Mathematical Biology
1/93
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
94 Terms
Continuous-time Models
Population dynamics that occur continuously in time
Ordinary differential equations
The Malthusian Model
Describe and predict the growth of the human population in England
N′(t) = bN − dN = (b − d)N = rN,
The Verhulst Model
Growth of a population is assumed to slow down as the population becomes larger due to crowding effects
N’(t) = r(1-(N(t)/K))
Steps of nondimensionalisation
Determine units of all parameters
Rewrite dependent and independent variables as nondimensional quantities
Find suitable choices for the undetermined constant
Steady State
Also referred to as equilibria of the system
Values N = N* that satisfy f(N*) = 0.
States the system cannot leave -N′(t) = 0 if N = N*
Types of steady states
stable if f’(N*) < 0
unstable if f’(N*) > 0
inconclusive if f’(N*) = 0
Hysterisis
history-dependence
The state a system is in not only depends on the current environmental conditions but also on which states the system has been in previously
Tipping point
A small environmental change leads to large changes in an ecosystem that cannot be reversed
Delay-differential equation
Accounts for delays in a system
dN(t)/dt = f (N(t), N(t − T))
Discrete-time models
Used to describe biological systems in which the time-continuous nature of ODE models is inappropriate
Difference equations
Nt+1 = f(Nt)
Nt+1 = rNt , r ≥ 0.
Steady state of difference equations / types
N* = f(N*)
Linearly stable if |f’(N*)| > 1
Linearly unstable if |f’(N*)| < 1
Types of (un)stability for discrete-time models
oscillatory stable if −1 < f′(N*) < 0
monotonically stable if 0 < f′(N*) < 1
oscillatory unstable if f ′(N*) < −1
monotonically unstable if f ′(N*) > 1
Cobweb diagram
Graphically determine steady states, stability and other solution behaviour
Enzyme reaction
enzymes combine with a substrate to form a complex that is subsequently degraded into products and the enzymes
Catalysts
they accelerate or inhibit a chemical reaction but are not degraded by doing so
The Law of Mass action
The rate of a biochemical reaction is proportional to the product of the concentrations of the reactants
Singularly perturbated problem
Problems in which “the most important term” is multiplied by a small constant ε ≪ 1
The implication is significant, setting ε = 0 does not provide an appropriate approximation
Method of matched asymptotic
Split the system into two simpler subsystems, analyse both and combine the results in a meaningful way.
Oscillations
Not all reactions lead to a gradual decay of a substrate and gradual increase of a product
Periodic solutions = limit cycles
The Poincaré-Bendixon Theorem
if there is a confined set in the system’s phase space that contains only a single steady state, and if that steady state is unstable, then solutions converge to a limit cycle
Confined set
A subset of the phase space that solution trajectories cannot leave
Stability of Steady states
If both eigenvalues λi ∈ C, i = 1, 2 of the Jacobian evaluated at the steady state (x, y) = (x∗,y∗) have negative real part, maxi=1,2 Re(λi) < 0, then the (x∗,y∗) is stable. If at least one eigenvalue has positive real part, maxi=1,2 Re(λi) > 0, then the steady state is unstable.
Hartman-Grobman Theorem
trajectories in a nonlinear system close to a steady state are well-approximated by trajectories of the corresponding linearised (about the same steady state) system, provided eigenvalues have non-zero real part.
Competitive interaction
occur when one population reduces the net growth of the other and vice versa
Interference competition
Direct competition for resources (e.g. fighting)
Uses the competitive Lotka-Volterra model.
Competitive Exclusion Principle
Coexistence of species is possible if every species exerts a higher competitive pressure on individuals in its own population than on individuals in other populations
(intraspecific competition among each species is stronger than the interspecific competition it exerts on its competitor)
Exploitation competition
occurs when species compete for a single limiting nutrient.
Tilman’s R* rule
The species that reduces the resource density to the lowest level at steady state outcompetes the other
Cooperation/ symbiosis
Occurs when one population promotes the growth of another and vice versa.
For example, birds that feed on berries help seed dispersal. Thus, the interaction is beneficial for both populations.
Predator-prey and host-parasite interactions
One population (the predator or parasite) benefits from the interactions with the other species (the prey or host).
The prey species, in turn, is harmed by these interactions.
Often feature oscillations with a slight lag
Jury Conditions
The eigenvalues λ1, λ2 ∈ C of the matrix J ∈ R2×2 satisfy |λ1| < 1 and |λ2| < 1, if and only if det(J) < 1, and 1 + det(J) > |tr(J)|
Compartmental models
The total population (humans, animals, plants) is split into different compartments, and over time, individuals can “move” around the compartments
SIR Model and Assumptions
three compartments: susceptibles (S), infected (I), and recovered (R)
S can become infected through contact with I, I can recover, and R immune from the infection.
assume that no birth and death occurs in the population
that immunity of recovered individual does not wane
Force of infection
The rate at which susceptibles become infected is λ = βI in this SIR model
Basic Reproduction number
R0 describes the average number of secondary infections caused by a single infected individual in an otherwise susceptible population
R0 > 1 leads to an epidemic
Extensions to the SIR model
Latent (L): individuals who have been exposed to the disease but are not yet infectious
Quarantined (Q): infected individuals that are subject to quarantine measures
Chronic (C): chronically ill individuals
Diseased (D): individuals can die
Vaccinated (V): individuals who are vaccinated
Individuals with maternally-derived immunity (M): offspring is initially immune to disease due to maternal antibodies
Vaccination and the SIR model
Assume that a proportion 0 ≤ p ≤ 1 of the population receives a vaccine and that this vaccine provides immunity for all recipients.
vaccination program prevents an epidemic provided that R0vacc < 1 ⇔ p > 1 − γ/βN = 1 − 1/R0 .
Vector-borne diseases
Many diseases are caused by parasites that spend periods of their life cycle in another organism, typically referred to as a vector
They can enter the blood stream of a human due to a bite from an infected mosquito
In turn, mosquitos become infected by biting an infected human. Thus, human-to-human infection occurs via mosquitos (vector)
Assumptions of vector-borne model
No recovered class → no immunity
disease dynamics are much faster than the human population dynamics and thus disregard any birth or death processes.
vectors are assumed to reproduce and die at same rate all newborn vectors are susceptible
2 Vector-borne steady states
disease-free steady state (iH, iV ) = (0, 0)
endemic steady state (iH, iV ) = (i∗H, i∗V ), where i∗H = (αHαV − 1)/ (αV(αH + 1)), i∗V = (αHαV − 1)/ αH(αV + 1)
Stability of vector-borne
(iH, iV ) = (0, 0) is stable if and only if αHαV < 1.
endemic steady state (iH, iV) = (i∗H, i∗V) is stable if and only if αHαV > 1
Ecological models
Provide information on the dynamics of (interacting) populations with pre-defined properties do not change over time.
Do not count for evolution.
Adaptive dynamics
A method that allows us to describe how model parameters evolve over time. Its main idea is to determine parameter values that represent a possible endpoint of evolution.
Evolutionary and Convergence stable
A strategy that cannot be invaded by other strategies
The eco-evolutionary dynamics can converge to it.
Fitness gradient
if ∂g/∂m |m=r
> 0 then the trait evolves towards larger values
< 0 then the trait evolves towards smaller values.
Evolutionary equilibrium
Whenever the fitness gradient is zero there is no directional selection in either direction occurring.
Can be stable (fitness maximum) if ∂2g/ ∂m2|m=r=r ∗ < 0, or unstable (fitness minimum) if ∂2g/ ∂m2|m=r=r ∗ > 0.
Pairwise Invasion plot
Plot the sign of the invasion fitness g(m; r) against r and m, with the resident trait r on the x-axis and the mutant trait m on the y-axis.
Plotted in 2d, with regions of g(m; r) > 0 and g(m; r) < 0 being distinguished through different shadings
Evolutionary stable on PIP
Evolutionary equilibria occur for trait values at which drawing a vertical line onto the PIP does not cause a change of sign(g(m; r)) as it passes through the diagonal m = r.
If sign(g(m; r)) ≤ 0 along the vertical line, then the evolutionary equilibrium is stable
If sign(g(m; r)) ≥ 0 along the vertical line, then the evolutionary equilibrium is unstable
Requirement for convergence stable
Close to the evolutionary equilibrium, directional selection for c < r∗ is positive and directional selection for c > r∗ is negative.
Convergence stable on PIP
If, for r = m < r∗ the invasion fitness is positive above the diagonal and negative below, and if for r = m > r∗ the invasion fitness is negative above the diagonal and positive below, then the evolutionary equilibrium r ∗ is convergence stable, and otherwise unstable.
Outcomes of evolution model
evolutionary stable and convergence stable - equilibrium presents a possible outcome of evolution
evolutionary stable and convergence unstable - no influence on long-term outcome
evolutionary unstable and convergence stable - evolutionary branching point
evolutionary unstable and convergence unstable - no influence on long-term outcome
Spatio-temporal dynamics
Consider both space and time
Can capture clusters of individuals
This leads to partial differential equations (PDEs) or integrodifference equations
Conservation equation
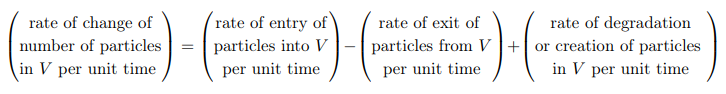
One-dimensional conservation eqn
The flux that changes the total population in V is that entering through the cross-section at x and leaving through the cross-section at x+ ∆x.
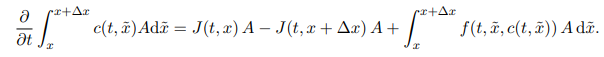
Fickian Diffusion
The random motion of individual molecules (random walk)
Nonlinear diffusion
diffusion parameter depends on population density (e.g. overcrowding)
Convection or Advection
Directed movement (e.g. pollen movement in a particular wind direction)
Taxi
Directed movement in response to an external chemical or physical signal, for example bacteria sensing and moving towards a food source.
Chemotaxi, Haptotaxi, Thermotaxi
movement directed by a chemical gradient
movement directed by a gradient in adhesive substances (often bacteria moving in response to extracellular matrix elements)
movement directed by a gradient of temperature
Infinite domain
the density is not influenced by the boundary
used when systems considered are so large that it can be safely assumed that dynamics in large parts of the system are unaffected by the boundary – for example vegetation dynamics in the desert.
Periodic BC
model densities are periodic functions
used when, for example, particles move on a torus, or also to mimic infinite domains in numerical simulations.
Dirichlet BC
density (concentration) is fixed at the boundary (but can still be a function of time and space)
For example, consider two reservoirs of chemical reactants placed at the ends of the domain, that are held at densities (concentrations) c1(t) and c2(t), respectively.
No-flux BCs
Particles cannot escape from the domain
Non-homogeneous Neumann BC
Flux on the boundary is given by some non-zero function.
Mixed Robin BC
A combination of Dirichlet and Neumann BCs. (Can be homogeneous or non-homogeneous)
Biological Waves
A quantity of interest (population density, chemical concentration, etc.) spreads throughout space over time.
e.g. spread of grey squirrels throughout UK
Travelling wave
If x ∈ R denotes space and t ∈ R denotes time, a function u(x, t) is a ____ if it can be expressed as
u(x, t) = u(z), z = x − vt, v ≠ 0
Pulse, Front, Periodic
Travelling ___: u(x, t) → a, as x → ±∞
Travelling ___: u(x, t) → a, as x → −∞, u(t, x) → b, as x → +∞ and a ≠ b.
___travelling wave: u(x, t) is a travelling wave that is a periodic function in x-
Fisher’s Equation
one-dimensional reaction-diffusion equation comprising a logistic growth term and standard diffusion
apply to any setting in which a population follows approximately logistic growth and moves randomly.
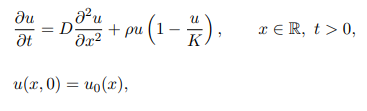
Bendixson-Dulac Theorem
If there exists a function φ(W, P), with φ ∈ C 1 (R 2 ), such that ∂(φF)/∂W + ∂(φG)/∂P has the same sign (≠ 0) in a simply connected region (region without holes), then the system dW/dz = F(W, P) , dP/dz = G(W, P) has no periodic solutions in this region.
Phase diagram
draw axis
draw nullclines
identify steady states = intersection of x/y nullclines
nullclines separate phase space into distinct regions, indicate signs of the rate of change
sketch a few example trajectories
Bifurcation diagrams
do model analysis
choose bifurcation parameter
draw axis with bifurcation parameter (horizontal) and solution measure (vertical)
Draw stable steady states
(optional) draw unstable steady states
add information on other asymptotic states (e.g. max/min)