PKP Kap. 9
1/7
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai | Chat |
|---|
No analytics yet
Send a link to your students to track their progress
8 Terms
What role does biology play in personality psychology research?
Role: empirical foundation (≠ competing personality theory); provides scientific findings that every personality theory must explain; supports trait theories while expanding them through evolutionary psychology and neuroscience.
Core evidence: Phineas Gage → frontal cortex damage → intelligence/language/memory preserved; personality radically altered (responsible → impulsive, irresponsible) → specific brain systems contribute causally to personality.
Research focus: biological foundations of personality via: (1) Temperament, (2) Evolutionary psychology, (3) Behavioral genetics, (4) Mood, emotion & the brain, (5) Plasticity.
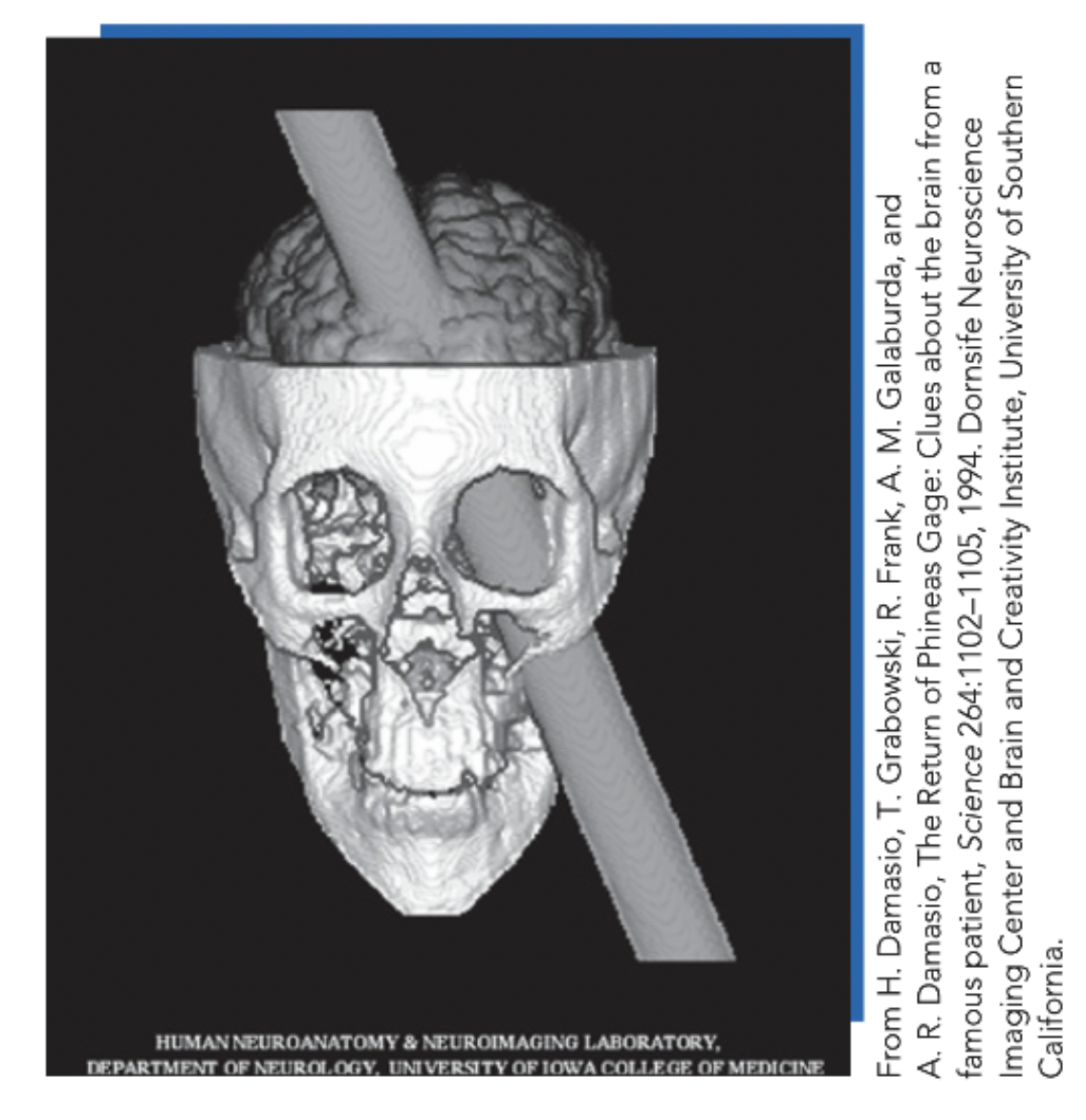
Describe temperament research.
Temperament: biologically based individual differences in emotional/motivational tendencies evident early in life (emotional reactivity; mood; arousal; self-soothing) → suggests biological origins before substantial social experience (but keep prenatal experience in mind for homosexual study!).
(A) Historical development: early taxonomy + causal explanations
Hippocrates/Kant: four temperaments explained by bodily fluids (rejected);
Gall: phrenology (correct: functional specialization; incorrect: isolated localization—brain functions via interconnected systems);
Darwin: evolution + emotional expression → foundation for evolutionary psychology;
Mendel: modern genetics;
Kretschmer/Sheldon: body type → temperament/personality (weak empirical support; methodological flaws);
Pavlov: nervous-system strength (maintaining functioning under high stimulation) + conditioned reflexes → precursor of biological stress reactivity.
(B) Modern longitudinal research:
Thomas & Chess (NYLS):
(1) easy (adaptable; positive); (2) difficult (negative; poorly adaptable); (3) slow-to-warm-up (low reactivity; cautious)
→ infant temperament predicts later adjustment (difficult → most problems; easy → fewest);
goodness-of-fit: optimal development depends on match between temperament and parenting (≠ universally optimal parenting).
Buss & Plomin:
inherited dimensions: (1) emotionality; (2) activity; (3) sociability → stable across development; substantial genetic influence (especially MZ twins);
limitation: parental-report bias (e.g., overestimating twin similarity). → Shift toward biologically grounded temperament dimensions; influenced later trait models (Big Five); lacked direct biological measures.
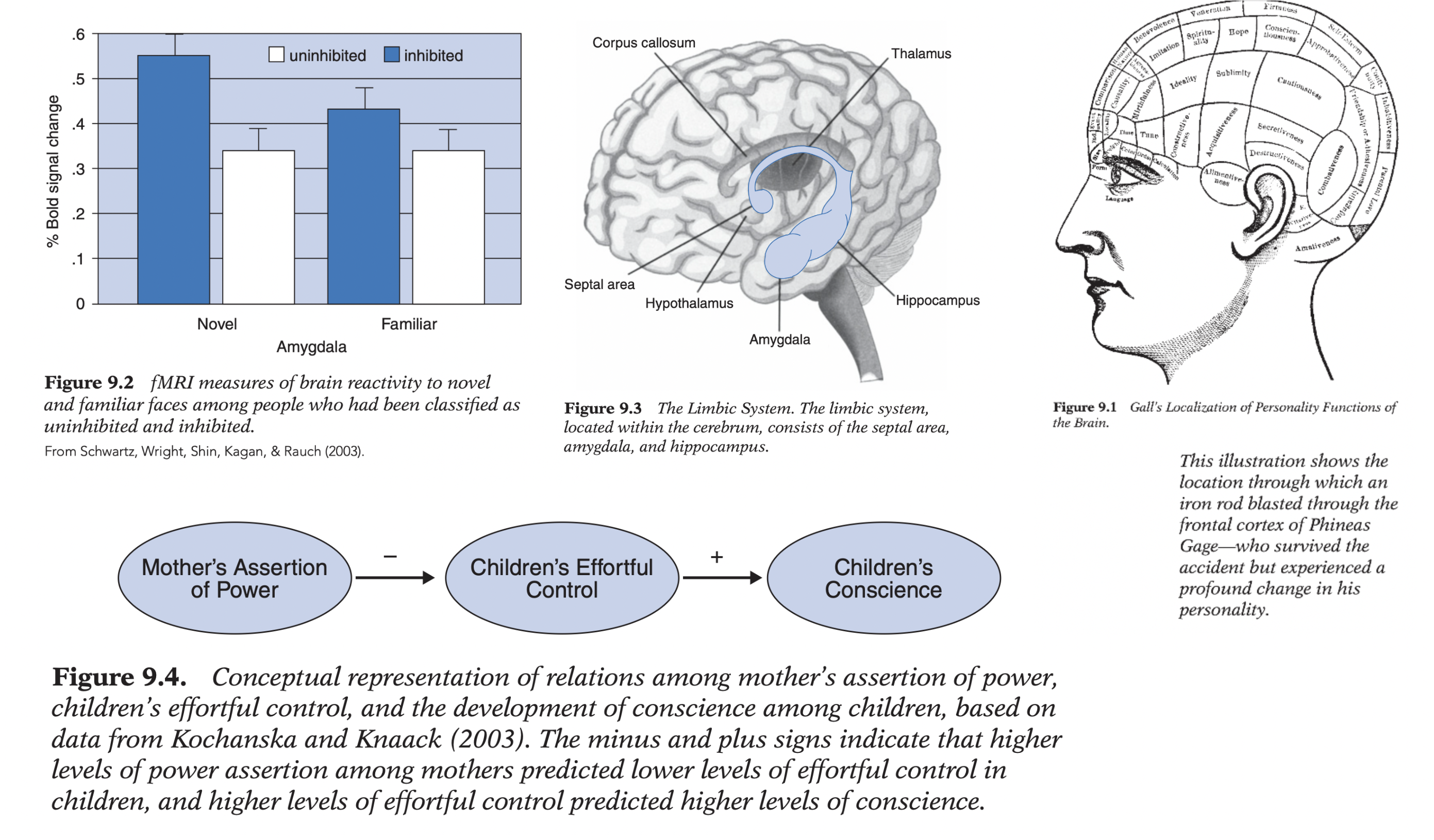
Kagan’s biological temperament model: objective laboratory observations (≠ parent reports);
(1) inhibited: fear; avoidance; restraint; physiological arousal to novelty vs. (2) uninhibited: curiosity; sociability; positive affect.
Hypothesis: high-reactive (!) infants → inhibited; low-reactive → uninhibited. 4-month testing: ~20% high-reactive; ~40% low-reactive; ~40% intermediate. Longitudinal follow-up (14 mo; 21 mo; 4.5 y; 8 y): substantial continuity (high-reactive → greater fear; HR/BP; less smiling/talking; low-reactive → confident/social).
Continuity ≠ determinism: (a) supportive, (b) non-overprotective parenting with (c) reasonable demands reduced fearful outcomes; BUT no high-reactive infant became consistently uninhibited; low-reactive → inhibited only rarely → inherited temperament biases developmental direction (predisposition ≠ destiny).
Biological mechanisms: high-reactive infants form distinct categorical group (~10% consistently inhibited; ≠ purely continuous dimension).
Amygdala: novelty/threat detection; frontal cortex: regulates amygdala/emotion. fMRI: inhibited at age 2 → greater amygdala activation to unfamiliar / novel (but not non-novel!) faces at ~20 y → long-term biological stability.
Animal knockout studies: stathmin gene influences amygdala functioning (gene present → greater fearfulness; absent → reduced fear). Limitations: amygdala ≠ fear center only (multiple functions); amygdala damage ≠ absence of emotion; novelty may activate amygdala more than fear; maternal social support moderates genetic prediction (high support weakens genetic risk) → biology provides predispositions expressed within developmental context; temperament retains built-in flexibility across life.
Effortful control & conscience:
Rothbart: effortful control = suppress dominant response to perform adaptive/subdominant response (delay gratification; inhibit impulses).
→ Kochanska: temperament × parenting → conscience (Freud’s superego analogue). Longitudinal behavioral measures (slow walking; whispering; delay candy) + observed parenting + later conscience tasks (cheating; obedient vs. naughty dolls).
Findings: authoritarian/high power assertion → ↓ effortful control; ↑ effortful control → ↑ later conscience; effortful control mediated parenting → conscience relation. → Moral development emerges from interaction of biologically based self-regulation and environmental experience (biology × parenting; mediation).
Describe the research in evolutionary psychology.
Evolutionary psychology: explains ultimate causes (why mechanisms evolved via natural selection for survival/reproductive success) ≠ proximate causes (current biological mechanisms producing behavior) // Main assumptions (Buss; Cosmides; Tooby; Pinker):
(1) Personality = evolved psychological mechanisms solving recurrent ancestral adaptive problems.
(2) Adaptations evolved in hunter-gatherer environments (modern mismatch possible; e.g., fat preference → obesity);
(3) Domain-specific mechanisms (specific problems ≠ general traits; e.g., jealousy ≠ childcare; fear of ancestral threats ≠ general fear)
(4) Mind = multiple specialized modules (social exchange; parenting; mating; exclusion…) ≠ single general processor. → Contrasts with trait theory (specialized adaptive mechanisms ≠ broad context-free traits).
Social exchange & cheating detection (Cosmides): reciprocal exchange repeatedly essential → cheating detection adaptive. Abstract logical reasoning (“if P then Q”) → poor performance; identical logic framed as social contract/cheating (e.g., taxes) → high accuracy. Replicated across literate/non-literate cultures → evidence for universal evolved module. Neuropsychology: frontal cortex/amygdala damage → normal general reasoning; impaired social-contract reasoning → specialized cognitive system for cooperative exchange.
Sex differences: Parental Investment Theory (Trivers): women invest more biologically (pregnancy; limited fertility; childcare); men face paternity uncertainty.
(1) Predictions: men prefer youth; physical attractiveness; chastity (reproductive value; paternity certainty); women prefer earning capacity; ambition; industriousness; protection/resources. Buss:>10,000 participants; 37 samples; 33 countries + islands;
→ broad support (men consistently valued youth/attractiveness; chastity only moderate support; women valued financial resources/ambition).
(2) Jealousy hypothesis: men → sexual infidelity (paternity threat); women → emotional infidelity (resource threat).
→ questionnaires supported predicted distress; physiological measures replicated; strongest effect for men with prior committed relationships (women: nonsignificant).
Evaluation/Criticism: universal evolutionary mechanisms predict cross-cultural consistency;
Eagly & Wood: sex differences smaller in gender-equal societies (women ↓ emphasis on earning capacity; men ↓ emphasis on domestic skills) → supports biosocial model (biology × social roles) ≠ biological determinism.
Miller et al.: men and women showed highly similar overall mate preferences.
DeSteno; Harris: jealousy effects largely disappeared when replacing forced-choice methods (MCQ) with separate ratings/real experiences; both sexes responded more strongly to sexual infidelity; men generally reacted more strongly to sexual content BUUT regardless of infidelity (!)
→ Evolution contributes to behavior, but evidence for universal, hardwired sex differences is mixed/inconsistent; biology and social context jointly shape behavior.

Describe behavioral genetics research.
Behavioral genetics: investigates genetic contributions to individual differences (genes influence personality INDIRECTLY via biological development; ≠ genes for Extraversion/Neuroticism) // Methods:
Selective breeding: experimentally breed animals for traits (e.g., fearfulness; alcohol responsiveness) → demonstrates genetic effects; manipulate environments experimentally (not possible in humans).
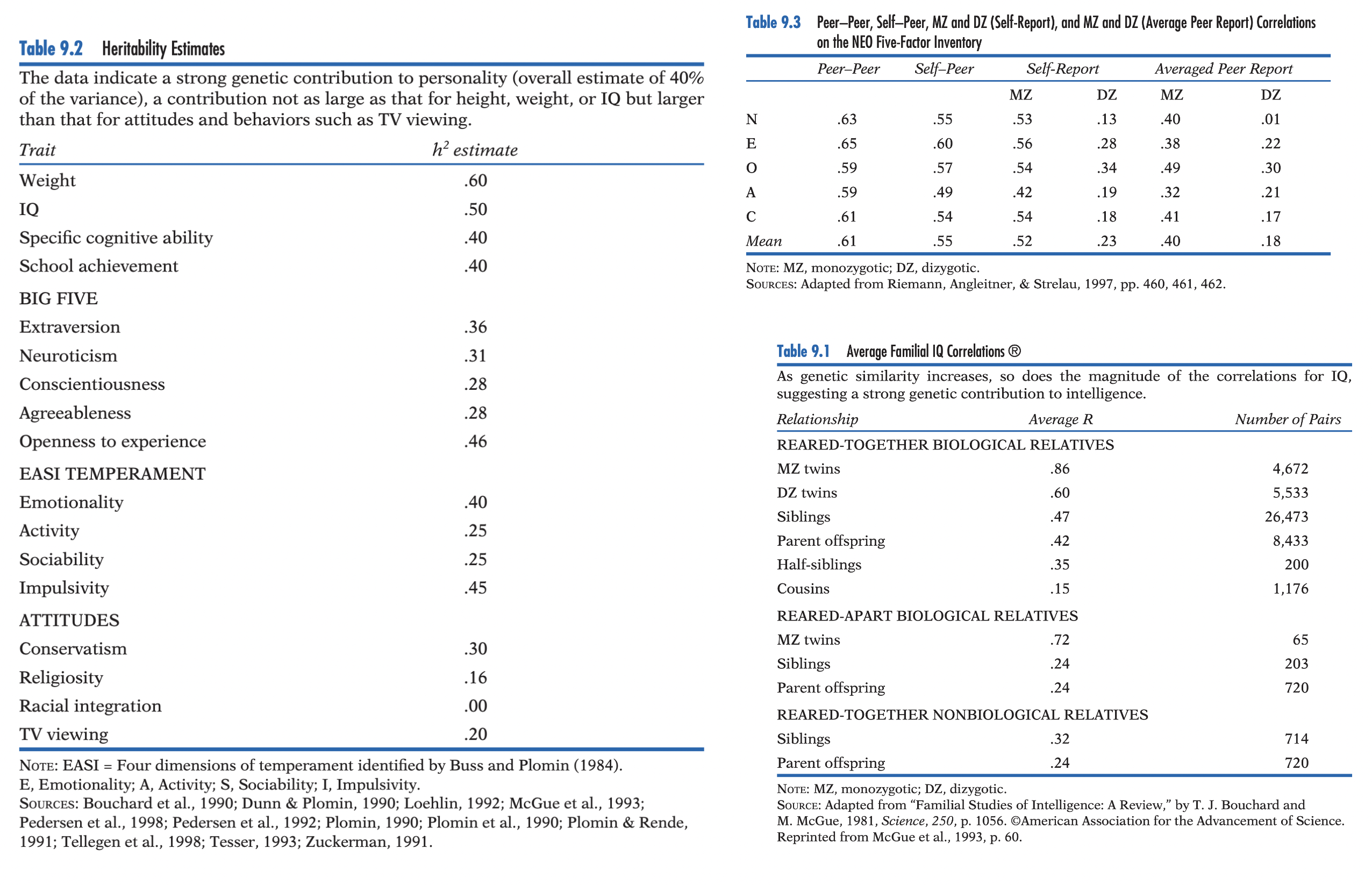
Twin studies: MZ (~100% shared genes) vs. DZ (~50%; ordinary siblings); MZ > DZ similarity → genetic influence; MZ reared apart → isolates environmental effects. MZ reared apart: r ≈ .45–.50; nearly identical to MZ reared together → substantial genetic contribution.
Adoption studies: compare adopted children with biological vs. adoptive relatives → biological resemblance = genetic effects; adoptive resemblance = environmental effects.
Heritability (h²): proportion of population variance attributable to genetic differences (remainder = environmental variance + residuals); population statistic (varies across traits/populations/environments; e.g., war ↓ h² by increasing environmental variation or different for different populations!); ≠ percentage of an individual’s trait caused by genes; h² = 0 possible despite genetic necessity (e.g., universal abilities with no person-to-person variation).
Findings: personality moderately heritable (~40–50%); genetic influences across virtually all traits (Big Five; IQ; emotionality; activity; sociability; impulsivity; attitudes such as conservatism). Self-report limitation addressed by peer ratings → independent peer reports replicated Big Five genetic findings (slightly lower genetic estimates) → greater validity/generalizability across measures.
(!) Caveats: (1) heritability ≠ immutability (environment can modify highly heritable traits); (2) heritability = nomothetic (population differences) ≠ idiographic (individual prediction); (3) genetics operate via gene × environment interactions, not genetic determinism; e.g. Molecular genetics: identifies specific genes underlying outcomes (≠ estimating overall h²).
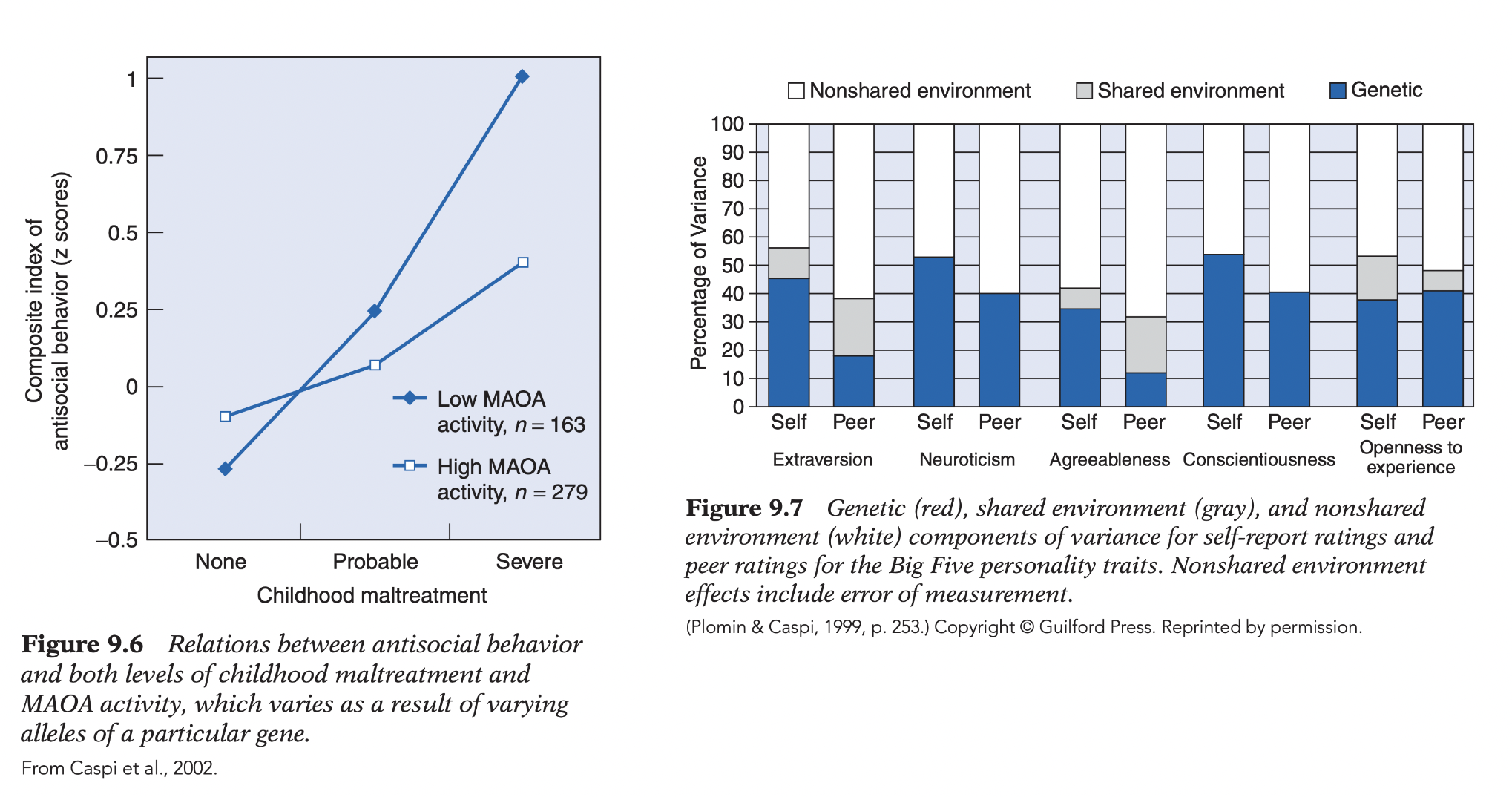
MAOA study: high-activity allele buffered childhood maltreatment → lower antisocial behavior than low-activity allele.
Serotonin transporter study: low-serotonin variant + many stressful life events → higher depression risk; gene alone insufficient.
→ Both demonstrate gene × environment interaction.
Gene–environment interaction: genes and environment = inseparable partners (≠ nature vs. nurture).
Cooper & Zubek: maze-bright/maze-dull rats × enriched/impoverished environments → enrichment benefited genetically dull rats; deprivation mainly harmed genetically bright rats → same environment produces different outcomes depending on genotype.
Environmental influences: shared environment (makes siblings similar) vs. nonshared environment (makes siblings different).
Findings: shared family environment contributes little to personality; ~40% of variance reflects nonshared environment. Big Five: substantial heritability across all traits (Openness independent of IQ); multiple measures separated measurement error from nonshared environment; peer ratings largely replicated self-reports
Reiss: nonshared effects partly arise because genetically different children evoke different parental treatment.
Three gene–environment interactions:
Passive: same environment → different effects depending on genotype.
Evocative: genetically influenced traits evoke different responses from others.
Active: individuals increasingly select/create environments matching genetic predispositions.
Describe the research on brain systems.
Focus: identifies neural/biochemical mechanisms underlying personality (emotion; temperament; self; morality); biology = dynamic system (≠ fixed cause only).
(!) Hemispheric asymmetry (Davidson): (L) left prefrontal → positive affect; enthusiasm; approach motivation; (R) right prefrontal → negative affect; distress; avoidance.
EEG: greater (L) left baseline activation → stronger positive emotions to pleasant films; greater (R) right activation → stronger negative emotions to unpleasant films.
Clinical evidence: depression → ↓ (L) left-anterior activity; left frontal damage → depression; (R) right frontal damage → mania;
distressed infants (maternal separation) → greater (R) right activation.
Worry ≠ anxious arousal: worry shows relatively greater (L) left-frontal activation → distinct neural mechanisms (≠ single Neuroticism factor).
Anger: despite negative valence → (L) left activation (approach/confrontation) → hemispheric asymmetry reflects approach vs. avoidance, not simply positive vs. negative emotion.
Neurotransmitters & temperament:
(1) Dopamine: reward/reinforcement; learning; pleasure; approach; positive affect; energy; impulsivity; incentive motivation; excess → schizophrenia; deficit → Parkinson’s disease.
(2) Serotonin: mood regulation; emotional stability; impulsivity; aggression. Low serotonin → depression; anxiety; OCD; alcoholism; impulsive aggression; pessimism. SSRIs prolong serotonin action.
(3) Cortisol: stress hormone; adaptive acutely; chronic elevation → depression; memory impairment; inhibited children (Kagan) ↑ cortisol at age 5 (effect absent age 7).
(4) Testosterone: dominance; competitiveness; aggression // high testosterone promotes competitive/aggressive behaviour, while winning competitions (even coin tosses) increases testosterone and losing decreases it → bidirectional cause–effect relationship.
Clark & Watson’s 3-factor temperament model → No one-to-one neurotransmitter–trait mapping; each transmitter influences multiple traits; personality reflects interacting biological systems. (A) Differentiation/localization: specialized brain regions vs.(B) organization/system: personality emerges from coordinated interaction of interconnected systems:
(A) PE (Positive Emotionality): Extraversion-like; energetic engagement; dopamine; left hemispheric activation.
(B) NE (Negative Emotionality): Neuroticism-like; threat sensitivity; emotional instability; low serotonin; right hemispheric activation; heightened amygdala responsiveness.
(C) DvC (Disinhibition vs. Constraint): self-regulation (≠ affective valence); high = impulsive; sensation-seeking; reward-driven; low = cautious; controlled; future-oriented; serotonin (primary) + dopamine + testosterone.
Plasticity: biology both cause and effect of experience.
Monkeys: dominance ↑ serotonin; loss ↓ serotonin.
Testosterone: promotes competition and increases after winning (even coin tosses), decreases after losing.
Juggling study: 3 months learning → ↑ gray matter in motion-processing regions → adult brain structurally plastic.
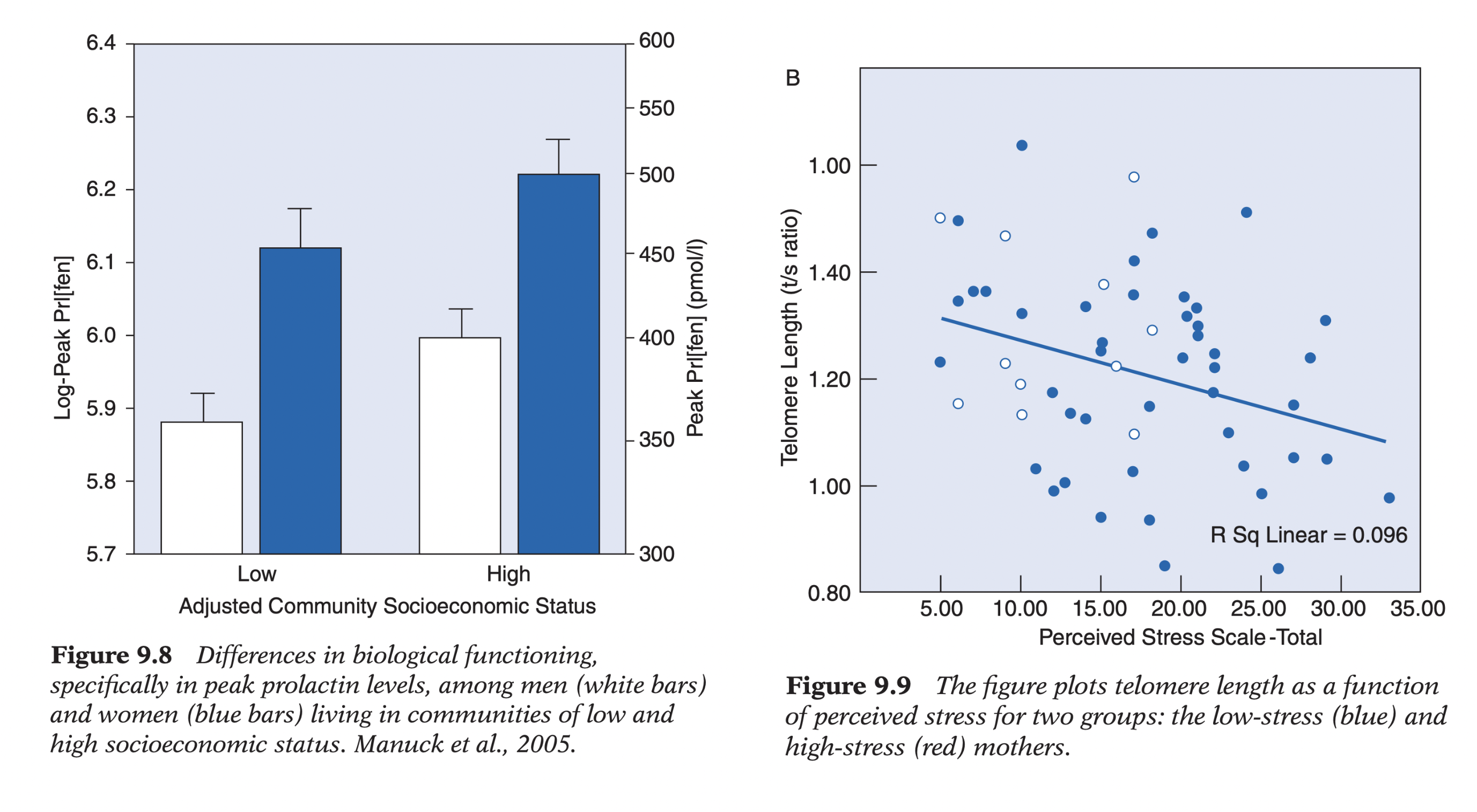
SES study: lower socioeconomic communities → reduced serotonergic responsiveness (serotonin agonist; prolactin measured), independent of IQ/Big Five → environment alters neurobiology.
Higher-level functions:
Self: fMRI; self-judgments (vs. Bush vs. perceptual judgments) → selective medial prefrontal cortex (mPFC; vgl. Rogers) activation → self-referential processing recruits specialized neural systems.
Morality: moral dilemmas activate emotion-related brain regions more than nonmoral reasoning → moral judgment integrates emotion + cognition (≠ purely rational).
2 Interesting studies:
Stress & telomeres (Epel): telomeres = chromosome end caps shortening with cell division (biological aging marker). Higher perceived + objective stress (especially mothers of chronically ill children vs. normal/healthy ones) → significantly shorter telomeres (~9–17 years greater biological age) → chronic stress modifies cellular biology; strong evidence for biological plasticity.
Prenatal environment: probability of male homosexuality ↑ with number of older biological brothers, independent of being raised together (!) → shared family environment excluded. Maternal immune hypothesis: successive male pregnancies trigger maternal immune response → altered prenatal environment → fetal brain development. → Prenatal biology contributes alongside genes and postnatal environment.
Tree (PKP Ch. 9) 1/2

Tree (PKP Ch. 9) 2/2

Tree (PKP Ch. 9) 3/3