SIGNAL TRANSDUCTION FULL DECK
1/69
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
70 Terms
Cell signaling (multicellular)
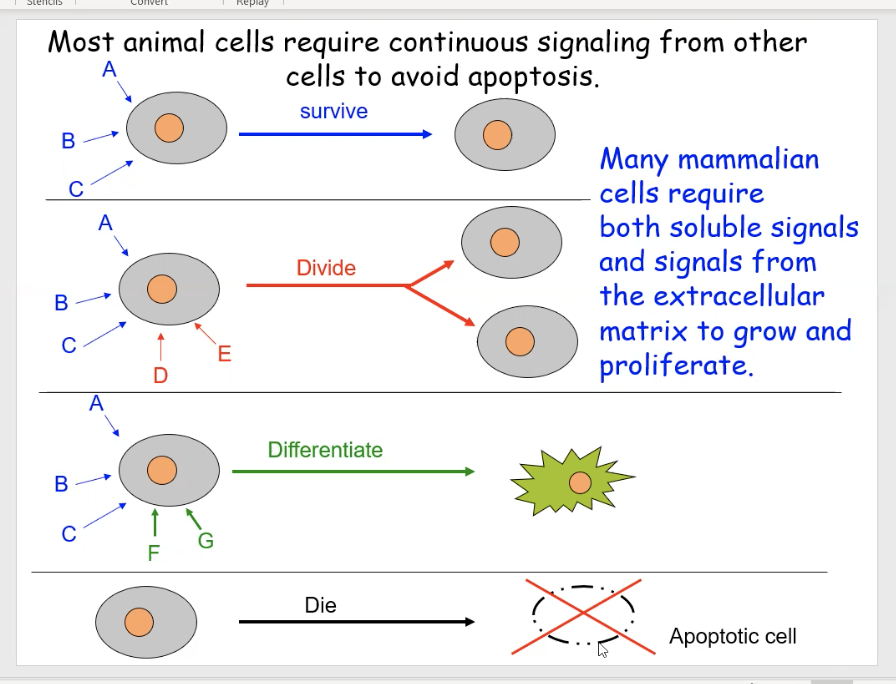
Cells require constant signaling from their environment, other cells, and their own interior
The diagram shows different examples of cells getting various combinations of signals, which enable them to perform specific functions.
Cell 1 is getting signals from signals A,B,C to tell it to survive
Cell 2 is getting signals from A, B, C AND D, E, which tells it to divide
Cell 3 is getting signals from A, B, C AND F, G telling it to mature
Cell 4 has no signals which means it should undergo apoptosis
unicellular cell communication
Single-celled organisms can communicate with each other
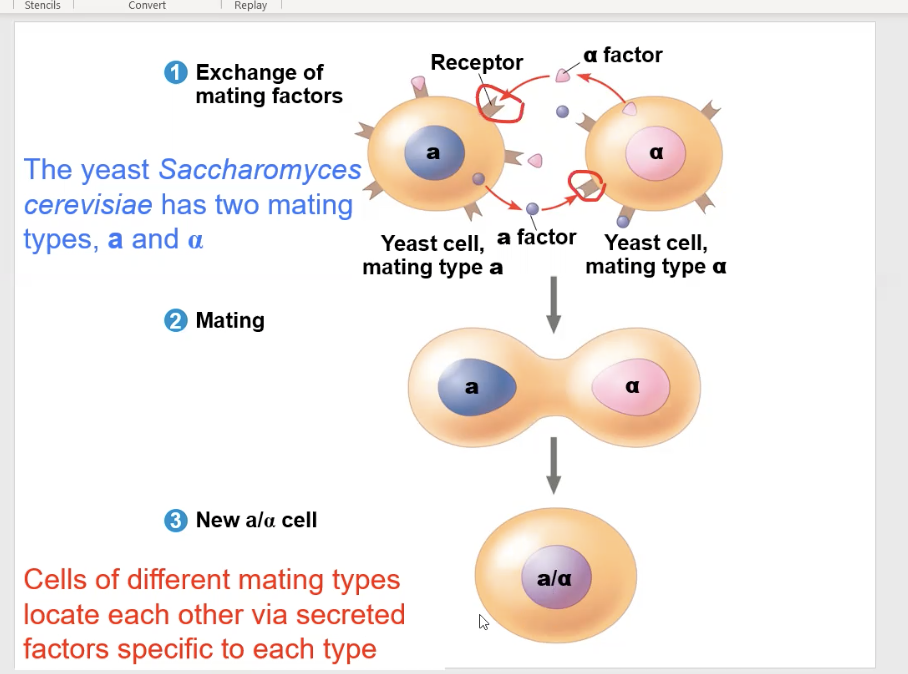
yeast cell communication
They communicate by secreting soluble factors on each other. also known as mating factors
The Saccharomyces cerevisiae yeast has two different factor types, the “a” type and the “α” type
The “a” type and “α” type yeast can only mate with each other, so they send each other the mating factors, and can only bind to each other’s receptors
Quorum sensing
Prokaryotes, single-celled organisms, can use this to cell signal
a form of bacterial communication where cells release signal molecules to sense their population density
local signaling
whens cells communicate with nearby cells, or with cells in direct contact with them.
Direct contact local signaling
Gap junctions in animal cells, plasmodesmata in plant cells, and Transmembrane proteins for cell-to-cell recognition all allow for direct local signaling
Plasmodesmata
channels in land plant and algal cell walls that connect the cytoplasm of neighboring cells, allowing for direct transport and communication of molecules like sugars, ions, proteins, and RNA
gap junctions
Protein channels in animal cells that connect the cytoplasm of adjacent cells, allowing direct passage of ions, small metabolites, and signaling molecules without entering the extracellular space
cell-to-cell interactions: Transmembrane proteins
Many transmembrane proteins function as gateways to permit the transport of specific substances across the membrane
Non direct contact local signaling
paracrine signaling and Autocrine signaling
paracrine signaling
Animal cells may use secreted messenger molecules (molecules travel short distances) to communicate with the target cell
The target cell must have a specific receptor for that messenger molecule
synaptic signaling
a specific type of paracrine signaling. An axon will extend to target cell, and release neurotransmitters.
Autocrine signaling
the secreting cell will send a signal, and it binds back on itself on its own receptor
long distance signaling
endocrine signaling
endocrine signaling
the secreting cell will send signals into the blood stream and travel down the blood vessel to the target cell
signaling molecules
can be either polar or nonpolar.
Nonpolar molecules easily travel through the plasma membrane. The signal molecule will enter the target cell and bind to a receptor inside the target cell. when the receptor binds to the signal, the receptor will go through a. conformational change (steroids are generally non-polar, e.g., cortisol)
polar signal molecules will find their receptor embedded in the plasma membrane (polypeptides and amino acids generally polar, e.g., secretin and epinephrine, respectively)
ligand
signal molecule
4 main types of receptors
Intracellular receptors
ion channel receptors
G-protein-linked receptor
Protein kinase (enzyme-linked) receptors
Intracellular receptors
located in the cytoplasm and nucleus
binds to nonpolar hydrophobic signal molecules
is often a transcription factor (After the ligand binds to the receptor, the receptor will go through a conformational change, which allows the ligand+receptor unit to change location, into the nucleus to aid in transcription)
Ion channel receptors
transmembrane receptor
When a ligand binds to the receptor, a conformational change causes the channel's "gate" to open, allowing specific ions (like Na⁺, K⁺, Ca²⁺, or Cl⁻) to flow across the cell membrane
“ligand-gated ion channel” (the presence of a ligand determines if the channel is open or closed)
Neuron signaling is an example
G- protein linked receptor
a receptor bound to a G protein
Protein kinase receptors
kinase is an an enzyme that puts a phosphate group on something
Upon binding to a ligand, the kinase receptor will transport phosphate groups to proteins
signal transduction pathway
an external signal, such as a hormone or neurotransmitter, triggering a series of molecular events, that forms the cells response to the signal.
basically a ligand binds to surface receptor→intracellular events occurs→these events trigger the the cellular response
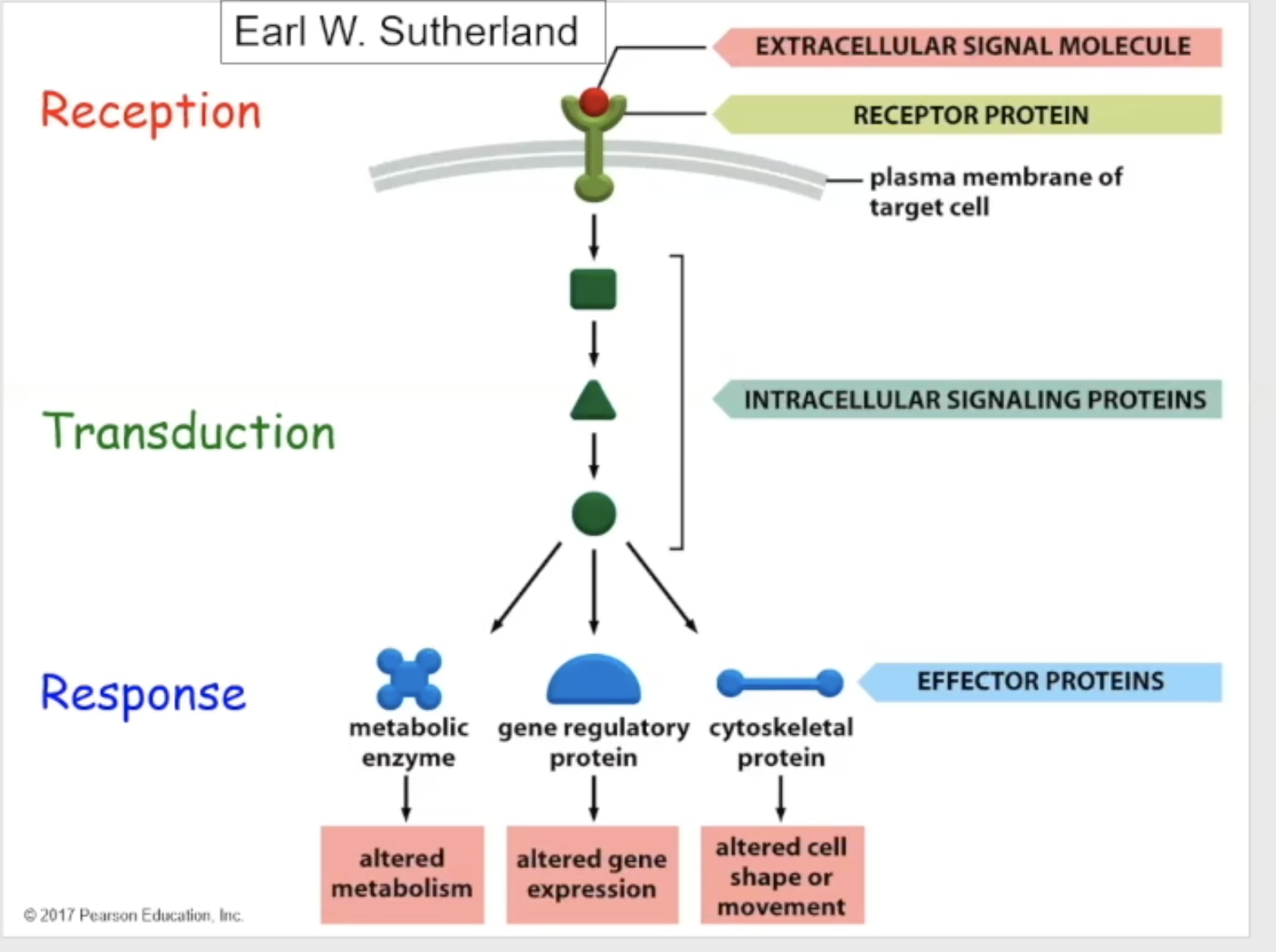
Signal transduction parts
Reception- the target cell detects that the signaling molecule has bound to the receptor
transduction- signaling/relay proteins work as intermediates, activating and changing, and communicating with one another, to tell the cell that the binding has occurred
response- the final response (such as a change of enzyme, alteration of gene expression, or change in cell shape to move somewhere else)
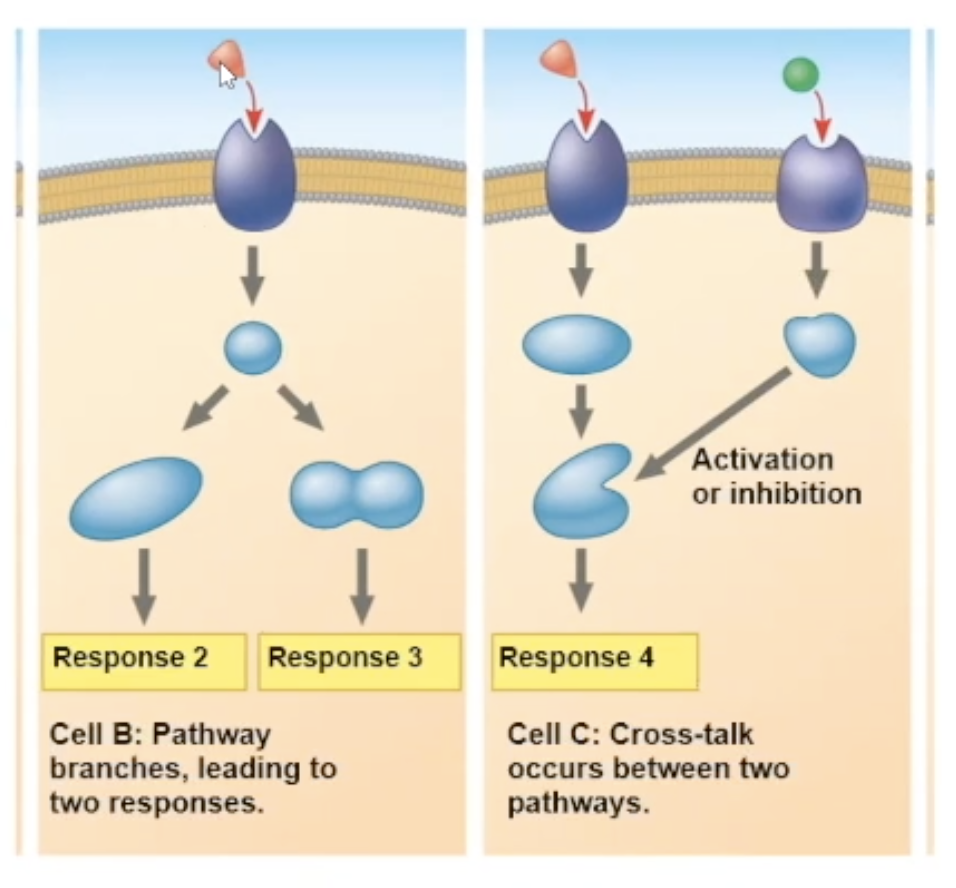
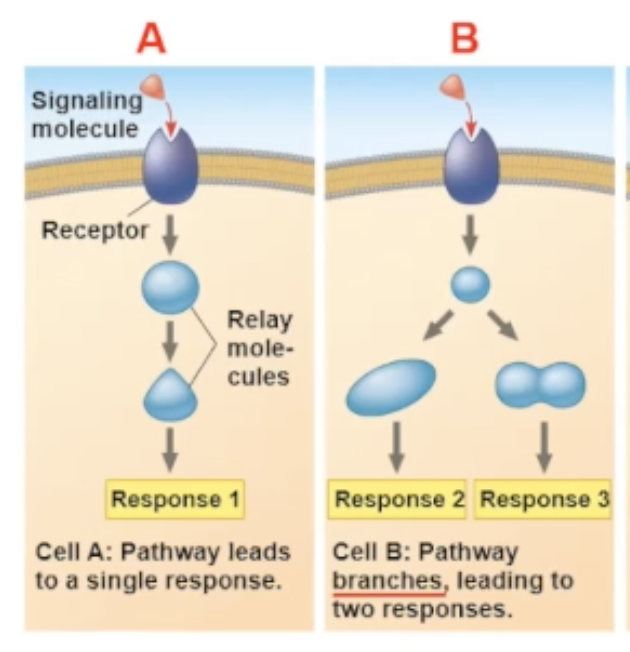
multiple responses
there are many different response paths that a binding can cause.
One pathway can fork off into two different responses
2 different signaling molecules pathways can communicate, and create a unique response
Reception
After the ligand binds to the surface receptor, the target cell detects that this binding has occurred
The receptor molecule (ligand + receptor) will go through a conformational change
transduction
the relay molecules will interact with each other one by one
G protein coupled receptor
transmembrane receptors that get coupled with G proteins
Usually, the polypeptide chain of this receptor folds on itself 7 times through the membrane
largest family of cell surface receptors
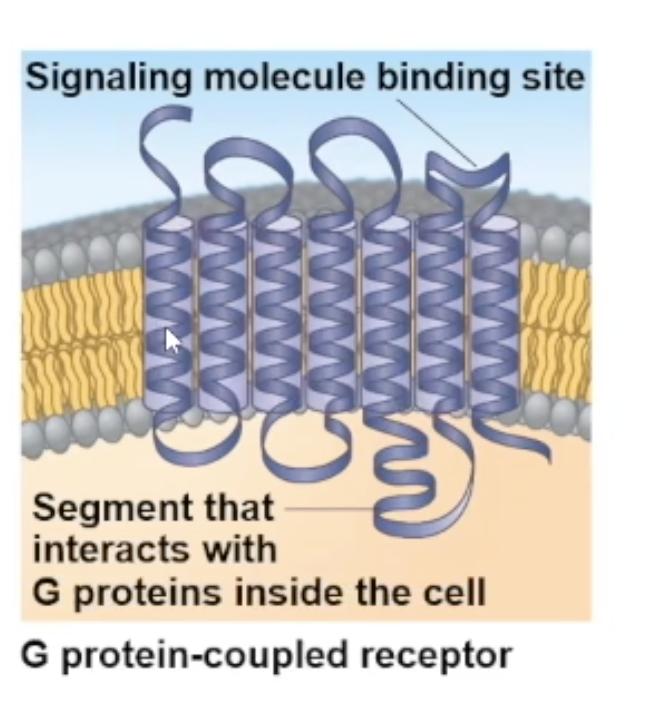
G proteins
G protein bound to GDP is off (inactive)
G protein bound to GTP is on (active)
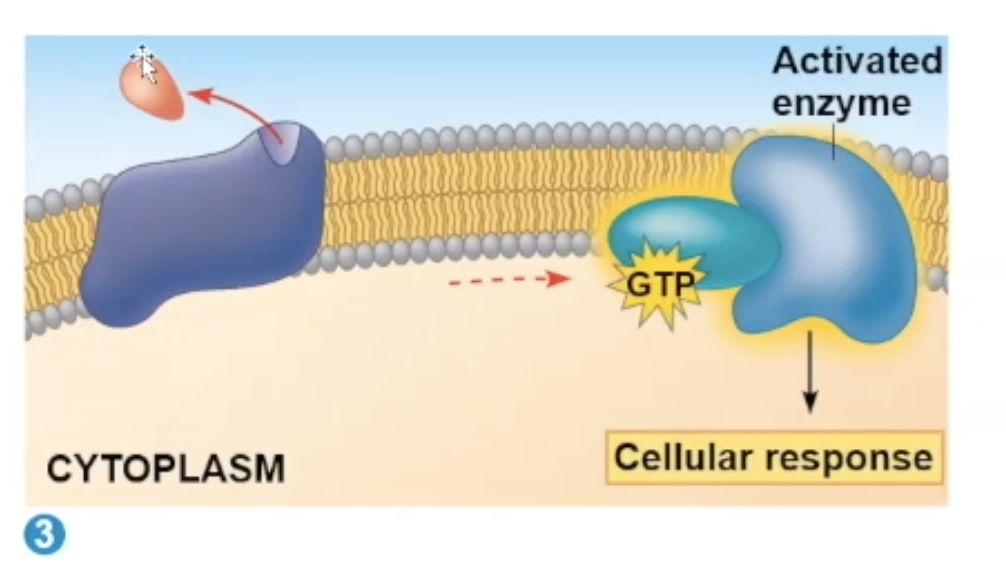
G protein receptor process
When a ligand (epenephrin is a common ligand for this process) binds to the receptor, it goes through a conformational change, which allows the receptor to have contact with a G protein.
The contact will allow the G protein to drop its GDP and gain GTP.
The G protein will move to an enzyme, activating the enzyme.
Adenylyl cyclase
one enzyme that the G protein can bind to
this enzyme can generate many '“second messengers'“ after being activated
second messengers
small non-protein molecules or ions that are water-soluble. They spread throughout the cell very quickly by diffusion. It can bring its information to a protein inside the cell.
cAMP molecules and calcium ions are common secondary messengers
The first ligand that binds to the G protein receptor (e.g., epinephrine) is a first messenger
Cyclic AMP (cAMP)
one of the most commonly used second messengers
The Adenylyl cyclase enzyme will chop 2 phosphates from an ATP (the 2 phosphates will just leave)
The remaining phosphate connects to the 5’ and 3’ carbons on the ribose sugar of the ATP, creating cAMP, aka the second messenger
Another enzyme called phosphodiesterase will further break down cAMP, creating AMP
Protein kinase A
One of the main targets of the cAMP secondary messenger
cAMP binding will allow protein kinase A to activate
When protein kinase A gets activated, it phosphorylates other proteins/molecules
phosphorylation cascade
A kinase will get activated and phosphorylate other kinases, which will trigger that kinase to get activated and phosphorylate another kinase. This cycle continues, however many times, until the final kinase phosphorates an inactive protein, creating an active protein that can create a cellular response
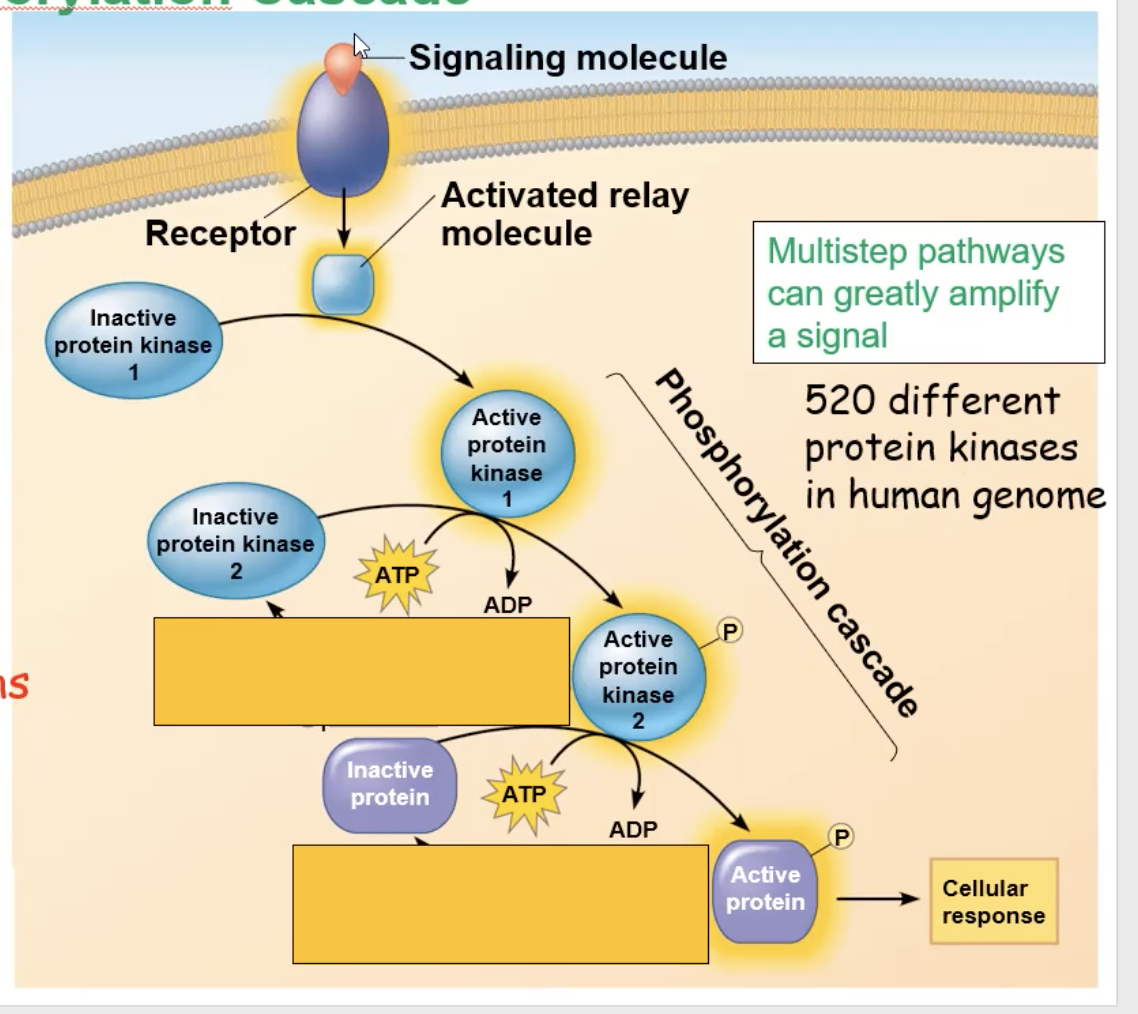
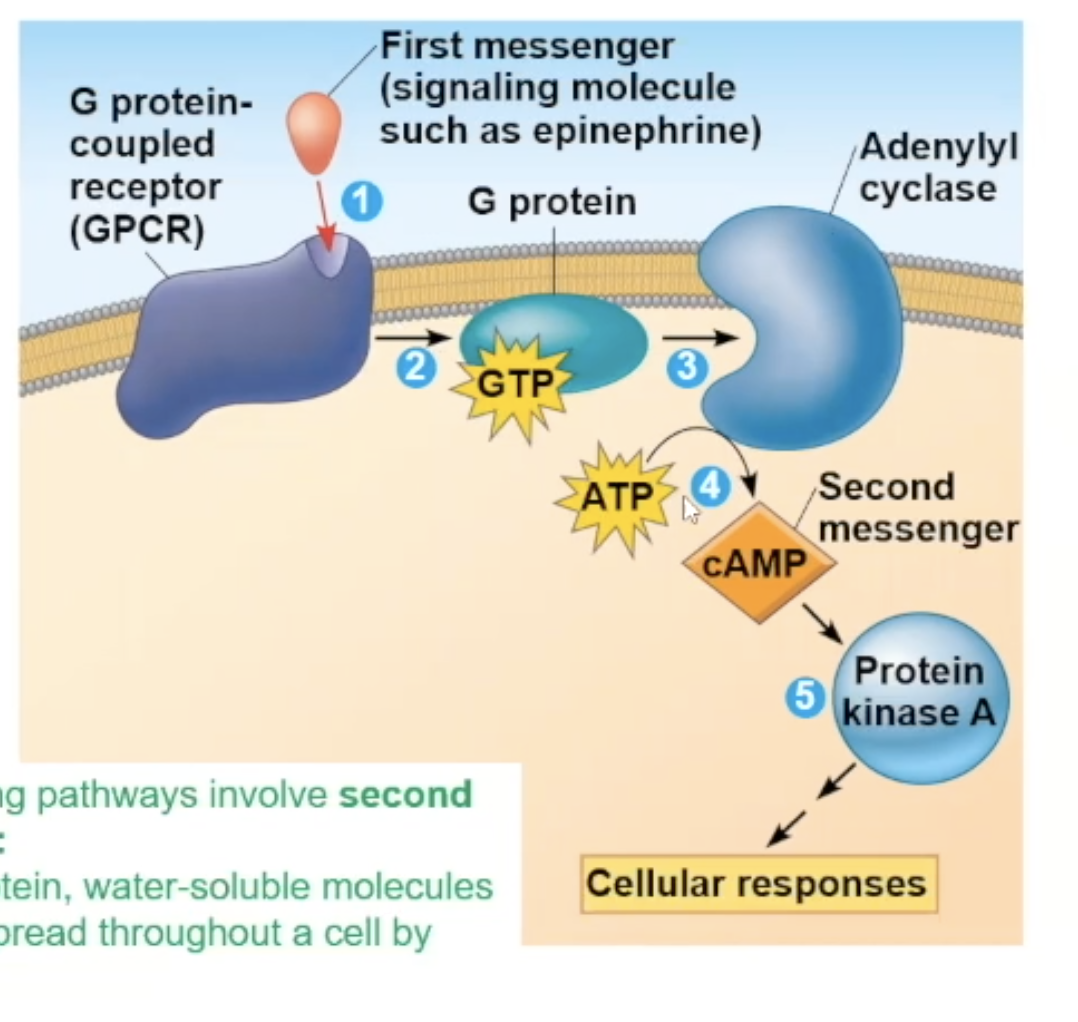
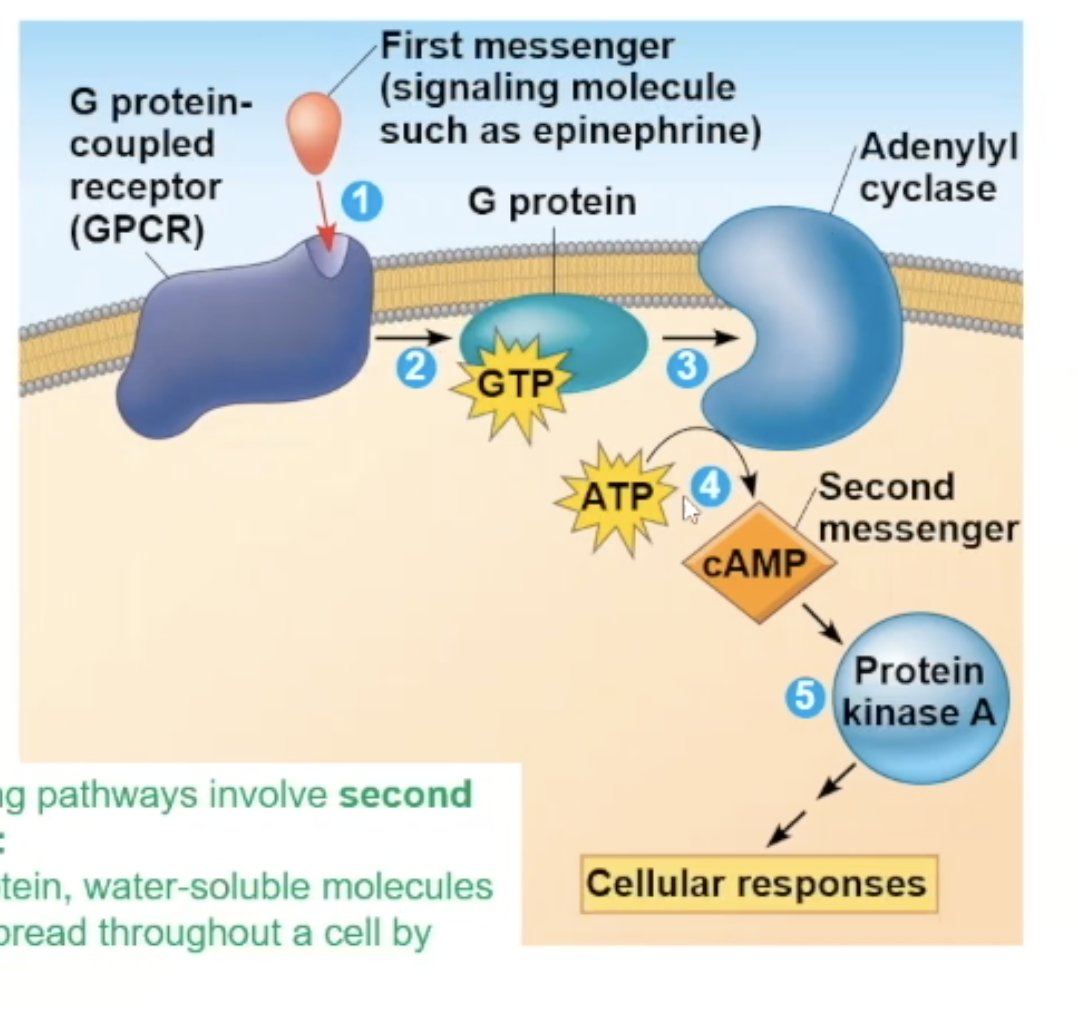
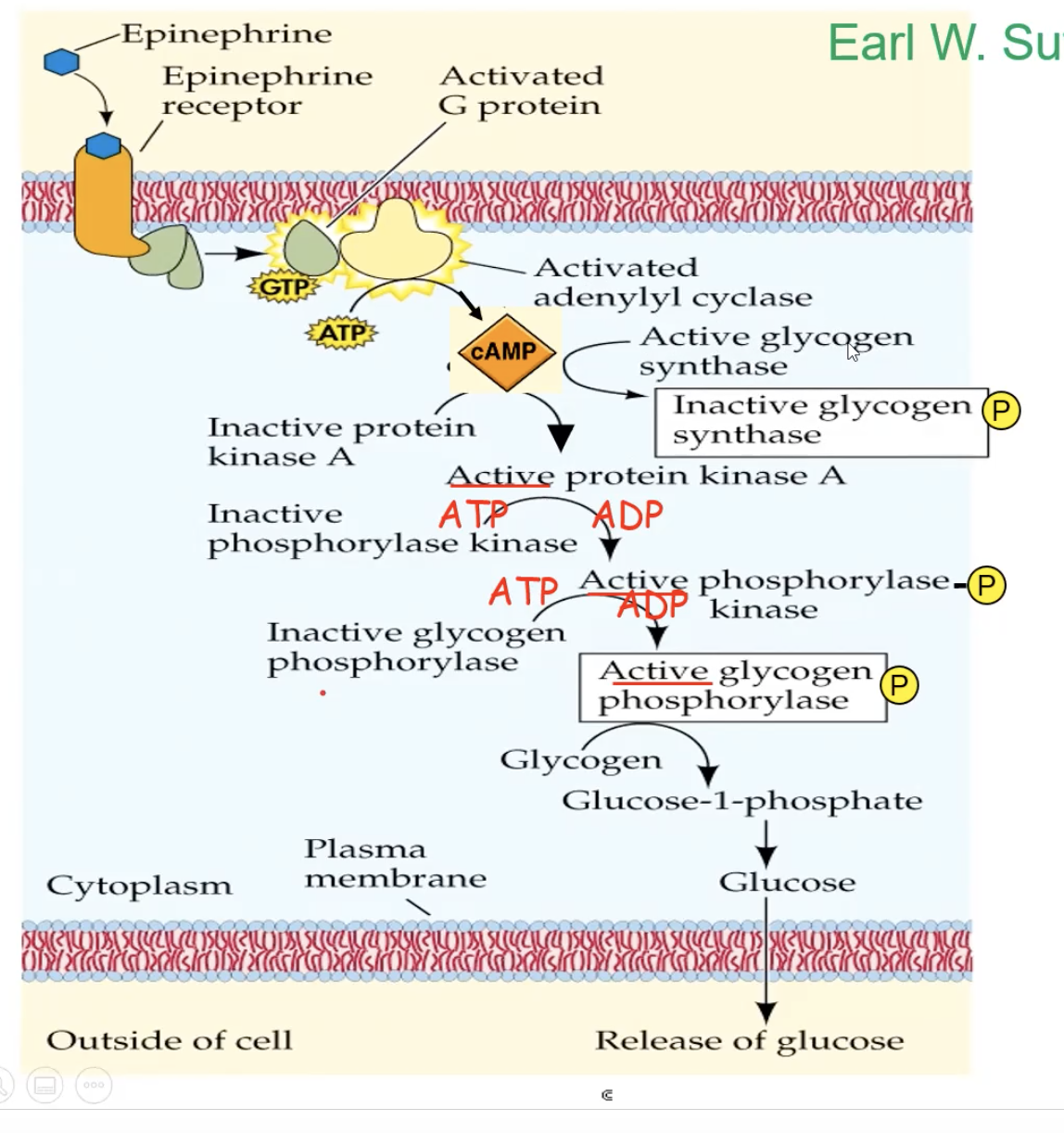
Full cycle of Epeniphrine binding to G protein receptor
epinephrine binds to the G protein receptor, creating a conformational change
The conformational change triggers the receptor to make contact with a G protein
The contact will make the G protein drop its GDP. and pick up GTP, which also activates the G protein
The activated G protein will go to an adenylyl cyclase enzyme, activating the adenylyl cyclase
Adenylyl cyclase, once activated, takes ATP and turns it into cAMP (secondary messenger)
cAMP binds to inactive protein kinase A and activates it
A phosphorylation cascade occurs, and the protein kinase A will phosphorylate phosphorylase kinase, which activates it, which then phosphorylates an inactive glycogen phosphorylase, which activates it, and the active glycogen phosphorylase will take glycogen and break it down and eventually release glucose from the cell.
At the same time, glycogen synthase gets inactivated to stop the production of glycogen
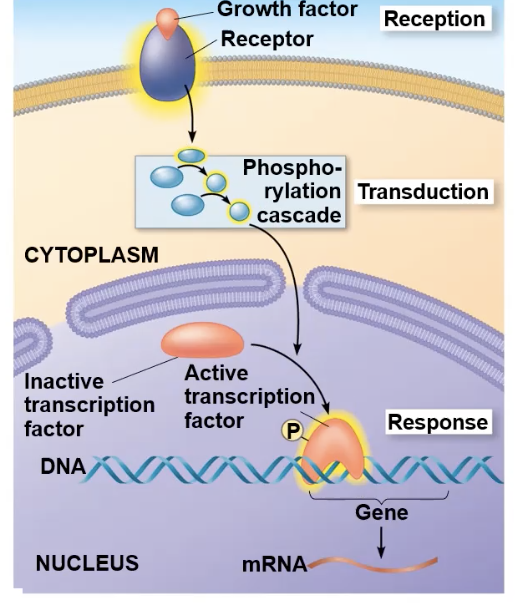
result of phosphorylation cascades
Phosphorylation cascades don’t just have to end with breaking molecules down.
They can also lead to the final kinase entering the nucleus and alter a transcription factor
phosphorylation cascade and transcription factors
The kinase series gets activated
the last kinase in the cascade enters the nucleus and binds to. an inactive transcription factor, creating an active transcription factor
The activated transcription factor binds to a specific DNA sequence and activates gene expression
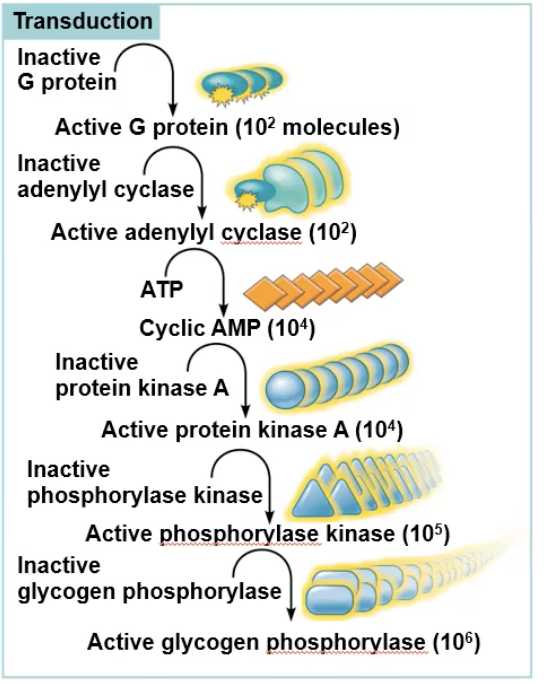
Amplification
A single binding can cause amplification, which is when a small initial stimulus leads to a large and significant cellular response
Example: 1 epinephrin binding can create around 100 active G proteins that diffuse out and activate 100 enzymes, which will produce 1000 cAMP, which will bind to 1000 pK-A, which will phosphorylate 100000 phosphorylase kinase, which will phosphorolyze 1000000 glycogen phosphorolase, which will break down and release 100000000 glucose molecules out of the cell.
reversable modifications
stop a signal after its serves its purpose, allowing it to return to its resting state.
protein phosphatase is an enzyme that assists in reversible modifications
protein phosphatase
enzymes that work to cut off added phosphate group in phosphorylation. This deactivates the proteins and stops its role in the phosphorylation cascade
Calcium ion as a second messenger
Ca+ is small, water-soluble, and freely diffusible, making it a good second messenger
There is a low concentration of it in the cytosol because Ca+ gets pumped out of the cell, as well as pumped inside storage organelles like the mitochondria or ER
When Ca+ concentration DOES rise in cytosol, any protein dependent on calcium concentration can become activated
Protein Kinase C is a protein responsive to calcium concentration
Calcium as secondary messenger
many pathways use calcium instead of cAMP or in addition to cAMP, as a secondary messenger
Ca+ is small, water-soluble, and freely diffusible, making it a good second messenger
There is a low concentration of it in the cytosol because Ca+ gets pumped out of the cell, as well as pumped inside storage organelles like the mitochondria or ER
When Ca+ concentration DOES rise in cytosol, any protein dependent on calcium concentration can become activated
Protein Kinase C is a protein responsive to calcium concentration
Where is Ca2+ located in the cell?
mitochondria or ER until the channel opens
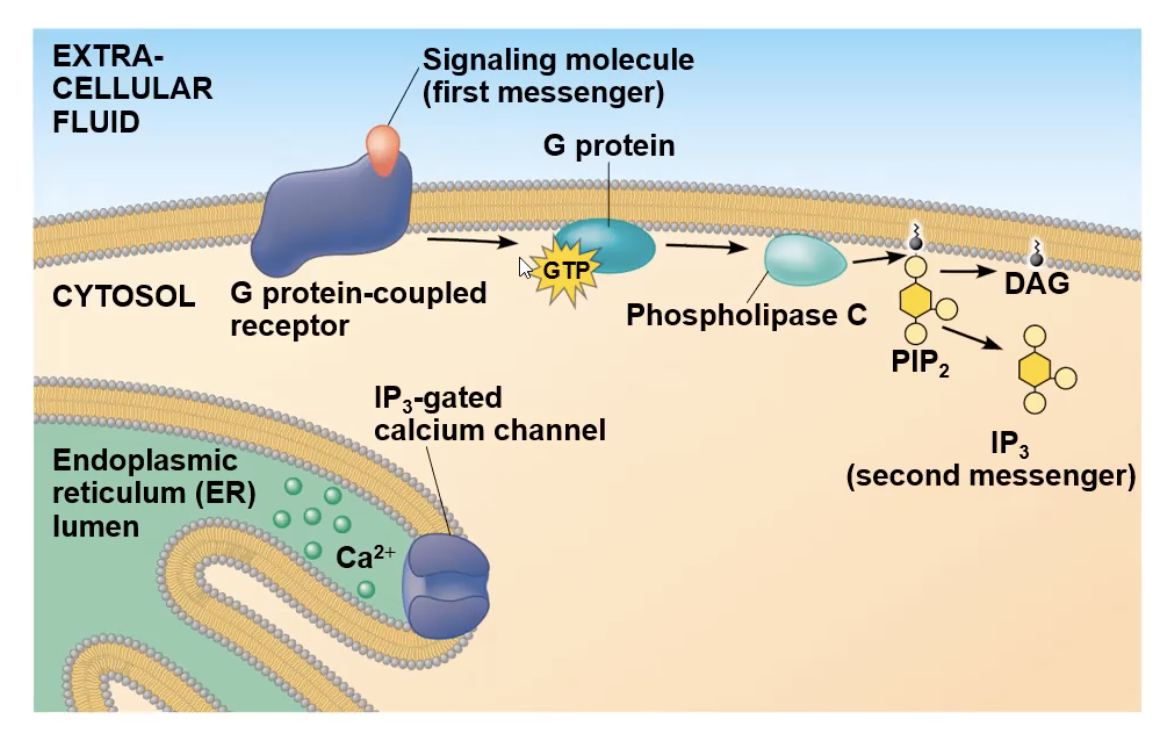
phospholipase C
an enzyme linked to a pathway involving Ca ions.
can cut phospholipids
After first messenger binds to G protein receptor, the G protein will interact with the receptor and drop its GDP and gain GTP, which activates it to be mobile, and attach to phospholipase C.
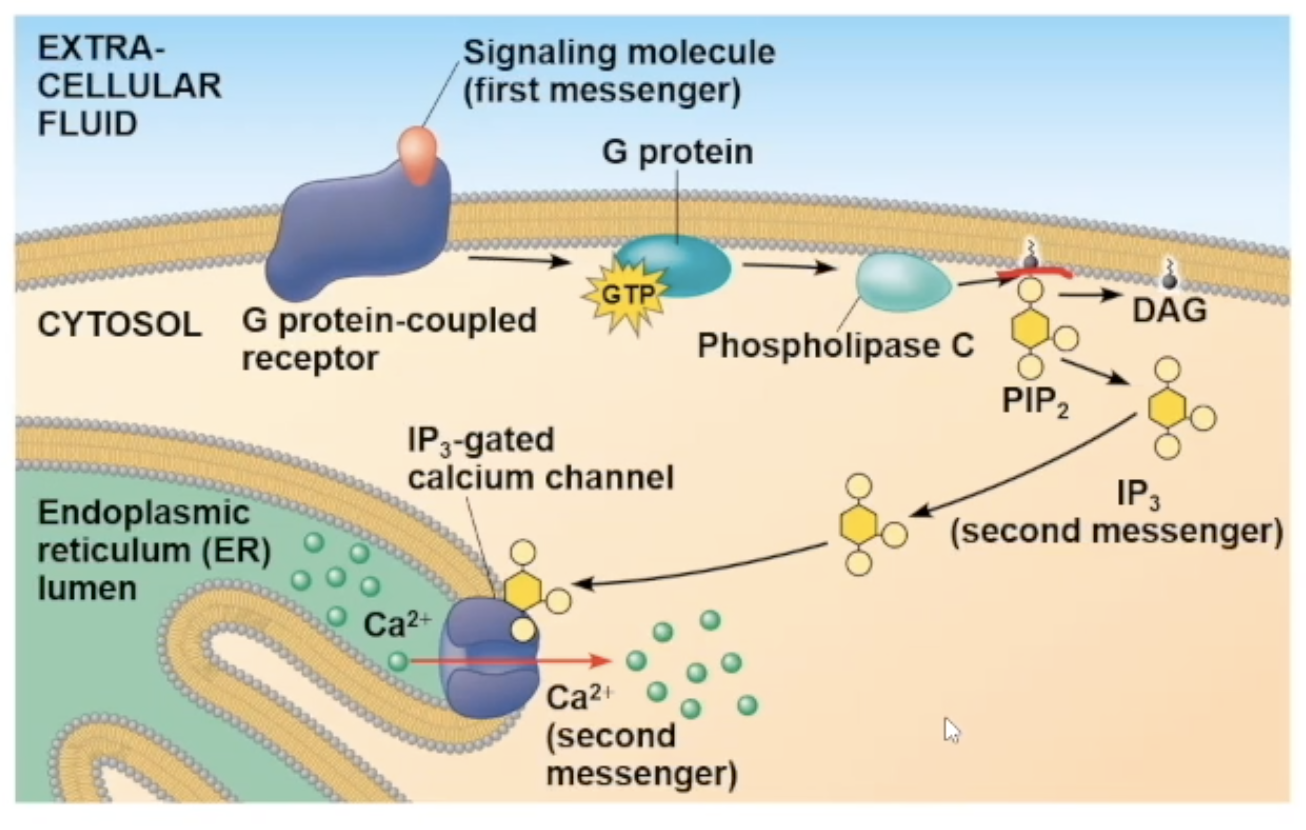
Full Ca pathway
signal molecule attaches to receptor
receptor goes through a conformational change and will interact with G protein
interaction with G protein causes it to lose its GDP nad gain GTP, causing it to be mobile and travel to phospholipase C
phospholipase C activates, cuts PIP2 molecule creating IP3 molecule (the second messenger)
IP3 diffuses to a gated calcium channel and binds to it. the binding opens the channel
The channel opening allows the Ca2+ ions inside the ER to travel out to the cytosol
The ions can bind to many different protein types, and depending on what it bind to cause a specific cellular response
protein Kinase receptor
class of cell surface receptors that play a crucial role in signal transduction pathways
resposible for protein phosphorylation
a specific one we will focus on is the receptor tyrosine kinases (RTK)
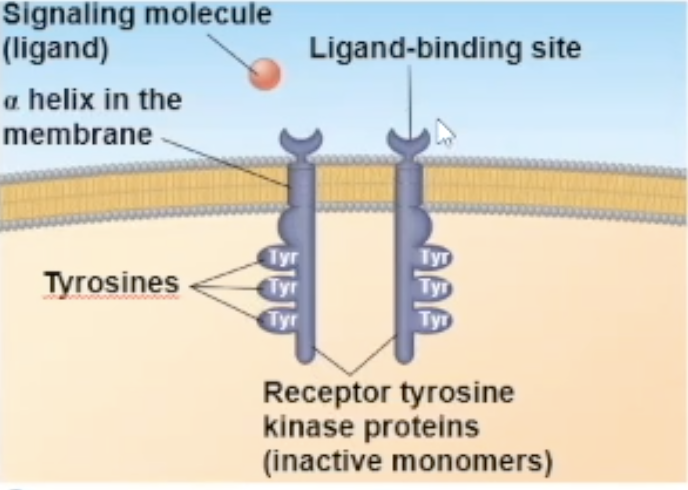
Receptor tyrosine kinase (RTK) structure
one RTK monomer has
a binding site that the ligand can bind to
a side in the cytoplasm that has RTK proteins on them to help with its enzymatic fuction
The side in the cytoplasm has tyrosine proteins on them
RTK binding
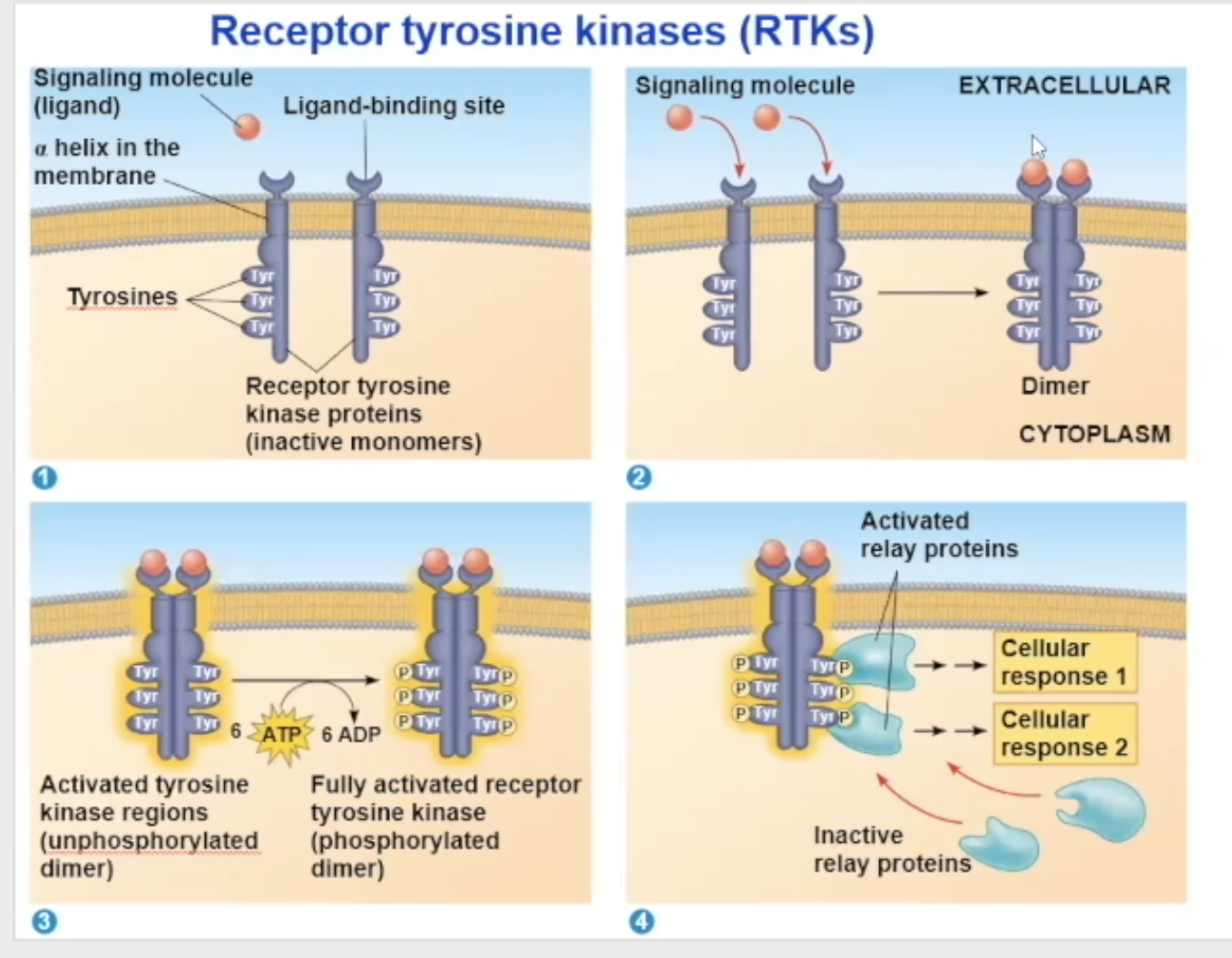
Either 2 signal molecules can bind to 2 independent RTK molecules, causing them to combine and turn into a dimer, OR 1 signal molecule can bring the two RTK molecules together and bind to both at the same time
receptor tyrosine kinases (RTK) process
Their unactivated state is independent, but when activated, it combines with another RTK receptor
The dimerization activates the molecule and goes through autophosphorylation, which means the two monomers put phosphate groups on each other’s tyrosine proteins
the phosphates come from ATP, so at the end of the auto-phosphorylation, it converts its used ATP to ADP
Now in its fully phosphorylated form, the dimer can bind with relay proteins
the binding of the relay prtiens is the beginning of the tranduction
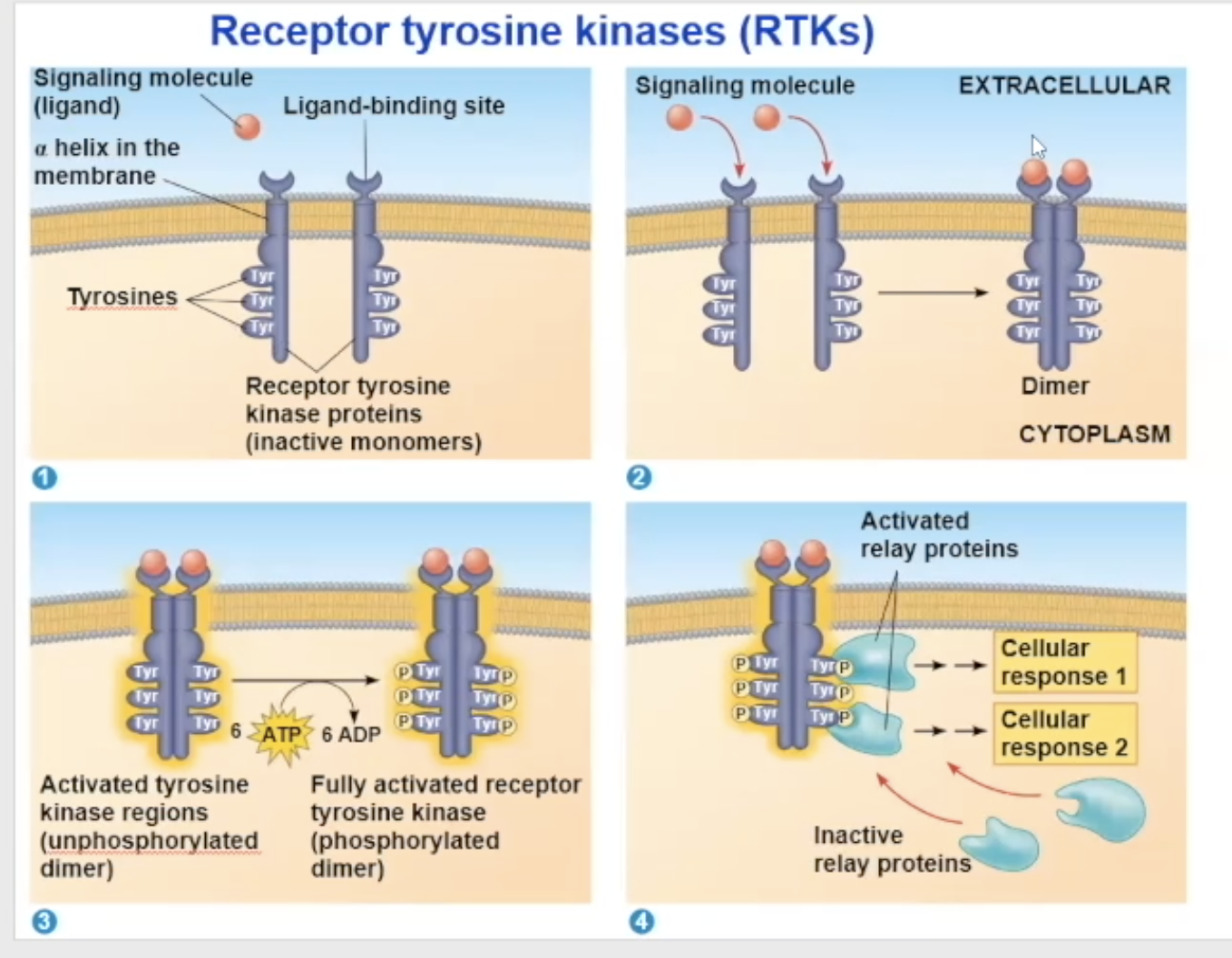
Scafollding peoteins
multiple protein kinases can combine together with a scaffolding protein
this increases the signal transduction efficiency because multiple proteins are already groped together and can bind to a receptor together
Termination of Signals
After a signaling molecule sends a signal which brings about a response, and then it needs a way to end the signal.
→ They can use
reversible binding
turning off enzymes
reversible binding
The two molecules (ligand and receptor) will join together and come apart
(The signaling molecule diffuses away from the receptor)
After inactivating all the proteins affected by the initial binding inactivates as well
for example an activated G protein with a GTP will turn off by hydrolyzing the GTP and gaining GDP
GTPase
when a G protein can hydrolyze its bound GTP (turning off on its own)
whena g protein is not a GTPase, it will get assistance from an enzyme to shut off
Termanation of signal- what happens to the ezyme
the protein will diffuse away from the enzyme, making it inactive again
all the molecules made by the enzyme gets destroyed
phosphodiesterase
An enzyme that destroys all the second messengers created by the enzyme after the enzyme is inactivated
What if the Termination of a signal does not occur? (example)
Vibrio cholerae (cholera) produces a toxin that modifies a G protein so its stuck in active form
this causes the protein to continually produce cAMP causing intestinal cells to secrete large amounts of salt into the intenstines
water follows by osmoses
an untreated person will lose salt and water and can die
how are cells different from eachother?
Cells are different from each other because they have different collections of proteins
They have different proteins within them because they have different genes being expressed
specificity of cell signaling
the arrival of the same signaling molecule in different cell types can bring about different effect
the different effects can be becuase
the proteins within the cells are different
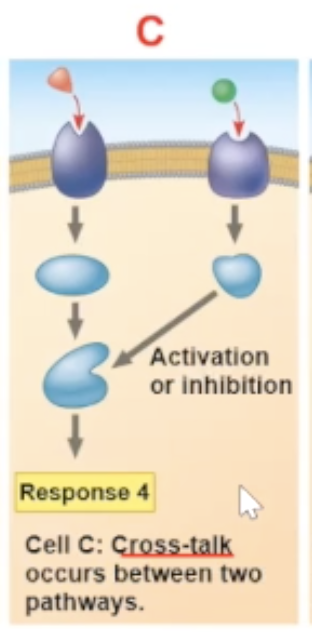
a cross talk between two signal molecules binding can cause a new pathway
the signal molecule binds to a different receptor
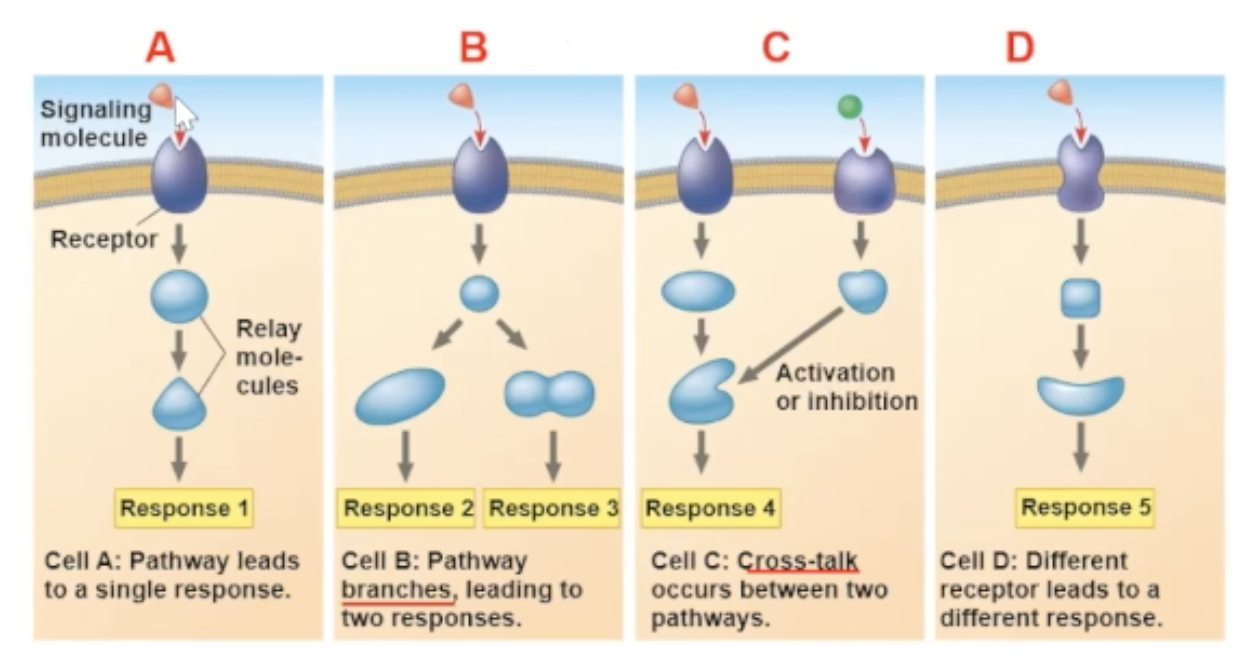
Why does the Same signal-molecule/receptor binding bring different pathways for different cell types?
The proteins in the cells are different.
Cross-talk
a cross-talk is when two different signaling molecules bind to their receptors, and the result of the two binds occuring leads to a new path
a cross-talk is another reason why different cells can have different responces to the same signal molecule binding
different receptor
different receptors will communicate with different relay protein
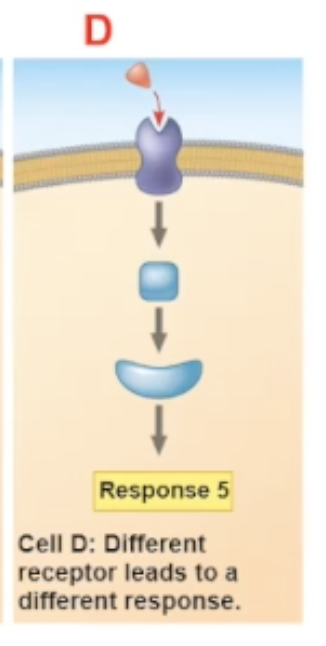
acetycholine binding with different cells
A. heart pacemaker cell- binds to G protein receptor cell. Upon binding it decreases the rate of firing
B. salivary gland cell- binds to G protein receptor cell. Upon binding, the cell secretes saliva
C. skeletal muscle cell- binds to an ion channel-coupled receptor. upon binding, it allows for contraction of the muscle
epinephrin binding to different cells
A. liver cell- binds to its beta receptor. upon binding breaks down glycogen and releases glucose
B. skeletal muscle blood vessel- binds to beta receptor. Upon binding, the vessel dilates
epinephrin binding to same cell different receptors
Receptors in the blood vessel
A. Beta receptor- upon binding, the vessel dilates
B. Alpha receptor- upon binding, the vessel contracts
Signal for apoptosis (example)
for soil worm caenorhabditis elegans
apoptosis is triggered by signals that activate a cascade of “suicide” proteins
when death signal is received, an apoptosis inhibiting protein (Ced-9) is inactivating allowing for the caspase “suicide” proteins to promote apoptosis
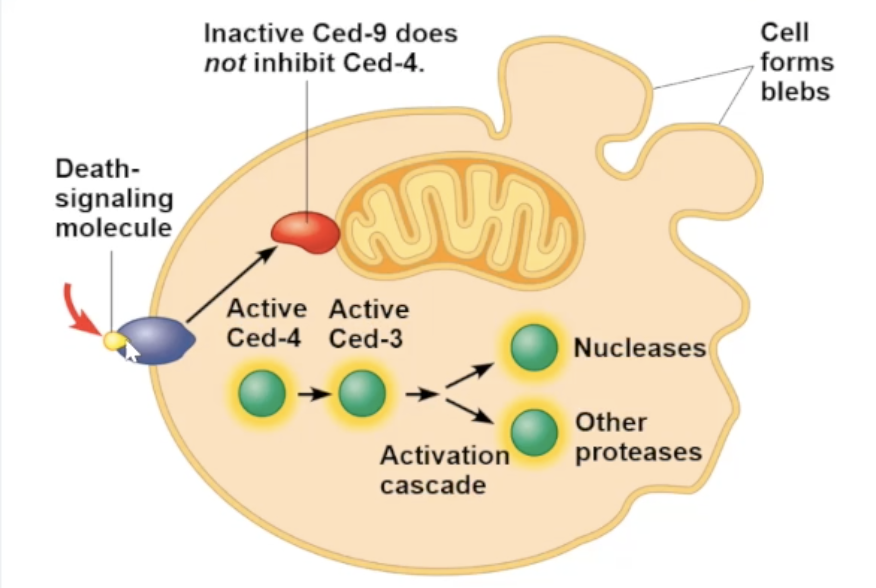
Ced-9 protein
protein inhibiting apoptosis. usually activated unless inactivated
Capases
proteins that carry out apoptosis
Apoptosis pathways and signals triggering them
humans have several different pathways to carry out apoptosis
apoptosis can be triggered by signals outside the cell or inside
internal signals include- DNA damage, protein misfolding
apoptosis steps
initial- sudden release of cytochrome c from the mitochondria into cytosol
second- the lipid asymetry of plasma membrane breaks down (phosphatital serine is found in the cytosol, but during apoptosis it is found outside the cell and other cells such as macrophages can consume the content)
final- the plasma membrane becomes permeable to small molecules