M7L2 - Microtubules
1/26
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No study sessions yet.
27 Terms
Microtubule Structure
Comprised of 13 protofilaments
Arrayed circularly to form a tube wall
They’re staggered to resemble a spiral
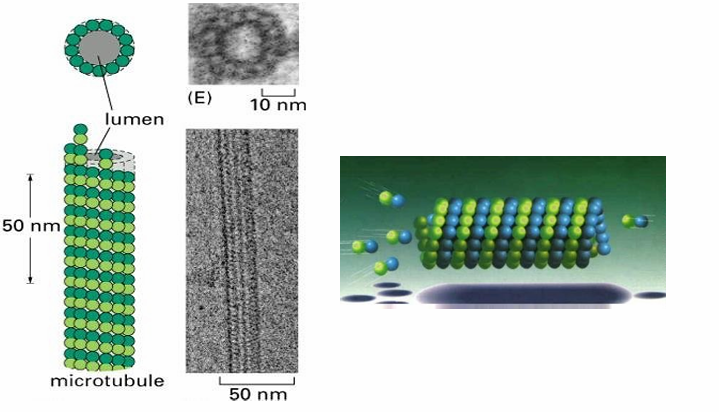
What are the basic subunits of each protofilament (microtubule structure)
Dimers of alpha and beta tubulin proteins
What are the GTP-binding properties of α- and β-tubulin subunits?
Both α- and β-tubulin bind GTP.
α-tubulin: GTP is tightly bound
never hydrolyzed
does not exchange with free nucleotides.
β-tubulin: GTP is loosely bound
hydrolyzed to GDP
exchanged for GTP in the cytosol.
How are tubulin subunits added and removed during microtubule assembly?
α- and β-tubulin subunits are added/removed as dimers.
αβ–GTP dimers have a higher affinity for the growing microtubule (more stable).
αβ–GDP dimers have a lower affinity and tend to dissociate from the filament.
Microtubule Polarity
They’re polar so the two ends have different characteristics and dynamics
(+) end = fast growing
(-) = slow growing
Within the dimers
the beta-subunit is closer to (+)
the alpha-subunit is closer to (-)
Microtubule Dynamics
Dimers with αβ–GTP are added to (+) end
Rescue phase
Dimers with αβ–GDP are released from shrinking filament
Catastrophe
GTP hydrolysis occurs within polymerized microtubule
Most of it consists of dimers containing αβ–GDP
(+) has GTP cap (unhydrolyzed) which favours growth
αβ–GTP dimers have a 4x slower disassociation rate in comparison to αβ–GdP
They thus have higher affinity for their neighbours and stay together
(+) end has dynamic instability
Oscillates between growth or shortening
High [GTP-tubulin] = polymerization
Low [GTP-tubulin] = depolymerization
EB1 Protein (Microtubule)
Plus-end binding protein
Prevents premature catastrophes
Acts as positive regulator of microtubule growth
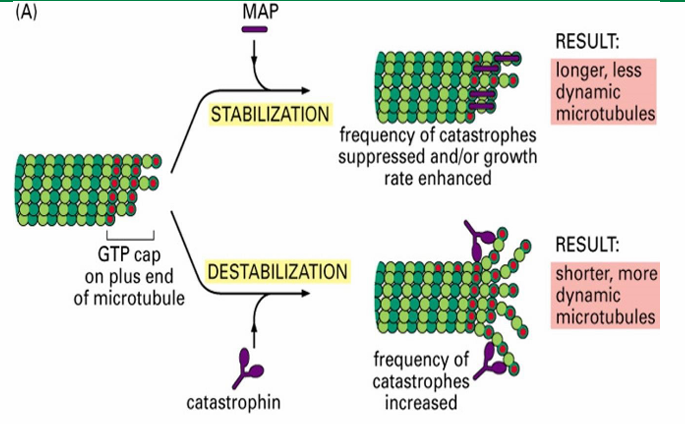
MAPs - Microtubule Associated Proteins
Proteins controlling the assembly and disassembly of microtubules
MAPs - Microtubule Associated Proteins (Function)
Interconnect microtubules to form bundles
Inc stability
alter rigidity
influence assembly rate
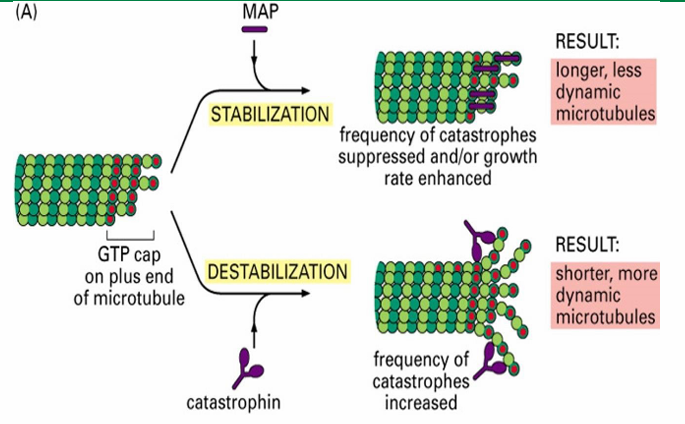
MAPs - Microtubule Associated Proteins (Two Groups)
Those that stabilize microtubules (Ex. Tau and EB1)
Those that destabilize microtubules (Ex. catastrophin)
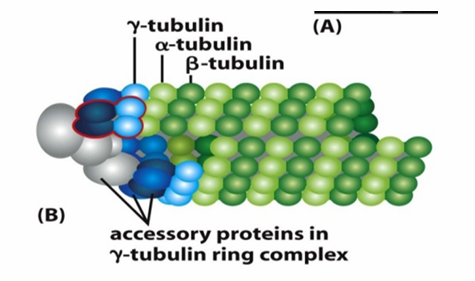
Microtubule Nucleation
Starting off growth
Involves γ‐tubulin which is present in smaller amounts in the cell compared to alpha/beta tubulin
Helps form γ‐tubulin ring complex (γ-TuRC)
Nucleates at (-) end of a new microtubule
Forms a template for the growing (+) end
γ-TuRC acts as a cap of the (-) end while microtubule growth occurs at (+) end
MTOC (Microtubule Organizing Center)
A specific location inside the cell where microtubule nucleation occurs
In animal cells, the MTOC is centrosome (red dot)
Located near nucleus
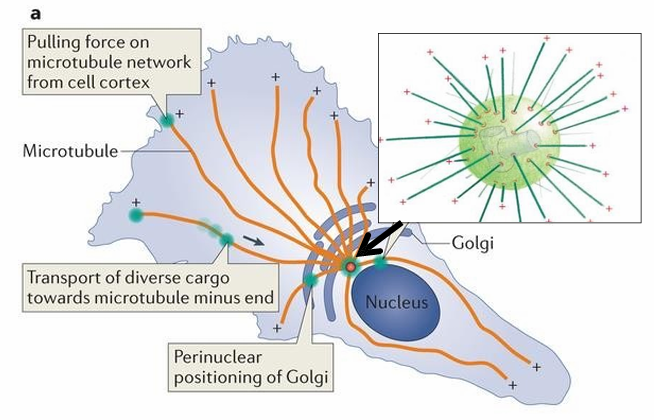
MTOC: Centrosome and yTuRC
Consists of 2 cylindrical structures called centrioles (inside centrosome which is in green)
Also has pericentriolar material (PCM) containing many γ‐TuRC complexes (red rings on green ball)
(-) end of microtubules are nucleated at the γ‐TuRC
(+) end are directed towards the cell periphery (shown as +)
MTOC role in Mitosis
The MTOC (centrosome) organizes microtubules that form the mitotic spindle.
The spindle’s microtubules attach to chromosomes to separate replicated sister chromatids.
Centrosomes are duplicated before mitosis, creating two MTOCs that move apart to opposite poles.
Microtubules nucleate from the γ-TuRC complexes at each MTOC, with plus ends growing outward.
Microtubule Toxins: Cholchicine
Useful in lab to arrest the cell cycle
Ex. cholchicine
Derived from meadow saffron
Inhibits polymerization
Binds and stabilizes αβ‐tubulin dimers
Prevents addition/loss of tubulin dimers
Arrests cells in metaphase without chromatid seperation
Microtubule Toxins: Taxol
Useful in lab to arrest the cell cycle
Taxol Function
Binds to β‐tubulin to increase affinity for (+) end
Prevents depolymerization
Prevents assembly of mitotic spindle to inhibit mitosis
Used in cancer treatment
Hard to synthesize in lab so it’s derived from pacific yew tree
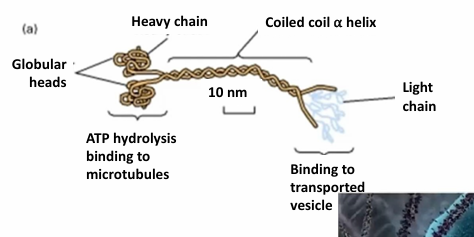
Kinesin Motor Protein
(+) directed transport on microtubules, so towards cell periphery away from MTOC
Tetrameric complex made of 2 heavy chains and 2 light chains
The globular heads (motor domains) cyclically bind to microtubules
Generates movement through ATP hydrolysis
The tails determine specificity of cargo binding
The tails are highly variable
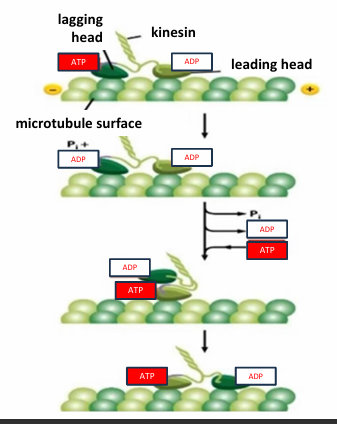
Kinesin Mechanochemical Cycle
The lagging head is bound to ATP
The leading head is bound to ADP
ATP kinesin has a higher affinity for the microtubule than ADP bound kinesin
The ATPase motor lagging head hydrolyzes ATP to ADP + Pi
Reduces affinity of lagging head for microtubule
ADP is exchanged for ATP in leading head
Increases affinity of leading head
The binding of ATP induces conformational change causing lagging head to swing in front to another microtubule binding site
This resets cycle to the top
How does kinesin move along microtubules?
Kinesin moves in a “hand-over-hand” fashion.
It has two motor heads (domains), and one is always attached to the microtubule.
The two heads work in a coordinated cycle, each in a complementary stage of ATP binding or hydrolysis.
In-vitro assays for kinesin movement
Nomarski Microscope
Following plastic beads tethered to kinesin
The track is anchored to the thing made from purified tubulin
Gliding mobility assay
kinesin are tethered to a glass slide at their cargo (Tail) ends
They can then move fluorescently labeled microtubules added to solution above slide
Dynein
(-) directed, moving towards MTOC
2 main forms: Cytoplasmic and Axonemal
Has 2 heavy chains and a variety of intermediate and light chains
Two forms of dynein
Cytoplasmic
Associated with microtubules
Direct movement of organelles and vesicles in cytoplasm
Axonemal
Found in structures powering movement of whole cells
Ex. cilia or flagella
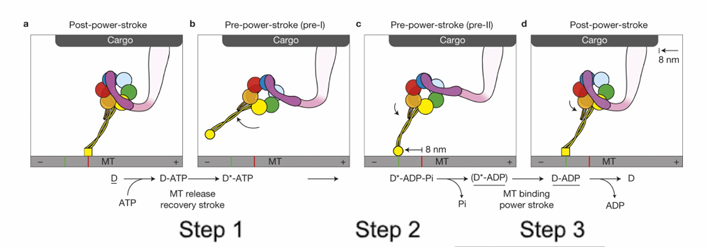
How does dynein move cargo along microtubules?
Movement is powered by a power stroke in the linker arm (near the cargo attachment site).
In a dynein dimer, the two motor units alternate power strokes, producing continuous movement.
Describe the steps of dynein’s ATP-driven power stroke
ATP binding releases motor head group from microtubule
ATP hydrolysis creates dynein-ADP+Pi that can now attach to the microtubule
The release of Pi powers the power-stroke of the liner
Pulls the cargo
Each power stroke, the cargo moves towards the (-) end by 8mm
Bidirectional Vesicle Movement: Neural Cells
Microtubules span the axons of neural cells
The (-) ends are anchored to MTOC
The (+) ends extend along the axons towards synapse cell membrane
Vesicles with NTs are carried from cell body to synapse along microtubules
Microtubule Tug of War
Model describing the movement of proteins if they’re bidirectionally transported
The final direction of movement is the winner of this ‘battle’
There are regulatory proteins controlling direction in response to cell signals
Change of Direction (Microtubule Transport) Application: Melanosomes in Fish
Melanosomes: Pigment-filled organelles
Movement of it changes skin cells in response to behavioural signalling
This movement is done by molecular motors carrying it to the cell periphery or center
Dynein: Move towards (-) end MTOC
Kinesin: Move towards cell periphery (+) end
Dispersion to periphery = cell appears darker
Concentrated in middle = Cell appears lighter
This is controlled by signals using cAMP as a secondary messenger