Biological Anthropology Exam #3
1/216
Earn XP
Description and Tags
Name | Mastery | Learn | Test | Matching | Spaced |
|---|
No study sessions yet.
217 Terms
We are hominins
Most scientists now use the term hominin to refer to modern humans and their direct ancestors, and hominid to refer to all of the great apes
Divergence and the last common ancestor
Use comparative method
Fossil record of modern apes is very poor
Lack of fossil evidence likely reflects habitat differences
Molecular evidence indicates that the divergence between the human and chimpanzee lineage occurred approximately 6-8 million years ago
Gene regulation
Indicates that differences between humans and the apes are more likely due to changes in gene regulation than the structural genes themselves
Evolutionary novelites of humans
Habitual upright walking (bipedalism)
Characteristics of the dentition
Elaboration of material culture
Significant increase in brain size
Long developmental period and long lifespan
Mosaic evolution
Different traits evolve at different points in time
Dentition
2.1.2.3 dental formula (same as in all Catarrhines)
Y-5 lower molar pattern
Canine reduction
Canine size and shape are associated with behavioral differences in apes
Reduction of sexual dimorphism in hominins → less male-male competition → different social interactions
Canine in apes
3rd Premolar (CP3) honing complex of apes
When the mouth is closed, the large canines interlock, each fitting into a space, called a diastema, in the tooth row of the opposite jaw
In the upper jaw, the diastema is in front of the canine, while in the lower jaw it is behind the canine
The lower premolar is larger than the premolar behind it
Has a single enlarged cusp
Its anterior or front surface is smooth and elongated, and it presents a sharpening edge for the canine
The 3rd premolar is often referred to as a sectorial premolar
Sectorial = adapted for cutting
Back surface of the upper canine is sharpened, or honed, against the blade-like 3rd premola

Canine size & sexual dimorphism
All great apes show sexual dimorphism in the canines
Great ape males engage in significant competition, often involving physical aggression, with other males over access to females

Ape vs. human dental arcade
Dental arcade = shape of the tooth row
Apes:
U-shaped dental arcade
Large canines, broad incisors
Diastema
Human:
Parabolic dental arcade
Smaller anterior teeth
No diastema
Dental reduction


Dentition: prognathism
Refers to the degree that the face projects forward, in front of the brain case
Chimps:
More facial prognathism
Humans:
Less facial prognathism
Small, shorter mandible
Muscles of mastication (chewing): temporalis
Extends from its origin on the vault of the cranium, passes behind the cheek bone (or zygomatic arch), inserts onto the top of the mandible near the jaw joint
Functions in closing the jaw
Smaller in modern humans
Muscles of mastication (chewing): masseter
Extends from the cheek bone to the bottom of the mandible
Functions in moving the jaw forward, and side to side
Smaller in modern humans
Apes skull and muscles of mastication (chewing)
Sagittal crest
Robust, flaring zygomatic bone
In modern humans, with small chewing muscles, the zygomatic is slender and not flaring
Temporalis
In gorillas, this muscle is very large, while the brain case is relatively small
Masseter
Masseter is enlarged, to accommodate high chewing force
Arises on the zygomatic arch and insets on the mandible
Modern human reduction in chewing muscles
Human bipedalism exceptionalism
Humans are the only primates – the only mammals – that do this all the time
Anatomy of bipedalism
Suspensory locomotion & vertical climbing
Increased mobility of extremities
Shoulder blade located on back
Forelimbs elongated compared to hindlimbs
Long and curved fingers for grasping branches
Knuckle-walking
Wrist joints are stabilized
African apes have long upper limbs (inherited from an ancestor with suspensory locomotion)
Dorsal position of the shoulder blade
Humans have dorsally positioned shoulder blades as well
Center of gravity
Fixed point, through which body weight is transmitted or balanced
When humans stand, the center of gravity is situated directly in the midline
Only minimal muscle activity is needed to maintain standing posture

Foremen magnum position
Positioned directly underneath the skull in humans

Body proportions
Intermembral index = [forelimb / hindlimb] x 100%
Chimpanzee ~ 110%
Human ~ 70%
Increased stride length
Vertebral column in modern humans
Cervical (neck) and lumbar (lower back) curvatures to maintain center of gravity over the pelvis
Larger size of the lumbar vertebrae to support body weight
Pelvis shape in modern humans
Wide, basin-shaped pelvis
Short, broad, curved iliac blades
Medial rotation of the ilium in humans
Repositioning of the gluteal muscles → Improved lateral stability during swing phase of bipedal walking

Modern human knee
Valgus angle of the knee
When humans walk, the foot falls directly below the center of gravity
Femur is oriented at an angle

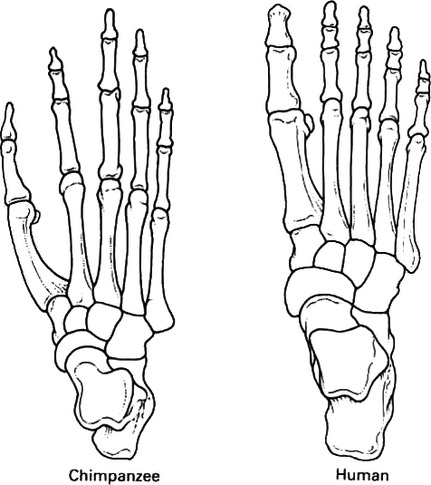
Modern human feet
Big toe is not opposed to the other four digits, and is enlarged in size
Enlarged heel (calcaneus)
Development of arches
Adaptive explanations for the origin of bipedalism: social factors
Ability to provision for the family in the context of the evolution of monogamous systems
Carrying tools, food, or infants
Provisioning family
Adaptive explanations for the origin of bipedalism: ecological factors
Moving across forested patches with higher energetic efficiency
Finding food and spotting predators
Primate evolution timeline

Miocene
Age of apes
Earliest human ancestors
Oligocene
Anthropoids (parapithecids, propliopithecids)
Eocene
Earliest definite primates (adapids, omomyids)
Paleocene
Possible primate ancestors (plesiadapiforms)
Timeline
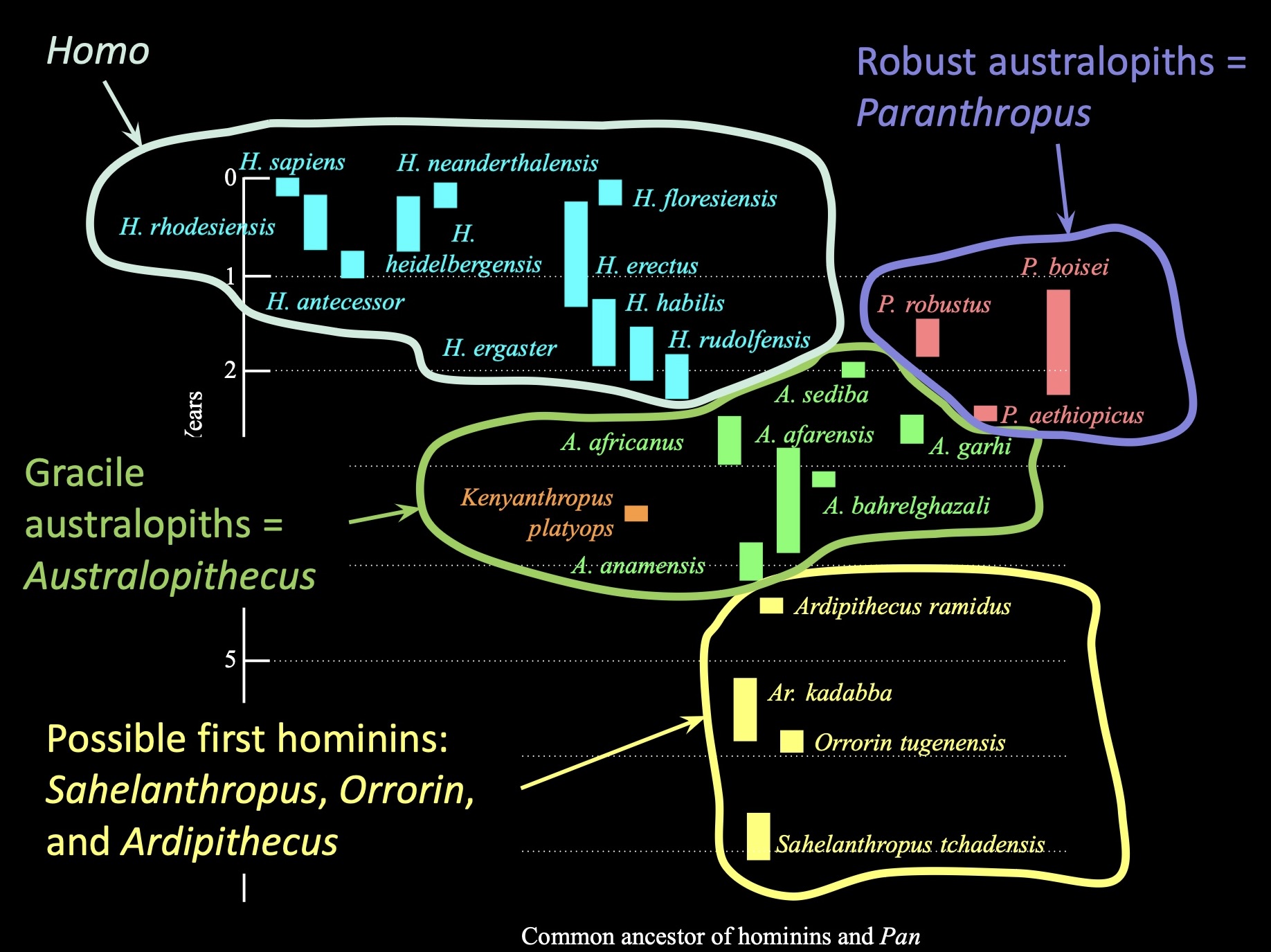
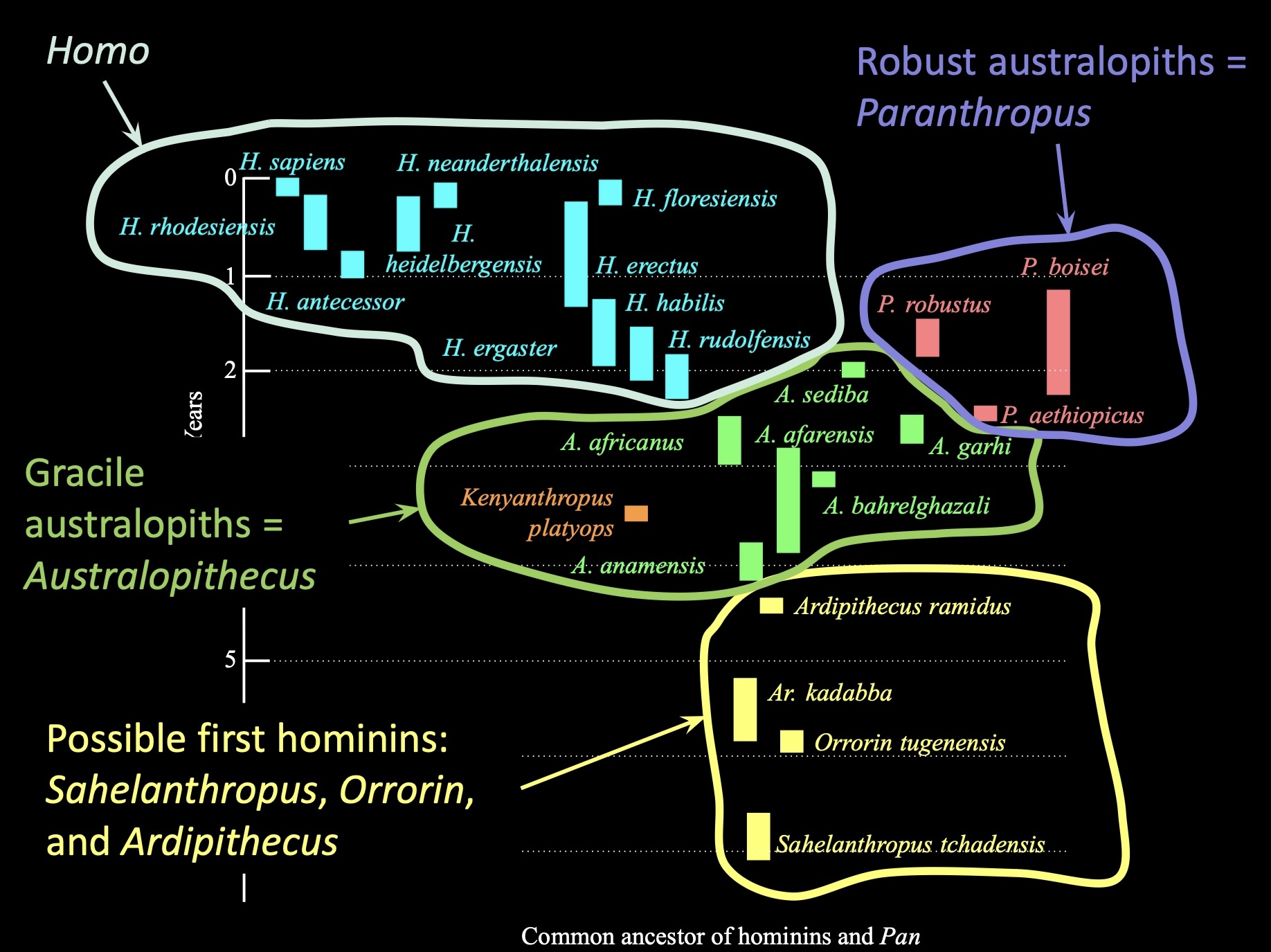
Between 7-4.4 million years ago, we have evidence in the fossil record of a small number of candidate species
Ardipithecus ramidus, Ar. kadabba, Orrorin tugenensis, and Sahelanthropus tchadensis
Geographical location of possible hominins
Early hominin fossil localities are primarily from east and south Africa
Nearly all fossil evidence from the first 4 million years of human evolution (7-3ma) comes from East Africa, from a region known as the Great Rift Valley
S. tchadensis
Chad
7-5.2 Ma
O. tugenensis
Kenya
6 Ma
A. ramidus
Ethiopia
4.4 Ma
A. kaddaba
Ethiopia
5.8-5.2 Ma
East Africa rift system
Area where three continental plates are pulling apart
Region yields lots of fossils
Most of these fossil sites have been well dates
Ash, chronometric dating techniques
Lake habitat


Possible earliest evidence of bipedalism: Sahelanthropus tchadensis
Chad
7-6 ma
More vertical face, higher skull vault
Smaller canines, no C-P3 honing complex
Possibly biped, position of the foramen magnum
Primitive in other respects (brain size, U-shaped dental arcade)


Possible earliest evidence of bipedalism: Orrorin tugenensis
Kenya
6 ma
Fragmentary cranial and postcranial remains
Femoral morphology is indicative of bipedal locomotion
Dental morphology is ape-like (large canines)

Ardipithecus
Intermediate canine size between apes and later hominins
2009, “evolution’s bad girl”
Environment, reconstructed by its discoverers to be a more closed woodland habitat
45% partial female skeleton
Ardipithecus kadabba
5.2 - 5.6 ma
Projecting and interlocking upper and lower canines
C-P3 complex was intermediate
Ardipithecus ramidus
4.4 ma
Combination of primitive and derived traits
Evidence of its ancestral position for the hominin clade for Ardipithecus discoverers
Evidence of the position of A. ramidus as an early ape or an evolutionary side branch for others
Relatively small brain, prognathism as in apes
More forwardly-placed foramen magnum - consistent with bipedalism
Reduced canine size
Loss of C-P3 honing
Minimal sexual dimorphism
Pelvis - possible evidence for bipedalism
Mosaic of characters for both bipedality and climbing
Short and broad ilium (unlike chimps)
Ischial surface is primitive
Foot retains a divergent big toe
Lacked other features for suspension/vertical climbing and knuckle walking seen in apes

Lovejoy’s model for the origins of bipedality

Major trends seen in australopiths (Australopithecus and Paranthropus)
Dentition – reduced canine size; larger cheek teeth; thicker enamel
Paranthropus chewing adaptation, postcanine megadontia
Locomotion/posture – adaptations for bipedalism
Relative brain size – not substantially enlarged compared to apes
Body size - smaller than modern humans, with more sexual dimorphism
Geographical location of australopiths (Australopithecus and Paranthropus)
Two kinds of bipedal ancestors with large teeth and small brains known from E. and S. Africa
Australopithecus (gracile)
Paranthropus (robust)
Gracile forms now also found in Central Africa
The genus Australopithecus
Australopithecus, meaning southern ape, was coined by a scientist in South Africa named Raymond Dart, after discovering the Taung child
Not accepted for decades because of Piltdown
Piltdown hoax
The Piltdown Man was a cranium and jaw, said to represent the fossilised remains of a previously unknown early human
Fragments consisted of parts of a skull and jawbone, said to have been collected in 1912 from a gravel pit at Piltdown, East Sussex, England
Eoanthropus dawsoni ("Dawson's dawn-man", after the collector Charles Dawson)
Exposed as a fraud by geochemical tests showing the mandible and cranium couldn’t have come from the same sediments (1953)
Orangutan and modern human
Australopithecus anamensis
4.2 – 3.9 ma
Kenya and Ethiopia
Derived traits compared to apes:
Reduced canines
Larger molars, thick enamel
Evidence of bipedalism
Compared to later hominins, retains some primitive traits:
More U-shaped dental arcade
Open woodland/gallery forest environment and open savanna environment
Recently discovered MRD VP 1/1 cranium “puts a face to a name” for this species
Project Leader Yohannes Haile-Selassie

Australopithecus anamensis tibia
4.2 – 3.9 ma, Kenya
Australopithecus anamensis
Earliest undisputed evidence of bipedalism
Enlarged proximal (top) end of the tibia
Likely part of an ancestor-descendant lineage with A. afarensis

Australopithecus afarensis
Best-known early hominin species
3.9 - 2.9 ma
East Africa
Kenya, Tanzania, and Ethiopia
Finds:
Lucy
Dikika, Ethiopia
3.3 ma
Apx. 3 years old
Was smaller-bodied than modern humans
Significant sexual dimorphism
A. afarensis: 56%
Gorilla: 50%
Lucy
Australopithecus afarensis
3.2 mya
40% complete skeleton of a single individual
13 more individuals found in the same area as her
Lumbar Curvature
Valgus Knee
Short toes
More forward placement of the foramen magnum
Short, broad pelvis; curved ilium
Enlarged heel, arches, non-opposable big toe
Australopithecus afarensis: skull
Sagittal crest
Small brain size relative to face size
Relatively prognathic
Strong nuchal (neck muscles) crest for neck muscle attachments

Australopithecus afarensis: teeth and jaw
Upper jaw
Canine further reduced
Posterior tooth rows near-parallel, converge slightly
Lower jaw
Lower 3rd premolar usually bicuspid (not sectorial)
Slightly larger cheek teeth
Thick molar enamel
Thick enamel on teeth suggest they may have eaten nuts, grains, or hard fruit pits, but also probably still eating fruits
Laetoli, Tanzania
3.6 ma
Trail of fossil footprints in Laetoli, Tanzania
Proves that a bipedal hominin lived in East Africa at the same time as A. afarensis
75 feet long trail made by three individuals who had crossed a thick bed of wet volcanic ash
Adducted big toe
Development of the arches
Deep depression of the heel

Arboreal features of Australopithecus afarensis
Relatively long and curved hand bones
Highly mobile shoulder joints and upwardly-oriented shoulder blade
Relatively long upper limbs compared to short lower limbs
Australopithecus afarensis: diverse habitats
Lived in environments ranging from more closed woodland to dry open grasslands
Ability to walk and climb trees would have allowed them to use resources from all of these environments
Early Australopithecus Fossil Sites
Chad (Bahr el Ghazal)
Australopithecus bahrelghazali
3.5 – 3.0 Ma
Hadar and Middle Awash, Ethiopia
Australopithecus afarensis
Kenya (Omo, W Turkana)
Australopithecus afarensis
Tanzania (Laetoli)
Australopithecus afarensis
Kenyanthropus platyops
East Africa
3.5 ma
Distinctive combination of flat lower face and small molars
Perhaps A. afarensis

Hominin localities – South Africa
Plio-Pleistocene
A. africanus
Makapansgat, Taung, Sterkfontein, Gladysvale
Paranthropus
Sterkfontein, Kromdraai, Drimolen, Swartkrans
Homo
Sterkfontein, Swartkrans
Australopithecus africanus: Taung child
3.5-2 ma
South Africa
Taung child, with endocranial cast
Grew up according to a schedule much like that of a chimpanzee
Thought have been attacked and killed by an eagle

Formation of South African sites
Cave sites, formed as layers of bedrock were dissolved and filled with sediments
No volcanic ash layers → chronometric dating is extremely difficult
Therefore using relative dating
“Osteodontokeratic culture”
Bone tooth horn, “killer ape” theory
Disproven by taphonomy
Dart
Assemblages represent bone accumulations – hominins were not thought to have actually lived in these caves
Australopithecus africanus: Sts 5, female
Slightly enlarged brain, & rounder cranial vault.
Less prognathism compared to A. afarensis
Less developed nuchal cresting
Reduced anterior dentition

Australopithecus africanus: Cranial capacity
Brain averaged 442 cc, which is slightly larger than A. afarensis
Still, significantly smaller brains than modern humans (just ~ 1/3 the size)

Australopithecus africanus: Sts 71, male
Cheek bones swept forward, nasal pillars
Robust mandible
Larger cheek teeth with thick molar enamel
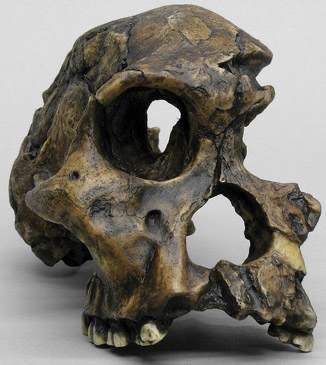
Australopithecus garhi
East Africa
2.5 ma
Small brain (450cc), projecting face, with large front teeth like more gracile forms, and very large back teeth like more robust forms
Limb proportions are unusual: long forelimbs (as in earlier forms), but also with relatively long legs (as in Homo)
Possible tool use
Found close to animals bones which show very clear signs of butchery with stone tools
Australopithecus sediba
2 ma
South Africa
Similarities with Australopiths:
Small brain size
Molar morphology
Small body size
Long upper limbs
Similarities with Homo:
Projecting nose
Smaller teeth and chewing muscles
Longer legs
Hand with precision grip
Berger and his colleagues have argued that A. sediba might be the long-sought species that gave rise to our genus (or a close relative of that species)

Major evolutionary trends at 2.5 M
Significant cooling trend around 2.6 ma
Expansion of open grasslands at the expense of closed forests
Homo
Increase in brain size and capacity for tool making
Decrease in prognathism, and postcanine tooth size
Increase in body size
Paranthropus
Hypermasticatory complex
Increase in cheek tooth size
Body size similar to Australopithecus
Global cooling trend

Global cooling trend associated with drier, more open habitats in tropical Africa
Robust australopiths: Paranthropus
Generate large bite forces at their molars, and made them extremely efficient at chewing
Large and forwardly-placed sagittal crest
Reorient chewing forces towards the back of the dentition
Enlarged cheek teeth with thick molar enamel; frontal dental reduction and crowding
Dished face



Paranthropus aethiopicus
East Africa
2.7-2.5 ma
Classic robust features
Well-developed sagittal and nuchal crests
Prognathism; “dish-shaped” face
Relatively small brain
Well-developed sagittal crest
Forward-projecting cheek bones (zygomatics)

Paranthropus boisei
East Africa
2.3-1.2 ma
Later robust australopith, “hyper-robust”
“Visor-like” (Darth Vader) face
Forward-projecting cheek bones (zygomatics)
Forwardly-placed sagittal crest
Reduced prognathism

Thick enamel
Anterior dental crowding; marked reduction in incisors and canines
Enlargement of premolars and canines; postcanine megadonty

P. boisei and P. robustus
Paranthropus boisei
Olduvai Gorge, Tanzania in East Africa
2.3-1.2 ma
Paranthropus robustus
Several sites in South Africa, Kromdraai and Swartkrans
2.0-1.5 ma
Both take this hypermastication trend to an extreme
Tough foods, especially during times of resource scarcity
Hard food objects, tough plants
Different types of foods, and that what they ate varied throughout the year – varied seasonally
The first “type fossils” of Homo habilis
1964, OH 24 (skull) and OH 7 (partial jaw)
East and south Africa
2.4-1.4 ma
Why Homo habilis
Found at Olduvai Gorge, Tanzania
Found with flaked stone tools
Slightly larger brain
Oldowan tools
Mary and Louis Leakey
Early homo



Larger cranial capacity
Slight reduction in size of the cheek teeth; more parabolic dental arcade; thinner enamel
Reduced prognathism, more gracile cranium
But, similar in body size to australopiths
Associated with stone tools
Anatomically, they are more similar to Australopithecus
But classification as Homo has been classically supported by their ability to use and make stone tools (first tool-making species)
Now, evidence that other non-Homo species may have used tools
Is Australopithecus habilis a more accurate classification?
Early homo: one or two species
H. rudolfensis
2.4 – 1.6 ma
Larger brain size (mean = 775 cc)
Australopith-like broad midface
Relatively larger molar teeth
Well-developed mandible for chewing muscles

H. habilis
2.4 – 1.4 ma
Smaller brain size (mean = 612 cc)
Broader upper-face than midface
Smaller molars
Less robust mandible
More australopith-like limb proportions
Cranial capacity
Range Average
(cm3) (cm3)
Human 1150 - 1750 1325
Chimpanzee 285 - 500 395
A. afarensis 320-500 420
A. africanus 420-500 440
P. aethiopicus 410 410
P. robustus 475-530 512
H. habilis 612 612
H. rudolfensis KNM-ER 1470 775
H. erectus (E. Africa) 900-1067 980
H. neanderthalensis 1125-1740 1415
Oldowan tool industry
Early Stone Age / Lower Paleolithic; 2.6 Ma – 200 Ka
Cores, flakes, and hammerstones
Uses:
Access the carcasses of animals to obtain meat
Break bones to obtain marrow
Secondary uses:
Plant processing
Woodworking
Cores
Lumps of stone from which pieces are removed
Can be used for a smaller variety of tasks, including chopping down a tree to make a digging stick or spear
Flakes
Small fragments removed from cores
Cutting through thick hide and butchering a large animal such as an elephant
Hammerstones
Rounded stone used to remove the flakes
Crack nuts, break open bones to get at the marrow inside
Swartkrans, South Africa tool use evidence
1.8-1.0 mya
Digging tools made of bone and horn cores
Termites
Evidence for selectivity
Paranthropus or Homo?


Bouri, Ethiopia tool use evidence
~ 2.5 mya
Percussion & cutmarks on bone
No tools found
Australopithecus garhi

Dikika, Ethiopia tool use evidence
3.39 mya
Cutmarked bones
Flesh removal
Marrow access
Not found in association with hominins or tools
But, provides evidence for stone tool assisted meat consumption before Homo

Lomekwi
3.3 ma stone tools from Lomekwi, Kenya
Well before earliest Homo

Hominin tool traits shared with chimps
Nut cracking
Stone tool use
Stone transport (<2km)
Stone selectivity
Small game hunting
Unique hominin tool traits
Stone transport >10km
Stone tool manufacture
Using tools to make tools (woodworking)
Plant processing (USOs)
Large game acquisition, carcass processing
Earlier hominins may have used tools that do not have any trace in the fossil record → wood or other organic materials
Hunters or scavengers
Hominins probably practiced both hunting and scavenging
Opportunistic omnivores – incorporating a variety of plant foods, insects, and meat
Homo erectus sensu lato
Homo erectus, H. ergaster
1.8 Ma - ca. 30 Ka (300/200,000)
Cranial capacity ~ 880cm3
First hominin to have a body stature and limb proportions more similar to modern humans
First hominin species for which we have fossil evidence to migrate out of Africa into temperate regions of Asia and perhaps Europe
Suggests that this species was committed to life on the ground
May have been the first species to control fire


Pleistocene (1.8 Ma – 10 Ka) Climate Oscillation
Colder and more variable climate starting at 1.8 mya
Interval marked by repeated glacial cycles, known as the Ice Age
Huge volumes of water in continental ice sheets
Exposed land bridges connecting continents
Homo erectus/ergaster morphology
Cranial
Long, low and flat
Face
Broad flat face; less prognathic than H. habilis
Projecting nose
Brain
Larger
Postorbital constriction
Avg cranial capacity = 1,000 cc (range over time from 750 – 1250 cc)
Homo erectus: one or two species
Homo ergaster
Africa (~1.9 - 1.0 Ma)
Homo erectus
Asia (1.8 Ma - ca. 30 Ka)
Geographical variation in Homo erectus
Asian forms (Classic Zhoukoudien traits) - “Homo erectus”
Sangiran 17, Indonesia
Sagittal keel
Thick brow that forms a single torus, very pronounced
Receding forehead
Sharp occipital angle (pronounced torus)
African forms (with some exceptions) - “Homo ergaster”
KNMER 3733
No keel
Brows over each orbit, more curving
Less receding forehead
Less of an occipital angle
Footprints from 1.5 Ma, Ileret, Kenya (2009)
Indistinguishable from the footprints made by modern humans, indicating a modern human-like form of bipedal locomotion
Thought to have been made by Homo erectus

Turkana Boy
KNM-WT 15000
Homo ergaster
Africa
Adolescent skeleton, estimated at ~ 8 years of age based on dental development
At an estimated stature of 5’3” (~ 6’ as an adult), indicates a significant increase in overall body size compared to Australopithecus
Similar body proportions to modern humans
Acheulean tool industry
1.6 Ma – 200,000 years ago
H. erectus
More sophisticated technology
Symmetrical, biface tools
Retouching, soft hammer percussion

Handaxe proportions follow a mental template (unlike oldowan tools)
Tools have very regular proportions, standardized form
Proportions hold for Africa, Near East, Europe
Requires more complex cognitive abilities

Acheulean intelligence
Requires more complex cognitive capabilities than the Oldowan
Mental representation of a target image
Advanced planning to arrive at that product, and ability to modify technique to achieve that goal
Handaxe usage
“Swiss army knife”
For processing large animal carcasses
Tip cuts through joints & meat
Wood and other plants
Cores (as flake dispensers)
Not utilitarian purposes?

Control of fire: cooking
Makes food more digestible
Less food is needed to get the same amount of nutrients
Some toxins can be neutralized
Control of fire: warmth
Important in cold and seasonal environments out of Africa
May have been crucial to allow migrations to Asia and Europe
Control of fire: cave occupation
Allows the use of caves as shelters
Also important in the colonization of European and Asian environments
Control of fire: predator protection
Especially important for large groups
Might be predated upon at night