Evolution of Life-History Traits, Social Behaviors and Symbioses, Eusociality and Coevolution
1/32
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
33 Terms
Extreme reproductive strategies are uncommon but can be adaptive. Why is it adaptive for thrips egg mites and kiwi birds to reproduce the way they do?
Trade offs - E diversion
Thrip Egg Mites
eat mom
Optimal offspring:
sexually mature - early
higher quality offspring.
Mature at birth + already inseminated. Hatch inside mother and mate with brother. makes a clutch of eggs and dies at 4 days old being eaten alive by her offspring.
Kiwi Birds
Higher quality offspring → big, robust, likely to survive.
Mom 6lbs Egg 1lb
Month to make each egg
can be on their own after a week.
Male incubates egg for 3 months - loses 20% of his bodyweight.
What are some trade-offs between energy allocated for reproduction vs. other functions and size vs. number of offspring? Can you give examples?

For example, an individual can allocate energy to growth for a long time, which may enable it to reach a larger size and ultimately produce more offspring. This benefit of size, however, is balanced by a cost. The time required to grow is time during which predators, diseases, or accidents may strike. An individual that takes the time to grow large thus incurs a greater risk of dying without reproducing at all. Whenever there is a trade-off between different components of fitness, we expect natural selection to favor individuals that allocate energy and time with an optimal balance between benefits and costs, thereby maximizing lifetime reproductive success.
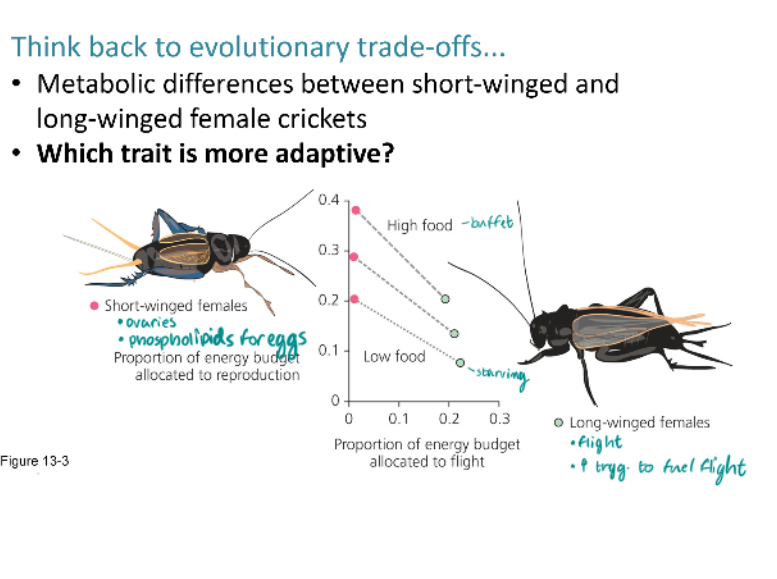
Is a specific trait always more adaptive? Or can reproductive traits shift in their advantageous-ness from season-to- season or from environment-to-environment?
Whenever there is a trade-off between different components of fitness, we expect natural selection to favor individuals that allocate energy and time with an optimal balance between benefits and costs, thereby maximizing lifetime reproductive success. Because different balances are optimal in different environments, environmental variation
Explain how senescence varies among animal groups. Why didn’t we discuss senescence in plants?
Aging, or senescence, is a late-life decline in an individual’s fertility and probability of survival
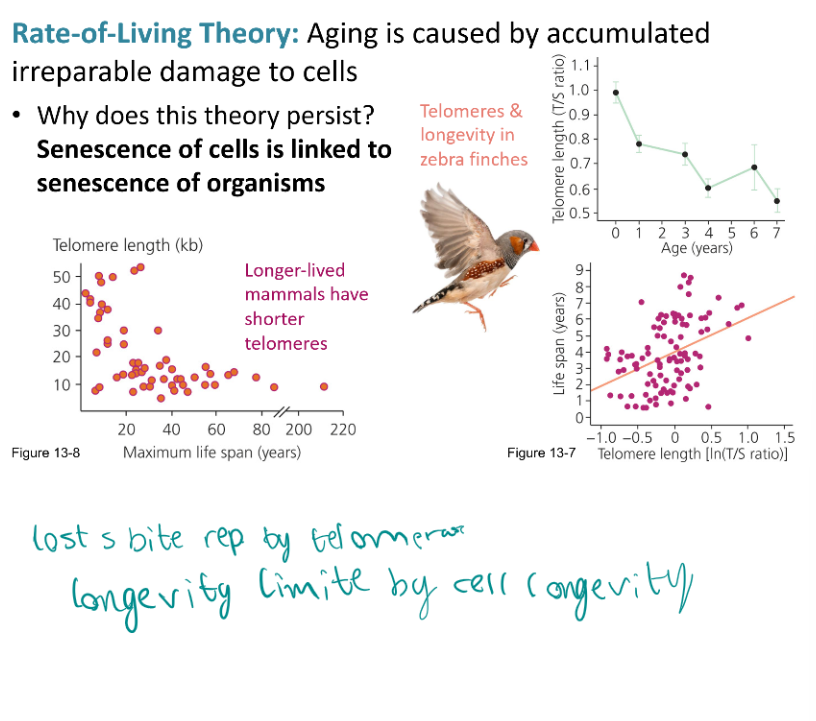
Explain the two theories for why aging persists in the face of selection, discuss evidence for and against those theories
Rate of Living Thory - aging is caused by accumulated irreparable damage to cells. Evolutionary constraint - populations lack the genetic variation to respond any further to selection against aging.
Each organism has reached the limit of biologically possible repair
Predictions:
Aging rate shoud be correlated with metabolic rate. (maxed our ability to repair)
Species should not be able to evolve longer life spans.
Aging is caused by accumulated irreparable damage to cells. Trade-off between the allocation of energy to reproduction versus repair.
Senescence of cells is linked to senescence of organisms.
Evolutionary Theory - Aging not caused by gradual damaging of cells, but by failure of organisms to repair this damage, due to deleterious mutations or trade-offs between repair and reproduction.
the failure of organisms to completely repair such damage. This failure leads to gradual decay and ultimate collapse.
the failure to completely repair damage is ultimately caused by either deleterious mutations or trade-offs between repair and reproduction.
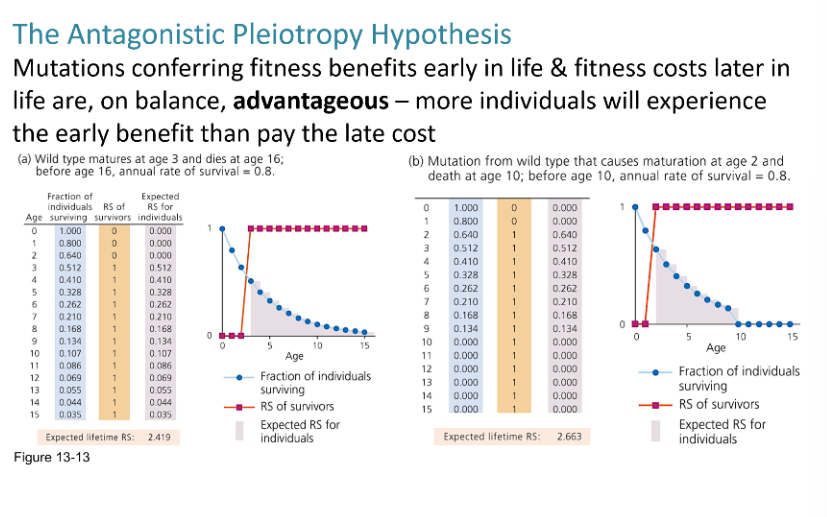
Antagonistic Pleiotrophy
Hox gene does multiple things.
Would you expect a mutation that confers a fitness advantage early in life, but leads to an earlier death, to increase in frequency? Why or not?
Yes, a mutation that improves early-life fitness can increase in frequency if it boosts reproduction, even if it causes earlier death later on. This is because natural selection is stronger on traits that affect reproductive success than on those that act after reproduction.
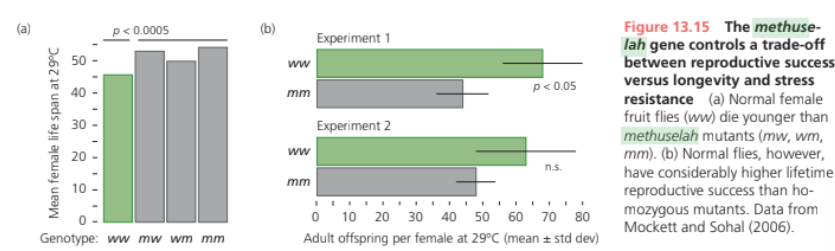
Explain the methuselah gene example – is it really a cost-free mutation?
Homozygotes for a mutant allele with reduced expression live 35% longer than normal flies. In addition to increasing longevity, it enhances resistance to starvation, heat, and the herbicide paraquat.
The mutants also lay fewer eggs during early adulthood, resulting in lower lifetime reproductive success. Compared to the mutation, the normal allele of methuselah thus appears to trade stress resistance and longevity for reproductive fitness.
What are two hypotheses for why women undergo menopause? What’s the evidence for and against each hypothesis?
Artifact Hypothesis: nonadaptive artifact of modern lifestyle.
Advocates of the artifact hypothesis point out that archaeologists reconstructing the demography of ancient peoples have often concluded that in premodern cultures, virtually all adults died by age 50 or 55 (see Hill and Hurtado 1991). If death by age 50 or 55 was the rule for our hunter-gatherer ancestors, then the modern situation, in which individuals often live into their 80s and 90s, is unprecedented in our evolutionary history. Menopause cannot be an adaptation, because our hunter-gatherer ancestors never lived long enough to experience it.
Critics of the artifact hypothesis point out that in contemporary hunter-
gatherer societies, many individuals live into their 60s and 70s (Figure 13.22).
These data may be more reliable indicators of the demography of our hunt-
er-gatherer ancestors than are archaeological reconstructions (Hill and Hurtado
1991; see also Austad 1994; Gurven and Kaplan 2007). If a substantial fraction
of our female hunter-gatherer ancestors lived long enough to experience meno-
pause, then menopause needs an evolutionary explanation.
Grandmother Hypothesis: adaptation associated with contribution of grandmothers to feeding grandchildren.
Advocates of the grandmother hypothesis note that human children depend
on their mothers for food for several years after weaning. This is true in contem-
porary hunter-gatherer cultures, particularly when mothers harvest foods that
yield a high return for adults, but are difficult for children to process (Hawkes et
al. 1989). Thus, a woman’s ability to have additional children may be substan-
tially limited by her need to provision her older, still-dependent children. Fur-
thermore, as a woman gets older, several relevant trends are likely to occur: (1)
The probability that she will live long enough to be able to nurture another baby
from birth to independence declines, (2) the risks associated with pregnancy and
childbirth rise, and (3) her own daughters will themselves start to have children.
The grandmother hypothesis suggests that older women may reach a point at
which they can get more additional copies of their genes into future generations
by ceasing to reproduce themselves and instead helping to provision their weaned
grandchildren so their daughters can have more babies. In other words, grand-
mothers face a trade-off between investment in children versus grandchildren.
Does selection favor a clutch size that produces the most surviving offspring? If not, why would a bird lay fewer eggs?
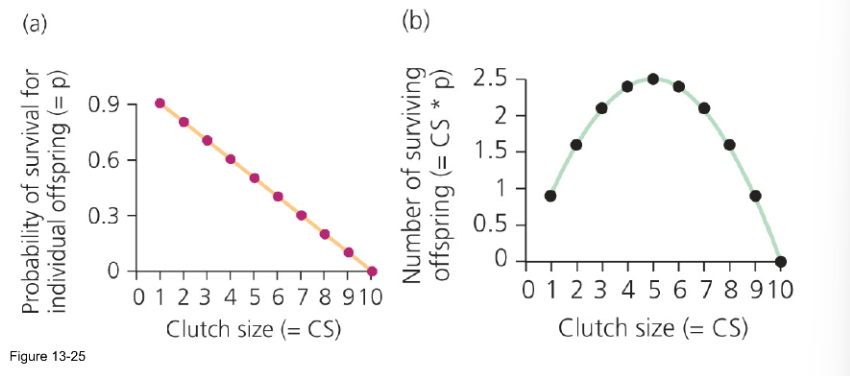
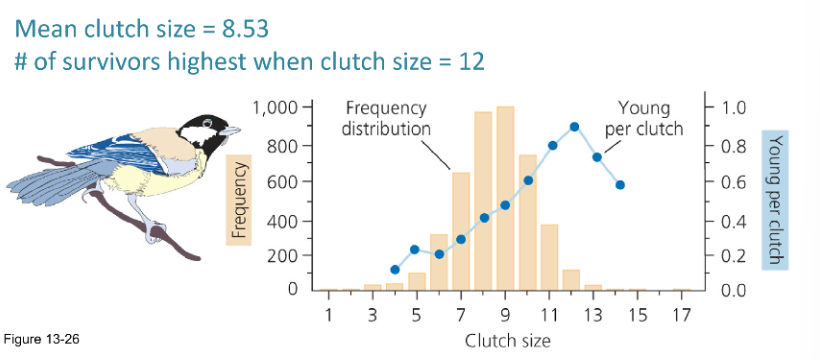
Selection doesn’t always favor the largest clutch size—it favors the size that results in the highest number of surviving offspring over time. A bird may lay fewer eggs if a larger clutch would reduce the survival or quality of the chicks, or compromise the parent's ability to reproduce in the future.
Selection does not always favor the clutch size that produces the most surviving offspring in a single year. Birds may lay fewer eggs because larger clutches can reduce offspring survival, lower offspring quality or future reproductive success, or increase costs to the parents that reduce their own survival and long-term fitness.
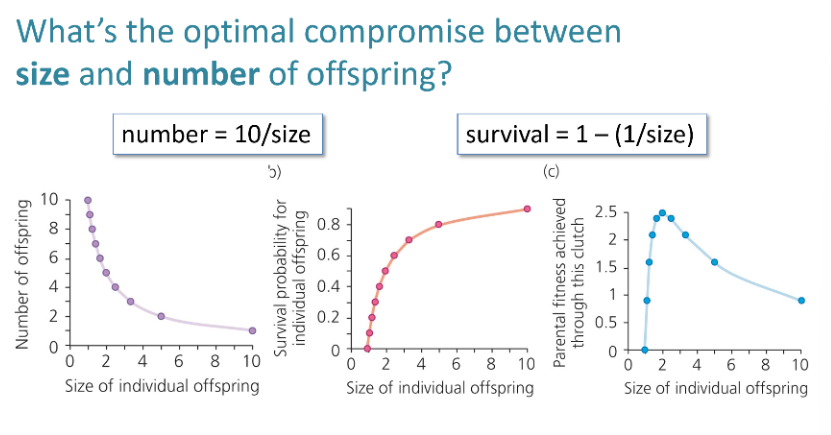
Know the example for Stator limbatus – why would females produce larger offspring on poorer hosts plants?
Female Stator limbatus beetles lay larger eggs on poorer host plants because larvae have a lower survival rate there, and larger eggs improve their chances. This plastic response is adaptive: while larger eggs reduce the total number laid, they significantly increase survival in tough environments, balancing the trade-off between offspring size and number.
Explain the concepts of genomic imprinting & when we would (and wouldn't) see that in an animal system
Genomic Imprinting: Alleles such as IGF-II result in embryos outcompeting other males’ embryos. Genomic imprinting occurs when male and female alleles contain distinct chemical markers and are transcribed differently.
Genomic imprinting is when a gene's expression depends on whether it is inherited from the mother or the father, due to chemical "marks" added during gamete formation. It often evolves in animals with placental development, like mammals, where paternal and maternal genes may have conflicting interests over maternal resource allocation to offspring. For example, paternal genes may push for more growth (e.g., IGF-II) to favor their own offspring, while maternal genes may limit this to conserve resources for future reproduction. Imprinting is expected in species with internal development and multiple paternity, but not in species where embryos are provisioned before fertilization or where all offspring share the same father.
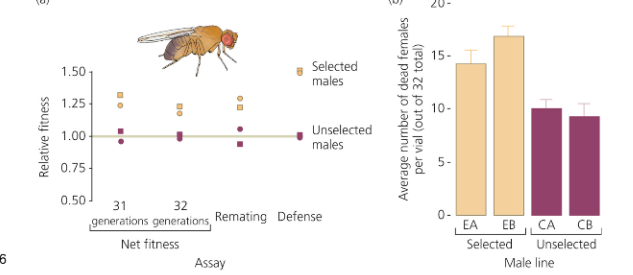
What is sexual coevolution? What happened in promiscuous Drosophila that were then made monogamous? Why?
Sexual Coevolution: Sexual selection may favor beneficial adaptations in one sex that are detrimental to the other sex.
Sexual coevolution is the evolutionary back-and-forth between male and female traits driven by conflicting reproductive interests. In promiscuous Drosophila, males evolved traits like toxic seminal fluid to boost their reproductive success, even though these traits harmed females by increasing mortality. When these flies were made monogamous for many generations, males evolved less harmful traits, and females lost resistance to those toxins—because both sexes now shared aligned fitness interests. This shift demonstrates how mating systems shape sexual conflict and coevolution.
Why is semelparity advantageous for B. schlosseri, and when/why is iteroparity advantageous for the same species?
In Botryllus schlosseri, semelparity is advantageous in spring and early summer when environmental conditions favor rapid growth and a single, large reproductive effort, maximizing fitness before potential threats arise. Iteroparity becomes advantageous in late summer when competition from Botryllus leachi increases, as iteroparous colonies are better able to survive and reproduce multiple times despite environmental stress. This seasonal variation in selective pressures maintains both life-history strategies in the population through genotype-by-environment interactions, preserving genetic variation despite strong natural selection.

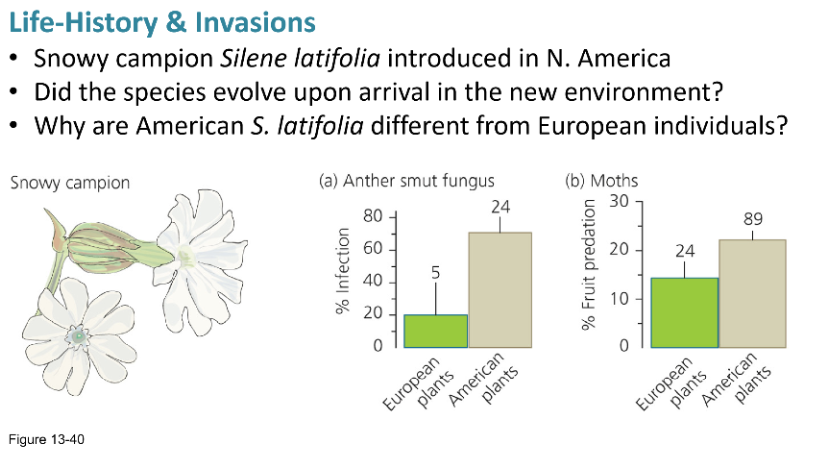
What happened to Silene latifolia when it arrived in N. America, and why didn’t that happen in Europe?
When Silene latifolia (snowy campion) arrived in North America, it escaped many of its natural enemies—such as a fungus that infects its anthers and seed-eating moths—that kept its population in check in Europe. Freed from these pressures, the plant evolved rapidly: North American individuals germinate earlier, grow faster, produce more flowers, and survive better than their European counterparts. This shift was due to a life-history trade-off, where the absence of enemies allowed the plant to invest less in defense and more in reproduction, boosting its fitness and enabling it to become a weed. In contrast, this did not happen in Europe because its natural enemies remained present, maintaining selective pressure for defense and preventing such a shift in resource allocation.
Sociality

Know some examples of the four basic kinds of social interaction, and be able to classify a described interaction as one of those 4 basic type
Mutually beneficial nesting in the greater ani bird Crotophaga major
Genetically unrelated adults nest together and all adults care for the eggs.
Communal breeding increase reproductive success - cooperative defense of eggs
Selfish Cannibalizing of eggs in cane toads
cane toad tadpoles eat the eggs of their conspecifics
tadpoles that eat eggs grow faster and survive at higher rates
can bait traps with bufogenins to attract invasive cane toad tadpoles
Altruism by alarm calls in ground squirrels
Classic example of mutualism is only mutualistic in some situations
whistle in response to air attacks
trill to alert for ground attacks
Spite by bacteria producing bacteriocins
A member of one species Xenorhabdus boveinii protects itself (individual), but kills members of its own species in the process.
Production of the bacteriocin also hinders growth of the producing individual.

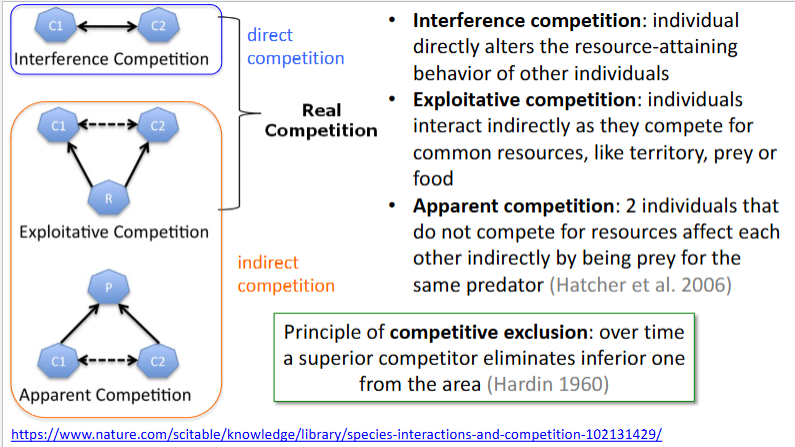
What is the Principle of Competitive Exclusion?
Intraspecific Competition - Competition between individuals of a single species, related to population density
• Competition for mates
• Competing for food and other resources
• Competing for space or defending a territory
Interspecific Competition - Competition between individuals of different species
• Competing for resources like food or light (both, if you’re a plant or coral!)
• Competing for space (e.g. anemone colonies on a rock, heads of coral, territorial animals, etc.)
Why was J.B.S. Haldane (supposedly) prepared to lay down his life for 8 cousins or 2 brothers?
J.B.S. Haldane famously quipped that he would be willing to lay down his life for “two brothers or eight cousins” to illustrate the concept of kin selection, a foundational idea in evolutionary biology.
According to Hamilton's Rule, an altruistic behavior (like sacrificing oneself) can evolve if the cost to the individual is outweighed by the benefit to the recipient, discounted by how closely related they are. The rule is often written as:
r × B > C
Where:
r is the coefficient of relatedness between the actor and the recipient,
B is the benefit to the recipient,
C is the cost to the actor.
In Haldane's example:
A brother shares 50% (r = 0.5) of his genes.
A cousin shares 12.5% (r = 0.125) of his genes.
So:
Saving 2 brothers gives a genetic payoff of 2 × 0.5 = 1.0
Saving 8 cousins gives 8 × 0.125 = 1.0
That equals the loss of his own 1.0, making the altruistic act "genetically" worthwhile in both cases. Haldane’s quote was a clever and memorable way to explain the mathematics behind kin selection.

How is kin selection dependent upon indirect fitness?

Examples of Kin Selection
Conflicts can arise from social behavior:
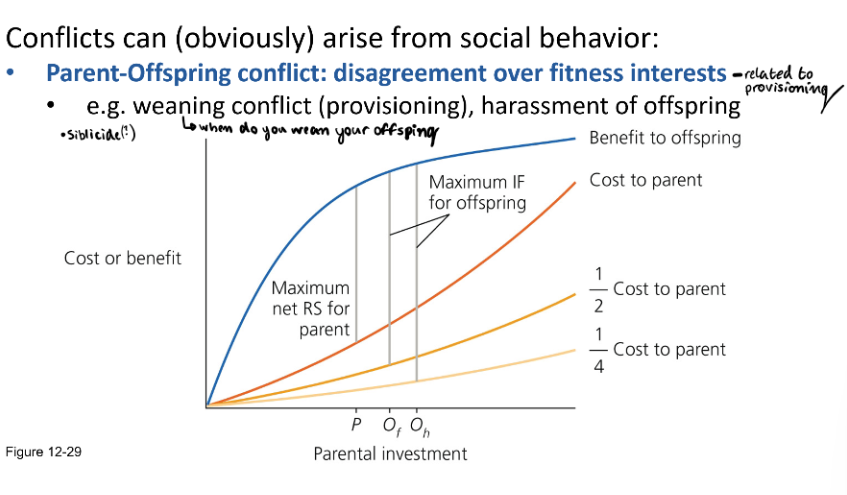
Parent-Offspring Conflict: disagreement over fitness interests - related to provisioning.
Weaning conflict (provisioning - who gets what), harassment of offspring.
Siblicide in some species of birds & mammals
Blue-footed boobies try to prevent siblicide and have more complex siblicide dynamics than masked boobies.
Clutch of ~ 2 chicks. BFB will help their siblings survive. BFB parents will save their chicks. MB wonn’t stop siblicide.
MB. Have older siblings. 10 days between births. Pushes younger sibling out of nest so that they can have all of the provisions.
Kin selection depends on indirect fitness because it favors behaviors that help relatives reproduce, thereby passing on shared genes. While direct fitness comes from an individual’s own offspring, indirect fitness comes from aiding relatives’ reproduction. Kin selection acts when the gain in indirect fitness outweighs the cost to direct fitness, increasing overall inclusive fitness.
Non-kin cooperation and Conflict
Reciprocity = exchange of favors that ultimately benefit both participants, e.g. baboons, humans.
Mean cooperation with money experiment. Kids in classes given 20 dollars. can give one up to give all 4 memebers of group 0.4 of a dollar. if everyone does this - end up with 36.
With punishment - donate more.
Without punishment - donate less.
Given a certain number of offspring for a recipient and a host of an altruistic behavior, as well as a measure of relatedness (r) be able to plug those in to the equation for Hamilton’s rule to determine if the frequency of the altruistic allele will increase
To determine whether an altruistic allele will increase in frequency, you can use Hamilton's Rule:
r × B > C
Br - C > 0
Where:
r = coefficient of relatedness between the actor (host) and recipient
B = benefit to the recipient (measured in extra offspring or fitness units)
C = cost to the actor (host) in the same units
Steps to Apply Hamilton's Rule:
Identify r: Use standard values:
Parent–child or full sibling: r = 0.5
Half-sibling, grandparent–grandchild, aunt/uncle–niece/nephew: r = 0.25
Cousin: r = 0.125
Plug in values:
Multiply the benefit B (extra offspring the recipient gains due to the altruistic act) by r
Compare the result to the cost C (offspring the host loses)
Interpret:
If r × B > C, the altruistic allele is favored by natural selection.
If r × B ≤ C, the allele will not increase in frequency.
Example:
An individual sacrifices having 1 offspring (C = 1) to help a full sibling produce 3 extra offspring (B = 3). Are they likely to increase the frequency of the altruistic allele?
r = 0.5 (full sibling)
B = 3
C = 1
Apply Hamilton's Rule:
0.5 × 3 = 1.5 > 1
✅ Altruism is favored, and the allele can increase in frequency.
Would you like to try a practice problem using this rule?
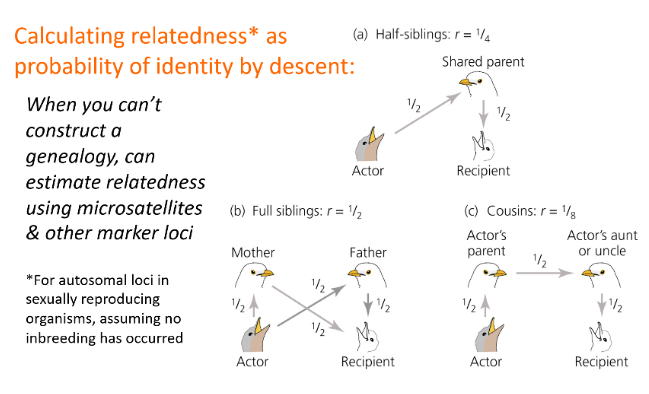
Calculating relatedness as probability of identity by descent
Know some examples of kin selection, and whether human beings practice kin selection
Ignoring spouses, who received 37% of the wealth left behind, the average will writer bequeathed
55.3% of his or her wealth to kin, versus
7.7% to non-kin
What’s a greenbeard allele? Can you give examples of greenbeard alleles and how they work?
Greenbeards - genes that can identify the presence of copies of themselves in other individuals, and cause their bearer to behave nepotistically toward those individuals
Dawkins imagined an allele that simultaneously causes its carriers to grow green beards, to recognize greenbeards on others, and to behave altruistically toward them. Carriers of such an allele would not have to distinguish full siblings versus half-siblings versus cousins and adjust their behavior accordingly. Under the greenbeard effect, alleles for altruism would not have to play the odds. Instead, they could bet on a sure winner.
Queller and colleagues studied a gene in the slime mold Dictyostelium discoideum called csA, which acts like a greenbeard allele. This gene produces a surface protein that causes amoebae to stick to others with the same protein, allowing them to recognize and preferentially associate with others carrying the same allele. In natural soil conditions, wild-type csA cells stick together more effectively, helping each other aggregate and form spores—thus showing discriminating altruism toward genetically similar individuals. This meets all three criteria of a greenbeard allele: it produces a trait, enables recognition of that trait in others, and leads to altruistic behavior toward those with the trait.
Define Competition, Mutualism, Commensalism, parasitism, predation, herbivory

Name some costs and benefits of being social
Costs of Being Social:
Increased competition for food, mates, or nesting sites
Higher risk of disease and parasite transmission
Greater visibility to predators
Risk of exploitation (e.g., cheating or freeloading within the group)
Benefits of Being Social:
Cooperative defense against predators (e.g., alarm calls, mobbing)
Improved foraging (e.g., shared information or group hunting)
Help with offspring care (e.g., communal nesting, babysitting)
Increased mating opportunities and access to mates
Thermoregulation (e.g., huddling for warmth)
Would you like examples of social animals that show these trade-offs?
Parasitism

Commensalism

Mutualism

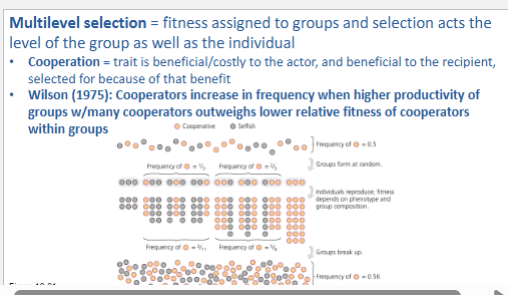
Understand multilevel selection and how it relates to human morality
Know the hypotheses of what traits facilitate eusociality (haplodiploidy & monogamy)
Eusociality - social system characterized by overlapping generations, cooperative brood care, and specialized reproductive & non-reproductive castes.
3 Hypotheses:
Haplodiploidy hypothesis: Haplodiploidy (males developing from unfertilized eggs, affecting relatedness) resulting in eusociality.
Monogamy hypothesis: Certainty of ½ relatedness leads to eusociality
Ecology & Life History Hypothesis: Complex brood care (nest building, care for larvae) leads to eusociality.
Association between nest building, larval care, and eusociality: hard for females to breed w/o help from a group, young dependent on parental care for long period of time.
What fossil evidence exists for sociality (and potentially eusociality) in hymenopterans?
Fossil evidence for sociality—and possibly eusociality—in hymenopterans comes primarily from amber-preserved fossils and trace evidence from the Cretaceous period (~100 million years ago). Some amber fossils show morphological caste differences, such as distinct worker and queen forms in ancient ants and bees. Additionally, groupings of individuals within the same amber piece suggest communal living, and fossilized nest structures indicate complex nesting behavior. These traits align with those James Hunt (1999) identified as key to eusocial evolution: complex nest building and extended care of larvae. While the behavioral details can’t be directly fossilized, these features strongly imply early sociality—and potentially the roots of eusociality—in ancient hymenopterans.

What’s coevolution? Explain this with respect to pollination biology and to malaria parasites
Coevolution: two taxa inluence each other’s evolution
Generally associated with extreme symbiotic relationships
Mutualism: Between pollinators and their plant symbionts.
Paarasitism: Between internal or external parasites and their hosts
Coevolution is the process by which two or more species reciprocally influence each other's evolution over time. Each species exerts selective pressure on the other, leading to adaptations that are often tightly linked.
Pollination Biology:
In pollination, coevolution occurs between flowering plants and their pollinators (like bees, birds, or moths). For example, plants may evolve nectar, bright colors, or specialized flower shapes to attract certain pollinators, while pollinators evolve long tongues, behavior patterns, or body structures to access and collect nectar efficiently. A classic example is the Madagascar star orchid and the hawk moth with an exceptionally long tongue—each evolved in response to the other.
Malaria Parasites:
In the case of malaria, coevolution occurs between Plasmodium parasites, mosquito vectors, and human hosts. The parasite evolves to avoid the human immune system and efficiently reproduce within both hosts. In response, humans have evolved genetic defenses such as the sickle-cell allele, which offers resistance to malaria. This arms race continues as the parasite evolves countermeasures to human defenses, and humans develop further resistance traits or medical interventions.