bio 211, lecture 13 - cell junctions and matrix
1/80
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced |
|---|
No study sessions yet.
81 Terms
clefting
smooth clump of epithelia, then involutes and forms wedges that split the epithelia into many distinct clumps
steps for the process of forming a neural tube
fertilization of the oocyte
many rounds of cell division without any growth
gastrulation
hollow ball of cells
epithelia sculpts
sides close to form epithelial tube
convergent extension
happens when cells reorganize so that they shrink (converge) in one axis and elongate (extend) in another axis
cadherin receptors
in the PM; bind to other cadherins in neighboring cells
mechanical stress
transmitted from cell to cell by cytoskeletal filaments anchored to cell-matrix and cell-cell adhesion sites
ECM
directly bears mechanical stresses of tension and compression
cell matrix adhesions
found at the bottom of the epithelial cells; connect to actomyosin filaments
desmosomes and hemidesmosomes
plug into IFs
provide structural strength to the tissues
not a big role in generating and transmitting forces
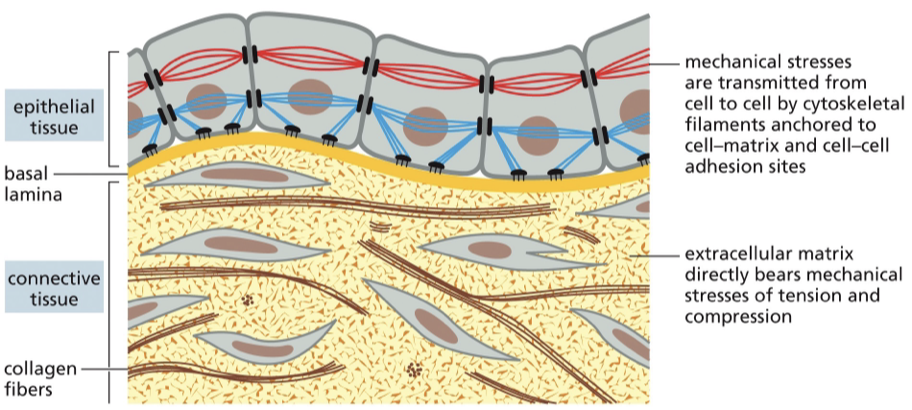
basement membrane / basal lamina
epithelial cells sit on this
function is to provide structural support for the tissue
provide physical barrier to separate outside from inside
what are the 2 ways to attach to the basement membrane?
actin-linked cell matrix adhesions
hemidesmosomes
actin-linked cell matrix adhesions
integrin-containing complexes that plug into talin and vinculin
transmit forces from actomyosin network to outside, and vice versa
hemidesmosomes
also attached to ECM at basement membrane
filament system they’re plugged into is IFs
job is to provide structural strength
gap junctions
do not give structural strength
put the cytoplasm of adjacent cells into communication with each other
have gap of sufficient diameter that macromolecules can freely flow from one cell to another
relationship between calcium and gap junctions
Ca concentration in the cytoplasm is increased, and activates Ca-dependent signaling events
gap junctions allow Ca to flow into adjacent cells
desmosome
attached to IF on the inside of the cell
on the outside of the cell, they form junctions on neighboring cells
give structural strength to the tissue
mediate strong adhesion
what is the difference between desmosomes and hemidesmosomes?
desmosomes are junctions involved in intracellular adhesion of epithelial cells
hemidesmosomes are junctions involved in adhesion of epithelia to basement membranes
tight junctions
connect the cells so tightly that not even ions can flow past
prevent the flow if big and small molecules across epithelial layers
tight junctions between the cells
only way for nutrients or other molecules to get across an epithelial layer and reach the bloodstream is to go through the cell
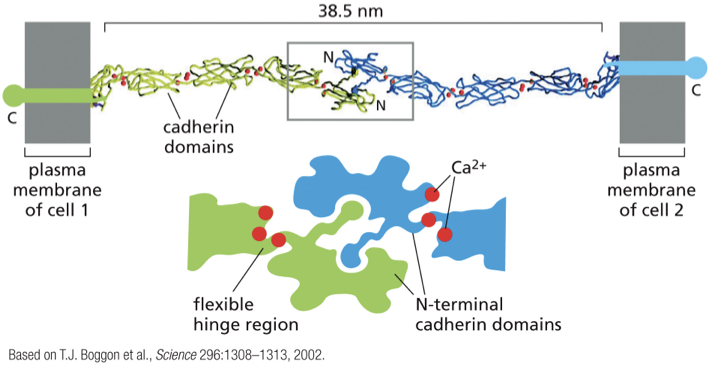
homophillic binding
cadherins bind to each other
Ca2+ ions are required in the extracellular space for homophilic binding to occur
E-cadherin
transmembrane glycoprotein which connects epithelial cells together at adherens junctions
P-cadherin
cell adhesion protein that helps maintain tissue structure and polarity
N-cadherin
transmembrane protein that helps cells stick together in a calcium-dependent way
how is calcium involved in making connections?
outside the cell, there is lots of Ca in the environment
Ca binds to cadherins at the hinge regions that occur between cadherin domains
this makes the protein more rigid and less flexible (rigidity is important in forming stable connections to neighboring cadherins)
what happens if you get rid of calcium?
the cadherin connections cannot form, and existing connections rapidly fall apart
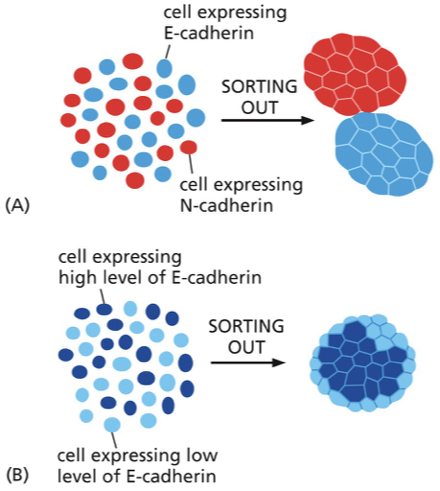
“sorting out” — different cadherins
mixed population of cells containing different types of cad
after a few days, they will have formed distinct clumps where it is all one cad in one clump and another cad in the other clump
why does sorting out occur? (different cadherins)
as cells bump into another one with the same cad, it becomes a firm connection, and they don’t let go (essentially an irreversible connection)
if they bump into a cell with a mismatched cad, they can let go and keep searching
“sorting out” — different levels of cad
cells with different amounts of cad are able to interact with each other, but the cells expressing more cad can form more connections with each other, so they get sorted out
why do the cells with different amount of cad get sorted out?
to have the most surface area in contact with the high-expressing cells (more connections this way and more stable arrangement)
cells with less cad will be kept to the perimeter because they form less junctions this way
cells with less cad are (more/less) adhesive
less
what is the overall purpose of sorting out?
allow for a higher degree of organization within the developing epithelial tissue
neural tube formation in later stages
e-cad is enriched at the surface epithelia and down-regulated as the neural tube begins to form
neural tube expresses many cads due to “sorting out”
cells that express the same cads form these clumps within the tissue — segregated from other epithelial cells within this neural tube
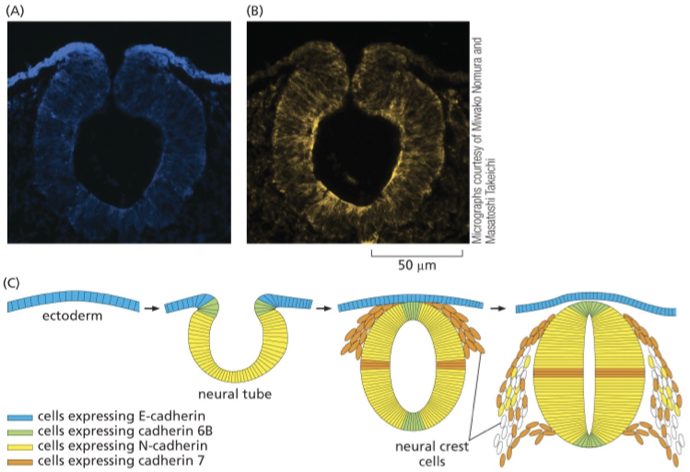
what happens if cad expression is messed up?
organization is lost
p120-cat and b-cat
bind the cytoplasmic tail of cad and help traffic the receptor to the PM and stabilize the initial homophilic binding
p120-cat
helps cad get expressed at the PM
b-cat
helps connect the a-cat to the cad
considered an adaptor (one side connected to cad and the other side connected to a-cat)
a-cat
provides the bridge from the cad receptor to the actomyosin filaments
binds to b-cat and connects e-cad to actin filaments
vinculin
brings additional actomyosin filaments to the complex following the application of force to the complex
applying force from outside the cell
pulling on a neighboring cell and causing tension on the cad
applying force from inside the cell
activate RhoA to activate contractility then tug on the cad form inside the cell
applying force from inside the cell or outside the cell triggers…
a positive feedback loop of force which leads to growth and strengthening of the cad junctions
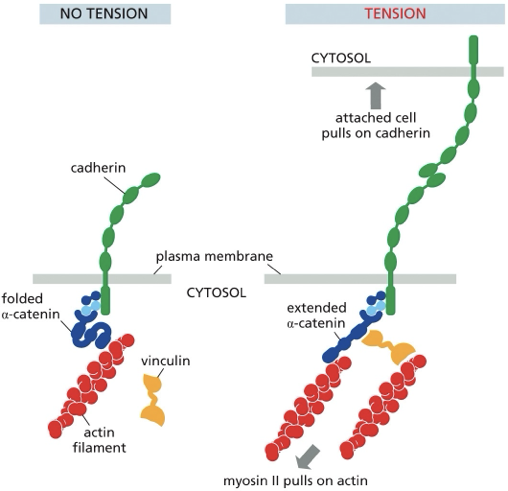
what does a-cat do in response to force?
stretches
reveals new vinculin binding sites that were previously hidden
allows actomyosin fivers to get plugged into complex and more force to be applied
step 1 of cell-cell adhesion
initial cell-cell contact
tissues move toward each other and contact each other for the first time
both express the same cad
form homophilic interaction across the gap between cells
cadheins cluster at initial contact site
step 2 of cell-cell adhesion
rac activation and arp2/3 nucleation
initial contact activates Rac1 at those sites
Rac1 activates arp2/3 at sites of initial contact
arp2/3 makes branched protrusions
Rac and Arp2/3 push the PM forward so it can form more connections with the underlying matrix through nascent adhesion and adhesion maturation
local rac activation and rho inhibition leads to loss of cortical tension
step 3 of cell-cell adhesion
rac1 turned off an RhoA turned on to strengthen adhesions
Rac1 activated, then RhoA turned off through cross-talk pathways
rac1 finishes expanding zone of contact, and GAP shuts it down (allows RhoA activity to increase at newly expanded site of contact
RhA activity is also responsible for strengthening adhesions
expansion of adherens junction
when rac1 activity is decreased…
RhoA activity is elevated
what happens when a-cat is extended?
force-dependent change in conformation reveals the new vinculin binding site
now have a second actomyosin fiber getting plugged into a-cat
positive feedback where you get massive expansion of contractility attached to growing/strengthening cads
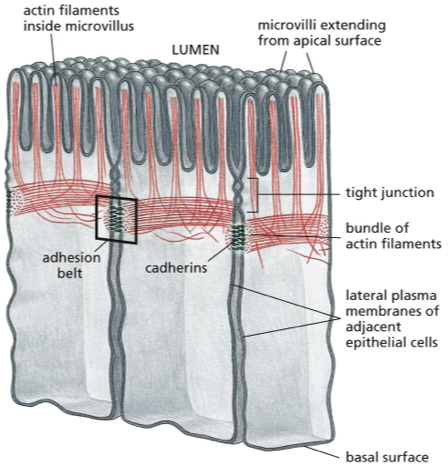
what are you left with at the end of the cell-cell adhesion process?
epithelial tissue with mature cell-cell junctions that are localized just below the apical surface (close to the top, but not at the very top)
cad role between cells
connected into “belt” of actomyosin contractility, which can generate further force in response to more activation signals
tissue shaping
spatially and temporally regulated actomyosin contractility and adhesion in epithelial cell monolayers
cell-cell contractility
belts of actomyosin bundles connected by homophilic e-cad binding
cell-matrix contractility
integrin-based cell-matrix adhesion connected by actomyosin stress fibers
order of layers in the cell
epithelial cells
basement membrane
collagen and fibroblasts
stroma
no BM
more loose and fibrile protein network
mostly made of type 1 collagen
cells can move through the fibers easily, unlike the BM
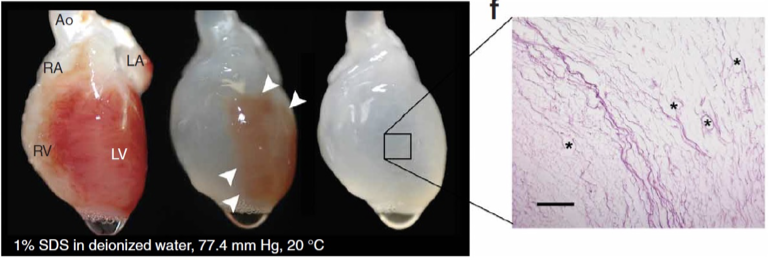
steps of ECM tissue formation
heart perfused with detergent to remove cells
only the ECM remains
adding neonatal cardiac cells to the matrix = beating heart
how do cells move and function in wound healing?
in wound healing, cross-talk between cells depositing matrix and the matrix that the cells deposit tell the other cells where to go, what to do, and how to differentiate
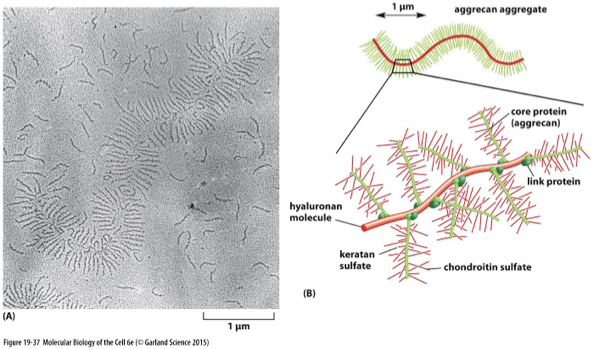
proteoglycans and GAGs
large, highly charged polysaccharides (GAGs) can be linked to protein (proteoglycan)
shock absorber
enriched in cartilage
fill spaces during development
polysaccharides attract _____ to ______
water, balance the charges
fibrous proteins are found in _____
dermis and BM
type 4 collagen
fibrous protein
component of BM
type 1 collagen
component of loose reticular stromal matrix
fibrillar collagen
protein that provides structure and stability to tissues like bone, cartilage, and skin (most abundant protein in the body)
glycoprotein
found in BMs
more protein than it is sugar
fibronectin
critical for development and wound healing
dynamic matrix protein which can be deposited in the context of wound healing/development to enhance adhesion of fibroblasts
laminin
cooperate with type 4 collagen to weave together the sheet-like BMs
nidogen
helps maintain BM and tissue architecture; also has role in cell adhesion, migration, and signaling
proteoglycans are composed of ….
GAG chains covalently linked to core proteins
linking to form core proteins
core proteins are synthesized in the ER
polysaccharides added in the Golgi
mature proteoglycan secreted into the extracellular space
aggrecan
enriched in cartilage
acts like shock absorber to protect joints from mechanical stress due to high-degree of hydration
single collagen fiber
triple helix of 3 different collagen proteins woven together — α, β, γ
bundling of collagen
bundled into thicker, stronger collagen networks
one way of being organized into higher-order structure
fibrils grouped to make fibers
alignment of collagen
all the bundles are going the same direction
can switch by layer; controlled by deposition
fibronectin
interact with integrin (cell)
interact with collagen and fibrin (makes up blood clots)
acts like signal (development, wound healing)
fibronectin & wound healing
wound: vessel severed
fibronectin spills out into the surrounding stroma (matrix surrounding the blood vessels; enriched in type 1 collagen)
fibronectin binds to collagen and to nearby cells
cells activated and provided with additional adhesive sites to move along the fibro-bound collagen
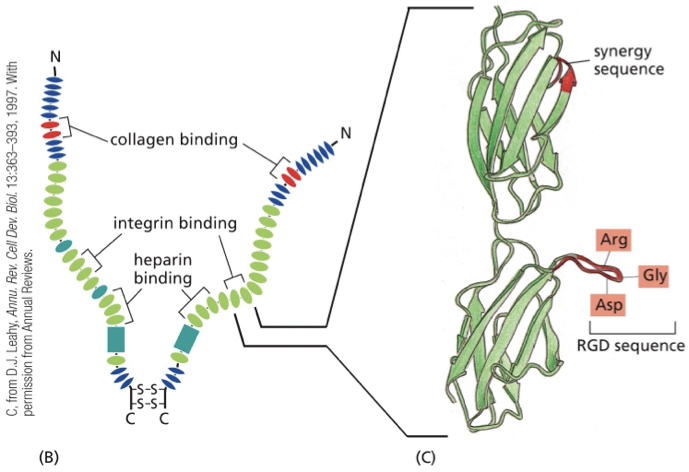
why does BM look like a sheet?
the 2 matrix proteins that make up the BM (type 4 collagen and laminin) have many different binding sites for other matrix proteins and cells
the more binding sites there are ….
more tightly woven together the network will be
laminin is a ____, and can bind _____
trimer
integrins, other laminins, and other matrix proteins (it’s a trimer)
type 4 collagen
forms many additional connections compared to type 1 collagen
comes together with other proteins to form the sheet-like structure that is woven together tightly to be a barrier
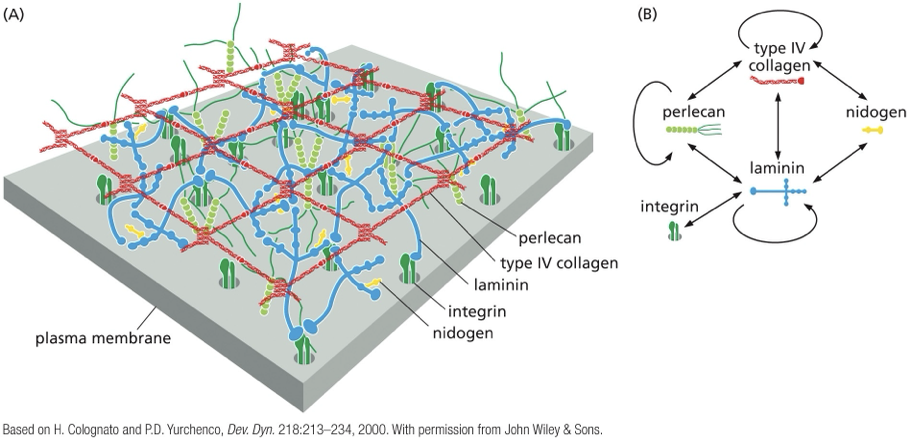
inherent complexity
the systems are interconnected by cytoskeleton and signaling cross-talk
hyaluronan
large GAG which is produced at the cell surface by a complex of enzymes
why is hyaluronan important?
important in development and wound healing by providing a transient space-filling material that can maintain the shape of a tissue while cells migrate
proteoglycans, GAG chains, core protein
core protein synthesized in the ER
polysaccharides added in the Golgi
mature proteoglycan secreted to the extracellular space