Neural Processing of Vision II : LGN to Visual Cortex
1/33
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
34 Terms
key 2 points to take away from the visual pathway
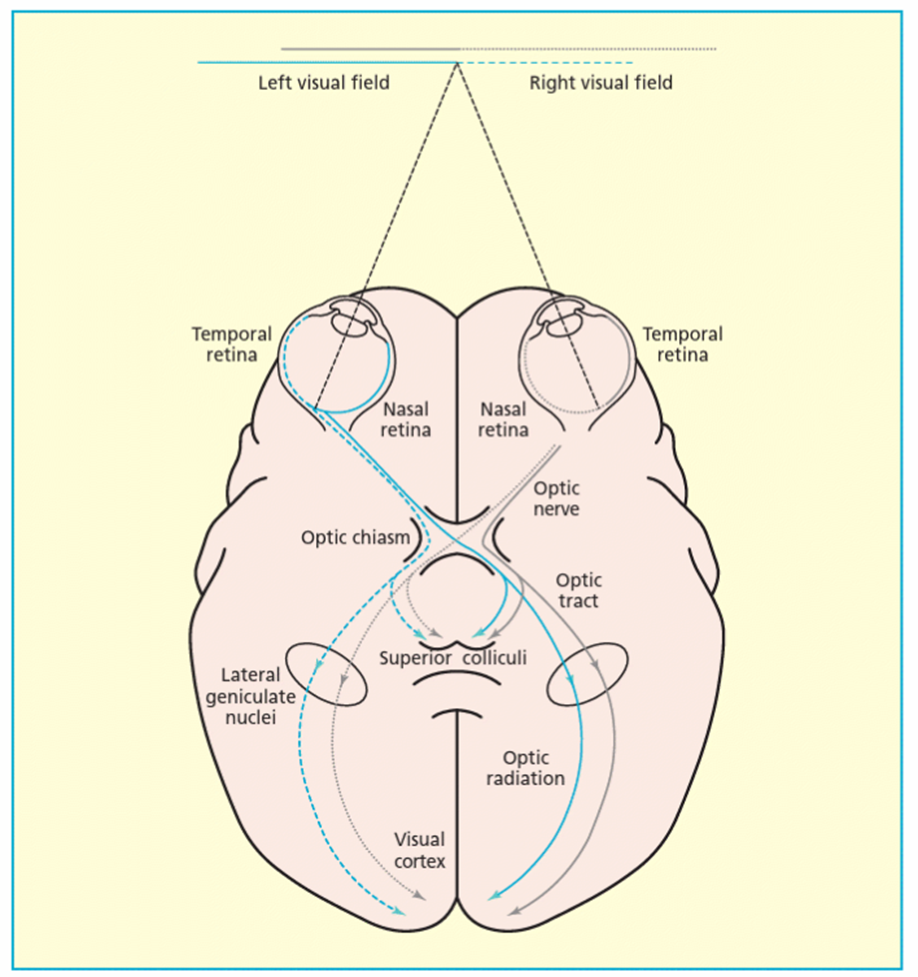
Information from right visual field goes to left visual cortex and vice versa
nasal fibers will cross but temporal fibers won’t in order for that to work ^^^
extra: Info that hits the temporal retina comes down optic nerve and stays on same side of optic chiasm – if info hit nasal retina those fibers go out optic nerve and optic chiasm and go to LGN and V1
sketch the visual pathway from retina to cortex
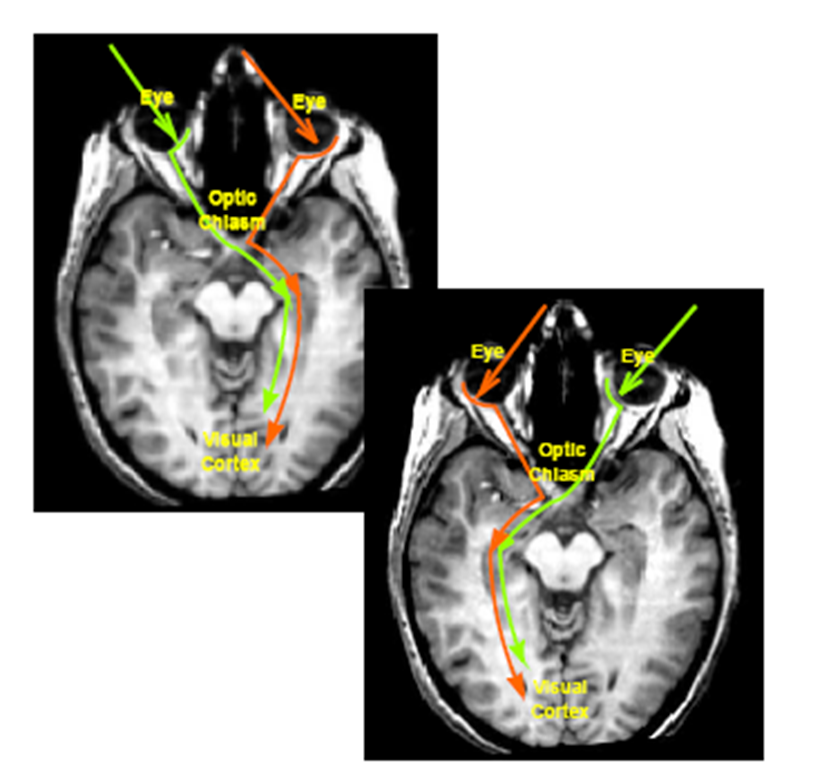

both eyes, the right SC, the right LGN, the right V1
just think of it being the opposite - so if information is coming from L VF then it will go the RIGHT and VV
where do the optic nerve fibers from the retinal ganglion cells go - visual pathway
Lateral geniculate nuclei (LGN) — 90% terminate here in macaque.
Superior colliculi— 10% terminate here in macque
some, but very few may also go to:
Suprachiasmatic nuclei—diurnal rhythm
Pretectum—pupil diameter
Pregeniculate—may be diurnal rhythm, unclear
Accessory optic system—maybe gaze stabilization, unclear
explain basic function of the superior colliculus
play a role in integrating sensory information (visual, auditory, somatosensory) into motor signals that help orient the head toward various stimuli and lead to action
Has vision, sound and touch – integrates across these 3
explain structure of the superior colliculus (4)
Layered structure (7 layers): top 3 layers have visual input, lower layers have auditory and somatosensory input
Visual input comes from retina AND V1, V2, V3, V4 and MT.
Cells are retinotopically organised (the brain keeps the same spatial layout as the retina so it can interpret visual scenes accurately, like a mental map of the image)
Visual cells in superficial/intermediate layers have ill-defined ON and OFF regions responding to just about any visual stimulus eg. bars of any orientation, spots, light flashes (so concerned not so much with “what” an object is, but “where” it is) - Not good at analyzing form but
good for knowing where something is
importance of the superior colliculus / role (2)
SC has an important role in visual orienting reflexes
SC is important for control of visually guided saccadic eye movements (frontal eye fields also important)
explain the basic structure of the LGN - layers
•The LGN has a very distinct, layered appearance (in humans it has 6 layers)
geniculate “with bent knee” – appearance
Layers 1 and 2 – larger cells – magnocellular layers
Layers 3,4,5,6 – smaller cells – parvocellular layers
Inter-layers – even smaller cells – koniocells
magnus “large”; parvus “small”; konis “dust”
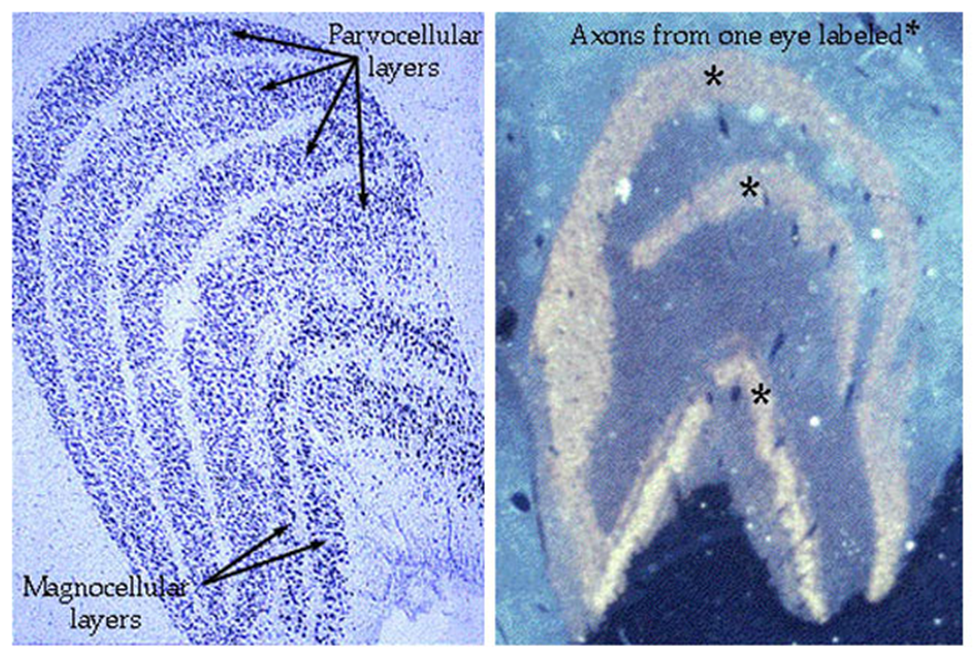
Info from 2 eyes is kept separate in distinct layers
Fibres strictly segregated at LGN
explain projection of rGCs to the LGN
P rGCs project to parvocellular layers of LGN
M rGCs project to magnocellular layers
K rGCs project to very small koniocells between the major layers
explain how the layers of the LGN are said to be “in register” / retinotopic organisation
each layer contains an orderly representation or map of the retina (a retinotopic map)
so that axons carrying information from adjacent parts of retina (visual field) connect to adjacent geniculate cells
explain difference and similarity between the LGN cells and rGCs in terms of receptive fields
Similar to rGCs, LGN cells have approximately circular receptive fields and centre and surround components interact antagonistically (ON- and OFF-centre cells)
surround of LGN receptive field exerts stronger inhibitory effect on centre than it does at the rGC level – highlights difference between light and dark slightly
Thus LGN cells serve to amplify differences in illumination i.e. contrast and edges
explain the differences between the cells in the layers of the LGN
Differences between cells in parvocellular and magnocellular layers are similar to those initiated in P cells and M cells of the retina.
Functions of Parvo and Magno cells clearly segregated at LGN, suggesting that they serve different roles in serving visual perception
describe the functional characteristics of the parvo cells in the LGN
Nearly all are differentially sensitive to colour of light imaged in RFs – they are “colour opponent” eg. +R/-G or +G/-R
RFs are 2-3 times smaller than magno cells at any particular eccentricity; important for analysing spatial detail
describe the functional characteristics of the magno cells in the LGN
Cells not differentially affected by colour; response depends on relative intensity of centre and surround.
Respond vigorously to fast abrupt fluctuations in light intensity within their RFs
larger RFs
describe the functional characteristics of the konio cells in the LGN
occupy interlayer regions of LGN
in LGN, they have relative large RFs compared to those for P cells and M cells (but very small RF centres)
they show selectivity for B versus Y, colour processing
there are afferents from SC and some respond to sound and touch as well as visual stimulation
what can receptive fields at the level of the LGN code for
can code for size AND colour
what is the function of the LGN
Pre process information before it is sent to the cortex – enables a more efficient onward journey to V1
But also a receiving area from V1 to let the LGN know when it is ready for more signals or not - visual cortical feedback accounts for 30% of synapses
modulate incoming information, integrating across large retinal areas and emphasizing sudden changes in stimulation
Important regulatory function - regulating strength of signals sent to cortex and pre-sorting information into significant streams
explain what the V1 one is - basic
Area V1 (striate cortex) is MAIN receiving area for LGN neurons
There are at least 30 distinct visual areas beyond V1 – called extra-striate cortex
describe the layers of V1 (4)
Anatomically split into 6 main layers (numbered 1-6), though layer 4 is subdivided into 4a, 4b and 4c
layer 4c further subdivided into 4ca and 4cb
Most of input arrives in layer 4c – magnocellular pathway terminates in 4ca; parvocellular pathway terminates in 4cb !!! - need to know this
K cells terminate in layers 2, 3 and 4a
explain retinotopic mapping/organisation in V1 compared to in other structures (LGN retina SC and V1 all retinotopically mapped) (3)
the layout of the retina is preserved in the primary visual cortex (V1) — like a visual "map" of what your eyes see
Each point on the retina (where light lands) connects to a specific point in V1
difference with V1 is cortical magnification - the central part of your vision (the fovea) is given much more space in the brain than the peripheral parts - central information is magnified - to allow for high-resolution vision
explain the main differences in the RFs at the level of V1 compared to LGN
RFs are NOT circular in V1 - elongated
can now code for orientation at V1 due to the elongation of the RFs
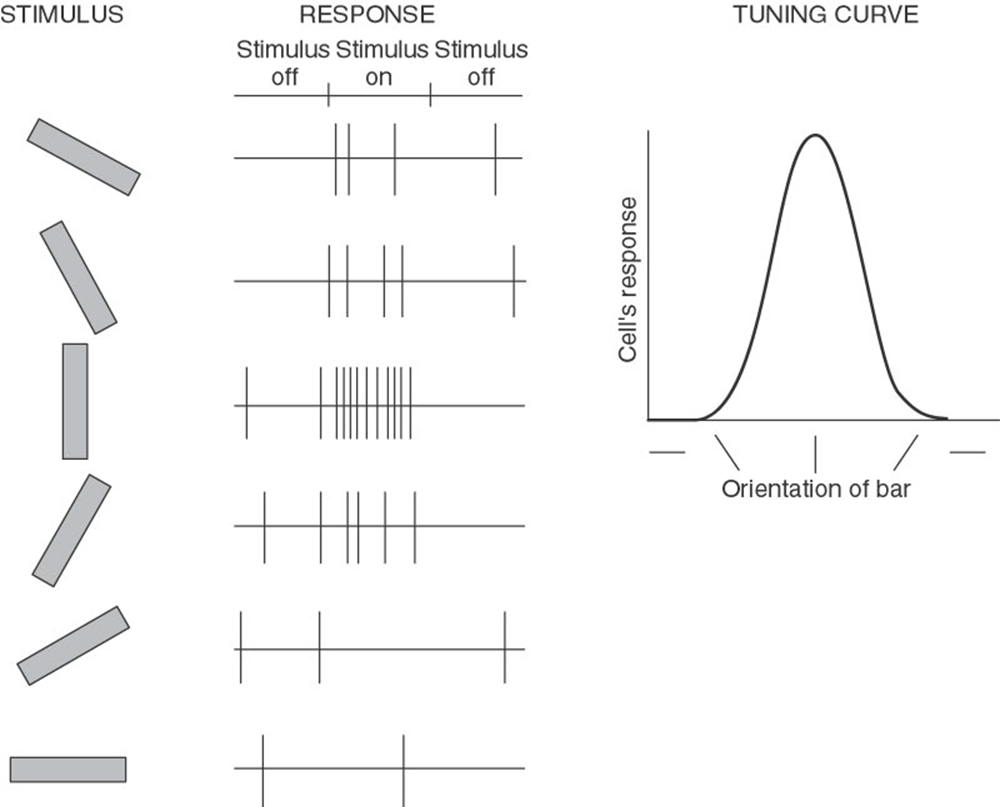
explain orientation selectivity in RFs in V1 (4)
Any particular cell will respond only if orientation of edge or line falls somewhere within rather narrow range – about 15 deg to either side of optimal
Cells in layer 4c, less orientation selective than those in other layers
Put stripes that match to its preferred orientation – fires a lot // If don’t – wont fire at all – if 90 degrees off
Orientation sensitivity is strong
what are receptive field profiles in V1 split into - 3
3 types of cells: simple cells, complex cells and hyper complex cells
explain characteristics of simple cells (5)
Have elongated/not circular RFs with elongated excitatory and inhibitory regions
Respond best to a) a bar or b) an edge of correct orientation and size in correct position on retina
Thus selective for orientation, size and position
Display little or no spontaneous discharge
well defined ON and OFF regions
explain characteristics of complex cells (5)
Have larger RFs and do not exhibit distinct ON and OFF regions - cannot be mapped out
Respond best to rapidly flickering or moving stimuli - Exhibit sensitivity to motion and direction
have a preferred orientation and size like simple cells, but are not so specific for stimulus position - Can respond to stimuli regardless of location within their receptive field
Respond well to edges and bars of light but are less selective for position
Because their responses are not simply given by linear summation of sensitivities of subregions, they are described as non-linear
explain the characteristics of hyper complex cells / end stopped cells
Responses depend not only on contour orientation but contour length as well - line also has to be a certain length for it to fire a lot
Maximum response is to bar whose length and width “fits” the RF – extending the bar length beyond this optimal value dramatically reduces cells response
End-stopping found amongst both simple and complex cells – though originally thought to be property only of hypercomplex cells
summary of simple, complex and hyper complex cells
Cell Type | Orientation Selective? | Position Sensitive? | Motion Sensitive? | End-Sensitive? |
|---|
Simple | ✅ Yes | ✅ Yes | ❌ Not much | ❌ No |
Complex | ✅ Yes | 🚫 Less | ✅ Often | ❌ No |
Hypercomplex | ✅ Yes | 🚫 Less | ✅ Often | ✅ Yes |
explain orientation selectivity in terms of the oblique effect ()
If you measure the orientation selectivity of every cell in v1 - MORE cells are biased to H and V orientations than obliques
= higher sensitivity for H and V than obliques
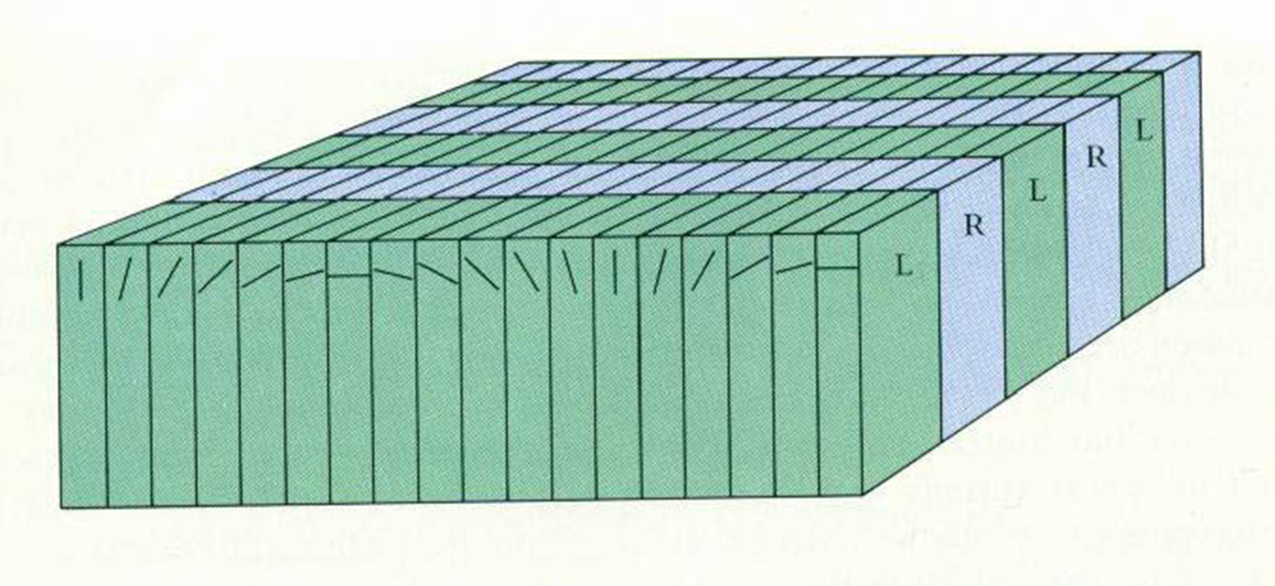
explain binocularity in cortical RFs - first time we see binocularity in the visual pathway
information comes to 4c separately from RE and LE layers from the LGN - once it leaves 4c ANY cell will respond to BOTH the RE AND LE
Once leave layer 4, ocular segregation gives way to binocular integration – with few exceptions, all cells outside layer 4 are binocularly driven
in total what can we code for at V1 - RFs
Coding for direction of motion develops in V1 also – by combining 2 cells
so we can calculate direction of motion – as well as colour and size and orientation
explain the organisation of V1
orientation: columnar organization of preferred stimulus orientation – all cortical cells in each column have similar preferred orientation – orientation column
ocular dominance: columnar organisation of preferred eye or ocular dominance columns (remember most cortical cells are driven by both eyes)
colour:
RFs BEYOND layer 4c can code for what…
colour, size, orientation, direction of motion and binocular vision
summary of visual pathway journey
rGCs synpase to LGN (10% to SC) = P to parvo, M to magno and K to konio – synapses coming back from VC TO LGN
LGn synpase to VC – V1
Describe what hypercolumns are
Full range of orientation - both eyes are represented- aggregation of adjacent columns