Controlling Gene Expression
1/24
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
25 Terms
Mutation
Changes to exons in the DNA code result in changes to the protein if they change the amino acid that is coded for. The DNA code is degenerate, as more than one codon can code for the same amino acid, which means that if you change one base it may result in the SAME amino acid being coded for.
Mutations happen randomly but the likelihood of one occurring can be increased by physical or chemical factors that affect DNA (e.g exposure to some type of ionising radiation or carcinogenic chemical e.g asbestos, alcohol, tobacco smoke) we call these mutagens or mutagenic agents.
The consequence of a mutation is that there may be a change to the codon, which may change the amino acid. This could result in a faulty protein e.g a change in the base sequence may change the primary structure of the protein which could have knock on effects to the secondary and tertiary structure of the protein. If the protein is an enzyme the active site could change shape which would mean it could no longer form E-S complexes.
Not all mutations are negative, positive mutations which give organisms advantages that help them to increase their survival and reproduction rate compared to others in their species can lead to evolution.
Types of mutation:
Mutation Type | Image/Diagram |
Substitution – where one or more bases are swapped for another (can result in no change as amino acid sequence could stay the same – a silent mutation) | |
Inversion – a sequence of bases is reversed (can result in no change as amino acid sequence could stay the same – a silent mutation) |
|
Insertion (addition) – one or more bases are added (can cause a frame shift) | |
Deletion – one or more bases are removed (can cause a frame shift) | |
Duplication – one or more bases are repeated (can cause a frame shift but can also result in increasing repeating subunits and cause genetic diseases e.g Huntington’s which is caused by a duplication mutation that increases the number of repeats of the base sequence CAG) | |
Translocation – a sequence of bases is moved from one location in the genome to another, this could be movement within the same chromosome or to a different chromosome. This happens when something similar to crossing over occurs but between non-homologous chromosomes |
Frame Shift Mutations (deletion, insertion, duplication)
These mutations can have a huge effect on the base sequence of a gene because they change the number of bases in the DNA code causing a shift to all the base triplets that follow (downstream) so that all codons after the mutation are changed and the DNA code will be transcribed differently.
Stem Cells
Stem cells retain the ability to divide repeatedly and differentiate (become specialized) into a range of cell types. Stem cells become specialized because they only transcribe and translate parts of their DNA. Which genes are expressed is determined by the conditions the cell is in, this causes proteins to be transcribed which modify the cell causing it to become specialized for a particular function. There are two broad categories of stems cell: adult and embryonic. Stem cells can be described as:
TOTIPOTENT STEM CELLS (OMNIPOTENT) | PLURIPOTENT STEM CELLS | MULTIPOTENT STEM CELLS | UNIPOTENT STEM CELLS |
Cells that can differentiate into embryonic and extra-embryonic tissues. | Cells that can differentiate into any of the three germ layers (ectoderm, endoderm, mesoderm), so any tissue in the body. | These can divide to form different cell types but not all. | Cells that can only form one other type of cell. |
They form the embryo and the placenta. | Occur in early embryos of mammals, cambium of plants. | Example is stem cells in bone marrow that form most cells of the immune system, red blood cells and platelets. | Example: Cardiomyocytes can divide to form other heart muscle cells. |
All genes in the nucleus are able to be activated. | The vast majority of genes are still able to be activated. | Some genes have been switched off but they have the genes for some different cells still available. | Lots of genes have been switched off via regulation of transcription factors |
Embryonic Stem Cells
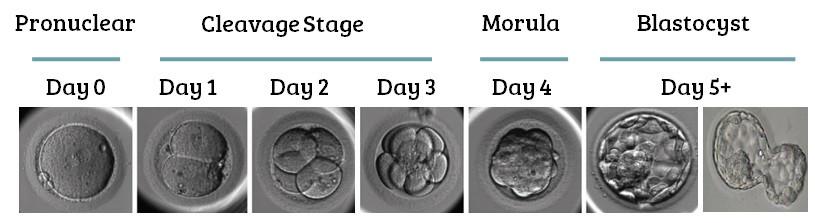
They are taken from the embryo- this is the only stage in human development where cells need to be totipotent as the embryo must divide to produce new cells which will eventually form all the specialized tissues in the body. In the very early embryo, cells are totipotent. Cells collected from the inner mass of a blastocyst (4-7 days) are pluripotent as they have already begun to differentiate.
Due to their potency/plasticity embryonic stem cells can be used to grow new organs and tissues for treatment of a variety of conditions. This could save or improve the quality of the lives of many people and reduce the need for organ donation in the future. However, their use is controversial, and research is tightly regulated in the UK to decide on the maximum age of embryos allowed for research and to prevent human cloning.
Objections to use of embryonic stem cells in medicine:
Unused IVF embryos can be donated to research instead of being destroyed. Some people object to this as it is the destruction of a potential foetus, some people believe the embryo has the right to life.
Egg cells frozen for IVF which have not been fertilised can be stimulated to divide – this causes less objection as they would not survive after a few days if implanted into a womb and would not create a foetus.
It is possible to use adult stem cells for some treatments which people feel is more ethical, however they cannot develop into all the specialised cell types that embryonic stem cells can.
Adult Stem Cells
These are obtained from the body tissues of an adult. They are found in bone marrow and can be removed with a simple operation that carries little risk. Adult stem cells are multipotent so they cannot differentiate into as many cell types as embryonic stem cells can. Bone marrow transplants have already been successful to replace faulty bone marrow that produces abnormal blood cells e.g lukaemia. Stem cells have also been used to treat paralysis caused by spinal cord injuries by replacing nerve tissue.
Induced Pluripotent Stem Cells (iPS Cells)
These stem cells are created in a lab from specialised adult cells after ‘reprogramming’ them to become pluripotent. The cells are made to express transcription factors (proteins which control the expression of genes) which are normally expressed by pluripotent stem cells. They “switch on” genes that are normally expressed in cells that have pluripotency. This could allow adult cells to become as plastic as embryonic stem cells and could be obtained without the same ethical issues. It is also possible that they could be made from a patients own cells which would be genetically identical to the patient – this would allow tissues/organs to be grown that wouldn’t be rejected and reduce the wait time for transplants.
Future Stem Cell Therapies
Clinical trials are already undergoing to look into the use of stem cells to replace diseased or damaged organs such as the bladder or to replace damaged tissue e.g in the trachea (a donor windpipe can be stripped of tissue and used as a cartilage base for stem cells to grow new tissue on) or damaged heart tissue after a heart attack or heart disease.
Recap of Protein Synthesis:
Transcription of the gene in the nucleus using RNA polymerase to convert DNA to mRNA
Pre-mRNA strand is spliced introns removed
mRNA leaves nucleus via nuclear pore
mRNA binds to ribosome on the RER
Translation of the mRNA into a polypeptide
Modification, folding and packaging of the protein in the golgi.
A gene is expressed or ‘switched on’ when it is transcribed into mRNA and then translated into a protein. Only a few genes in a cell will be expressed at any one time – this is how cells differentiate and become specialised.
Transcription Factors:
Before transcription can begin a gene needs to be stimulated by a transcription factor. Transcription factors are proteins which travel into the nucleus and control the rate of transcription by either activating or repressing RNA polymerase. Each transcription factor binds to a specific region of the DNA known as the promoter region. The promoter region is the binding site of RNA polymerase and therefore the starting point for transcription of that gene.
Transcription factors can turn genes on and off as they can either activate or block the functioning of RNA polymerase, repressors will bind to the promoter region and prevent the RNA polymerase from binding therefore stopping transcription of that gene.
Transcription factors can be switched off by an inhibitor molecule. This can bind to the transcription factor, preventing it from attaching to the promoter region. Without the transcription factor the gene may not be transcribed (if the TF is an activator) or the gene may be expressed (if the TF is a repressor).
Oestrogen as a Transcription Factor
A hormone response element (HRE) is a short sequence of DNA within the promoter of a gene, that is able to bind to a specific hormone receptor complex and therefore regulate transcription.
1 – 2 To affect transcription oestrogen has to bind to a transcription factor called an oestrogen receptor forming an oestrogen-oestrogen receptor complex. (not all cells have these receptors so not all cells are affected by oestrogen)
3 The oestrogen-oestrogen receptor complex moves from the cytoplasm into the nucleus
4 The complex binds to the HRE in the promoter of the gene, activating or repressing transcription
Preventing Translation using interfering RNA (RNAi)
RNAi molecules are small lengths of non-coding RNA (they do not code for proteins). They regulate gene expression by affecting translation instead of transcription. They are double stranded (unlike other RNAs!) and stop mRNA that has been transcribed from being translated into a protein. There are two types:
Short interfering RNA (siRNA) – in animals only
Micro RNA (miRNA) – in plants and animals
How they siRNAs and miRNAs (in plants) work:
Double stranded siRNA associates with proteins in the cytoplasm and unwinds
One of the siRNA strands is chosen and the other degrades (breaks down)
Single strand of siRNA binds to the target mRNA (as it is complimentary to the base sequence in a section)
The proteins associated with the siRNA cut the mRNA into small fragments so it can no longer be translated. The mRNA fragments then move into a processing body which degrades them.
This has the potential use for “gene silencing” of inherited genes which cause disorders. Patients could be treated with siRNA molecules that are complimentary to mRNA transcribed from faulty genes to prevent faulty proteins from being expressed.
How do miRNAs work in mammals:
miRNAs are not fully complimentary to the target mRNA so they are often less specific and can target more than one mRNA molecule. miRNA has to go through processing stages in the cytoplasm to be a single strand like siRNAs: from a folded strand to a double strand then two single strands, one of these is degraded as with siRNA.
Once the miRNA is bound to the mRNA it does not cause degradation instead it simply blocks the translation by preventing a ribosome from binding. The mRNA is then moved to a processing body where it can be stored or degraded.
Gene Expression and Cancer
Control of gene expression also plays an important role in cancer. The rate of cell division is controlled by genes, if mutations occur in these genes then uncontrolled cell division can occur. Uncontrollable cell division causes the cells to keep on dividing and they layer on top of each other forming a tumour – a mass of abnormal cells. Tumours that invade and destroy surrounding tissues are called cancers. There are two types of genes that control cell division:
PROTO-ONCOGENES | TUMOUR SUPPRESSOR GENES | |
These genes can cause cancer when they are activated or turned on. | These genes can cause cancer when they are inactivated or turned off. | |
Proto-oncogenes are genes that normally help cells grow. When they mutate they can become permanently activated. When this happens, the cell grows out of control, which can lead to cancer. A mutated proto-oncogene is known as an oncogene. | Tumour suppressor genes are genes that normally help cells slow down cell division, repair DNA mistakes, or tell cells when to die (a process known as apoptosis or programmed cell death). When tumour suppressor genes don't work properly, cells can grow out of control, which can lead to cancer. | |
For example, Ras is one of the on/off switches in a series of steps in a major pathway that eventually leads to cell growth. When Ras is mutated, it encodes for a protein that causes an uncontrolled growth-promoting signal. | For example, abnormalities of the TP53 gene (which codes for the p53 protein) have been found in more than half of human cancers. This protein acts as a tumour suppressor, which means that it regulates cell division by keeping cells from growing and dividing too fast or in an uncontrolled way.. |
Mutations Causing Cancer
Mutations are changes to the base sequence of DNA, these can be caused by mutagens such as ionising radiation from UV rays or carcinogenic chemicals like those found in cigarette smoke. Some people can also inherit mutations in genes that make them more likely to suffer with some cancers.
Example: About 5-10% of breast cancer cases are thought to be hereditary. The most common cause of hereditary breast cancer is an inherited mutation in the BRCA1 or BRCA2 gene. In normal cells, these genes help make proteins that repair damaged DNA. Mutated versions of these genes can lead to abnormal cell growth, which can lead to cancer.
If you have inherited a mutated copy of either gene from a parent, you have a higher risk of breast cancer.
Women with one of these mutations are more likely to be diagnosed with breast cancer at a younger age, as well as to have cancer in both breasts.
Tumours
Cancer cells are different to normal cells in their structure and function, they will either die through apoptosis or they can be destroyed by the immune system which recognises them as ‘foreign’ or different to normal body cells because they:
Have large dark nuclei sometimes more than one
Irregular shape
Do not produce proteins needed to function properly
Different antigens on their surface membrane
Do not respond to the usual growth regulation process
Tumours need their own blood supply but often they suffer from low oxygen as the blood supply cannot satisfy the tumours oxygen demand as the rate of cell division is so high. There are two types of tumour:
Non-cancerous Slow growing Do not metastasize but can put pressure on organs or cause blockages Cells have a normal shape | Cancerous Fast Growing Invade and destroy other tissues and metastasize (spread) around the body in the lymphatic system May have abnormal shape. |
Oestrogen and Breast Cancer
Increased exposure to oestrogen over an extended period of time is thought to increase a woman’s risk of developing breast cancer. There are a couple of reasons that some women may be exposed to more oestrogen than others:
Taking birth control or HRT (drugs that contain oestrogen which help control menstruation or treat symptoms of the menopause)
Starting menstruation early in life
Starting menopause late in life
Risk of developing breast cancer also increases with age. As women age, the fat cells in their breasts tend to produce greater and greater amounts of an enzyme called aromatase. Aromatase promotes the production of oestrogen. Consequently, with age, the levels of oestrogen present in women's breasts increases. The exact cause is still not fully understood but there are some theories about how high oestrogen causes some breast cancers:
Stimulates more breast tissue cells to divide increasing the chance of mutations
Cancer cell replication could be further promoted by oestrogen causing faster tumour growth
Treating Cancer
Cancer treatments can control the rate of cell division in cancer cells by targeting the cell cycle to try and stop it. This will kill the tumour cells and prevent them from dividing further. However, the treatments are not able to distinguish between cancer cells and other normal body cells that divide a lot e.g hair cells, stem cells in bone marrow and cells that line the small intestine (this explains side effects of cancer treatment e.g hair loss!). Cancer cells have a faster rate of division, so these cells are more likely to be affected by the cancer treatment. Examples of cancer treatments include:
Surgery is normally conducted where possible to remove a specific tumour if it hasn’t spread but this can lead to breakage and spreading of cells if not done carefully.
Immunotherapy and targeted therapy can help direct treatments to cancerous cells to reduce side effects and damage to the body.
Chemotherapy which prevents the synthesis of enzymes required for DNA replication. This stops the cell cycle before the S phase as it cannot take place. If DNA cannot be replicated cells will not be able to divide.
Radiotherapy and some chemotherapy drugs can damage DNA, if DNA is damaged the cell will not pass the check points in S phase. This will force the cell to kill itself (apoptosis). Cells will not divide/mitosis is prevented
Drugs which prevent spindle fibres from forming will prevent mitosis stage as one cell will contain double the DNA and one cell will contain no DNA. This prevents the cells from functioning or dividing again.
Regulation of Gene Expression II- Recap: DNA Structure
In eukaryotes DNA is held in the nucleus surrounding proteins called histones. They package and order the DNA into structural units called nucleosomes. The histones associate (bind to) the DNA and help the DNA to condense into chromatin. Condensed DNA (chromatin) that is tightly bound to histones cannot be transcribed, the unwinding allows access for transcription.
Genome vs Epigenome
Histones are covered in chemicals: “tags”. These tags are known as the epigenome – ‘epi’ comes from the Greek and means “on”. These tags determine the shape of the histone-DNA complex and they help to control the transcription of genes. Epigenetics is the study of inheritable changes in gene function that have not been caused by changes to the base sequence of DNA.
Genome | Epigenome |
The sequence of bases in the entire DNA molecule of an organism – this sequence is fixed | The tags on the histone proteins which are associated with DNA – these tags can be changed easily |
Sequence can only be changed by mutation | Tags can respond to external factors and change gene expression without affecting DNA sequence |
Mutations in DNA sequence can be inherited | Epigenetic tags are often removed in gametes to allow embryos to be totipotent. But sometimes they remain allowing changes to gene expression caused by environmental influences can be inherited |
Environmental factors can influence the genome by causing mutations, but these are specific | Environmental factors can influence the epigenome, but they can be wide ranging |
Epigenetic Control of Gene Expression:
There are two ways that epigenetics can control gene expression using the epigenetic tags of methyl and acetyl groups:
Increasing the methylation of DNA (adding a methyl group -CH3 to DNA). This alters the DNA structure preventing transcription factors and enzymes from binding to it (i.e switching it off). DNA methyltransferase enzymes attach the methyl group to cytosine bases on the DNA molecule.
Acetylation of histones (adding or removing -COCH3 group). When histones are acetylated the DNA loosens from the histones making it less condensed. This means the transcription factors and enzymes can bind to the DNA and transcribe it. When acetyl groups are removed from the histones the chromatin condenses and transcription factors and enzymes cannot access the DNA so it is not transcribed. Histone deacetylase (HDAC) enzymes are responsible for removing acetyl groups, while acetyltransferase enzymes add acetyl groups. There are several enzymes that control methylation and acetylation. They work together to increase or decrease expression of different genes. These have major roles in development of cancer, inheritance of certain environmental factors and development of other diseases.
Epigenetics and Disease
Abnormal methylation (too much or too little) of proto-oncogenes and tumour suppressor genes can cause abnormal cell growth and cancers to develop.
Hypermethylation of tumour suppressor genes prevents them from being transcribed. The proteins they normally produce which slow down cell division will not be made. This can cause cells to divide uncontrollably by mitosis and form tumours.
Hypomethylation of proto-oncogenes causes them to act as oncogenes and produce proteins that encourage cell division. This can stimulate cells to divide uncontrollably by mitosis and form tumours.
Epigenetics can be used to treat diseases as these changes are reversible unlike DNA sequence mutations. Any drugs which counteract epigenetic changes must be very specific to their target genes as some of these changes are necessary and important to normal function and happen in all cells in the body. Some examples:
Drugs that stop DNA methylation can prevent genes from being switched off so they can be transcribed and produce proteins that when missing cause symptoms of the disease e.g Prader-Willi Syndrome, Fragile -X Syndrome, and Angelman Syndrome.
HDAC inhibitor drugs such as romidepsin can be used to prevent decreased acetylation of histones which can stop some genes from being turned off. They work by inhibiting the activity of decacetylase enzymes so that the gene remain uncondensed and can be transcribed.
How environmental factors can affect epigenetics
Epigenetic changes in gene expression caused by a response to environmental changes e.g pollution, availability of food or drought (in plants) experienced by an organism have been shown to have been inherited and be present in the children and sometimes grandchildren of that organism.
There are many environmental influences that can affect the epigenome, see the diagram on the right.
A good example has been demonstrated in rats. Some mother rats spend a lot of time licking, grooming, and nursing their pups. Others seem to ignore their pups. Highly nurtured rat pups tend to grow up to be calm adults, while rat pups who receive little nurturing tend to grow up to be anxious. It turns out that the difference between a calm and an anxious rat is not genetic, it's epigenetic. The nurturing behavior of a mother rat during the first week of life shapes her pups' epigenomes and this behaviour stays even after the rats become adults and affects whether they are high nurturing mothers or not when they have their own pups. This effect on the DNA was determined to be epigenetic not genetic as it was still seen in rats who were raised by “foster mums” who they shared no DNA with.
This was further evidence by the ability to reverse these traits in rats using drugs. You can take a low-nurtured rat, inject its brain with a drug (TSA) that removes methyl groups, and make it act just like a high-nurtured rat. The GR gene gets turned on, cells make more GR protein, and the rat takes on a more relaxed personality. You can also take a relaxed, high-nurtured rat, inject its brain with methionine (a source of methyl) and make it more anxious.
Inheriting epigenetic changes are an advantage as it allows the inheritance of behaviours which will make offspring automatically adapted/prepared to deal with the environmental changes when they are born.