Neural basis of working memory
1/23
Earn XP
Description and Tags
LTM vs STM, Prefrontal cortex as a possible STM storage buffer, what vs where in WM, evidence from multivoxel pattern analysis against the PFC storage buffer hypothesis. role for the PFC in attention to internal representation
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
24 Terms
recap - HM (1954) - bilateral removal of the temporal lobe
severe amnesia → inability to form long term memories for event & facts
preserved STM memory
preserved LTM memory
recap - Lesion to parietal/occipital cortex (1969)
reduced digit span (STM approx 2 items)
preserved LTM
double dissociation between LTM & STM
key distinction between STM
Fuster (1974)
studied the neural mechanisms of working memory by showing that neurons in the PFC exhibit sustained activity during the delay period of a task where monkeys had to remember the location of food
Neurons in PFC show sustained, elevated response during the delay period of a working memory task
found storage memory happens in dorsolateral pfc
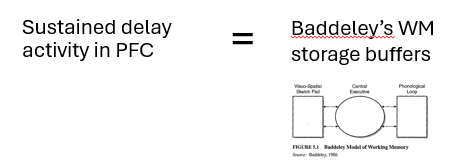
Goldman-Rakic (1987) - standard model of working memory
suggests that this PFC reflects the neuronal instantiation of Baddeley’s working memory storage buffers
sustained activation in PFC during delay period of a WM task reflects a neuronal WM template
a temporary representation of to-be-remembered information
became dominate view of the PFC in working memory for some time
Evidence from monkey neurophysiology for standard model
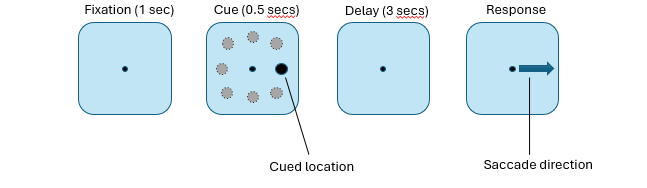
monkey single neuron recordings → oculomotor delayed response task
monkeys saw cue on left/right of fixation & had to maintain their eye gaze at centre for 3 seconds & then make an eye movement in direction of the cue
had to hold direction of cue in their memory for 3 seconds
found that single neurons in PFC showed direction-specific firing during delay period of this task (Between cue & response)
this particular neuron fired strongly when cue pointed to the upper left location
interpret this as showing a direct neurophysiological correlate of WM template, a temporary representation of spatial location indicated by the cue
Supporting human neuropsychological evidence for a role of PFC in working memory (Petrides & Milner 1982)
administered a task called self-ordered task to P w/frontal lobe lesions
P instructed to touch 1 picture per sheet of paper & not to touch the same picture twice until they had through 12 sheets
task requires them to remember from 1 sheet of paper to the next which images they have touched previously
found P w/frontal lobe lesions were disproportionately impaired on this task indicating that P had a deficit w/WM
interpreted this as evidence that the PFC hold representation of to-be-remembered info over short period of a WM
What vs Where in working memory
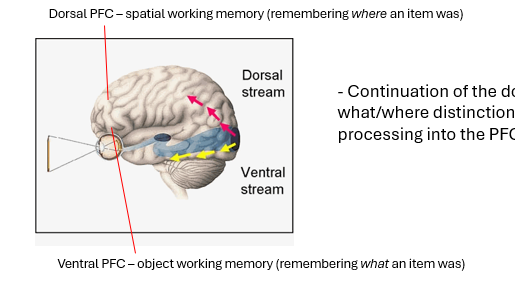
one of key tenets of the standard model is that there are 2 types of visuospatial WM → one for objects & spatial locations
extended what we know about visual object recognition into PFC
the idea that the 2 visual streams extend into the PFC and enable remembering this information over short periods.
locating objects in space & for identifying objects
The evidence for this idea came predominantly from monkey neurophysiology studies
Evidence from monkey neurophysiology for what/where dissociation in working memory
Majority of evidence for ventral/dorsal dissociation in PFC for objects vs spatial working memory came form monkey
same paradigm of spatial delayed response task but with pattern version → had to remember the identity of the pattern (pockadot = down eye movement)
monkeys were trained to perform an oculomotor delayed response task → saw a cue that instructed them to make an eye movement in a particular direction then had to remember that info before making the response
key change to the task was that in addition to standard spatial cues (a cue appearing in actual location to which monkeys had to make an eye movement) & pattern cues (pattern appeared in centre of screen & instructed monkeys to make an eye movement in a particular direction)
response in both trial types was same but type of information to be remembered on each trial differed.
In spatial trials monkey had to remember spatial location of the cue and in pattern trials monkey had to maintain the identity of the pattern
responses of 2 neurons during delay period during each part of the trial
B = shows response of a neuron in inferior convexity – lower, ventral part of PFC
C = response of a neuron in dorsolateral (upper, dorsal) part of PFC
neuron in inferior ventral PFC shows higher activation to patterns & lower activation to spatial cues
neuron in upper dorsal PFC shows higher activation to spatial cues & lower activation to patterns.
what vs where in working memory → support from PET (courtney et al)
evidence from fMRI for ventral/dorsal what/where distinction in WM
P were required to remember either identity or location of 3 faces & say whether test stimulus matched any of identities or locations
Activation for object WM task was seen in ventral PFC & activation for the spatial WM task was seen in dorsal PFC
Object WM task = remember identities of 3 faces → Activation in ventral PFC
Spatial WM task = remember locations of 3 faces → Activation in dorsal PFC
neural basis of WM → testing the claim that PFC stores temporary representations of remembered info
evidence against the idea PFC stores temporary representations of remembered info
Multivoxel pattern analysis (MVPA)
takes advantage of fine-grained pattern of activation in brain
Uses machine learning techniques to teach an algorithm about pattern of neural activation associated w/particular stimulus
Algorithm is then able to ‘decode’ what the subject is looking at simply by viewing their pattern of brain activity
if PFC is a storage then we should be able to predict
Linden et al (2011) - mapping brain activation & information during category specific visual working memory
P performed task requiring them to memorise/hold several objects in WM
On each trial, required to decide whether a single object was part of the memory set
4 categories of objects – faces, bodies, flowers, scenes
Trained pattern classifier to learn patterns of activation for each category
Tested ability of pattern classifier to predict which category subject was holding in WM on each trial
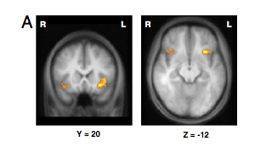
regions holding category-related information are exclusively in posterior part of brain → visual processing regions e.g fusiform face area & other regions of the ventral visual stream
same regions that are activated when these categories of object are presented to the subjects.
implication of this is that the same regions that enable us to process an object when we see it w/our eyes are also involved in storing temporary representations of those objects in WM
looked at whole brain
can predict what P are holding in the memory
but only in posteier brain regions
PFC does not seem to hold temporary representations of information in WM
These temporary representations appear to be stored in brain regions involved in sensory processing – in this example in the visual cortex
shows us what P are memorising
The implication is that the same regions that enable us to process objects when we see them are also involved in storing temporary representations of those objects in WM
PFC is organised according to a ventral/dorsal distinction for remembering what vs where an object is → evidence for what/where dissociation in working memory (Wilson et al, 1955-8)
neuron in the upper dorsal PFC shows higher activation to spatial cues and lower activation to patterns
Evidence against ventral/dorsal (what/where) distinction in PFC (Rao et al 1997)
There is now a considerable body of evidence supporting this idea – that the PFC does not store representations of stored stimuli.
lots of neurons share activation → flexible response properties which can change depending on task
Some of the most convincing evidence against the standard model originally came from monkey neurophysiology → found that single neurons in PFC can encode both the location of an object & its identity
Monkeys had to remember first identity of an object and make an eye movement to the same object in a different location, and then remember its location. They made an eye movement to the location lacking identity information.
Found that neurons could encode both object identity and location – suggests flexible adaptation of responses in PFC – neurons can adapt to represent whatever information is task relevant.
explaining PFC activation during WM - adaptive coding model of PFC (duncan)
PFC neurons show flexibility in their response tuning
whatever they are doing
unlike neurons in other brain regiosn response propteries of PFC neurons aernot fixed buy adapt to encode currenlty task relevant informaiton
why do we find acrivation in doslateral PFC during spacial → role of chunking (Bor & DUncan, 2003)
structured vs unstructured sequence spatial task
whether sequence forms a shape or not
P try to make it easier for themselves by storing information into a manageable chunk
higher activation in lateral for structured
the role of other processes
just becuase P w/frontal lesions can have issues with task
storage of previously touched item with supression of previously youcheditem
selective attention to a novel item
may enage in planning/stragtergy
Human and monkey studies converge to suggest the PFC is not organised according to the type of stimulus held in WM (what vs where)
In fact PFC may not store information in WM at all. What then is the role of the PFC in WM?
We have seen that the ability to follow complex task rules is a key component of fluid intelligence
can we decode information about items held in WM from patterns of activation in prefrontal/parietal cortex (Riggall & Postle, 2012)
looked into the question of where in the brain different types of information are represented during working memory
They scanned people performing a WM task where they had to memorise a moving dot array, and during the delay period were cued to either remember the direction or the speed of the dots.
Then they were shown a probe that was either a match or mismatch.
The key thing here is that when the cue appeared subjects had to follow a rule (attend to the speed or the direction of the dots in WM) and also hold in WM a specific stimulus (the particular speed at which the dots were moving, or the direction in which they were moving)
They found that they could decode which direction and how fast the dots were moving but only from visual cortex and temporal cortex. PFC provided no information about this.
In contrast, task instructions could be decoded from PFC and parietal regions.
This is consistent with the Linden study – stimulus specific information was stored in posterior, sensory specific cortex. However, information about task rules was stored in frontal and parietal cortex.
can decode from patterns from posterior PFC about what info is being held in wm
alternative view to standard model → WM as emergent property (Postle, 2006)
“If the brain can represent it, the brain can also demonstrate working memory for it.…working memory functions are produced when attention is directed to systems that have evolved to accomplish sensory-, representation-, or action-related functions.
From this perspective, working memory may simply be a property that emerges from a nervous system that is capable of representing many different kinds of information, and that is endowed with flexibly deployable attention”.
WM is a process of internal attention
working memory as attention to internal stimuli
fMRI evidence for internal attention hypothesis of WM (Higo et al. 2011) - consistent with idea of WM being attention for stimuli
provided direct evidence for the idea that the PFC performs an attentional role in WM
P held two objects in WM and were subsequently cued either to maintain both objects (non-selective attention condition) or a single object (selective attention condition) in WM
Finally asked to decide whether any of the objects in an array matched the object(s) they were holding in WM
Activation in PFC was greater for the selective condition than the non-selective condition
Connectivity analysis demonstrated that activation in PFC modulated activation in different occipitotemporal regions depending on which stimulus the subject maintained in WM
Combined TMS/fMRI showed that disruption of PFC activity caused distal effects on occipitotemporal activation, thus establishing direction of causation
PFC seems to send an attentional bias signal to sensory-specific regions to enhance processing of the task-relevant object during WM
distributed neuronal architecture of WM
lower level visual regions maintain the temporary representations of items held in WM (the templates) and PFC/parietal regions hold a representation of the task rules for manipulating this information (the central executive!)
in posterieror storage of specific items in WM
frontal attentional processing/maniputaltion of items in WM
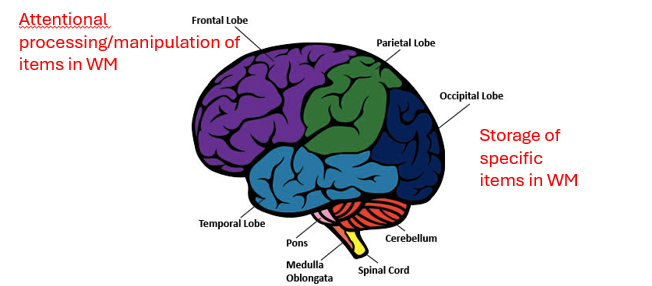
key points for neuronal WM
PFC neurons show sustained activity during the delay period of a WM task
However, this activity does not reflect storage of items in WM
Representations of items stored in WM are maintained in sensory-specific cortex
PFC seems to play a role in the processing of stimuli held in WM
This role may involve enhancing attention to internal representations of task relevant stimuli in working memory and manipulating such information according to a set of stored task rules
This ability plays a key role in fluid intelligence