior and dorsal-ventral body axes are already established in the developing egg
What are the three tagmas of a Drosophila fly?
Head
Thorax (T1-T3)
Abdomen (A1-A8)
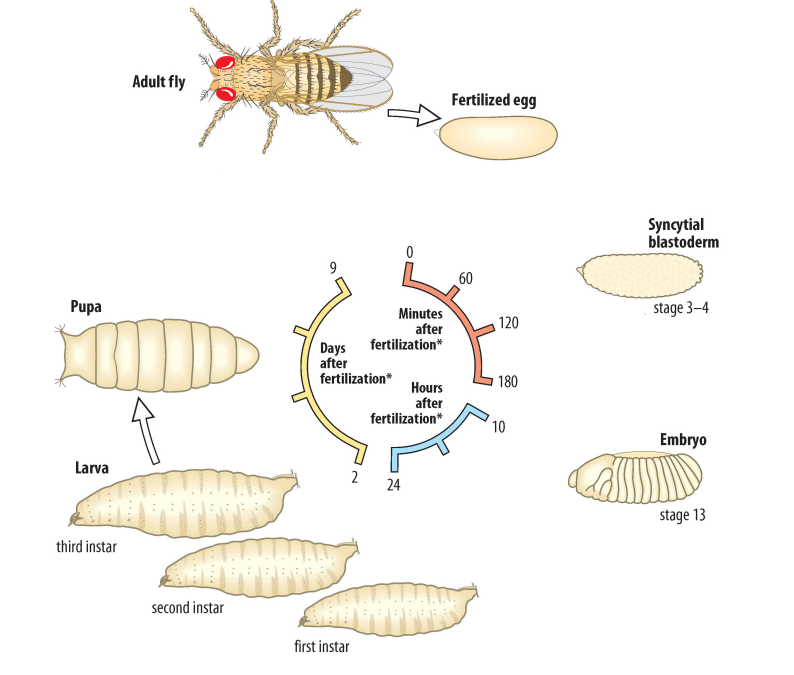
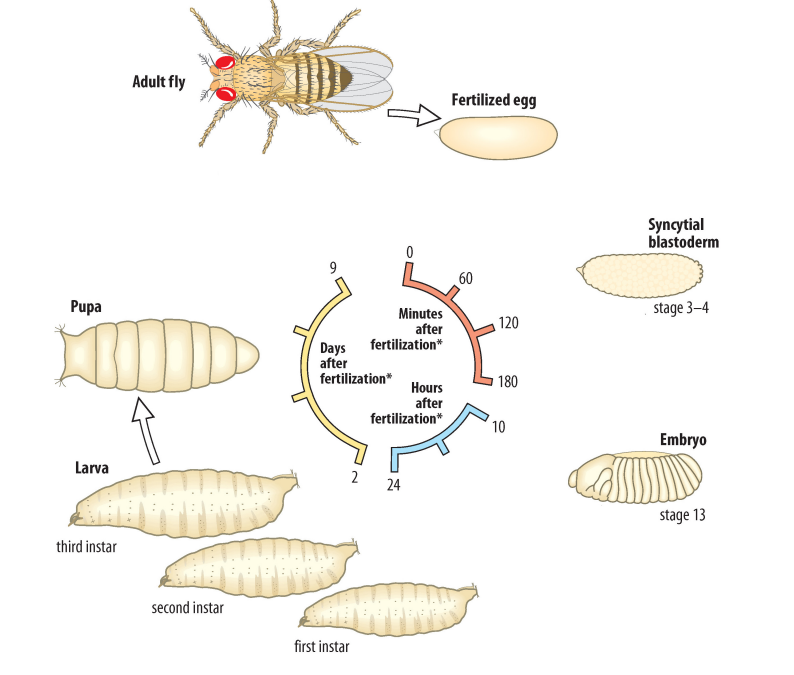
Describe the stages of the Drosophila life cycle.
Drosophila undergo internal fertilization and external development
Once a sperm cell enters the micropyle located at the anterior side of the centrolecithal egg, the sperm cell forms a plaque using its long flagellum, effectively blocking out other sperm from entering the micropyle
The fertilized centrolecithal egg undergoes meroblastic superficial cleavage to form a blastula
In insects, cells undergo the S phase and M phase during cleavage
Because cells do not undergo cytokinesis during cleavage, but continue to undergo DNA replication and mitosis, a syncytium (a giant cell with multiple nuclei) forms
The blastula undergoes gastrulation, during which the three germ layers (ectoderm, mesoderm, endoderm) are specified
The gastrula develops into an embryo and hatches, giving rise to a larva that passes through three stages (instars) of development
First instar
Second instar
Third instar
During each instar, the larva is highly active and continuously feeds to grow in size
During gastrulation, there is a retraction of the blastula towards the most posterior end
Once the larva reaches a certain size, it pupates to form a pupa and undergoes metamorphosis to become the adult fly
The entire process of Drosophila development (from fertilization to reaching sexual maturity) takes 10 days!
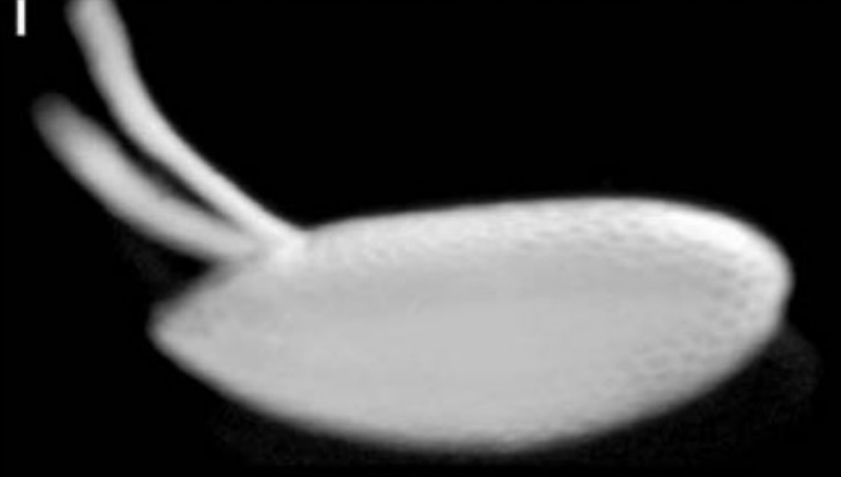
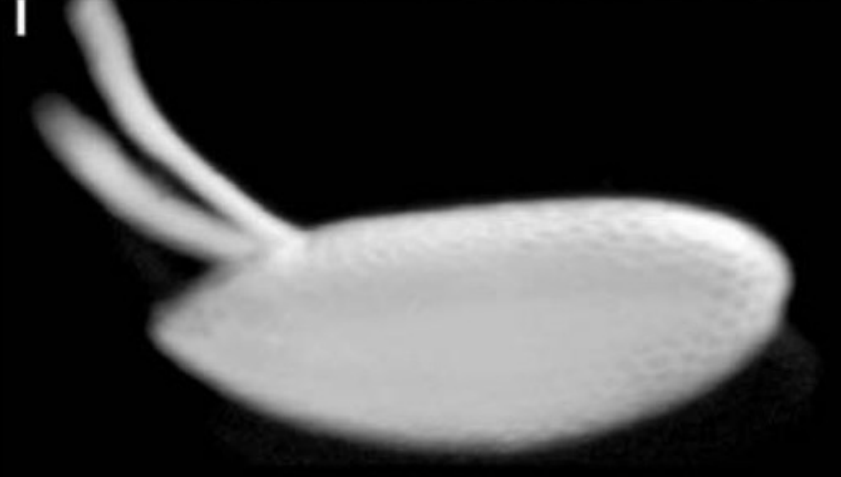
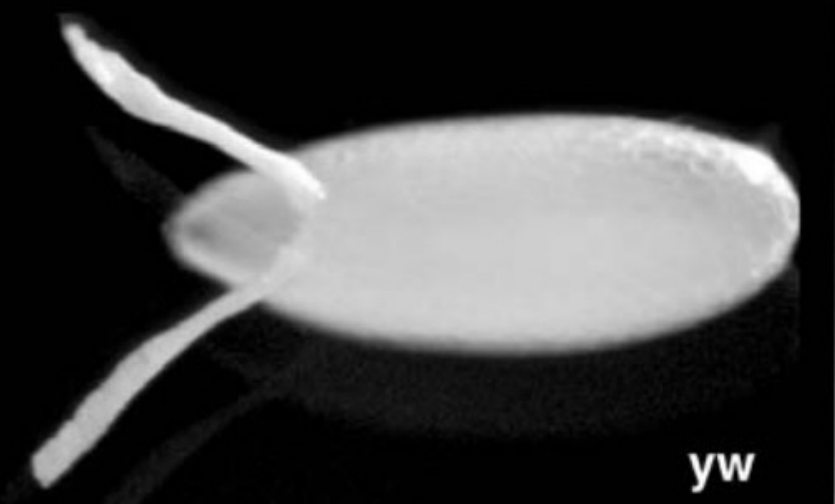
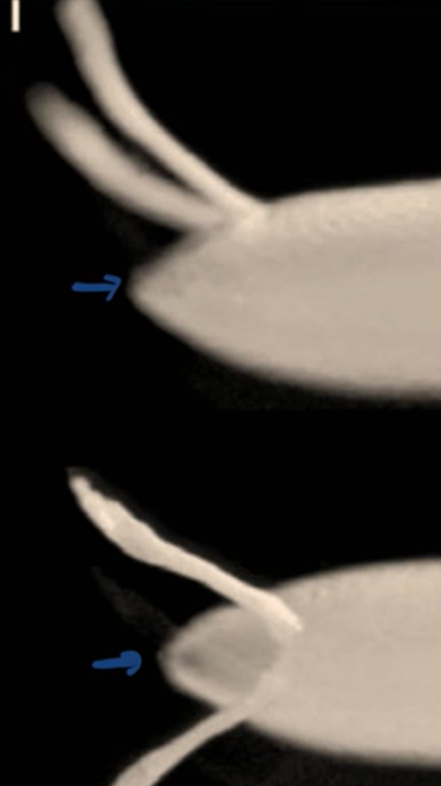
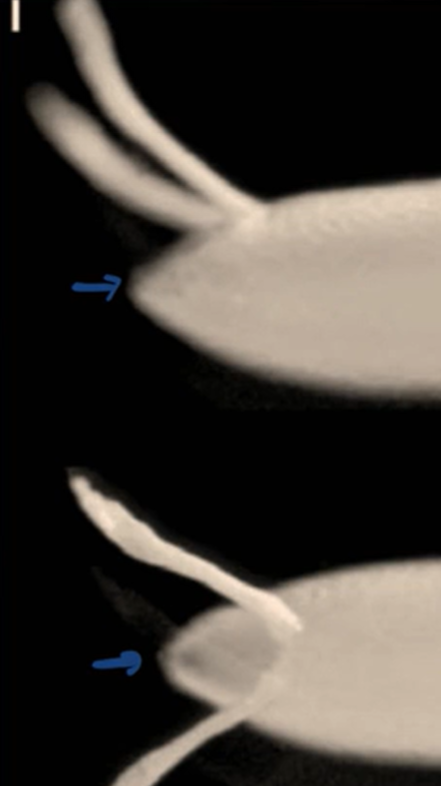
Describe what you see in the figure below.
A lateral view of the Drosophila chorion (egg shell)
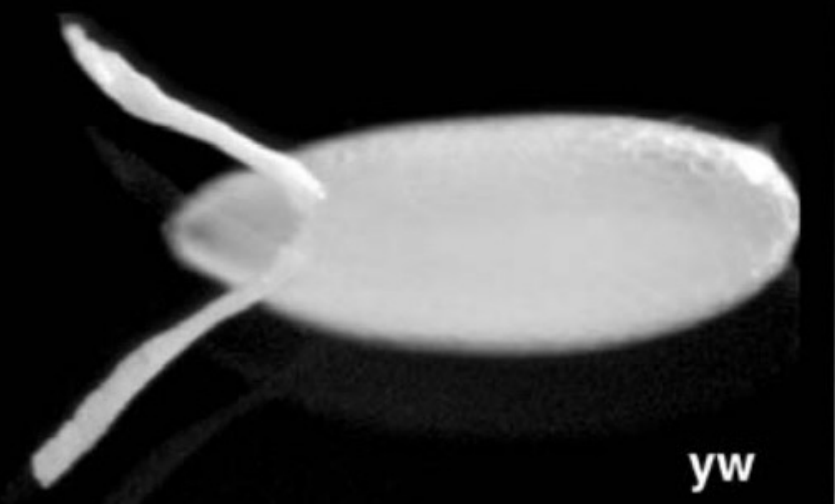
Describe what you see in the figure below.
A dorsal view of the Drosophila chorion (egg shell)
Chorion
The shell of the Drosophila egg
Rigid, acts as a physical barrier to sperm
Micropyle
A small opening at the anterior end of the chorion that allows sperm to penetrate
Once sperm has penetrated into the micropyle, it can stay alive for an extremely long time
Describe how polyspermy is prevented in Drosophila.
A Drosophila sperm cell has an extremely long flagellum (nearly 300 times longer than the flagellum of a human sperm cell)
Once a sperm cell enters the micropyle, its long flagellum forms a plaque that forms a physical plug, preventing other sperm cells from entering the micropyle and fertilizing the egg
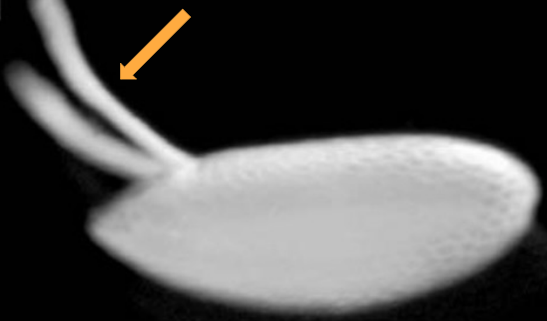
Dorsal appendages
A pair of tube-like structures that are present on the dorsal side of the embryo'
Involved in communication and gas exchange
Allow the embryo to breathe
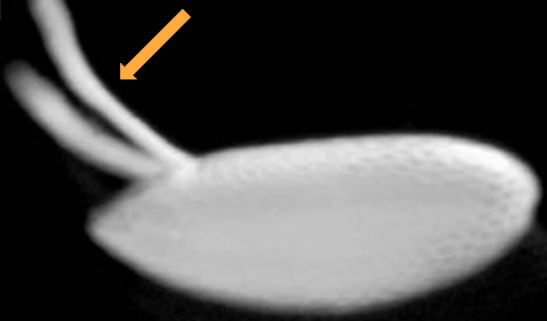

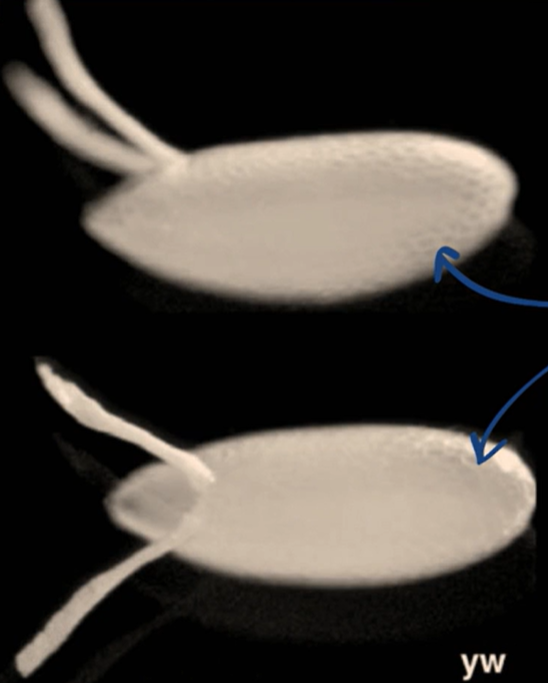
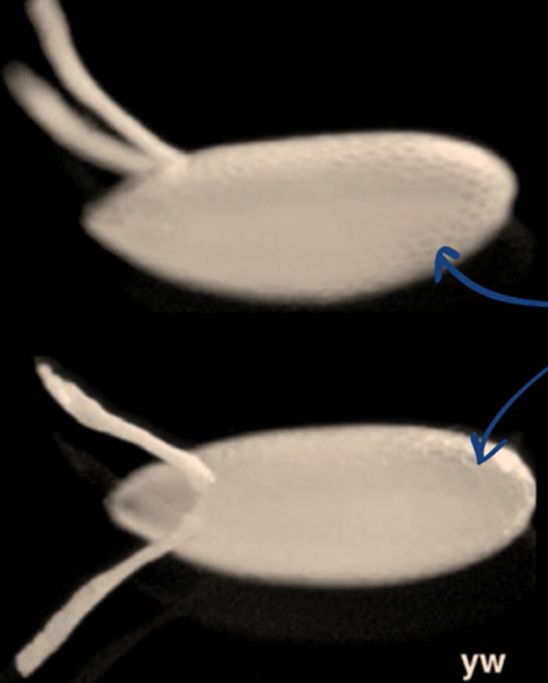
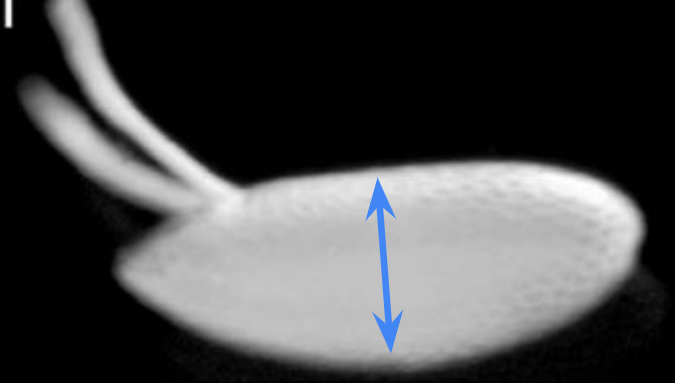
Which body axis is represented in the image?
The anterior-posterior axis

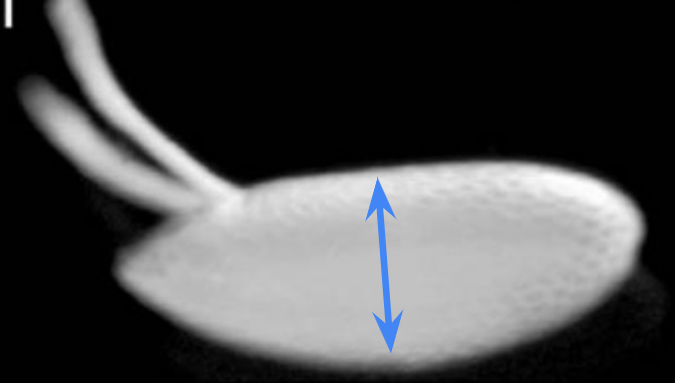
Which body axis is represented in the image?
The dorsal-ventral axis
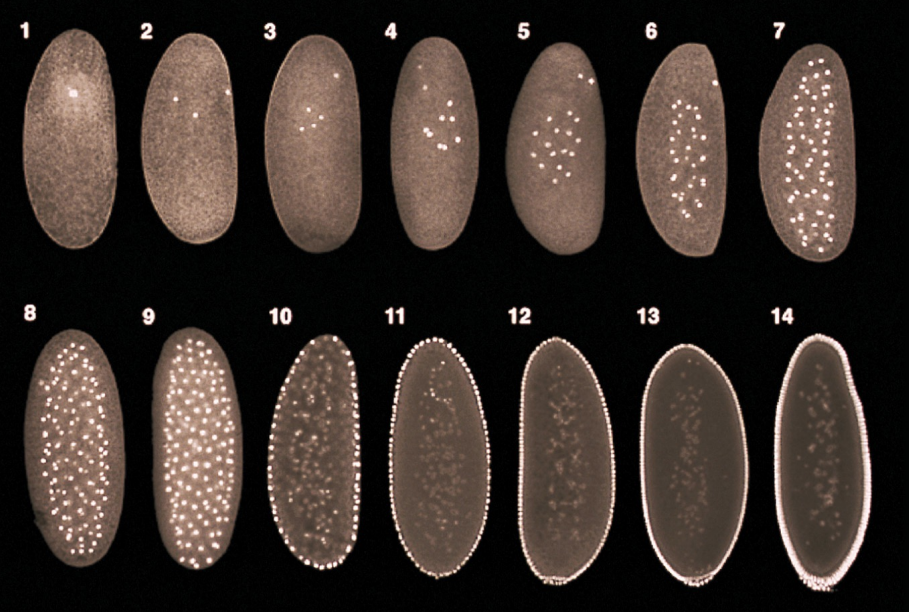
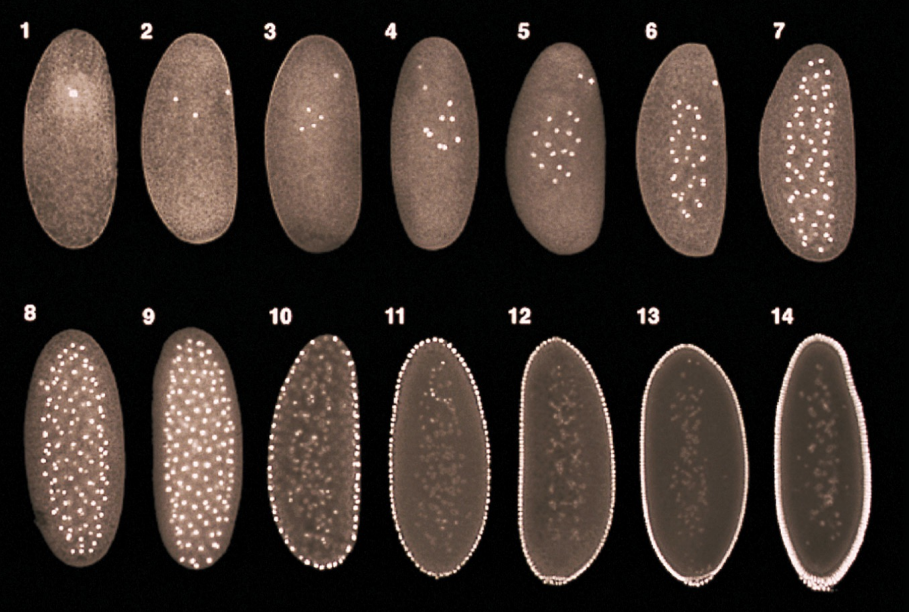
Describe how the process of meroblastic superficial cleavage in Drosophila generates a syncytium.
Cleavage in Drosophila:
Within the span of hours, the number of nuclei in the blastula progressively increase with each cell division
From the 1st-10th cell division, the nuclei divide and float around in the blastula
Around the 10th-11th cell division, the nuclei begin to migrate to the external surface of the blastula to form the syncytial blastoderm and the yolk concentrates in the interior of the blastula
From the 13th-14th cell division, a process of cellularization occurs, in which the cell membrane invaginates around each individual nuclei to form individual cells → this forms a cellular blastoderm (does not contain syncytium, but rather individual cells)
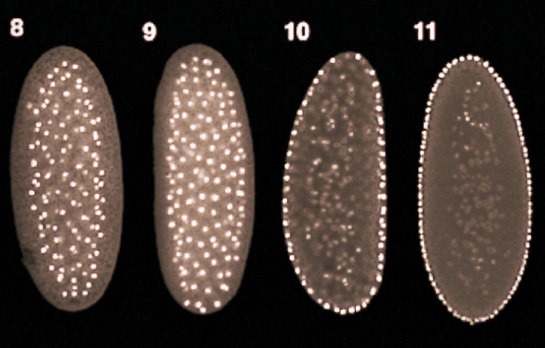
What process is occurring here?
From the 10th to 11th cell division during cleavage, freely-floating nuclei in the syncytium migrate to the external surface of the blastula to form the syncytial blastoderm
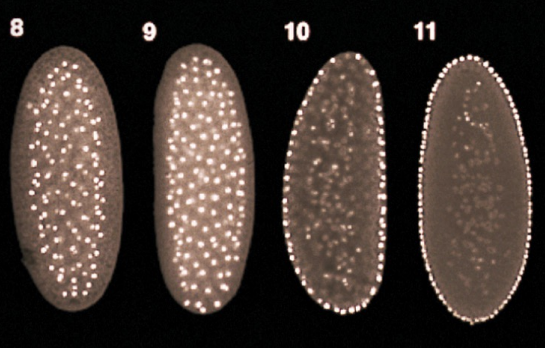
What process is occurring here?
From the 13th to 14th cell division during cleavage, cellularization occurs, in which the cell membrane invaginates around the individual nuclei of the Drosophila syncytial blastoderm to form individual cells
There is no longer a syncytium
The syncytial blastoderm becomes a cellular blastoderm
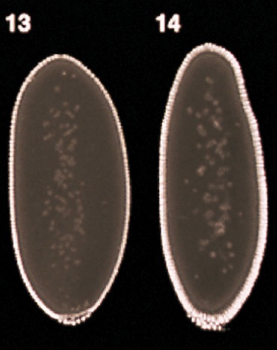
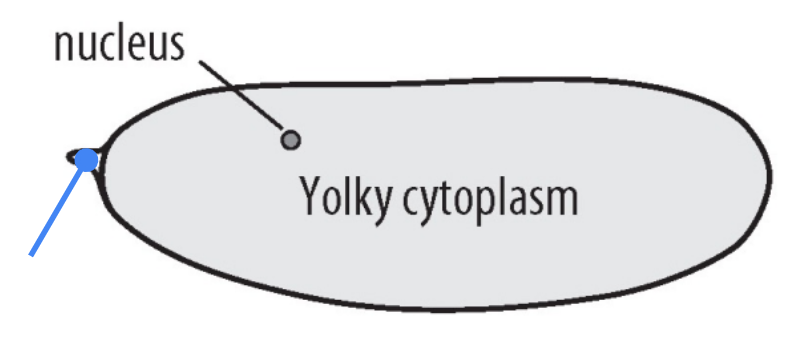
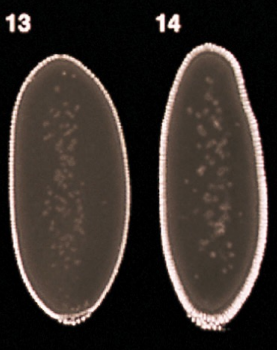
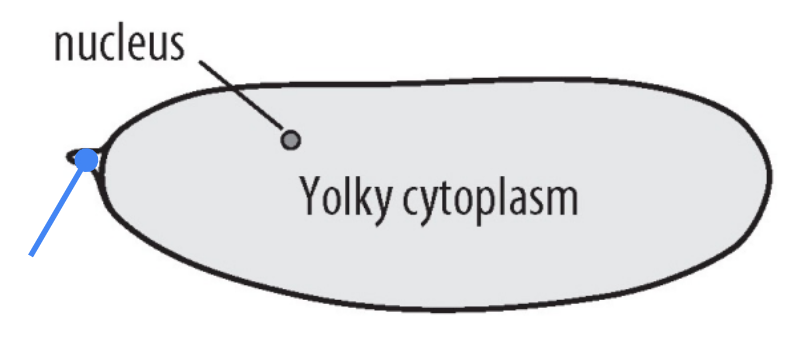
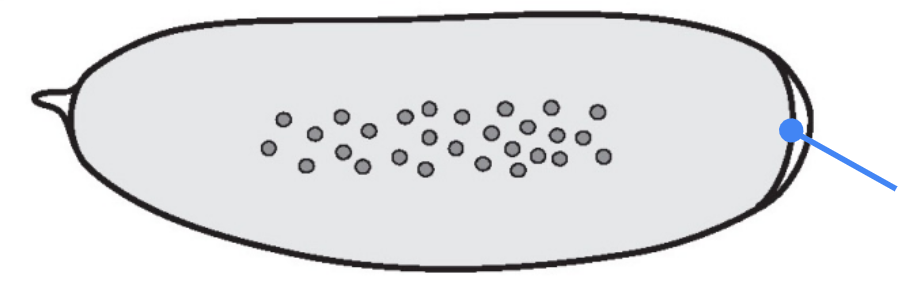
What part is shown in the figure?
Micropyle
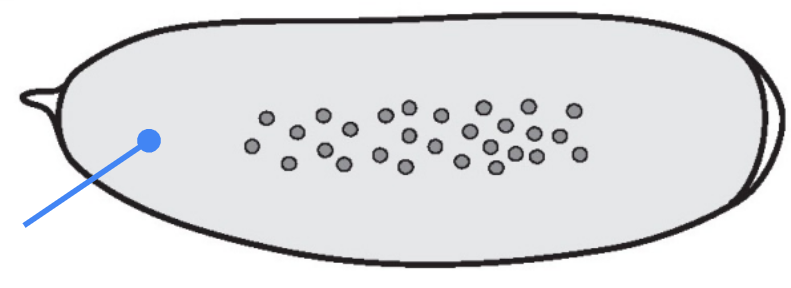
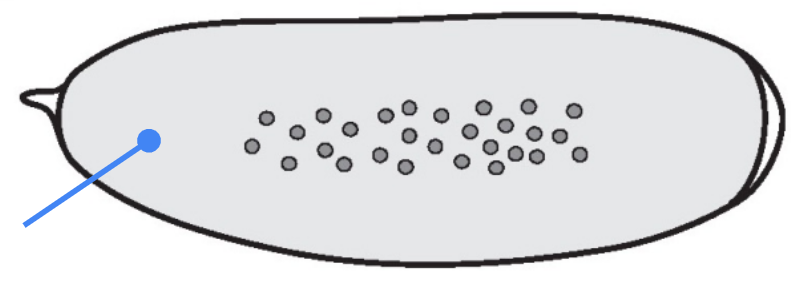
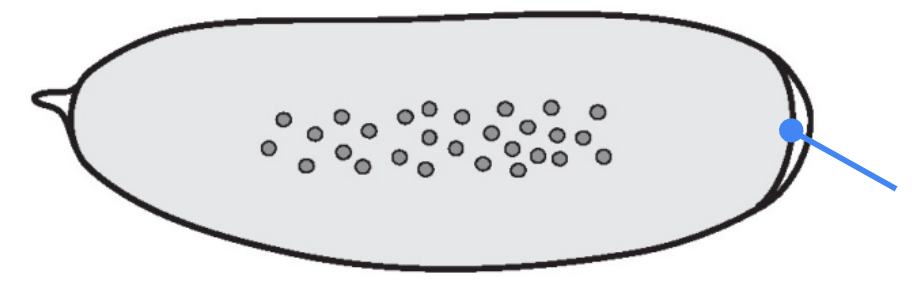
What part is shown in the figure?
Syncytium — a single large cell containing multiple nuclei formed by rounds of DNA replication and mitosis without cytokinesis
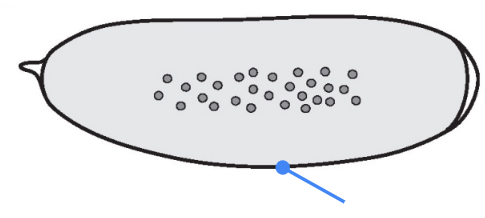
What part is shown in the figure?
Chorion
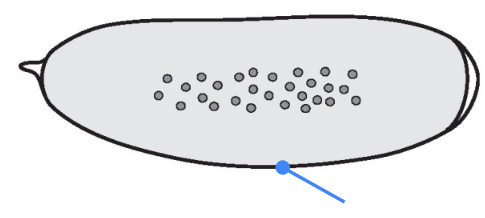
What part is shown in the figure?
Egg membrane

What part is shown in the figure?
Syncytial blastoderm (individual nuclei localized to the exterior surface of the egg)


What part is shown in the figure, and what process is it formed by?
Cellular blastoderm
The cellular blastoderm is formed by cellularization, during which the egg membrane invaginates around the individual nuclei to form individual cells

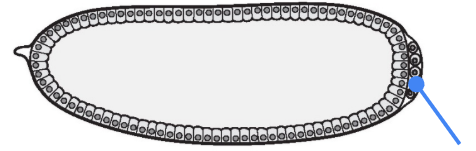
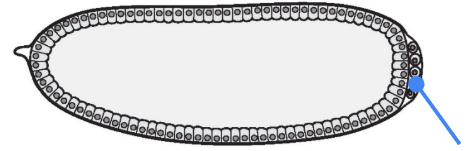
What part is shown in the figure?
Germ (pole) cells
One of the earliest decisions that the Drosophila embryo has to make is the specification and differentiation of the germ (pole) cells at the posterior side of the embryo
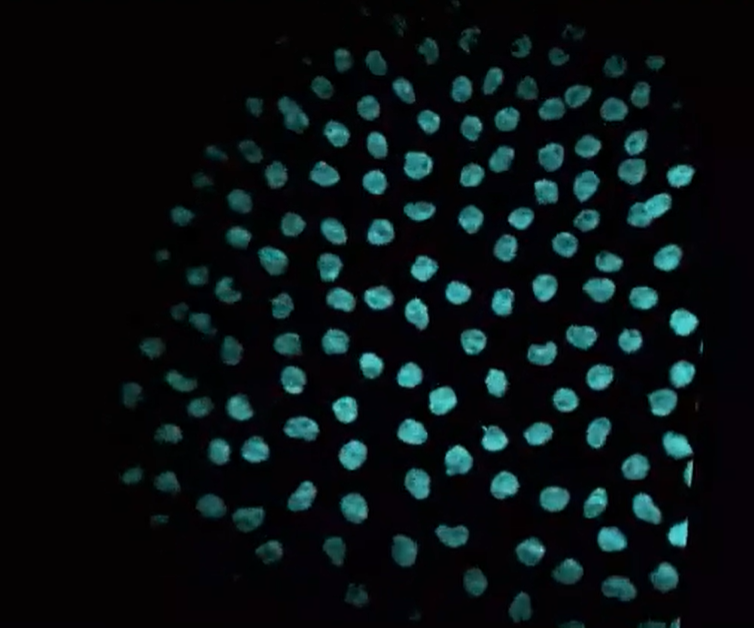
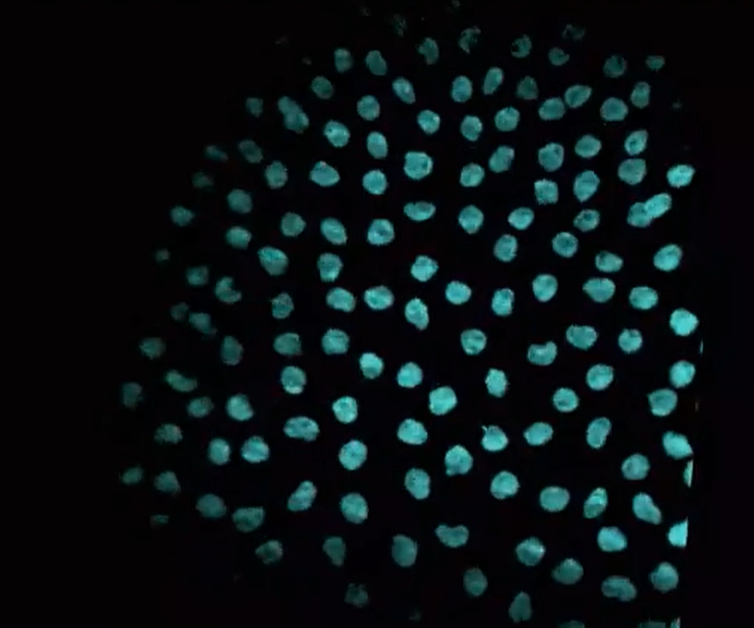
This video shows synchronized cell divisions occurring in the early Drosophila embryo. In the video, you can see the DNA being replicated and divided with each round of cell division. How was this video made?
Time-lapse imaging of a transgenic reporter line
Beta-actin promoter + H2B CDS + BFP CDS - pA
H2B encodes one of the histone proteins (H2B) that make up the histone octamer in chromatin
When is everything in the Drosophila embryo defined?
Everything in the Drosophila embryo is defined at the time of oogenesis (formation of the egg)!
The unfertilized oocyte already has the defined segments that will appear in the adult form
Syncytial specification
A type of specification in Drosophila in which cytoplasmic determinants (maternal mRNAs) are localized and positioned in the syncytium by the mother during oogenesis
Examples of maternal mRNAs:
bicoid mRNA = specifies the anterior side of the embryo
nanos mRNA = specifies the posterior side of the embryo
oskar mRNA = specifies the germ cells
Everything in the Drosophila embryo is defined at the time of oogenesis (formation of the egg), not during fertilization!
Compare syncytial specification with autonomous and conditional specification.
Autonomous specification:
A type of specification due to factors (ex. transcription factors) that are already present in the cell and act cell-autonomously, independently of neighboring cells
The presence of a transcription factor in a cell is sufficient to inform the cell, independent of context or environment
Involved in mosaic development, where cells in the embryo independently acquire their specialized fates based on intrinsic factors rather than interactions with neighboring cells
Conditional specification:
A type of specification due to signals from other cells and the environment that allows the cell to act quickly in response to stimuli
Involved in regulative development, which allows the embryo to regulate or compensate for the loss or damage of early cells (because cell fates are not yet determined, the remaining cells can alter their developmental paths and acquire different fates to replace missing parts)
Syncytial specification:
A type of specification in Drosophila in which cytoplasmic determinants (maternal mRNAs) are localized and positioned in the syncytium by the mother during oogenesis
The distribution gradient of a cytoplasmic determinant informs the identity and fate of individual nuclei depending on the relative location of the nucleus along the body axes of the embryo
In autonomous specification, a cell inherits specific factors (ex. transcription factors) that inform its identity and fate, independent of other cells. In conditional specification, a cell receives signals from surrounding cells and the environment, and can acquire different fates if its environment changes to regulate the overall course of development. In syncytial specification, individual nuclei receive information from a distribution gradient of cytoplasmic determinants (maternal factors) that are positioned in the syncytium.
Unlike in autonomous specification, the fate of a nucleus in syncytial specification depends on the specific maternal factors present in its environment, which in turn depends on its relative location along the body axes of the embryo. Autonomous specification is position-independent, but syncytial specification depends in part on the position of an individual nucleus relative to the entire embryo.
Unlike in conditional specification, the fate of a nucleus in syncytial specification depends on signals from mRNA gradients that diffuse throughout the embryo, and there are no boundaries because no cell membranes exist between the individual nuclei in a syncytium. Conditional specification involves signaling across the membranes of individual cells, while syncytial specification involves gradients within a shared cytoplasm.
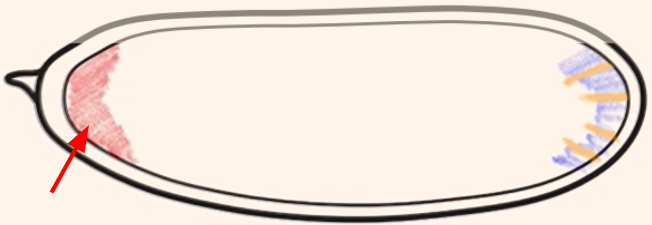
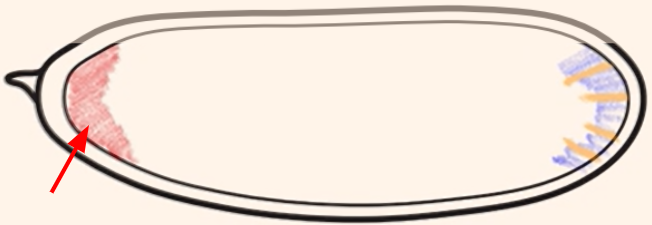
Which cytoplasmic determinant is shown in the figure?
Bicoid mRNA = specifies the anterior side of the embryo
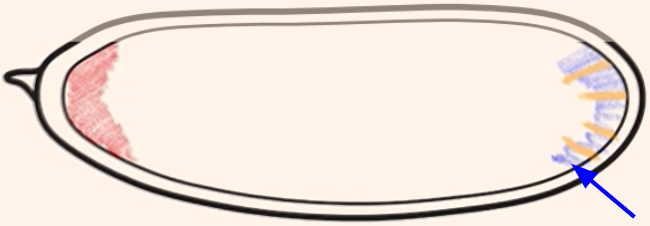
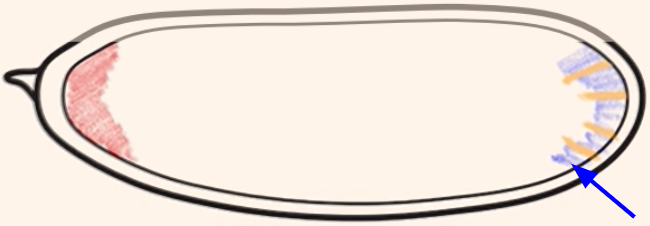
Which cytoplasmic determinant is shown in the figure?
Nanos mRNA = specifies the posterior side of the embryo
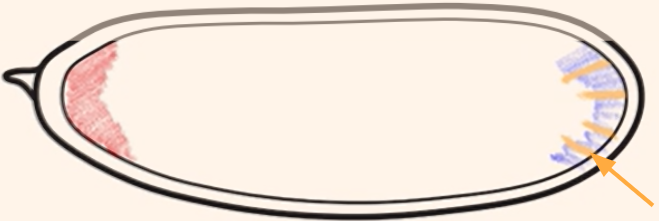
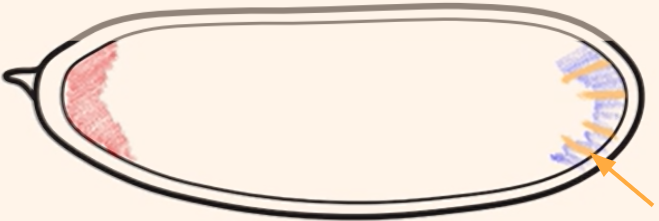
Which cytoplasmic determinant is shown in the figure?
Oskar mRNA = specifies the germ (pole) cells of the embryo
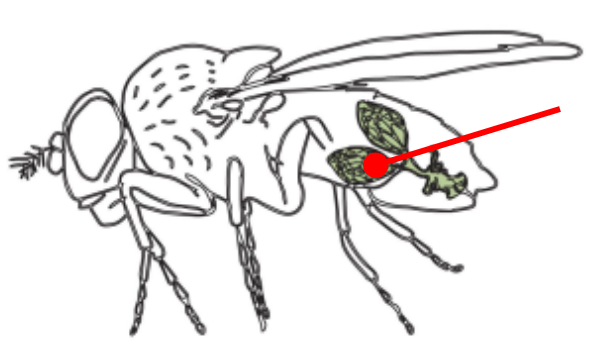
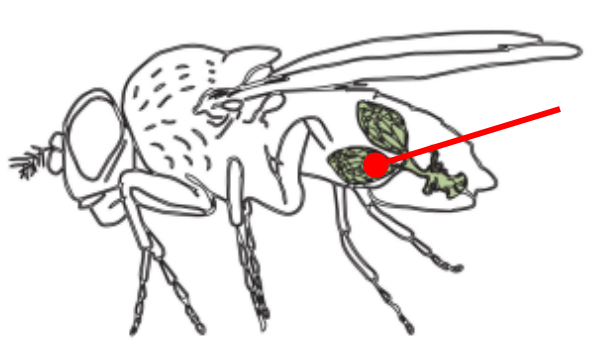
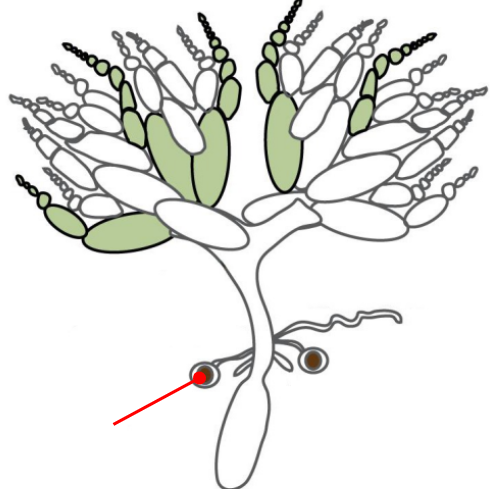
Ovaries
The gonads of a female Drosophila
The ovaries are connected by an oviduct and are made of units called ovarioles
The oviduct connects the ovaries to the uterus
The oviduct contains spermatheca, a specialized organ that allows the female to store sperm from the male for very long periods of time
The structure and function of the ovaries is what defines the axis of the animal!
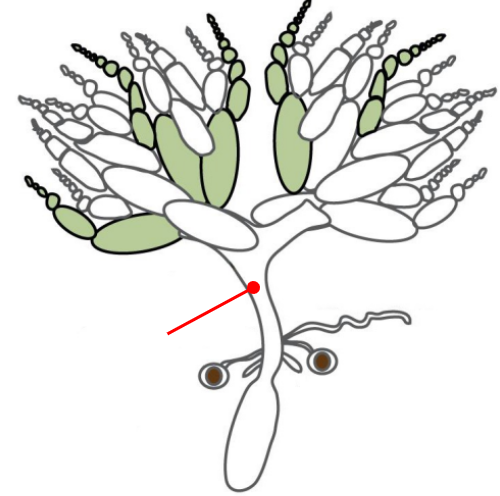
Oviduct
Connects the ovaries to the uterus
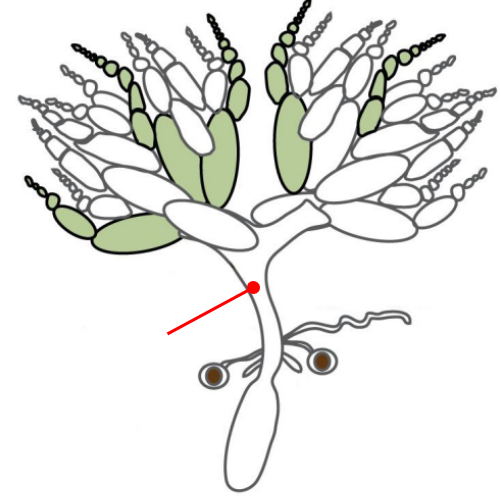
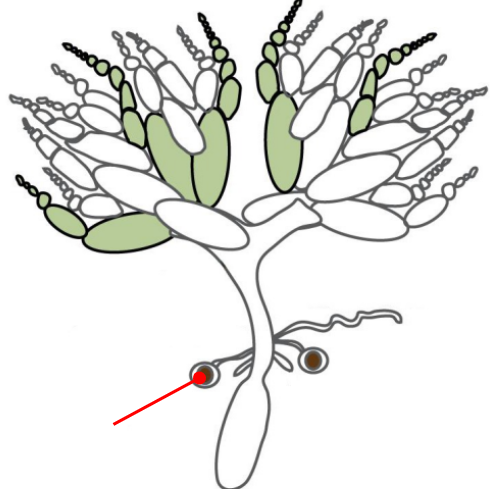
Spermatheca
A specialized organ connected to the oviduct that allows the female Drosophila to store the sperm from a male for very long periods of time
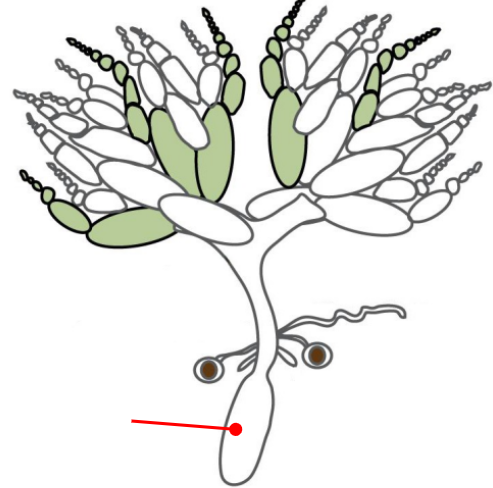
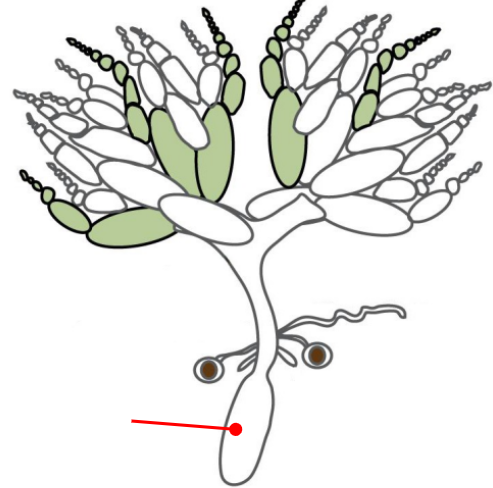
Uterus
Connected to the ovaries via the oviduct
Ovariole
The units that make up the ovaries
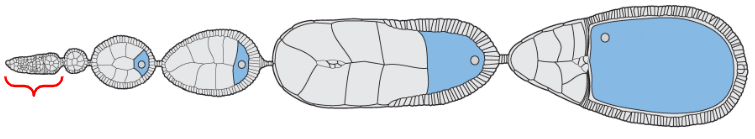
Oogonium
Stem cells present at the tip of each ovariole
Undergo 4 rounds of cell division to form 16 cells (1 oocyte progenitor and 15 nurse cells)
The 16 cells generated by incomplete oogonium division are connected and open to each other via ring canals
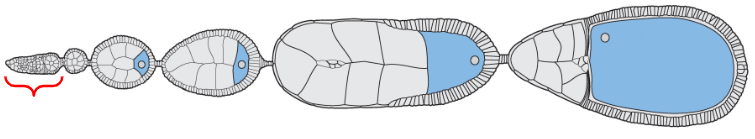
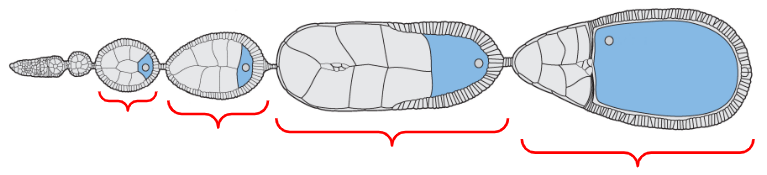
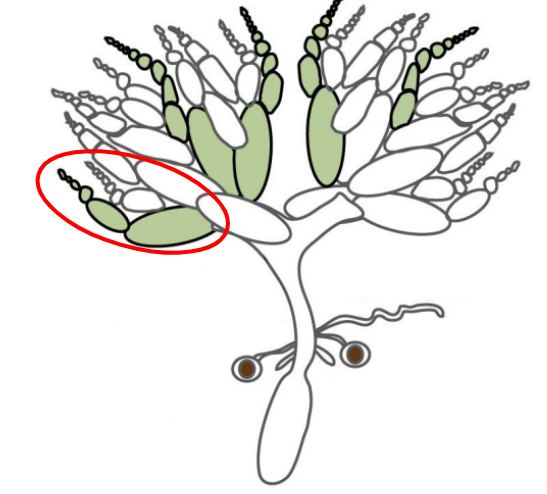
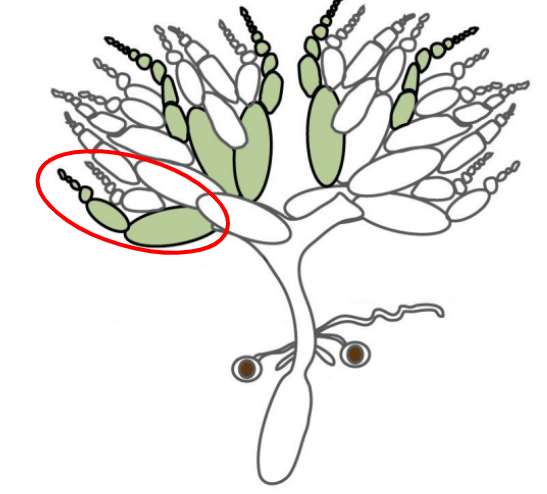
Egg chambers
Individual compartments within an ovariole that are surrounded by follicular epithelium
There are 15 nurse cells near the anterior end of the egg chamber
There is 1 large oocyte progenitor near the posterior end of the egg chamber
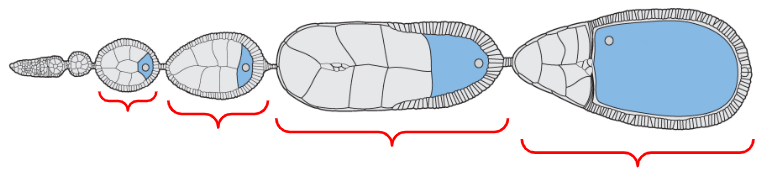
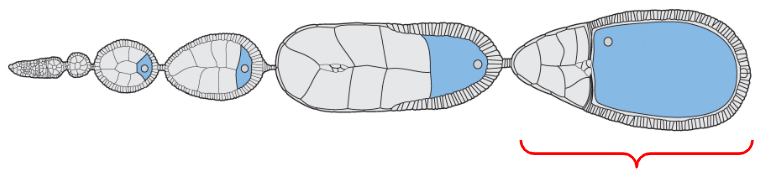
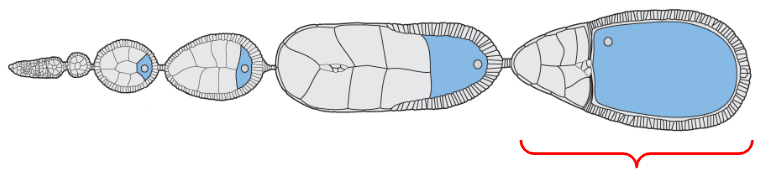
Mature form of the egg
The egg chamber located at the proximal base of the ovariole, closest to the oviducts
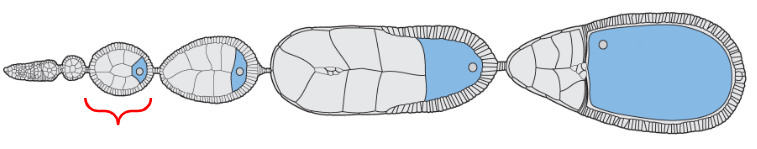
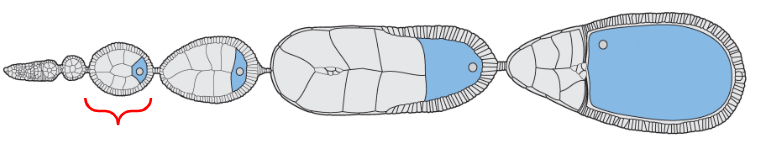
Earliest form of the egg
The egg chamber located near the distal tip of the ovariole but proximal to the oogonium (stem cells)
Ring canals
In Drosophila, the division of the oogonium (stem cells) is incomplete, resulting in cells that remain connected through a ring canal structure
A highly specialized opening that facilitates the transport of large molecules, such as mRNA transcripts
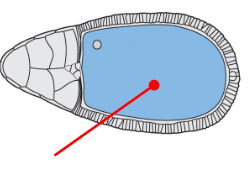
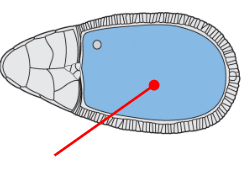
Oocyte progenitor
The progenitor of the egg (oocyte)
Completely transcriptionally silent until sadkfl;dsfl
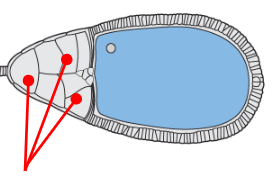
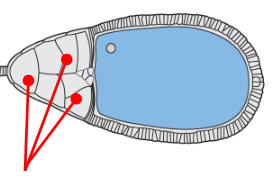
Nurse cells
Care and provide for the oocyte progenitor, transcribing various maternal mRNAs that will later be moved to the oocyte progenitor
Gurken
A maternal mRNA that is critical for specifying the dorsal, ventral, anterior, and posterior axes of the embryo
Drosophila is a great model organism for ______ genetic screens.
Drosophila is a great model organism for forward genetic screens (which involve random mutagenesis to create mutations throughout the genome, analysis for mutant phenotypes, and positional cloning linkage analysis to determine the identity of the gene that was mutated)
Nurse cells are located in the ____ side of the egg chamber.
Nurse cells are located in the anterior side of the egg chamber.
The oocyte progenitor is located in the ____ side of the egg chamber.
The oocyte progenitor is located in the posterior side of the egg chamber.
Describe how gurken mRNA is transported to the oocyte progenitor during oogenesis.
The embryo uses microtubules to transport gurken mRNA from the nurse cells to the oocyte progenitor
Within the egg chamber:
The plus ends of the microtubules are located at the anterior end (where the nurse cells are)
The minus ends of the microtubules are located at the posterior end (where the oocyte progenitor is)
The nurse cells transcribe gurken mRNA, which is delivered by dynein, a motor protein that moves from the plus (+) end to the minus (-) end of the microtubules
All 16 cells (the 15 nurse cells and 1 oocyte progenitor) are connected and open to each other through the ring canals
The ring canals facilitate the transport of gurken mRNA across cell membranes
Squid is a 3’ UTR-binding protein that is an intermediary between the gurken mRNA and the dynein motor protein
Squid binds to dynein and binds to the 3’ UTR of gurken mRNA, but Squid also inhibits the translation of gurken mRNA
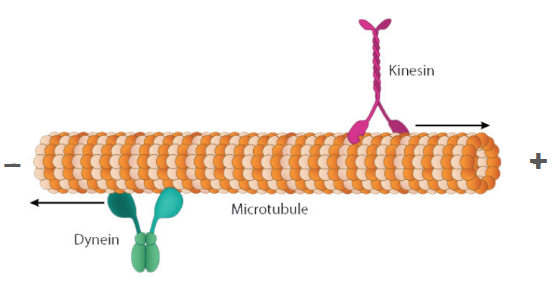
Compare the plus vs. minus end of a microtubule, and identify the direction that dynein and kinesin (microtubule motor proteins) travel in.
Plus end → the direction of growth and polymerization, where tubulin subunits are incorporated into the growing microtubule
Minus end → the location where the microtubule organizing center (MTOC) forms
Dynein → a microtubule motor protein that moves from the plus end to the minus end (+ → -)
Kinesin → a microtubule motor protein that moves from the minus end to the plus end (- → +)
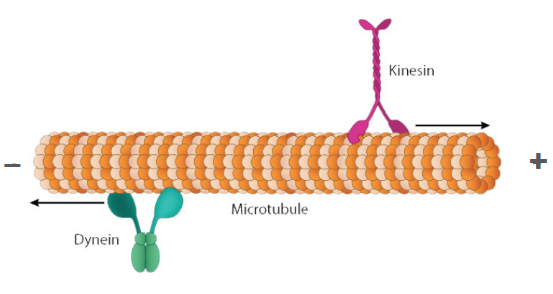
Plus end
The direction of microtubule growth and polymerization (where incoming tubulin subunits are added)
Minus end
The location of the microtubule organizing center (MTOC)
Kinesin
A microtubule motor protein that moves in the direction of the plus end (- → +)
Dynein
A microtubule motor protein that moves in the direction of the minus end (+ → -)
Centrosome
The element in the microtubule organizing center (MTOC) that nucleates the microtubules
Microtubules
Components of the cytoskeleton that act like highways, providing paths for the transportation of different components in the cell
Describe how the anterior-posterior axis and dorsal-ventral axis is established during oogenesis.
Gurken mRNA is produced by nurse cells and is transported on microtubules (with the help of the dynein motor protein and the Squid 3’ UTR-binding protein) in the minus-end direction, accumulating in the most posterior side of the oocyte progenitor and attaching to the nuclear envelope in the cytoplasm
At the most posterior end of the oocyte progenitor, there is local translation of gurken mRNA
Squid binds to the 3’ UTR of the gurken mRNA sequence, but also inhibits translation of gurken mRNA into protein
In order for translation of gurken to happen:
ORB binds to the gurken mRNA, triggering a conformational change in the gurken mRNA so that Squid dissociates
Casein kinase 2 (CK2) phosphorylates and activates ORB, and the activated ORB also recruits Wispy, a polyadenylate polymerase that adds a poly-A tail to the gurken mRNA
The addition of the poly-A tail to the 3’ end of the gurken mRNA by Wispy makes the mRNA more stable, protects it against degradation, and is the signal to initiate translation of gurken mRNA into protein
Gurken protein is locally translated in the cytoplasm of the posterior side of the oocyte progenitor and binds to the Torpedo receptor present on nearby cells, triggering the formation of the posterior follicular epithelium (the cells of the follicular epithelium are anterior by default)
Torpedo is a tyrosine kinase receptor expressed in all follicular epithelial cells, but only the Torpedo receptors that are in close proximity to the secreted Gurken protein will bind Gurken and be activated
After the specification and differentiation of the posterior follicular epithelium, the PFE sends an unknown signal that causes the microtubules in the oocyte progenitor to flip and reverse direction
Previously, the microtubules nucleated from the posterior towards the anterior side of the oocyte progenitor, away from the nucleus located at the posterior end (plus ends pointed away from the oocyte progenitor and towards the nurse cells)
Now, the microtubules have reorganized and nucleate from the anterior and lateral sides of the oocyte progenitor towards the nucleus, which is located at the posterior end (plus ends pointed towards the nucleus of the oocyte progenitor, minus ends pointed towards the lateral and anterior sides of the oocyte progenitor)
The reorganization of microtubules generates a huge amount of force in the cell that pushes the nucleus towards the anterior side of the oocyte progenitor
Dynein transports gurken mRNA (with Squid bound) in the minus end direction, towards the anterior side of the oocyte progenitor → gurken mRNA accumulates on the anterior side of the oocyte progenitor, in either corner
Dynein transports bicoid mRNA (with an unspecified protein bound) in the minus end direction, towards the anterior side of the oocyte progenitor → bicoid mRNA accumulates along the anterior side of the oocyte progenitor
Kinesin transports nanos mRNA in the plus end direction, towards the posterior side of the oocyte progenitor → nanos mRNA accumulates along the posterior side of the oocyte progenitor
Once the gurken mRNA is transported to one of the anterior corners of the oocyte progenitor, the gurken mRNA attaches to the nuclear envelope and there is local translation of gurken
ORB binds to the gurken mRNA, inducing a conformational change that causes Squid to release from the gurken mRNA
Casein kinase 2 (CK2) phosphorylates and activates ORB, and the activated ORB recruits Wispy, a polyadenylate polymerase that adds a poly-A tail to the 3’ end of the gurken mRNA
The polyadenylation of the gurken mRNA stabilizes the mRNA, protects it from degradation, and is a trigger for the local translation of gurken mRNA
Cells that receive the Gurken signal develop into the dorsal follicular epithelium (the cells of the follicular epithelium are ventral by default):
Gurken binds to the tyrosine kinase receptor Torpedo, and the activated torpedo phosphorylates Capicua (a transcriptional repressor of mirror), inactivating Capicua
The inactivated Capicua cannot enter the nucleus, so mirror (a transcriptional repressor of pipe) is expressed
Pipe is a sulfotransferase that modifies the ECM of follicular epithelial cells that have never been exposed to gurken
Because mirror is expressed, pipe is not expressed, so these cells differentiate into the dorsal follicular epithelium
Cells that do not receive the Gurken signal develop into the ventral follicular epithelium (the cells of the follicular epithelium are ventral by default):
Since Gurken is absent and does not bind to Torpedo, Torpedo remains inactive and does not phosphorylate Capicua (a transcriptional repressor of mirror), so Capicua remains active
Because Capicua is active, it can enter the nucleus and repress the transcription of mirror (a transcriptional repressor of pipe), so pipe is expressed
Pipe is a sulfotransferase that modifies the ECM and VE of follicular epithelial cells that have never been exposed to gurken
Because pipe is expressed, the ECM of these cells will be modified and these cells will develop into the ventral follicular epithelium
Squid
A 3’ UTR-binding protein that is an intermediary between the gurken mRNA and the dynein motor protein
Binds to dynein and binds to the 3’ UTR of gurken mRNA, but also inhibits the translation of gurken mRNA
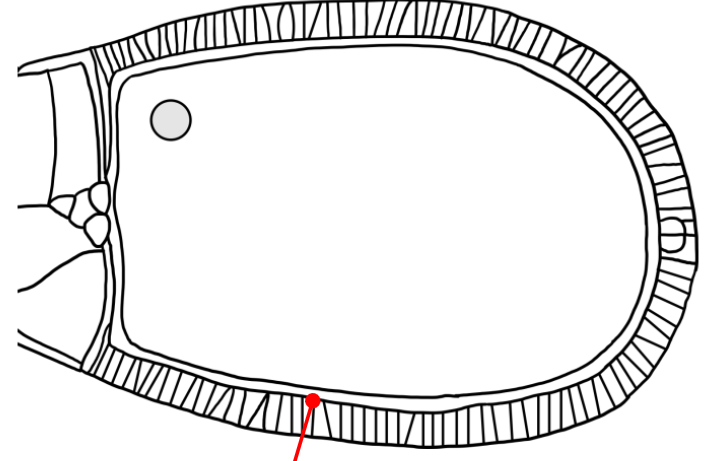
Which layer is indicated in the figure?
The cell membrane of the follicular epithelial cells
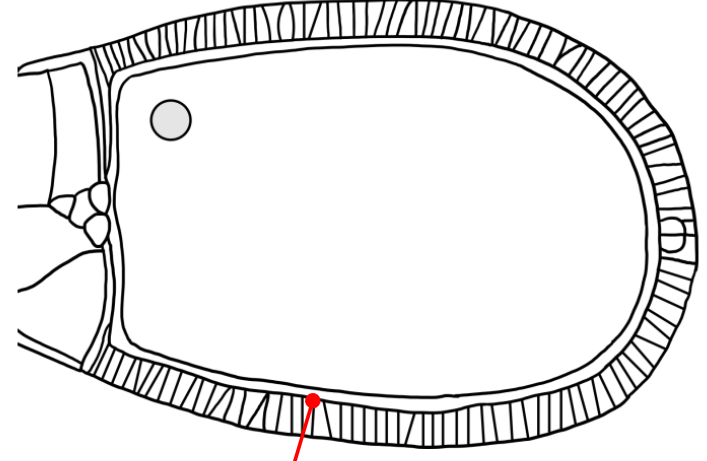
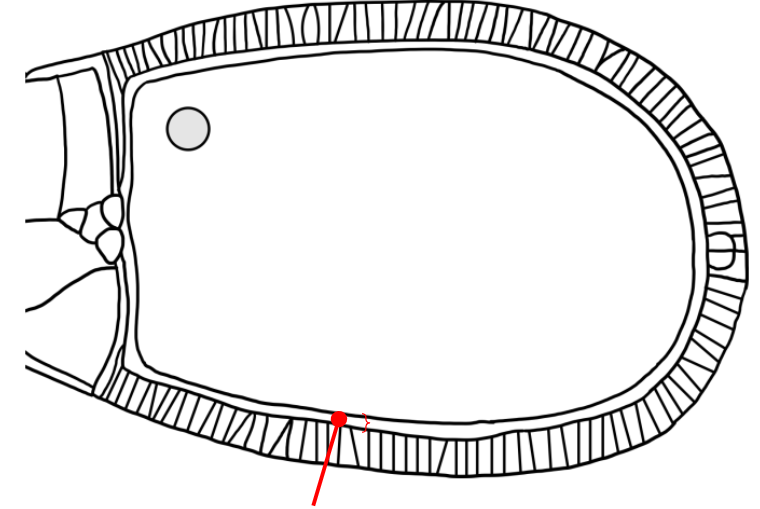
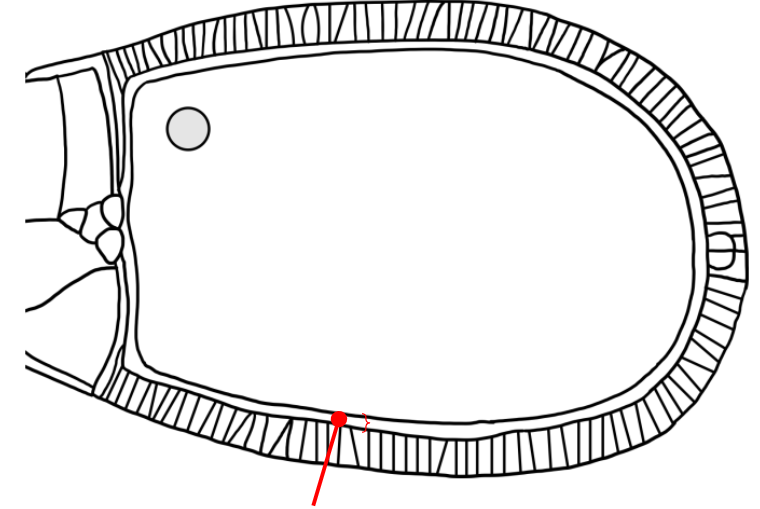
Which layer is indicated in the figure?
A space that is similar to the mesh-like vitelline envelope of the sea urchin egg
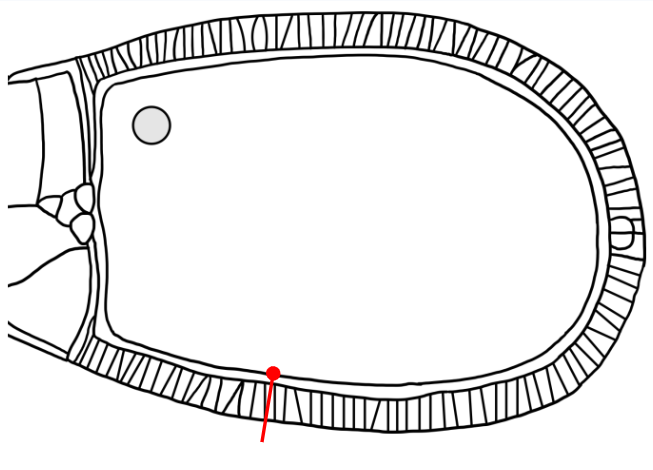
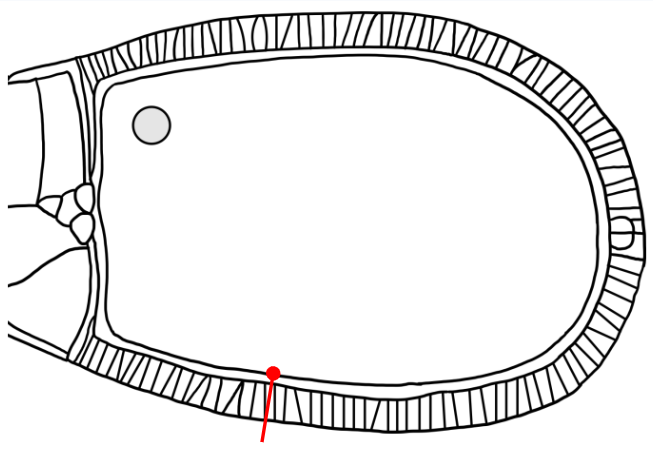
Which layer is indicated in the figure?
The egg membrane (the membrane of the oocyte progenitor)
Describe the specific cellular locations where the gurken protein binds to the torpedo receptor.
Gurken is a secreted protein translated in the cytoplasm (yolk) of the oocyte progenitor and secreted into the vitelline envelope-like layer. The torpedo receptor is expressed on the surface of follicular epithelial cells, and extends into the vitelline envelope-like layer, where gurken can bind as a ligand and trigger either the formation of the posterior follicular epithelium or the formation of the dorsal follicular epithelium.
Describe how the dorsal-ventral axis is established during oogenesis.
Once the gurken mRNA is transported to one of the anterior corners of the oocyte progenitor, the gurken mRNA attaches to the nuclear envelope and there is local translation of gurken
ORB binds to the gurken mRNA, inducing a conformational change that causes Squid to release from the gurken mRNA
Casein kinase 2 (CK2) phosphorylates and activates ORB, and the activated ORB recruits Wispy, a polyadenylate polymerase that adds a poly-A tail to the 3’ end of the gurken mRNA
The polyadenylation of the gurken mRNA stabilizes the mRNA, protects it from degradation, and is a trigger for the local translation of gurken mRNA
Cells that receive the Gurken signal develop into the dorsal follicular epithelium (the cells of the follicular epithelium are ventral by default):
Gurken binds to the tyrosine kinase receptor Torpedo, and the activated torpedo phosphorylates Capicua (a transcriptional repressor of mirror), inactivating Capicua
The inactivated Capicua cannot enter the nucleus, so mirror (a transcriptional repressor of pipe) is expressed
Pipe is a sulfotransferase that modifies the ECM of follicular epithelial cells that have never been exposed to gurken
Because mirror is expressed, pipe is not expressed, so these cells differentiate into the dorsal follicular epithelium
Cells that do not receive the Gurken signal develop into the ventral follicular epithelium (the cells of the follicular epithelium are ventral by default):
Since Gurken is absent and does not bind to Torpedo, Torpedo remains inactive and does not phosphorylate Capicua (a transcriptional repressor of mirror), so Capicua remains active
Because Capicua is active, it can enter the nucleus and repress the transcription of mirror (a transcriptional repressor of pipe), so pipe is expressed
Pipe is a sulfotransferase that modifies the ECM and VE of follicular epithelial cells that have never been exposed to gurken
Because pipe is expressed, the ECM of these cells will be modified and these cells will develop into the ventral follicular epithelium
Describe how the anterior-posterior axis is established during oogenesis.
Gurken mRNA is produced by nurse cells and is transported on microtubules (with the help of the dynein motor protein and the Squid 3’ UTR-binding protein) in the minus-end direction, accumulating in the most posterior side of the oocyte progenitor and attaching to the nuclear envelope in the cytoplasm
At the most posterior end of the oocyte progenitor, there is local translation of gurken mRNA
Squid binds to the 3’ UTR of the gurken mRNA sequence, but also inhibits translation of gurken mRNA into protein
In order for translation of gurken to happen:
ORB binds to the gurken mRNA, triggering a conformational change in the gurken mRNA so that Squid dissociates
Casein kinase 2 (CK2) phosphorylates and activates ORB, and the activated ORB also recruits Wispy, a polyadenylate polymerase that adds a poly-A tail to the gurken mRNA
The addition of the poly-A tail to the 3’ end of the gurken mRNA by Wispy makes the mRNA more stable, protects it against degradation, and is the signal to initiate translation of gurken mRNA into protein
Gurken protein is locally translated in the cytoplasm of the posterior side of the oocyte progenitor and binds to the Torpedo receptor present on nearby cells, triggering the formation of the posterior follicular epithelium (the cells of the follicular epithelium are anterior by default)
Torpedo is a tyrosine kinase receptor expressed in all follicular epithelial cells, but only the Torpedo receptors that are in close proximity to the secreted Gurken protein will bind Gurken and be activated
After the specification and differentiation of the posterior follicular epithelium, the PFE sends an unknown signal that causes the microtubules in the oocyte progenitor to flip and reverse direction
Previously, the microtubules nucleated from the posterior towards the anterior side of the oocyte progenitor, away from the nucleus located at the posterior end (plus ends pointed away from the oocyte progenitor and towards the nurse cells)
Now, the microtubules have reorganized and nucleate from the anterior and lateral sides of the oocyte progenitor towards the nucleus, which is located at the posterior end (plus ends pointed towards the nucleus of the oocyte progenitor, minus ends pointed towards the lateral and anterior sides of the oocyte progenitor)
The reorganization of microtubules generates a huge amount of force in the cell that pushes the nucleus towards the anterior side of the oocyte progenitor
Dynein transports gurken mRNA (with Squid bound) in the minus end direction, towards the anterior side of the oocyte progenitor → gurken mRNA accumulates on the anterior side of the oocyte progenitor, in either corner
Dynein transports bicoid mRNA (with an unspecified protein bound) in the minus end direction, towards the anterior side of the oocyte progenitor → bicoid mRNA accumulates along the anterior side of the oocyte progenitor
Kinesin transports nanos mRNA in the plus end direction, towards the posterior side of the oocyte progenitor → nanos mRNA accumulates along the posterior side of the oocyte progenitor