Ecology Unit 1
1/113
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced |
|---|
No study sessions yet.
114 Terms
Ecology
the scientific study of the relationships between organisms and their environment
Distribution
the geographic and ecological range of a population
Dispersion
the spacing of individuals with respect to one another
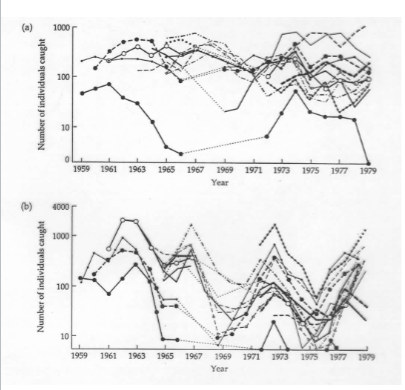
Mark-recapture methods
• Individuals (numbering M) are captured, marked, and released.
• We assume that they mix freely and completely with the rest of the population.
• Then we capture some individuals again
• Count the total number of individuals caught (n) and the number of those that were marked previously (x).
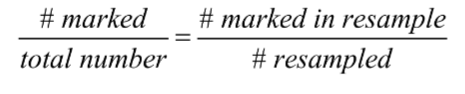
What problems exist with this mark-recapture method?
It made certain (poor) assumptions about:
-- distribution
-- dispersion
-- better with more marked individuals and larger re-sample
demographic transition
On August 20th, 1897, Sir Ronald Ross, while working as a military physician in India, demonstrated the malarial oocysts in the gut tissue of female Anopheles mosquito, thus proving the fact that Anopheline mosquitoes were the vectors for malaria.
\
**Ross also tried to predict the spread of malaria** in human populations. He created two linked equations, one each for infected humans (H) and for infected mosquitoes (M).
dH/dt = # new human infections - # recoveries or deaths
dM/dt = # new mosquito infections - # mosquito deaths
(Note the dN/dt = births – deaths form!)
\
b= per-capita birth rate
the probability of an individual having offspring during some time period
bN
Population birth rate
We can just multiple each of these per-capita rates by the populations size (N) to get a rate for whole populations
We can just multiple each of these per-capita rates by the populations size (N) to get a rate for whole populations
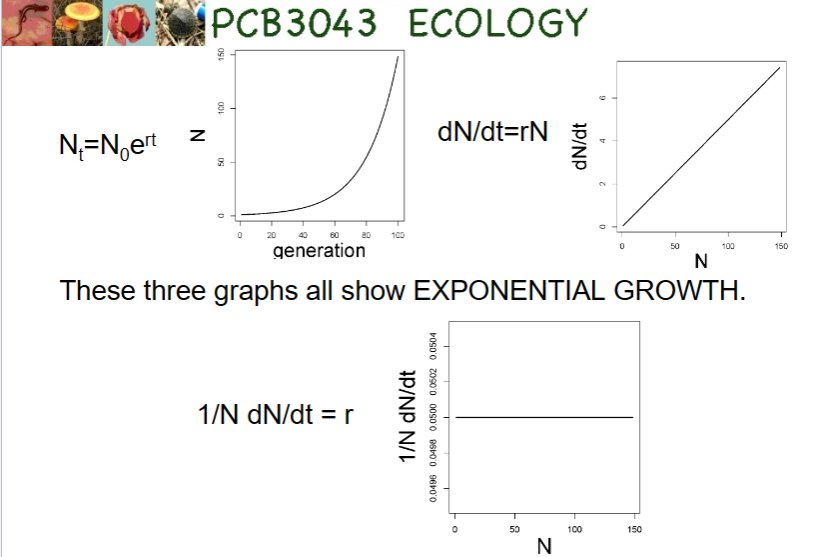
dN/dt = bN – dN \n (note confusing use of “d” for two different things here!)
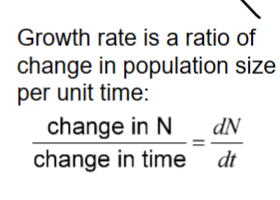
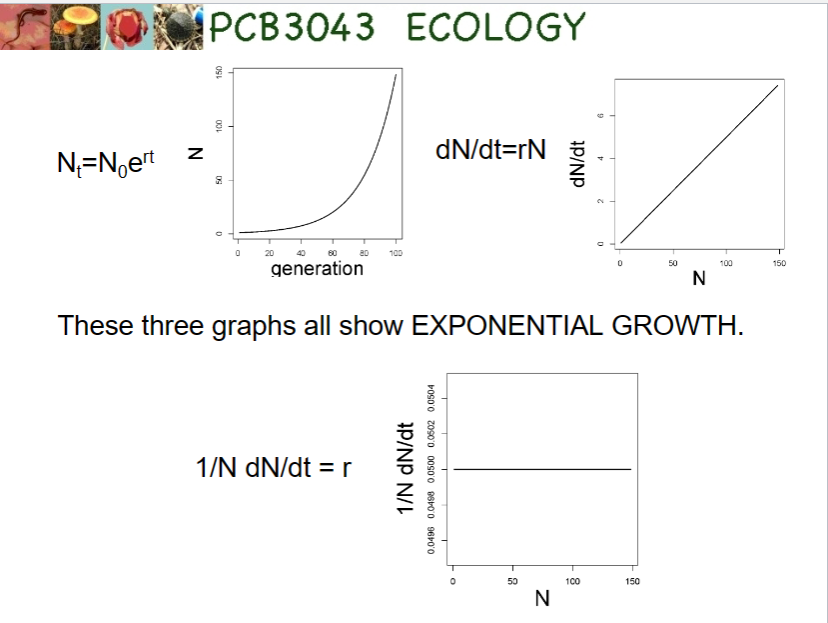
“little r”
r = births – deaths + immigration – emigration
per-capita growth rate
the intrinsic rate of increase
we get exponential growth with continuous growth if r remains constant and positive
closed population
If there is no movement of individuals between different populations, (no immigration or emigration) then the numbers of individuals are determined by local processes (that is, just births and deaths)
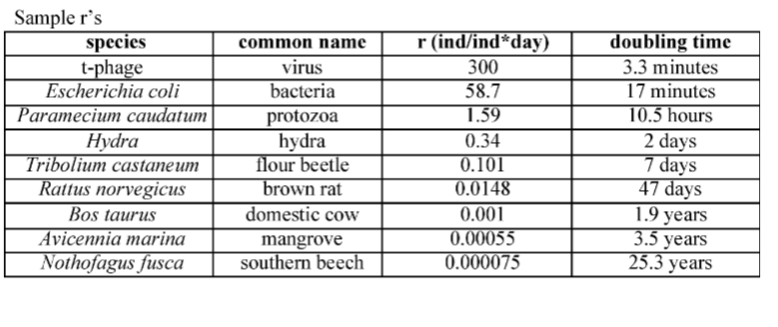
THE MAJOR PRINCIPLES OF POPULATION ECOLOGY
(1) Populations have the potential to grow rapidly.
(2) They usually don’t. Populations must be regulated such that rapid growth is a transient, but common phenomenon
\
His worked showed the two laws of population growth
Nt+1 = Nt x R0
Note that since it is multiplicative, if R0 = 1, the population is stable
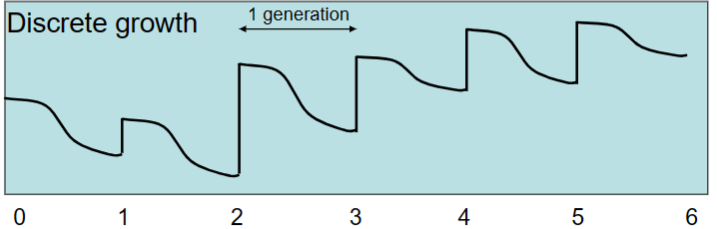
Continuous growth
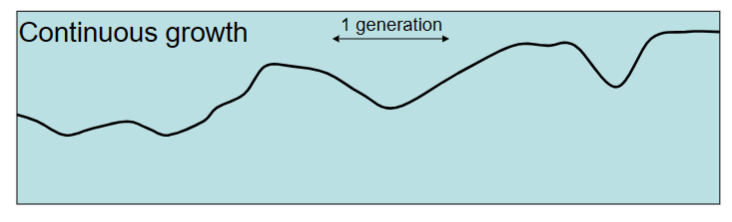
reproduction is not tied to seasons and can go on at any time. Individuals have might live just as long as before, but they can mate, make babies, and die all year round
R0
R0 (“R sub O”) is another growth rate, if R sub 0 is constant, exponential growth.
If R sub 0 is 1, population is stable
Discrete exponential population growth: Nt = N0 x R0t
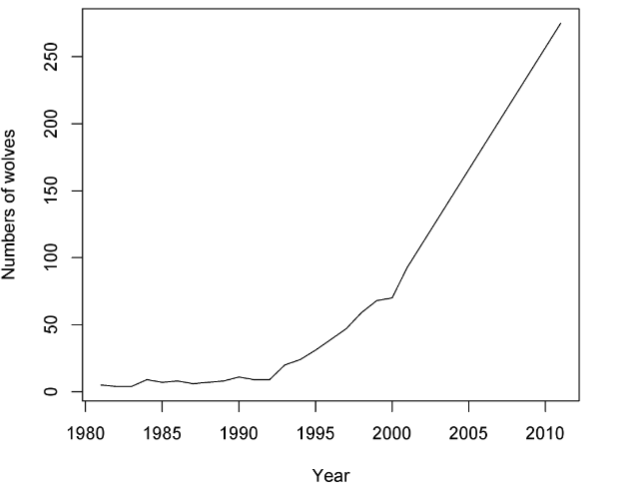
Case Study: growth in Scandanavian wolf populations (and bottleneck)
• A previously unknown pack of <10 wolves was discovered in 1983
• Without any external changes in management, the population began increasing in 1991 by 29% annually
• Genetic data suggest there were only two founding wolves, probably from Russia or Finland (not a relic population)
Case Study: human population growth (including Von Foerster doomsday prediction)
Until the last 20 years or so, it looked exponential, with no end in sight
Von Foerster thought the population would go to infinity on Nov 13 2026
He determined that the population growth rate was actually greater than exponential -- r is increasing
Case Study: Malthus studied growth in England (density dependent) and the US (more density independent)
Malthus thought that higher growth rates in the U.S. were due to better living conditions, he also thought that England’s fewer resources caused a lower growth rate and less morals
As temp increases, little r increases
K has highest value at **intermediate** temp
competition
a reduction in resource acquisition rate (feeding, foraging, parasitism, nutrient uptake) due to the action or presence of another individual that seeks to acquire the same resources
allee effects
mechanisms arise from cooperation or facilitation among individuals in the species, such that they do a bit better in groups.
Examples of such cooperative behaviors include better mate finding, environmental conditioning, and group defense against predators
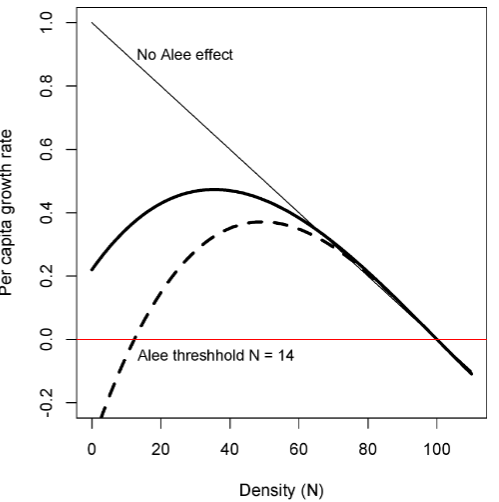
intraspecific competition
competition within species
interspecific competition
competition between different species
K
carrying capacity
minimum viable population size
Smallest population that can be predicted to have some probability (e.g., 95%) chance of persisting for some number of years (e.g., 100 or 1000 years)
environmental stochasticity
this when there are random environmental fluctuations affect the average birth and/or death rates, in turn causing variation in the population growth rate
demographic stochasticity
variation in population growth rates arising from differences among individuals in survival and reproduction
time lags
occur when population growth rates are determined not by the density at the present time, but by the density at some other time.
One example is gestation time – when times are good, animals may mate, but the effects on density may not occur until the offspring are born and achieve some size
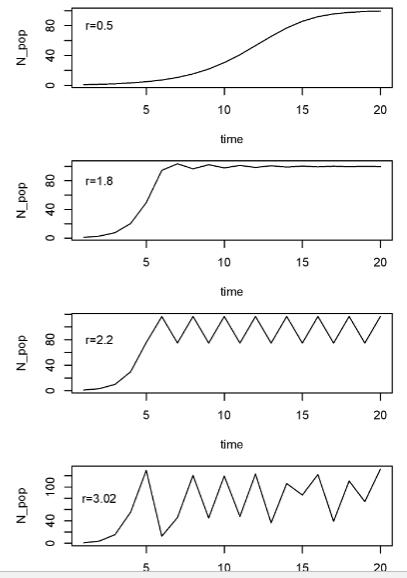
chaos
time lags can cause oscillations. As the lag or r increases, the population will first have damped oscillations, then fixed oscillations, then 2-point oscillations, then
CHAOS
not random
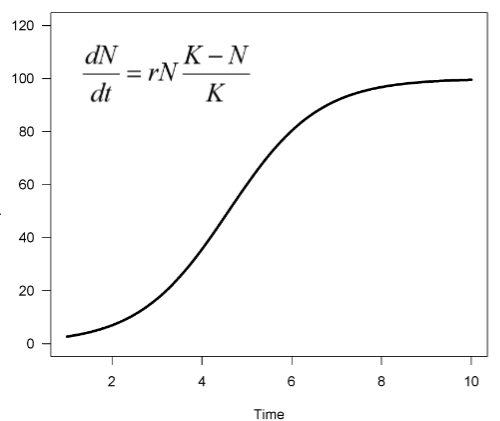
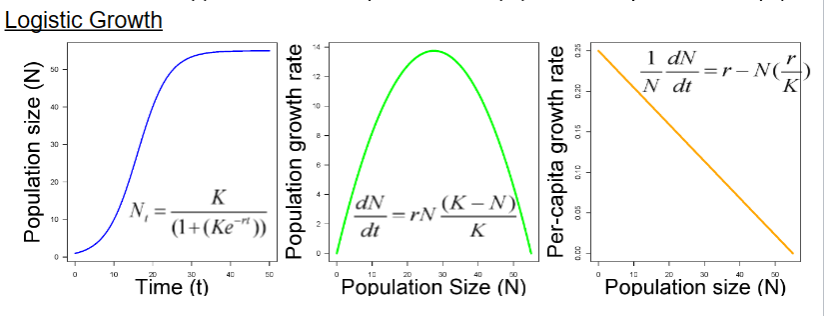
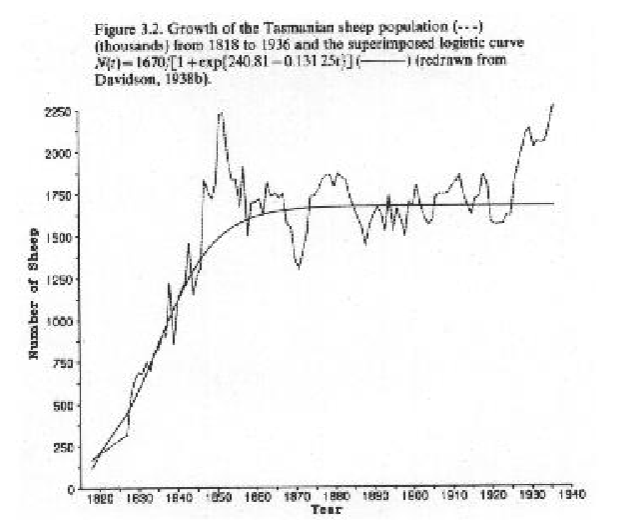
Logistic growth does occur in nature
example of barnacles and sheep
Assumptions of logistic growth (constant r and K)
In real life:
The carrying capacity is not constant (environmental stochasticity). Birth and death rates (or r and R) do not change in a linear way with density
Most populations are affected by both
density-dependent and density-independent
Logistic growth in Drosophila
Example of how genetics or environment can affect competition , r and the carrying capacity (K).
MVP example from Bighorn Sheep
Populations with fewer than 50 individuals went extinct within 50 years
Lionfish and density independence
Benkwitt transplanted lionfish onto 6 reefs at 6 different densities. I also established and maintained 4 reefs with 0 lionfish as controls. All treatments were started within a 2-week period.
Individual growth in length declined linearly with increasing lionfish density, while growth in mass declined exponentially with increasing density. There was no evidence, however, for density-dependence in recruitment, immigration, or loss (mortality plus emigration) of invasive lionfish. The observed density-dependent growth rates may have implications for which native species are susceptible to lionfish predation, as the size and type of prey that lionfish consume is directly related to their body size.
density vague
suggests that there is some average population level around which a regulated population fluctuates. As the populations is perturbed further from this average level, density becomes more important in “pushing” the population back towards the average.
How much it fluctuates depends on the relative importance of density dependent and independent factors.
density vagueness concept
It reveals that ALL
natural populations are really controlled by both density
dependent and density independent factors
lx or survivorship
survivorship from the beginning number
AKA #survived/# we started of with
cohort life table
are constructed by following an even age group (such as all the eggs in nests from one year) throughout their entire life. At any point in time, they are all the same age
show the probability of death of people from a given cohort
px or age specific survival
tells us which portion survives in the next time period
Density dependence vs. density independence is resolved by density vague
ALL natural populations are really controlled by both density-dependent and density independent factors
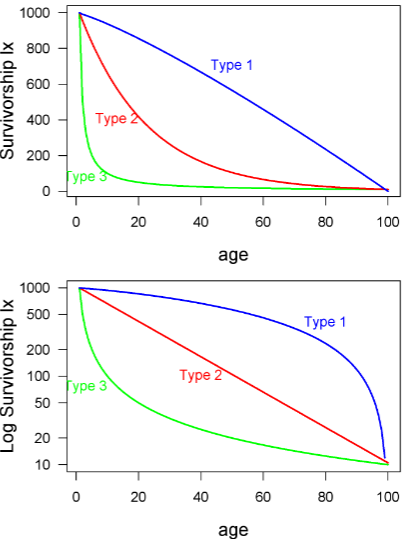
Y-axis for survivorship curves
the y axis may be plotted in several different ways. For example, the y axis may be actual numbers of individuals or proportions (this won’t affect the shape of the curve). Or, even more confusing, the y axis may be either log transformed or on a log scale -- this does change the shape!
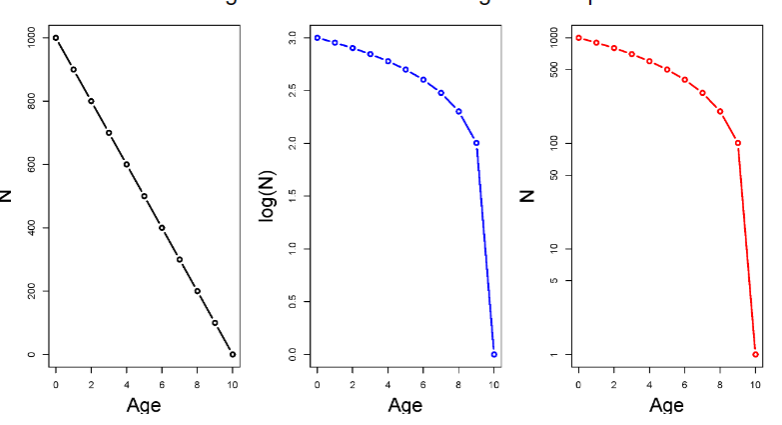
Sex-specific differences in survival
Females tend to have higher mortality during
childbirth years. However, females tend to live longer
Eisenberg’s snails
He conducted a simple experiment in 1966 to see if snails in a pond exhibited density dependent growth and to determine what factors might limit the snail population growth (e.g., competition or predation
food didn’t affect lx but did affect bx
Erickson’s dinosaurs
show type I survivorship curve
Type I survivorship
exhibit high juvenile survival and are common for mammals
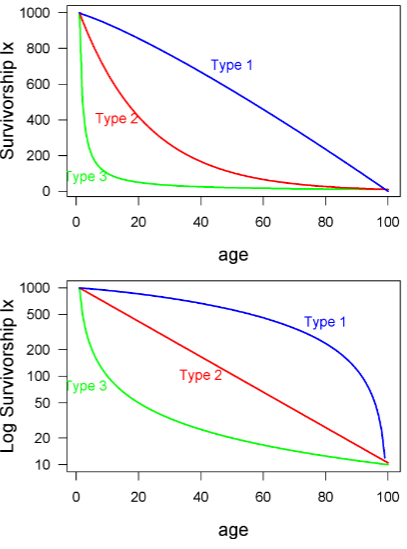
Type 2 survivorship
is constant probability of death (or survival) across age classes.
ex: birds
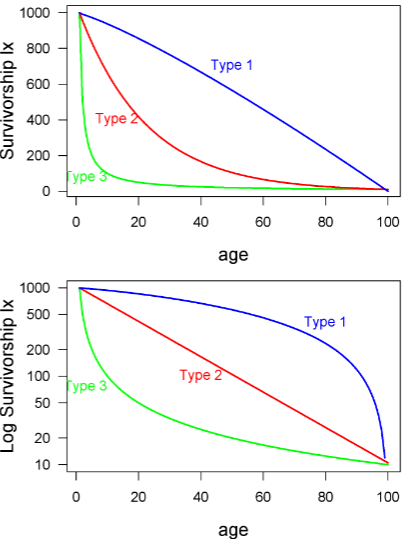
Type 3 survivorship
at younger age, higher chance of dying
ex: fish, insects
mx
average births per individual age x
lxmx
The lxmx for each age group tells us the reproductive contribution of each age group, weighted by the probably of surviving to reproduce
R0
sum of lxmx
If greater than 1= growing
If less than 1= decreasing
generation time
= mean age of mothers at the time of their mean daughter’s birth
= summation xlxmx / R0
stable age distribution
using R0 assumes that the population has achieved a stable age distribution.
Another way of determining R0
the sum of lxmx
Life table vs. population projection
Life tables do not have densities or abundances (N’s)!
However, we can now take this life table and predict what will happen to any given population through time. This is a population projection.
sensitivity
the slopes of these lines (in sensitivity analyses) are a measure of the sensitivity of the population growth to values in the life table
sensitivity analyses
This is used to see which birth or death rates most affect the overall population growth rate
How you get the stable age distribution and how it relates to R0
stable age distribution: the proportion of the population occupied by each age class remains constant. This pattern is seen in populations with a declining birth rate and a low death rate.
if either r or R0 is constant, then you have exponential growth IF you are at the stable age distribution
Remember how r and R0 differ
Increasing; r is constant and positive; R0 is greater than one
Decreasing: r is negative; R0 is less than one
Converting between r and R0
r = ln(R0)/G
R0 = e^rG
Emperor geese, sensitivity analyses
We need R0 greater than 1. So, the only way we could get that would be to increase juvenile survival or to increase adult egg production
R0 is most sensitive to changes in juvenile survival
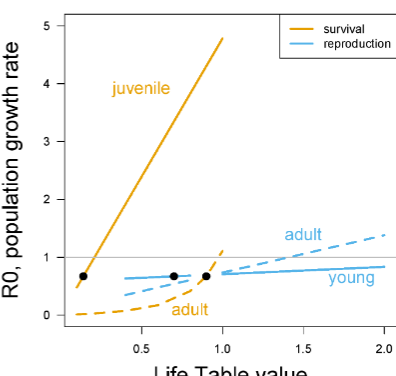
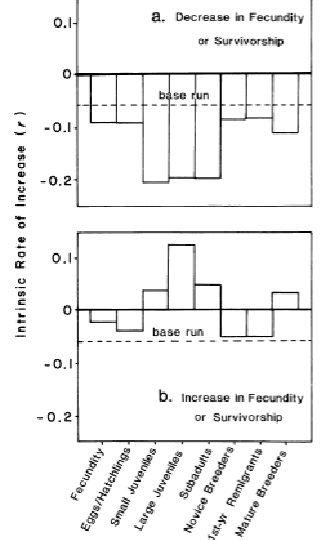
Sea turtles, sensitivity analyses
Looking at the bottom graph, the best thing is to save large juveniles
sink population
a net import of individuals (immigration > emigration)
source population
a net export of individuals (emigration > immigration)
rescue effect
The populations in some patches are maintained only through continuous migration, otherwise the population would go extinct. In effect, these sink populations are being continuously “rescued” from extinction
Regional population
a group of local populations
metapopulation
is a group of several local populations that are linked by immigration and extinction
Local population
make up a metapopulation
Going from studying population numbers to numbers of populations
instead of births and deaths, we focus on colonization of new patches and extinctions of patches
When do you have a metapopulation
at intermediate dispersal among patches)
Probabilities of patch colonization and extinction in b-d form
dP/dt= cp(1-P)-Ep
this equation states the growth tare of local populations can be determined by colonization’s and extinctions
Local vs. regional population persistence with number of local populations
the probability of regional persistence for one year is (1 - the probability that both patches go extinct during the year)
Persistence of x patches = 1 – (e)^x
Potential role of corridors
a way to link local populations
Hanski’s butterflies in dry meadows on islands
Some of the first work on metapopulations was by Ilka Hanski, who studied butterfly populations on dry meadows on islands near Finland
Populations in these patches aren’t static or unchanging. Instead, some populations may increase in size, while others decline. Some may go extinct, while others are being colonized
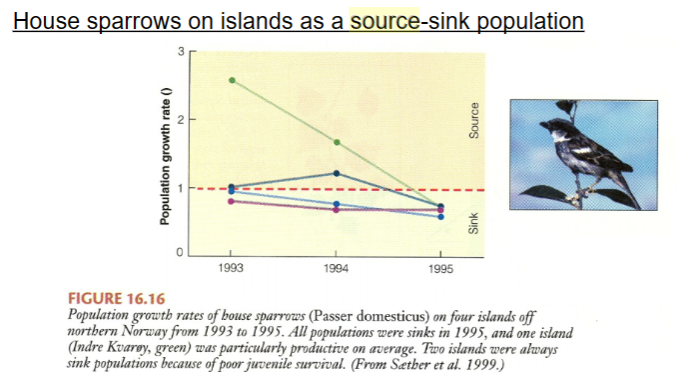
Sparrows on four island demonstrating sources and sinks
Sparrow populations on four islands off the coast of Norway, with migration among the islands. All islands maintained relatively constant number across the three years of this study, with migration from source populations “rescuing” sinks. The graph shows that some islands are more productive that others, acting as sources in good years. But, in severe years, all populations are sinks
Beetles in the Netherlands showing rescue effects
For the beetle Pterostichus versicolor, the abundances vary asynchronously and there were no extinctions over the 21 years.
For the beetle, Calathus melanocephalus, populations varied somewhat synchronously, leading to occasional local extinctions.