Metabolismo Lipídico Completo
1/17
Earn XP
Description and Tags
Todo o metabolismo de Lipoproteínas e Lipídios
Name | Mastery | Learn | Test | Matching | Spaced |
|---|
No study sessions yet.
18 Terms
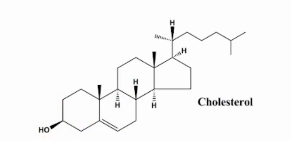
estrutura de colesterol
Estruturalmente, é uma molécula caracterizada por quatro anéis coplanares formando um núcleo esteroidal apolar que faz parte de uma cauda hidrofóbica e um grupo hidroxila polar, tornando-o anfipático.
Estrutura do Ácido Graxo Livre
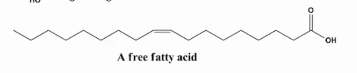
Os ácidos graxos livres são moléculas formadas por uma longa cadeia de carbonos, ligadas a um grupo carboxila (–COOH) na extremidade, que também conferem características anfipáticas.
Se organizam em micelas atraindo água, oque seria uma estocagem ineficiente.
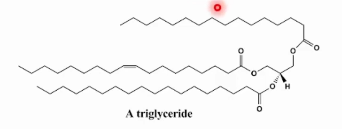
Estrutura do Triglicerídeo
3 Ácidos graxos esterificados em uma molécula de glicerol completamente apolar estocada na forma anidra.
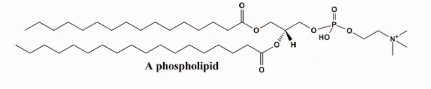
Estrutura do Fosfolipídio
duas cadeias de ácidos graxos ligadas a um glicerol e a um grupo fosfato, conferindo-lhes características anfipáticas. Eles são componentes fundamentais das membranas celulares, formando bicamadas.
Colesterol Utilização
Um esterol que é um importante componente das membranas celulares e precursor de hormônios esteroides e sais biliares.
Entrega e Remoção dos tecidos

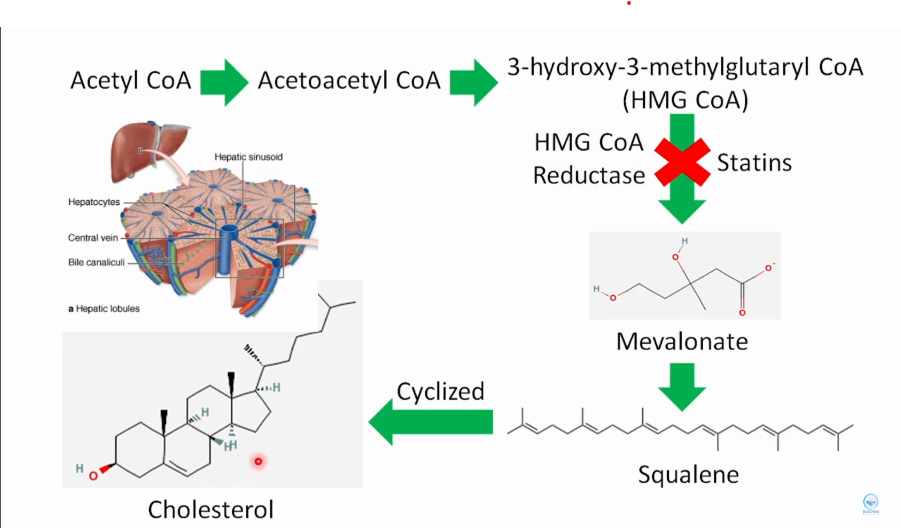
Síntese endógena de colesterol
HMG CoA Redutase é a enzima chave na via de síntese do colesterol, convertendo HMG-CoA em ácido mevalônico.
Sais biliares
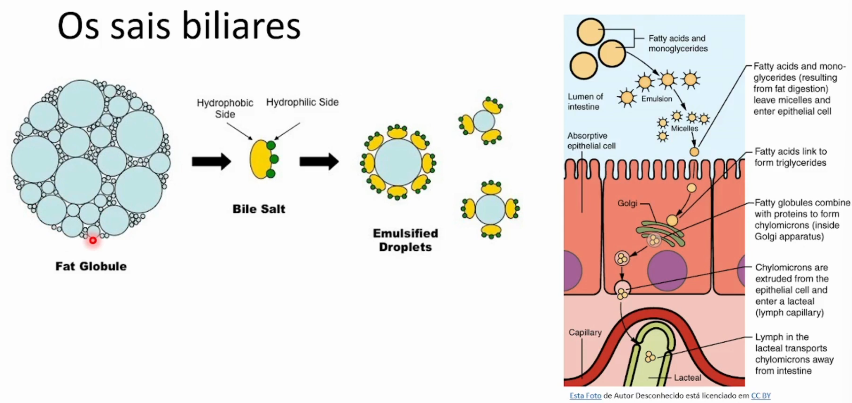
Moléculas anfipáticas derivadas do colesterol que emulsificam gorduras estabilizando pequenas gotículas e permitindo acesso de enzimas pelo aumento de superfície para a digestão e absorção de lipídios no intestino.
grandes gotas de moléculas gordurosas se emulsificam em micelas menores que são absorvidas pelo intestino delgado.
Bile
Mistura de sais biliares, fosfolipídios e colesterol, que ajuda na emulsificação e digestão de gorduras no intestino.
Colecistocinina (CCK):
hormônio gastrointestinal liberado após a ingestão de gorduras, estimulando a contração da vesícula biliar para liberar bile no intestino.
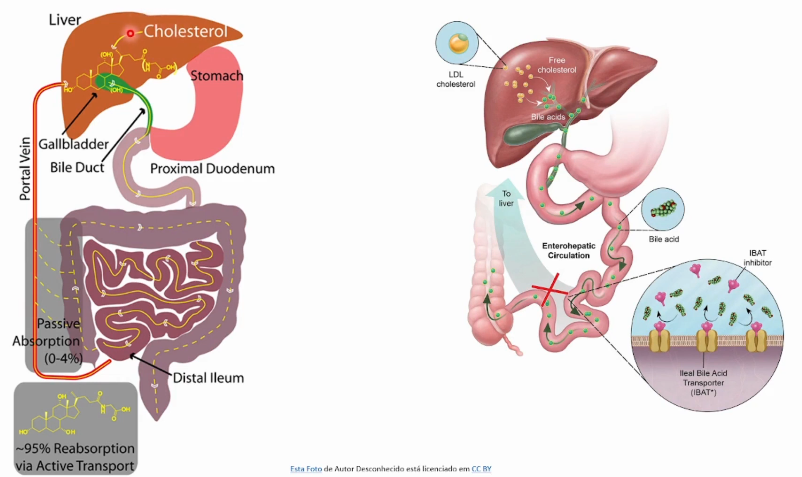
95% dos sais biliares são reabsorvidos pelo organismo via circulação porta hepática e reutilizados pelo fígado.
Sais biliares podem ser sequestrados por moléculas positivas que impedem sua reabsorção e ajudam na redução dos níveis de colesterol.
Como os lipídios são transportados na corrente sanguínea
Lipídios são transportados na corrente sanguínea principalmente através de lipoproteínas, que são partículas compostas de lipídios e proteínas, permitindo a solubilização e o transporte eficiente de gordura no plasma sanguíneo.
Monocamada de fosfolipídios com moléculas de colesterol livre e apolipoproteínas com função de estabilizar e endereçaras lipoproteínas para os tecidos.
Em seu centro possui triglicerídeos e ésteres de colesterol.
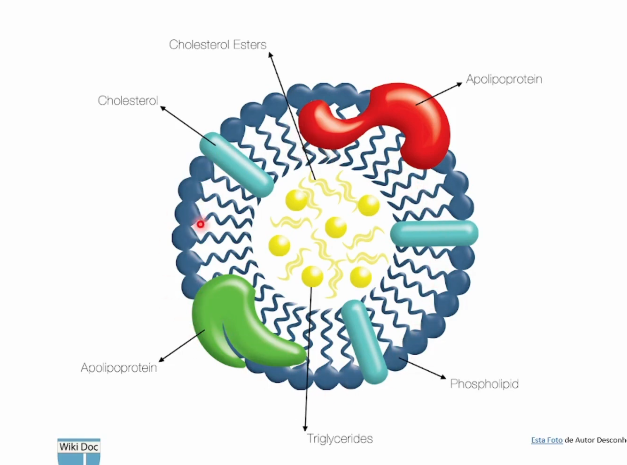
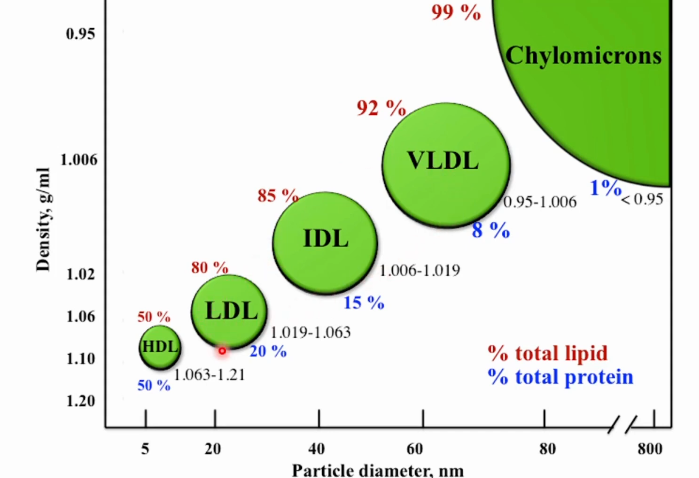
Diferentes Lipoproteínas
Quilomícrons (transporte de gorduras da via exógena)
VLDL, LDL,IDL (transporte de colesterol e risco cardiovascular), e HDL (transporta colesterol para o fígado).
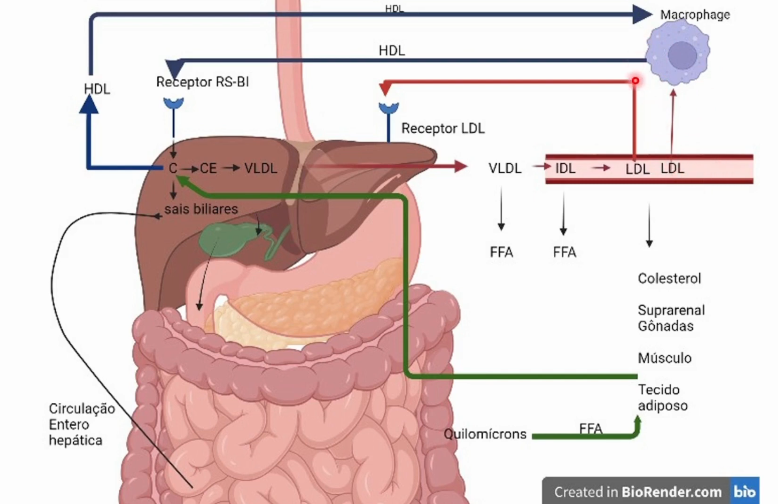
Como funciona o transporte de gorduras endógenas e exógenas no organismo
Quimiosensores no duodeno percebem a presença de moléculas gordurosas e há ejeção da bile no duodeno(CCK) que emulsifica as gorduras em micelas para que as Lipases tenham acesso a gordura, liberando ácidos graxos livres. Os ácidos graxos livres, colesterol e outras moléculas apolares são absorvidas pelos enterócitos e são empacotadas em lipopartículas de Quilomícrons que são muito grandes e não permeiam capilares diretamente carregando as gorduras da dieta, então entram no sistema linfático e posteriormente na circulação sistêmica para o tecido adiposo e músculo liberando ácidos graxos livres para produção energética, se tornando um quilomícron remanescente com menos ácidos graxos livres e colesterol e é endocitado no fígado. No fígado, libera colesterol livre nos hepatócitos que percebem a quantidade de colesterol da dieta e ativa ou inativa as enzimas de produção endógena de colesterol. O colesterol pode ser transformado em sais biliares que são armazenados e concentrados na vesícula biliar. O colesterol total tem parte produzida pelo fígado e parte dos quilomícrons remanescentes. É esterificado e empacotado em VLDL com colesterol e TAG. O VLDL é liberado pelo fígado e atinge a circulação sistêmica liberando ácidos graxos livres nos tecidos periféricos, se transformando em IDL que libera ácidos graxos livres nos tecidos periféricos pela ação da lipoproteína lipase presente no endotélio vascular dos músculos e tec. adiposo e se transforma em LDL que pode retornar ao fígado e internalizado pelos hepatócitos, ou fornecer colesterol para membranas plasmáticas ou síntese de hormônios esteroidais nas gônadas ou glândulas suprarrenais ou ainda ser oxidado na corrente sanguínea e fagocitado pelos macrófagos formando placas de ateroma. As placas de ateroma podem ser diminuídas pelo HDL, produzido pelo fígado com alto teor de proteína comparado a seu conteúdo de colesterol, atinge a circulação sistêmica e passa por tecidos periféricos que estão reciclando suas membranas e capta colesterol também das placas de ateroma, evitando o acúmulo de colesterol e ajudando a transportá-lo de volta ao fígado.
oque acontece após os quilomícrons estarem na corrente sanguínea
os quilomícrons na corrente sanguínea interagem com a Lipoproteína Lipase (LPL) do endotélio capilar, hidrolisando os triglicerídeos em ácidos graxos livres e glicerol, os AGL são reesterificados em triglicerídeos nos adipócitos ou oxidados nos miócitos para produção de energia, o glicerol é metabolizado pelo fígado e utilizado para produção de energia na via da glicólise e gliconeogênese ou utilizado para formação de TAG que é empacotado em VLDL e liberado na corrente sanguínea. À medida que perdem triglicerídeos, os quilomícrons remanescentes são endocitados pelos hepatócitos, (onde o colesterol sinalisa para inibição da síntese endógena de colesterol via bloqueio da transcrição da enzima HMG-CoA Redutase) é usado na produção de sais biliares e para juntamente com o colesterol endógeno ser empacotado em VLDL.
explique a lipólise
Hidrólise dos TAG presentes em gotículas de gordura armazenadas nos adipócitos em AGL e GLICEROL. O estado de jejum e estresse estimula a presença dos hormônios epinefrina, norepinefrina, glucagon e cortisol que estimulam a via da lipólise, que inicia da seguinte forma:
A enzima Lipase de Triglicerídeo Adiposa (ATGL) atua hidrolisando o TAG em AGL e DAG.
A enzima Lipase Hormônio Sensível (LHS) atua clivando o DAG em AGL e MAG.
A enzima Lipase de Monoacilglicerol (LMG) atua clivando o MAG em AGL e GLICEROL.
Os AGL são liberados na corrente sanguínea e transportados pela proteína albumina sérica aos músculos esqueléticos e cardíacos e Fígado. Nos músculos, os AGL são oxidados via beta oxidação para produção de ATP. No fígado, os AGL podem ser reesterificados em TAG (para empacotamento em VLDL), oxidados para produção de ATP, ou convertidos em corpos cetônicos.
O glicerol é liberado na corrente sanguínea e é metabolizado pelo fígado, onde pode ser utilizado para a gliconeogênese (principalmente em jejum) ou entrar na via glicolítica para produção de ATP.
Explique a Beta Oxidação
A beta oxidação começa com a conversão de um AGL no citosol dos hepatócitos ou miócitos em um Acil-CoA, catalisada pela acil-CoA sintetase. Este Acil-CoA atravessa a membrana externa da mitocôndria e chega ao espaço intermembranar. Ali, as moléculas de acil-CoA de cadeia curta atravessam a membrana interna e chegam à matriz mitocondrial. As moléculas de acil-CoA de cadeia longa são transportadas via carnitina, onde o grupo acil é transferido à carnitina, formando acilcarnitina. A acilcarnitina atravessa a membrana interna e chega à matriz mitocondrial. Na matriz mitocondrial, a acilcarnitina transfere o grupo acil para uma coenzima A, formando uma molécula de Acil-CoA.
Na matriz mitocondrial, a molécula de Acil-CoA sofre a beta-oxidação, onde cada ciclo promove a remoção de 2 carbonos na forma de acetil-CoA da molécula de acil-CoA. A beta oxidação é assim:
Oxidação: via ação enzimática, ocorre uma oxidação que promove a criação de uma dupla ligação do tipo trans entre os carbonos alfa e beta, produzindo um FADH₂.
Hidratação: via ação enzimática, a dupla ligação é hidratada (adiciona-se uma molécula de H₂O).
Oxidação: via ação enzimática, o grupo hidroxila do carbono beta é oxidado e forma-se uma carbonila, produzindo um NADH + H⁺.
Tiólise: via ação enzimática ocorre a clivagem da ligação entre os carbonos alfa e beta, adicionando uma molécula de coenzima A, gerando uma molécula de acetil-CoA e outra de acil-CoA com 2 carbonos a menos que o acil-CoA original.
O acetil-CoA é utilizado via ciclo do ácido cítrico para produção de ATP.
O FADH₂ e NADH + H⁺ são utilizados na fosforilação oxidativa para produção de ATP.
Explique a lipogenese
A lipogênese é ativada em superávit energético, quando a ingestão calórica excede as necessidades do corpo. Ocorre no citosol de células do fígado e tecido adiposo.
1. Disponibilização do Acetil-CoA:
O excesso de glicose da dieta é convertido em piruvato via glicólise. O piruvato entra na mitocôndria e é convertido em Acetil-CoA.
Quando as necessidades energéticas estão satisfeitas e o ciclo de Krebs desacelera (devido a altos níveis de ATP e NADH), o citrato (formado pela condensação de Acetil-CoA com oxaloacetato) acumula-se na mitocôndria.
Este citrato é transportado para o citosol, onde é clivado, regenerando Acetil-CoA.
2. Síntese de Ácidos Graxos:
O Ciclo de Alongamento:
Cada vez que a cadeia do ácido graxo é alongada em dois carbonos, ocorrem quatro reações principais:
Condensação: A cadeia de ácido graxo já existente se liga a dois carbonos do Malonil-CoA. O resultado é uma cadeia dois carbonos mais longa, mas com um grupo cetona.
Primeira Redução: O grupo cetona é reduzido a um grupo hidroxila. Esta reação requer elétrons, que são fornecidos pelo NADPH.
Desidratação: Uma molécula H₂O é removida da estrutura, criando uma dupla ligação entre os carbonos alfa (α) e beta (β).
Segunda Redução: A dupla ligação é novamente reduzida pela adição de hidrogênios, também fornecidos pelo NADPH. Isso resulta em uma cadeia de ácido graxo saturada, agora alongada em dois carbonos em relação ao início do ciclo.
Repetição e Produto Final:
A cadeia de ácido graxo alongada serve como ponto de partida para o próximo ciclo de adição de dois carbonos (novamente a partir de um Malonil-CoA).
Este ciclo de quatro etapas (condensação, redução, desidratação, redução) repete-se até que uma cadeia de 16 carbonos, como o palmitato, seja formada e então liberada.
3. Síntese de Triacilgliceróis para Armazenamento:
Os ácidos graxos sintetizados (ou provenientes da dieta) são ativados a Acil-CoA.
Estes são então esterificados a um esqueleto de glicerol-3-fosfato para formar TAGs.
Destino dos TAGs:
No fígado, os TAGs são incorporados em VLDL (lipoproteínas de densidade muito baixa) e secretados na corrente sanguínea para serem distribuídos aos tecidos periféricos.
No tecido adiposo, os TAGs são armazenados em grandes gotículas lipídicas como a principal reserva de energia do corpo.
explique a cetogênese
A cetogênese é o processo de formação de corpos cetônicos (acetoacetato, β-hidroxibutirato e acetona) a partir do Acetil-CoA. Este processo ocorre nas mitocôndrias dos hepatócitos.
Contexto Metabólico
Em situações como o jejum prolongado, as reservas de glicogênio se esgotam e a glicemia cai. Como resposta, a lipólise é intensamente estimulada no tecido adiposo, liberando grandes quantidades de ácidos graxos na corrente sanguínea.
Esses ácidos graxos são captados principalmente pelo fígado, onde sofrem β-oxidação, resultando em uma alta produção de Acetil-CoA. Simultaneamente, para tentar restaurar os níveis de glicose sanguínea, o fígado intensifica a gliconeogênese. Um substrato crucial para a gliconeogênese é o oxaloacetato, que também é um intermediário essencial do ciclo de Krebs (necessário para se condensar com o Acetil-CoA). Portanto, o desvio do oxaloacetato para a gliconeogênese diminui sua disponibilidade para o ciclo de Krebs.
Essa combinação de excesso de Acetil-CoA (pela alta β-oxidação) e deficiência relativa de oxaloacetato (pelo desvio para a gliconeogênese) impede que todo o Acetil-CoA seja oxidado no ciclo de Krebs, direcionando-o então para a via da cetogênese no fígado.
Dietas com restrição severa de carboidratos (cetogênicas).
Fluxo da Via Cetogênica
Duas moléculas de Acetil-CoA se condensam para formar acetoacetil-CoA.
O acetoacetil-CoA condensa-se com uma terceira molécula de Acetil-CoA para formar HMG-CoA.
O HMG-CoA é então clivado para produzir acetoacetato e uma molécula de Acetil-CoA.
O acetoacetato pode ser reversivelmente reduzido a β-hidroxibutirato, utilizando NADH como doador de elétrons.
O fígado sintetiza os corpos cetônicos (acetoacetato e β-hidroxibutirato), mas não os utiliza como fonte de energia devido à ausência da enzimas. Em vez disso, eles são exportados para a corrente sanguínea e transportados para tecidos como cérebro, músculo esquelético e músculo cardíaco, onde podem ser reconvertidos a Acetil-CoA e oxidados no ciclo de Krebs para produção de ATP.
receptores metabotrópicos do tipo gs,gi e gq
O sinal ativa o receptor acoplado à proteína G, que atua como um Fator de Troca de Nucleotídeos de Guanina (GEF). Este receptor promove a liberação de GDP da subunidade α da proteína G, permitindo que o GTP (presente no citosol) se ligue. A ligação do GTP ativa a subunidade α e promove sua separação do complexo βγ.
A subunidade αs atua ativando a enzima adenilato ciclase, que transforma ATP em AMPc. O AMPc ativa a PKA, que atua no metabolismo, transcrição gênica e canais iônicos.
A subunidade αi atua inibindo a enzima adenilato ciclase, diminuindo os níveis de AMPc e, consequentemente, a ativação da PKA. O complexo βγ da proteína Gi pode atuar na abertura de canais de potássio, permitindo seu efluxo e hiperpolarizando a célula.
A subunidade αq atua ativando a Fosfolipase C (PLC). A PLC cliva o Fosfatidilinositol 4,5-bisfosfato (PIP₂) da membrana em Diacilglicerol (DAG) e Inositol Trifosfato (IP₃). O DAG permanece na membrana e, juntamente com o cálcio, ativa a Proteína Quinase C (PKC). O IP₃ difunde-se pelo citosol e atua abrindo canais de cálcio regulados por IP₃ no Retículo Endoplasmático Liso (REL), promovendo o efluxo de cálcio para o meio intracelular. O aumento do cálcio intracelular, em conjunto com o DAG, ativa a PKC, que por sua vez modula o metabolismo, a transcrição gênica e canais iônicos. O cálcio também pode ativar outras vias, como aquelas mediadas pela calmodulina.
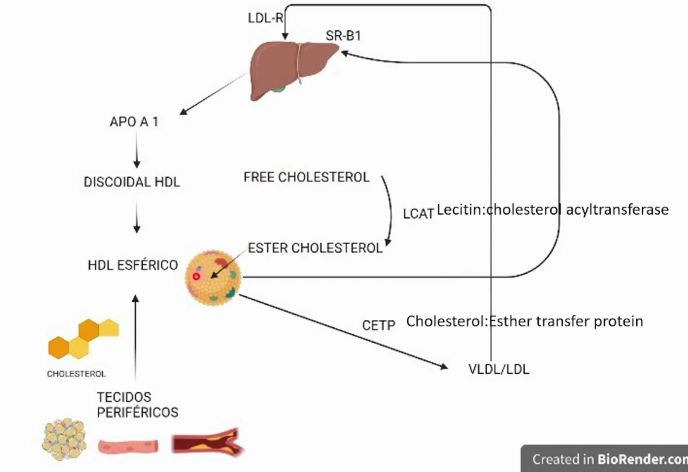
Explique o transporte reverso de colesterol
O HDL produzido pelo fígado possui Apolipoproteína APO-A1 estabiliza HDL.
A APO-A1 produzida e secretada pelo fígado ao atingir o meio extracelular se complexa com colesterol e fosfolipídios e forma uma partícula de pré-HDL com formato discoidal e baixa capacidade de captação de colesterol dos tecidos periféricos, o pré-HDL sofre ação da LCAT (lecitin colesterol aciltransferase), que catalisa a transformação de colesterol livre em colesterol esterificado que se acumula no interior do HLD que se torna maduro, capaz de captar colesterol dos tecidos periféricos e de placas ateroscleróticas diminuindo seu tamanho. O HDL cheio de colesterol retorna ao fígado via receptor SR-B1 e entrega moléculas de colesterol dentro dos hepatóicitos que fazem a eliminação correta do colesterol. Outra maneira é transferir o colesterol via CETP para os VLDL/LDL que vão ao fígado via LDL-R e são endocitadas pelos hepatócitos.