1) V ATP synthase du thylacoide
1/8
Earn XP
Description and Tags
204 du cours 1
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
9 Terms
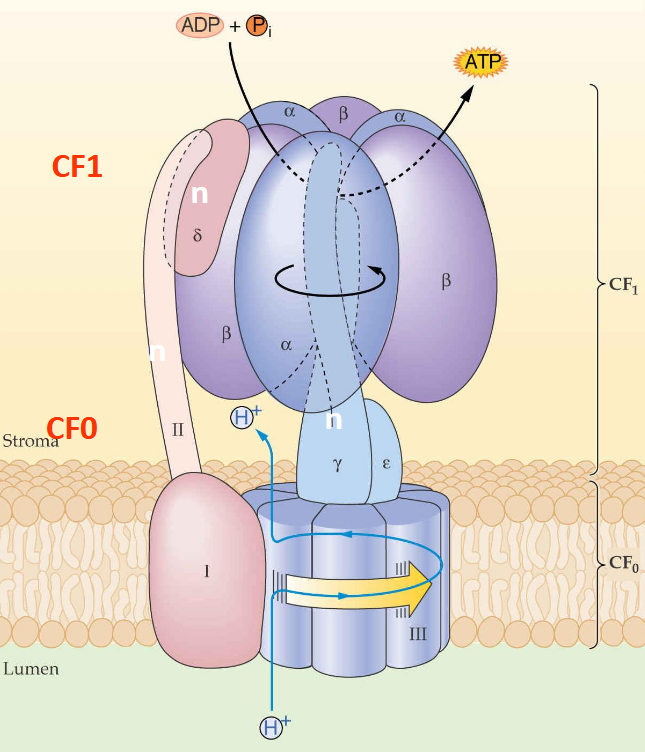
structure de l'ATP Synthase (F-Type ATP Synthase):
CF0 et CF1 :
CF0: Partie transmembranaire insérée dans la membrane du thylakoïde, formant un canal pour les protons H+H^+H+.
CF1: Partie catalytique située dans le stroma, responsable de la synthèse de l'ATP. → soluble.
Sous-unités :
Les sous-unités sont notées α, β, γ, δ, ε, I, II, III, IV.
γ (gamma) : régulée par changements redox (Fd)
δ (delta) : Assure le lien entre CF0 et CF1.
ε (epsilon) : Rôle de régulation, arrête l'activité catalytique dans l'obscurité.
III = port pour les H+
comment est fournie la force motrice ?
La différence de concentration de protons (gradient électrochimique) entre le lumen (pH 5) et le stroma (pH 8) génère une force motrice.
Les protons passent à travers CF0, ce qui induit la rotation de la sous-unité γ.
Cette rotation fournit l'énergie nécessaire pour convertir l'ADP + Pi en ATP dans CF1
ATP synthase role
dynamo / moteur pour produire ATP
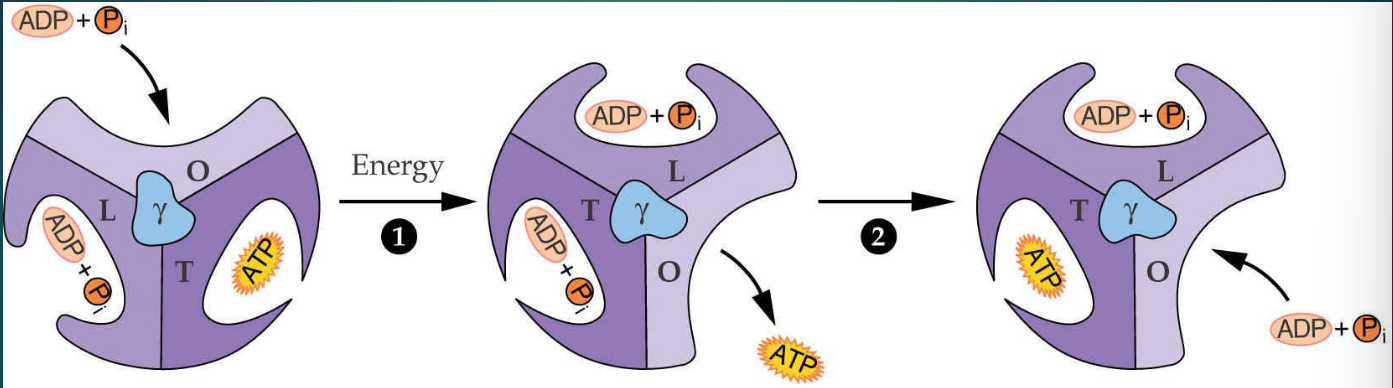
conformation de l’ATP synthase
Site O (Open) - Site ouvert :
Permet la libération de l'ATP déjà formé et l'entrée de nouveaux substrats (ADP + pi).
Faible affinité pour les ligands.
Site L (Loose) - Site lâche :
Capture et maintient l'ADP = Pi dans le site actif.
Les substrats sont liés sans être convertis en ATP.
Site T (Tight) - Site serré :
Conformation qui permet la formation d'ATP par condensation de l'ADP + pi
Le mouvement de γ est directement entraîné par le flux de protons (H⁺) qui traverse le complexe CF0, situé dans la membrane du thylakoïde.
expérience de jagendorf et Uribe
Les thylakoïdes isolés des chloroplastes sont placés dans un milieu acide (pH 4) pendant 60 secondes. Cela permet de créer un gradient de protons très élevé à l'intérieur des thylakoïdes (accumulation de H⁺ dans le lumen).
Changement de milieu :
thylakoides transférés dans un milieu basique (pH 8) avec :
ADP
Phosphate inorganique (Pi)
Alcalin pour neutraliser l'excès de protons
Résultat :
Immédiatement après le transfert (15 secondes), la synthèse d'ATP est détectée.
→ le gradient de protons (différence de pH entre l'intérieur et l'extérieur) suffit à générer de l'ATP, même sans apport direct d'énergie lumineuse.
L'ATP synthase utilise le retour des H⁺ du lumen vers le stroma pour phosphoryler l'ADP en ATP.

2 types de transfert d’e-
linéaire
cyclique
Ratio du transfert linéaire des e- via la chaine photosynthétique
ratio fixe de NADPH/ATP = 3/2.
Problème : Ce ratio ne correspond pas toujours aux besoins du cycle de Calvin (souvent besoin de plus d'ATP que de NADPH).
transfert cyclique des e-
PSII déconnecté : pas de production d'O2 ni de libération d'électrons à partir de l'eau.
Cycle cyclique autour du PSI :
Les électrons sont renvoyés vers le cytochrome b6f puis récupérés par le Plastocyanine (PC).
Ils retournent vers le PSI sans passer par le PSII.
Cela génère un pompage de protons dans le lumen → augmentation du gradient de protons → production d'ATP par l'ATP synthase.
Pas de NADPH produit.
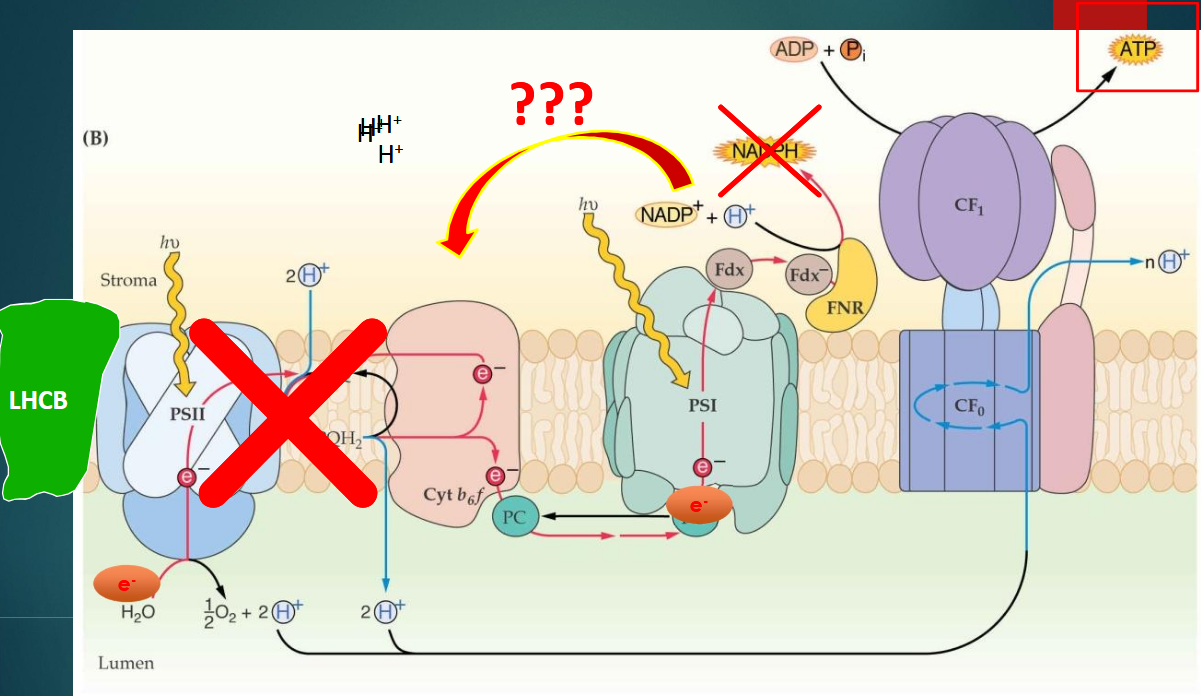
qu’est ce que la transition d’état entre transfert linéaire et cyclique
linéaire → Cyclique : La transition se fait pour ajuster le rapport ATP/NADPH en fonction des besoins métaboliques.
LHCII (Light Harvesting Complex II) :
Ces antennes peuvent se déplacer entre le PSI et le PSII pour moduler l'efficacité de capture de lumière.
Cela permet de compenser les déséquilibres énergétiques entre les deux photosystèmes.