retina & visueel
1/108
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced |
|---|
No study sessions yet.
109 Terms
OVERZICHT: te beantwoorden vragen

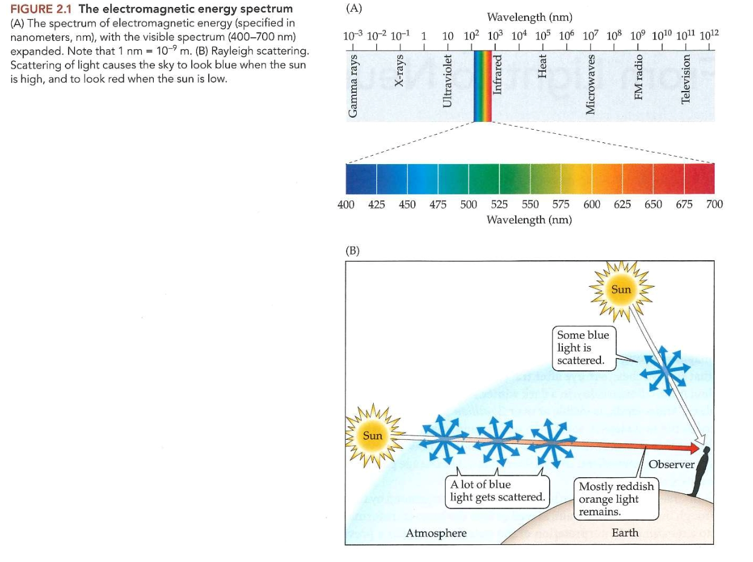
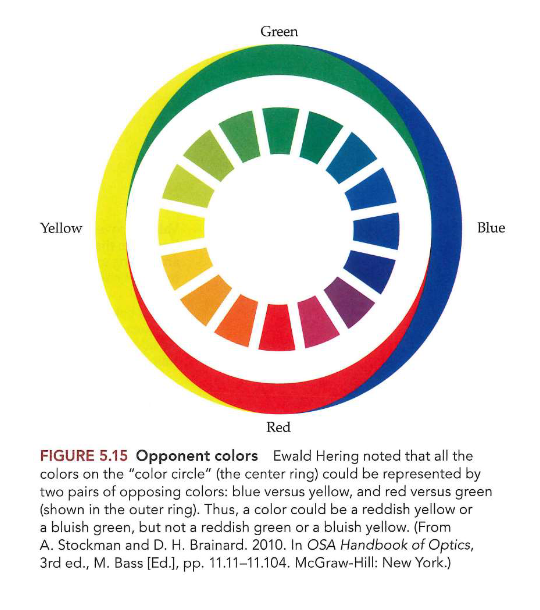
Elektromagnetisch spectrum:
Elektromagnetisch spectrum:
uit energie die varieert over zeer breed golflengtebereik
licht slechts klein deel van spectrum
zichtbare lichtgolven
golflengte tussen 400 – 700 nanometer (1nm = 10-9 m)
naarmate golflengte in zichtbare spectrum varieert → tint die we waarnemen verandert
400 nm → violet
650 nm → rood
ertussen: hele spectrum van regenboog
Lichtgolven zelf → niet gekleurd
pas nadat visuele systeem een inkomende golf heeft geïnterpreteerd, nemen we specifieke kleur waar
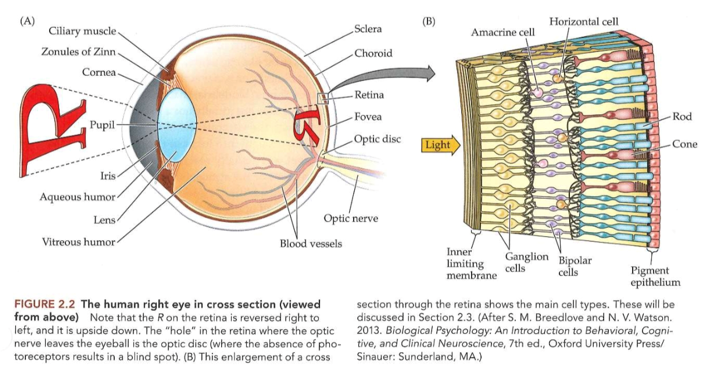
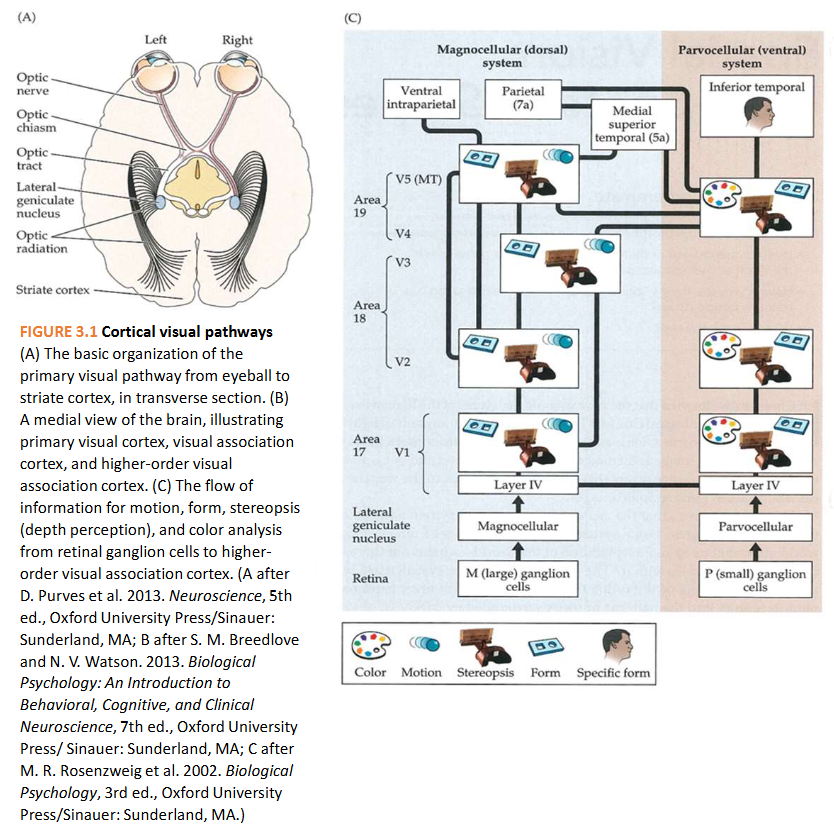
Ogen die licht capteren: het menselijk oog
→ doorsnede van voor naar achter door menselijk oog:
cornea
eerste weefsel waar licht tegenaan komt
= hoornvlies
transparant:
want zeer geordende reeks vezels
geen bloedvaten/ bloed (zouden licht absorberen)
grote hoeveelheid transparante sensorische zenuwuiteinden
dwingen ogen om te sluiten & tranen te produceren als hoornvlies gekrast raakt
voorste oogkamer
ruimte direct achter hoornvlies
gevuld met vloeistof = kamerwater
voorziet zowel hoornvlies als lens van zuurstof & voedingstoffen
lens
geen bloedtoevoer
volledig transparant
vorm van lens: bepaald door ciliaire spieren
pupil
licht moet hierdoor om lens te bereiken
= gat in spierstructuur (= iris)
belangrijke rol in beeldkwaliteit (zoals diafragma van camera)
bij weinig licht → pupil groot → scherptediepte (= bereik van afstanden waarover beeld scherp is) verminderd → slechte beeldkwaliteit
iris
geeft oog zijn kenmerkende kleur
regelt grootte van pupil → en dus hoeveelheid licht die netvlies bereikt
pupilreflex
afname van lichtsterkte → iris verwijdt automatisch om meer licht in oog toe te laten
toename van lichtsterkte → iris krimpt automatisch om minder licht in oog toe te laten
achterste oogkamer
= glasvochtkamer (ruimte tussen lens & netvlies)
glasvocht
gelachtig & over algemeen transparant
retina
= netvlies
licht wordt (normaliter) scherp gesteld
vanaf hier begint het zien echt → want hier lichtenergie omgezet in elektrische zenuwsignalen
= transductie
Ogen die licht capteren: accommodatie & verandering in functie van de leeftijd
om ver beeld op netvlies te focussen → brekingsvermogen van de 4 optische componenten van oog moet perfect afgestemd zijn op lengte van oogbol
optische componenten:
hoornvlies
kamerwater
lens
glasvocht
hoornvlies is sterk gebogen & heeft hogere brekingsindex dan lucht → krachtigste brekende oppervlak in oog
kamerwater & glasvocht helpen ook bij breken van licht
→ brekingsvermogen van deze 3 = onveranderlijk → dus niet te gebruiken om dichtbijzijnde objecten scherp te stellen
lens → brekingsvermogen kan veranderen door van vorm te veranderen
= accommodatie (verandering van focus)
bereikt door samentrekking van ciliarispier
lens via kleine vezels (ophangbanden = zonulae van Zinn) aan ciliarispier bevestigd
ciliarispier ontspannen → zonulae uitgerekt → lens relatief plat
oog zal dan scherpstellen op zeer verre objecten (bv. verre ster)
ciliarispier trekt samen → minder spanning op zonulae → lens staat bol
scherpstellen op iets dichterbij (bv. horloge/ smartphone)
hoe dikker lens → hoe meer ‘power’ → hoe dichterbij je kunt scherpstellen
power van de lens (P) = 1/f
f = brandpuntsafstand in meters
eenheid van power van de lens = dioptrie
→ vermogen om te accommoderen neemt af met leeftijd (± verlies van 1 dioptrie accommodatie per 5 jaar tot leeftijd van 30, en zelfs meer na leeftijd van 30)
tegen tijd dat meeste mensen tussen 40-50 jaar oud zijn:
niet makkelijk meer om 2,5 dioptrie te kunnen accommoderen die nodig is om scherp te zien op 40 cm
presbyopie = ouderdomsverziendheid → onvermijdelijk:
lens wordt harder
kapsel dat lens omgeeft (& waardoor lens van vorm kan veranderen) verliest elasticiteit
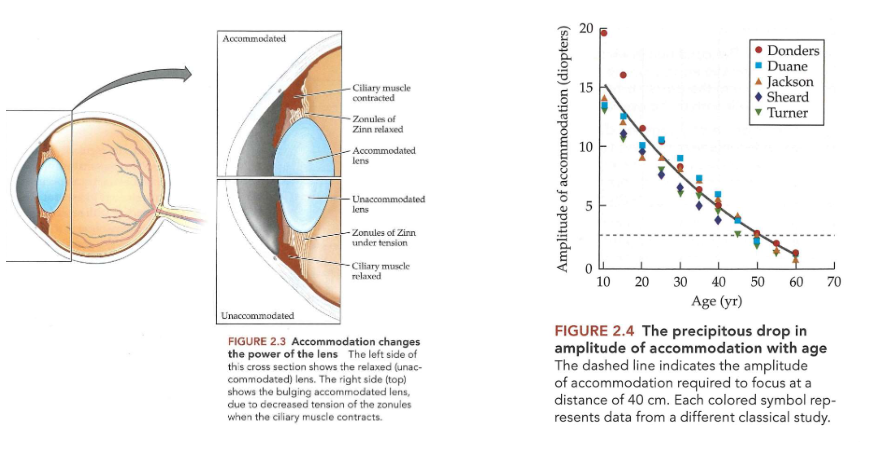
Ogen die licht capteren: cataract
= troebelingen van de lens
kan op verschillende leeftijden voorkomen & vele vormen aannemen
congenitaal cataract (aanwezig bij de geboorte)
relatief zeldzaam, maar kan desastreuze gevolgen hebben voor normale visuele ontwikkeling als het niet snel wordt behandeld wanneer het beeldkwaliteit thv netvlies ernstig verstoort
prevalentie van verworven cataract
neemt toe met leeftijd → rond 70 jaar bijna iedereen enig verlies van transparantie
behandeling:
troebele lens wordt verwijderd & vervangen door implantaat
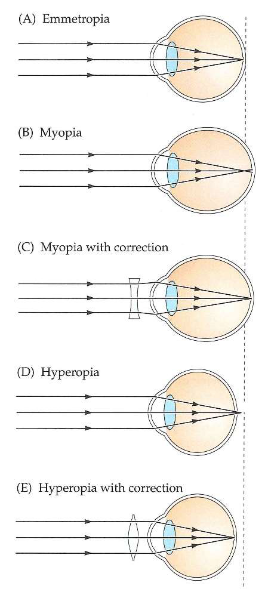
Ogen die licht capteren: optica van menselijk oog → emmetropie vs. refractieafwijkingen
emmetropie
brekingsvermogen van 4 optische componenten van oog (hoornvlies, kamerwater, lens & glasvocht) komen perfect overeen met lengte van oogbol
geen corrigerende lenzen nodig om objecten in verte te zien
refractieafwijkingen: wanneer oogbol te lang/ te kort is tov sterkte van 4 optische componenten
oogbol te lang → beeld van ster zal vóór netvlies worden gefocusseerd
ster → wazige vlek
myopie = bijziendheid
verre objecten niet scherp te zien
corrigeren met negatieve (min) lenzen → gaan stralen van sterrenlicht divergeren voordat ze oog binnendringen
oogbol te kort → beeld van ster achter netvlies gefocusseerd
hyperopie = verziendheid
ster → wazige vlek
corrigeren met positieve (plus) lenzen → gaan stralen van licht convergeren voordat ze oog binnendringen
Ogen die licht capteren: astigmatisme
hoornvlies → krachtigste brekende oppervlak in oog
levert ± 2/3 van focusvermogen van oog
bij emmetroop → hoornvlies = bolvormig (zoals voetbal)
indien hoornvlies niet bolvormig → kromming verschillend in horizontale & verticale meridianen (zoals rugbybal)
resultaat: astigmatisme
astigmatisme
verticale lijnen worden iets vóór netvlies scherp gesteld
horizontale lijnen worden iets achter netvlies scherp gesteld
→ of andersom
behandeling van astigmatisme:
lenzen met 2 brandpunten → lenzen met verschillende focussterkte in horizontale & verticale vlak

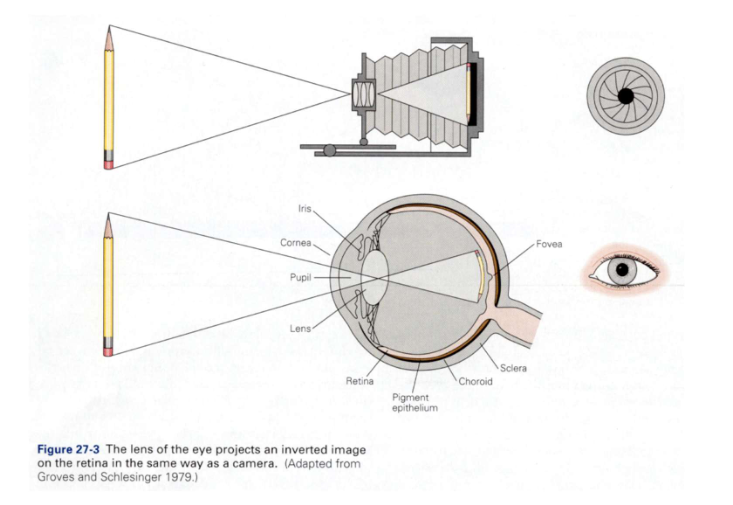
Ogen die licht capteren: oog vs. camera
gelijkenis:
allebei mechanisme voor reguleren van hoeveelheid licht (iris) & lens voor aanpassen van brandpuntsafstand
om zowel objecten dichtbij als veraf op netvlies scherp te stellen
verschil:
menselijke visuele systeem heeft taak om beeld te interpreteren
proces van zien begint bij netvlies/ retina → lichtenergie omgezet in neurale energie die door hersenen kan worden geïnterpreteerd
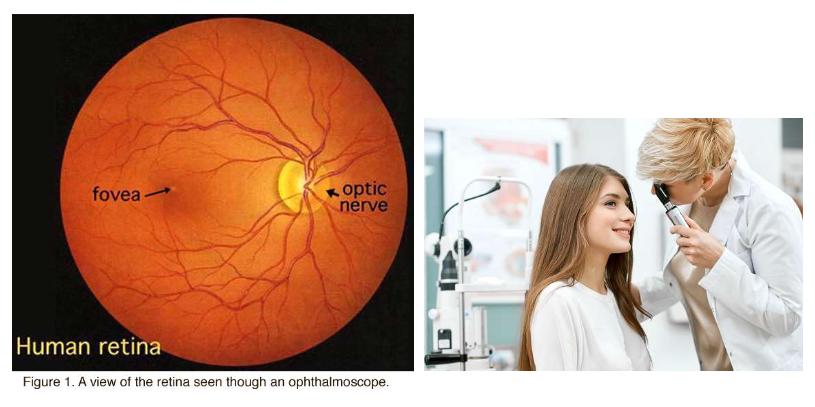
Ogen die licht capteren: de retina
ophthalmoscoop gebruiken om naar achterkant van oog te kijken
achterkant oog = fundus
witte cirkel = papil (optic disc)
punt waar slagaders & aders samenkomen en waar axonen van ganglioncellen het oog verlaten
dit deel van netvlies bevat geen fotoreceptoren → blind
= blinde vlek (fysiologisch)
fovea (vlekje van 1,5 mm) → in midden van netvlies
= plek van het scherpe zicht
nabij centrum van macula
centrale ~0,5 mm van fovea → geen bloedvaten → al het licht kan erdoorheen
geeft holte op de figuur
→ fundus = enige plaats in lichaam waar men slagaders & aders direct kan zien
belangrijk venster op gezondheid van vaatstelsel van lichaam
vasculaire ‘boom’ (= vertakte bloedvaten) spreidt zich op karakteristieke manier uit over netvlies, maar stopt net voor de fovea
Ogen die licht capteren: test voor de blinde vlek
blinde vlek in gezichtsveld normaal niet te zien:
we hebben 2 ogen → objecten waarvan beelden in blinde vlek van ene oog vallen, kunnen door andere oog gezien worden
door de oogbewegingen
hersenen die ontbrekende informatie aanvullen

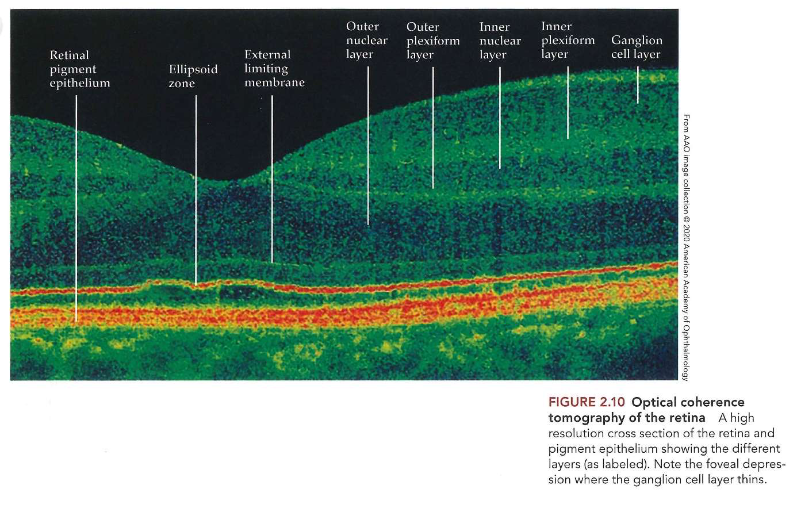
Ogen die licht capteren: optical coherence tomography (OCT)
netvlies bestaat uit meerdere lagen
± helft van dikte van een creditcard
laag pigmentepitheel achter laatste laag van retina
optical coherence tomography (OCT)
= niet-invasieve beeldvormingstechniek die gebruikt maakt van licht
laat oogarts toe om lagen van netvlies in dwarsdoorsnede te bekijken
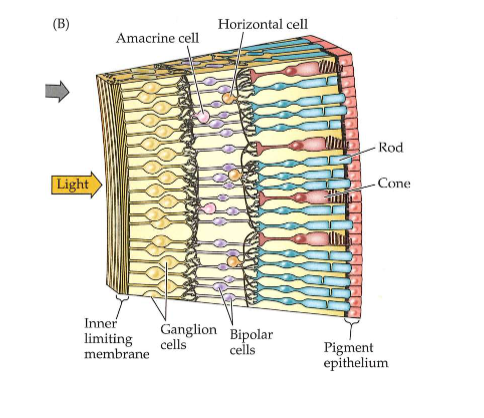
Ogen die licht capteren: neuronen in de retina
neuronen van retina vormen samen minicomputer → begint met interpreteren van informatie in visuele beelden
omzetting van lichtenergie in neurale energie → begint in achterste laag van netvlies
achterste laag: bestaat uit fotoreceptoren
fotoreceptoren nemen licht waar → stimuleren dan neuronen in tussenliggende lagen:
bipolaire cellen
horizontale cellen
amacriene cellen
→ neuronen maken dan verbinding met voorste laag van netvlies
voorste laag: bestaat uit ganglioncellen
axonen verlopen via optische zenuw naar hersenen
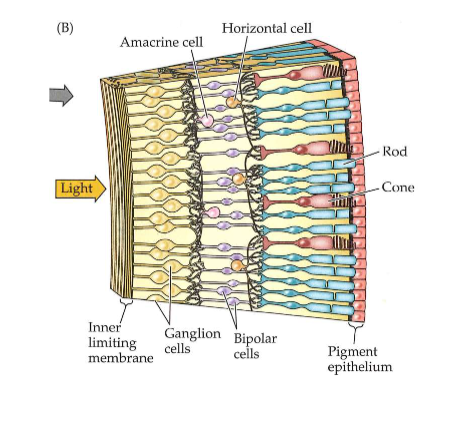
Ogen die licht capteren: neuronen in de retina → waarom bevinden fotoreceptoren zich aan de achterkant/ in laatste laag?
→ licht moet hierdoor door ganglioncellen, horizontale cellen & amacriene cellen voordat het de fotoreceptoren bereikt
MAAR: deze neuronen zijn grotendeels transparant, terwijl cellen in pigmentepitheel (leveren essentiële voedingsstoffen & recycling- (of housekeeping) functies aan fotoreceptoren) zijn ondoorzichtig
→ fotoreceptoren moeten naast beide componenten liggen:
naast pigmentepitheel → voor voeding & recycling
naast andere neuronen → om signalen door te geven
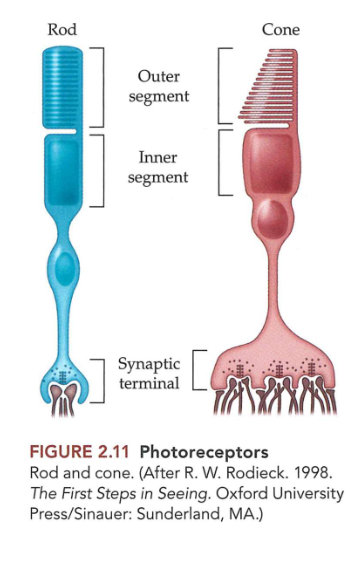
Ogen die licht capteren: fotoreceptoren
elk netvlies → ± 100 miljoen fotoreceptoren
fotoreceptoren
= neuronen die licht opvangen & zien initiëren door chemische signalen te produceren (signaaltransductie)
menselijk netvlies → minstens 2 soorten fotoreceptoren:
staafjes
kegeltjes
→ verschillende vorm, verschillend verdeeld over netvlies & vervullen verschillende functies
(synaps is met glutamaat)
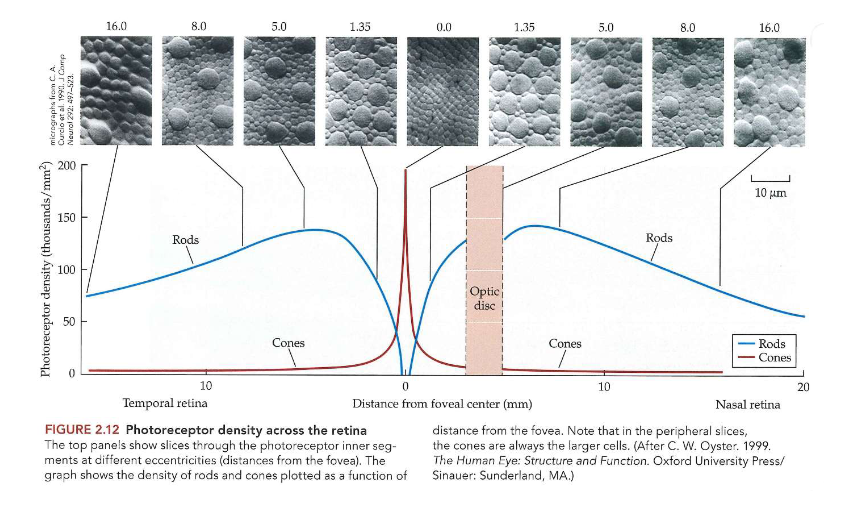
Ogen die licht capteren: densiteit van fotoreceptoren doorheen retina
algemeen: veel meer staafjes (± 90 miljoen in elk oog) dan kegeltjes (± 4-5 miljoen in elk oog)
2 soorten cellen hebben zeer verschillende geografische verdeling op netvlies
staafjes:
volledig afwezig in centrum van fovea
dichtheid neemt toe tot piek rond 20 graden, neem dan terug af
kegeltjes:
meest geconcentreerd in centrum van fovea
kegeltjes in foveale centrum: kleiner & dichter opeengepakt dan in andere delen van netvlies
gebied bevindt zich direct achter midden van pupil → over visuele hoek (= hoek thv oog) van ± 1 graad
dichtheid neemt dramatisch af met excentriciteit van netvlies (= afstand tot fovea)
kegeltjes worden groter & schaarser vanaf foveale centrum → kleine cellen die buiten fovea verschijnen zijn staafjes (zijn ongeveer even groot als kegeltjes in fovea)
→ grote cellen = kegeltjes (behalve op foto 0,0m)
→ fovea = kuiltje in netvlies dat gespecialiseerd is in zien van fijne details
geen probleem om te bewegen in het donker
komt doordat staafjes voldoende informatie geven voor navigatie
geen gat te zien wanneer we met staafjesvrije fovea direct naar een klein object kijken, ook al kan object zelf ‘verdwijnen’
MAAR: gezichtsveld lijkt naadloos te worden ingevuld door omringende netvlies
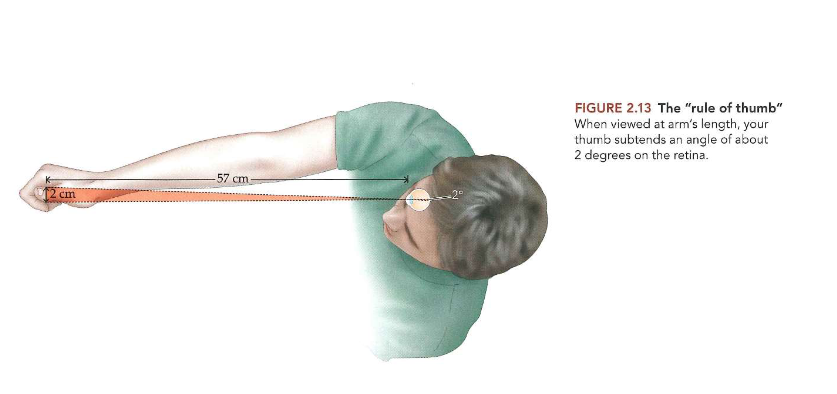
Ogen die licht capteren: de ‘rule of thumb’
→ om te weten hoe groot 1 graad is
vuistregel:
duim, bekeken op armlengte, maakt hoek van ± 2 graden op het netvlies (ervan uitgaande dat duim ± 2cm breed is & uitgestrekte arm zich ± 57cm van oog bevindt)
dus: als we direct naar een object kijken waarvan beeld kleiner is dan 1 graad, zal beeld op gebied van netvlies terechtkomen dat alleen kegeltjes heeft
Ogen die licht capteren: eigenschappen van fovea vs. periferie van netvlies
fovea
hoge gezichtsscherpte
gebruiken om objecten te identificeren, lezen & fijne details te inspecteren
perifere deel
voor detecteren & lokaliseren van stimuli waar we niet direct naar kijken (bv. zien van rijdende vrachtwagen in de ooghoek)
→ staafjes & kegeltjes werken beste onder verschillende lichtomstandigheden:
staafjes:
functioneren relatief goed onder omstandigheden van zwakke verlichting
= scotopisch zicht of nachtzicht
kegeltjes
heldere verlichting nodig (bv. zonlicht of kamerverlichting) om efficiënt te functioneren
= fotopische zicht
→ gebied in midden van fovea zonder staafjes: bij zwak licht is centrale 1 graad van fovea feitelijk blind
→ ander functioneel verschil tussen staafjes & kegeltjes: fotopigment
alle staafjes = hetzelfde type fotopigment
kunnen geen kleurverschillen signaleren
elk kegeltje → 1 van de drie verschillende fotopigmenten die verschillen in golflengten waarop ze licht het meest efficiënt absorberen
kegeltjes kunnen dus informatie over golflengte doorgeven → vormen basis voor kleurenzicht
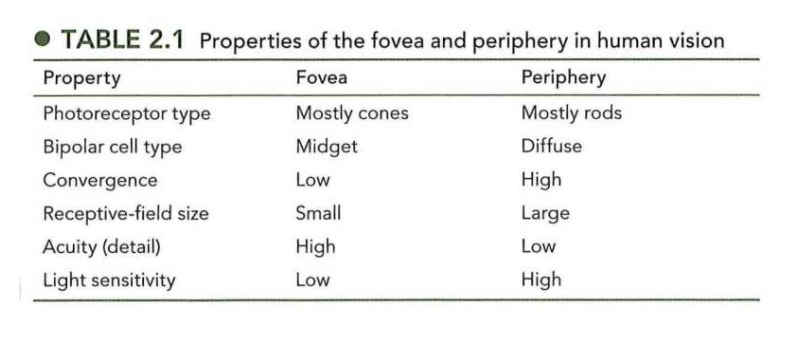
donker- & lichtadaptatie: donkeradaptatie
wanneer donkere kamer binnentreden vanuit fel zonlicht → aantal fotonen licht dat oog binnenkomt neemt af met factor van enkele miljarden (meer dan 12 logeenheden)
in het begin → moeilijk om iets te zien
na 30 min → zelfs slechts een paar fotonen waarnemen
donkerblauwe curve: verandering in drempel aan lichtintensiteit (= minste licht dat nodig is) om perifeer punt te detecteren
in begin: drempel hoog → lage gevoeligheid
na ± 20 min: drempel sterk verlaagd → gevoeligheid is verhoogd
vanuit donker terug naar zonlicht → vrijwel direct terug kunnen zien
donker- & lichtadaptatie: aanpassing aan verandering verlichting (4)
pupilgrootte
fotopigmentregeneratie
‘duplex’ netvlies
neurale circuits
aanpassing aan verandering verlichting (4) → pupilgrootte
zaklamp in iemands oog schijnen in slecht verlichte kamer → pupil vernauwt snel
diameter van pupil: kan variëren met factor 4
bij helder licht: 2mm
in het donker: 8mm
hoeveelheid licht in oog dat binnenkomt is evenredig met oppervlakte van pupil
=> viervoudige vergroting van diameter zorgt voor 16-voudige toename van hoeveelheid licht
MAAR: pupilverwijding helpt slechts voor een deeltje (maar enkele seconden) → want aanpassing in donker duurt vele minuten
dus: pupilgrootte is slechts klein onderdeel van totale vermogen van visuele systeem om zich aan te passen aan licht- & donkeromstandigheden
aanpassing aan verandering verlichting (4) → fotopigmentregerenatie
bij weinig licht: voldoende fotopigment beschikbaar → staafjes & kegeltjes absorberen zoveel mogelijk fotonen en reageren daarop
staafjes betere gevoeligheid dan kegeltjes bij weinig licht
fotopigmentmolecule gebruikt om foton te detecteren → ondergaat dan ‘bleaching’
moet geregenereerd worden voordat het opnieuw kan gebruikt worden om ander foton te absorberen
meer licht: aantal fotonen beginnen systeem te overbelasten → fotopigmentmoleculen kunnen niet snel genoeg worden geregenereerd om alle fotonen te detecteren die fotoreceptoren raken
→ langzame regeneratie is goede zaak voor vergroten van gevoeligheidsbereik
indien weinig fotonen → we gebruiken ze allemaal om te zien
indien te veel fotonen → we gooien er een deel van weg & gebruiken de rest
aanpassing aan verandering verlichting (4) → ‘duplex’ netvlies
staafjes: uitstekende gevoeligheid bij weinig licht → MAAR: raken overbelast wanneer achtergrondlicht matig fel wordt
dus: verlies aan beeldkwaliteit
kegeltjes: veel minder gevoelig dan staafjes (functioneren slecht bij zeer zwak licht) → MAAR: werkingsbereik veel groter:
10 fotonen per seconden → net genoeg licht om kleur te zien
honderdduizenden fotonen per seconde → bv. besneeuwde berg in fel zonlicht
→ staafjes om te zien bij weinig licht
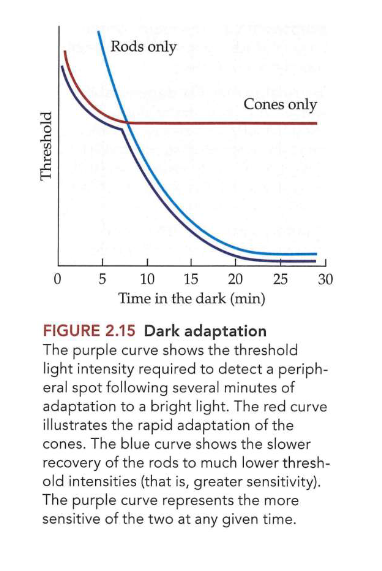
herstellen langzamer (blauwe curve op figuur over donkeradaptatie)
na ± 20 min zeer gevoelig voor zwak licht
wanneer volledig aangepast aan donker: licht dat dicht bij detectiedrempel ligt lijkt kleurloos
→ kegeltjes nemen werking over bij te veel licht om staafjes goed te laten functioneren
na aanpassing aan fel licht: kegeltjes herstellen snel gevoeligheid (rode curve op figuur over donkeradaptatie), raken dan verzadigd
niet erg gevoelig voor zeer zwak licht
aanpassing aan verandering verlichting (4) → neurale circuits
→ belangrijkste reden waarom we ons niet storen aan variaties in algehele lichtniveaus
ganglioncel: meest gevoelig voor verschillen in lichtintensiteit in ‘center’ & ‘surround’ van receptieve veld (= gebied op netvlies, en corresponderende gebied in visuele ruimte, waarin visuele stimuli de vuurfrequentie van neuronen beïnvloeden)
ganglioncellen: minder beïnvloed door gemiddelde lichtintensiteit
MAAR: zullen nog steeds met hogere snelheid vuren dan spontaan wanneer licht op gehele receptieve veld valt, zolang fotoreceptoren die ganglioncellen voeden niet volledig verzadigd zijn
→ ganglioncellen coderen dus patroon van relatief lichte & relatief donkere gebieden in netvliesbeeld
patroon van illuminatie (niet algemene lichtniveau) → primaire zorg van rest van visuele systeem
donker- & lichtadaptatie: ziekten van staafjes en/of kegeltjes
→ degeneratie van staafjes en/of kegeltjes
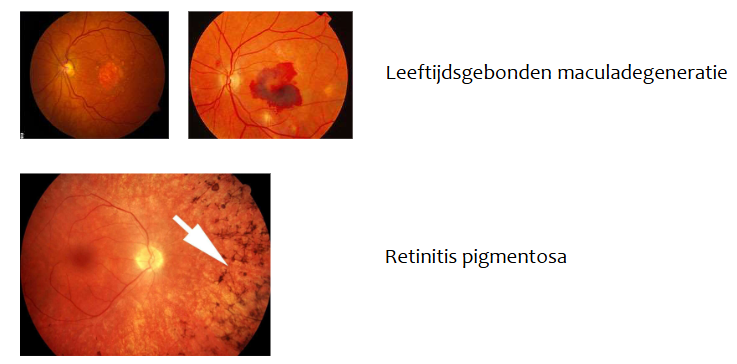
leeftijdsgebonden maculadegeneratie
ziekte door veroudering
tast macula aan
scherpe centrale zicht wordt vernietigd → moeilijk om te lezen, auto te rijden & gezichten te herkennen
retinitis pigmentosa
progressieve degeneratie van netvlies
tast nachtzicht & perifere zicht aan
vaak voorkomend in families → veroorzaakt door defecten in verschillende genen
retinale informatieverwerking: mechanisme
→ netvlies: uit 5 hoofdklassen van neuronen:
fotoreceptoren
horizontale cellen
bipolaire cellen
amacriene cellen
ganglioncellen
→ mechanisme:
lichttransductie door staafjes- & kegeltjesfotoreceptoren
laterale inhibitie door horizontale cellen
convergentie & divergentie van informatie via bipolaire cellen
communicatie met hersenen via ganglioncellen
center-surround receptieve velden
P- & M-ganglioncellen herbekeken
intrinsiek fotogevoelige retinale ganglioncellen in ontwikkelende netvlies
lichttransductie door staafjes- & kegeltjesfotoreceptoren
fotoreceptoren vangen licht op → produceren dan chemische veranderingen die cascade van neurale gebeurtenissen in gang zetten → eindigen in visuele waarneming
signalen sturen via synaptische uiteinden:
gespecialiseerde structuren voor contact met horizontale & bipolaire cellen
fotoreceptoren:
buitenste segment → grenst aan pigmentepitheel
binnenste segment
kleine fabriek, gevuld met mitochondriën
synaptisch uiteinde
visuele pigmenten
molecules die gemaakt worden in binnenste segment
worden opgeslagen in buitenste segment → geïncorporeerd in membraan
elke visuele pigmentmolecule bestaat uit:
eiwit (= opsine)
structuur bepaalt welke golflengten van licht de pigmentmolecule absorbeert
chromofoor
capteert de fotonen
= retinal/ retinaldehyde
afgeleid van vitamine A (= retinol) → wordt geproduceerd uit bètacaroteen
→ opsine & chromofoor zijn met elkaar verbonden
lichttransductie door staafjes- & kegeltjesfotoreceptoren → elke fotoreceptor: slechts 1 van 4 soorten visuele pigmenten
rhodopsine
in staafjes
voornamelijk geconcentreerd in stapel membraanschijven in buitenste segment
elk kegeltje → 1 van de andere 3 pigmenten
reageren elk alleen op lange, middellange of korte golflengten
lichttransductie door staafjes- & kegeltjesfotoreceptoren → ander type fotoreceptor die ‘leeft’ tussen ganglioncellen
leeft tussen ganglioncellen
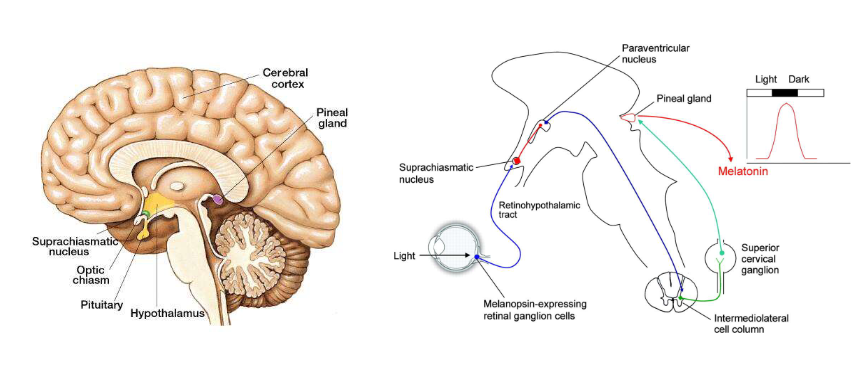
betrokken bij aanpassen van onze biologische ritmes aan dag & nacht van buitenwereld
gevoelig voor lichtniveau in omgeving
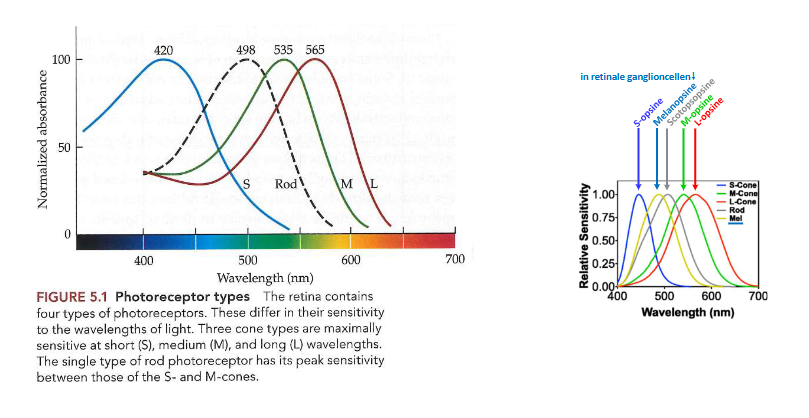
bevatten fotopigment melanopsine
sturen signalen naar suprachiasmatische nucleus via retinohypothalame tractus
suprachiasmatische nucleus = klein onderdeel van hypothalamus
fungeert als centrale circadiane klok die dagelijkse ritmes van slapen-waken & andere fysiologische functies reguleert
lichttransductie door staafjes- & kegeltjesfotoreceptoren → mechanisme fotoactivatie
foton bereikt buitenste segment van staafje → wordt geabsorbeerd door rhodopsine molecule
foton draagt energie over aan chromofoorgedeelte van visuele pigmentmolecule
= fotoactivatie/ bleaching
→ initieert biochemische cascade:
resulteert uiteindelijk in sluiten van celmembraankanalen die normaal ionen in buitenste segment van staafje laten stromen
sluiten van kanalen maakt binnenkant van cel negatiever → hyperpolarisatie
hyperpolarisatie sluit spanningsafhankelijke calciumkanalen bij synaptische terminal → concentratie vrij calcium in cellen neemt af
verlaging van calciumconcentratie vermindert concentratie van neurotransmitter glutamaat die in synaps vrijkomt → verandering geeft bipolaire cel het signaal dat staafje een foton heeft opgevangen
→ reeks gebeurtenissen duurt slechts milliseconden
(visuele pigmentmoleculen van kegeltjes werken op vergelijkbare manier)
→ hoeveelheid glutamaat die op bepaald moment aanwezig is in synaps tussen fotoreceptor & bipolaire cel, is omgekeerd evenredig met aantal fotonen dat door fotoreceptor wordt geabsorbeerd
fotoreceptoren werken dus niet op alles-of-niets-manier
geven informatie door aan bipolaire cellen via graded potentials
variëren in grootte (ipv all-or-none actiepotentialen, die overal in zenuwstelsel voorkomen)
lichttransductie door staafjes- & kegeltjesfotoreceptoren → pseudokleuring kegels in menselijke retina
de 3 fotopigmenten zijn niet gelijkmatig verdeeld over de kegeltjes
kegeltjes gevoelig aan korte golflengten (S-kegeltjes): 5-10% van totale kegeltjespopulatie
meer kegeltjes gevoelig voor lange golflengten (L-kegeltjes) dan kegeltjes die gevoelig zijn voor middellange golflengten (M-kegeltjes)
gemiddeld ± 2x zoveel L-kegeltjes als M-kegeltjes
MAAR: verhouding tussen L- & M-kegeltjes varieert enorm tussen individuen

laterale inhibitie door horizontale cellen
horizontale cellen
lopen loodrecht op fotoreceptoren
leggen contact tussen nabijgelegen fotoreceptoren
laterale verbindingen spelen belangrijke functionele rol in vorm van laterale inhibitie
laterale inhibitie:
signalen die retinale ganglioncellen bereiken → gebaseerd op verschillen in activering tussen nabijgelegen fotoreceptoren
belangrijke rol bij visuele perceptie & bij verschillende illusies (bv. Mach-banden & Hermann-raster)
amacriene cellen
ook deel van laterale route
lopen ook loodrecht op fotoreceptoren in binnenste lagen van netvlies
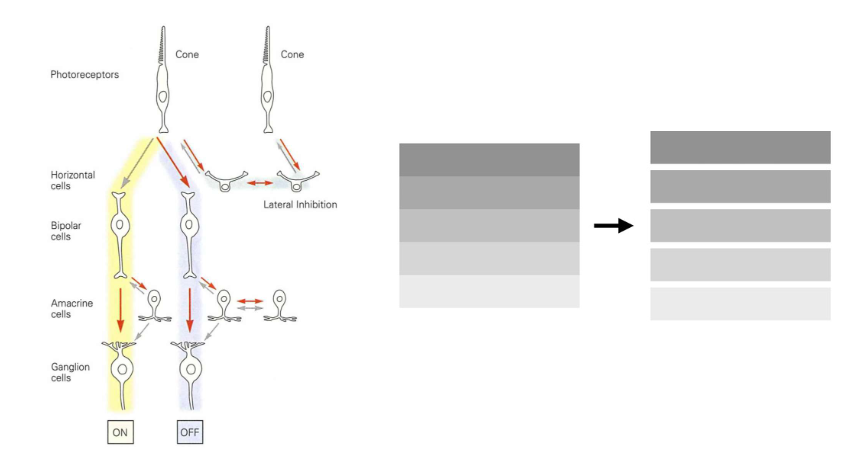
convergentie & divergentie van informatie via bipolaire cellen
verticale route: fotoreceptoren, bipolaire cellen & ganglioncellen
bipolaire cellen = intermediair
verschillende soorten bipolaire cellen & bedrading bepaalt informatie die van fotoreceptoren naar ganglioncellen wordt doorgegeven
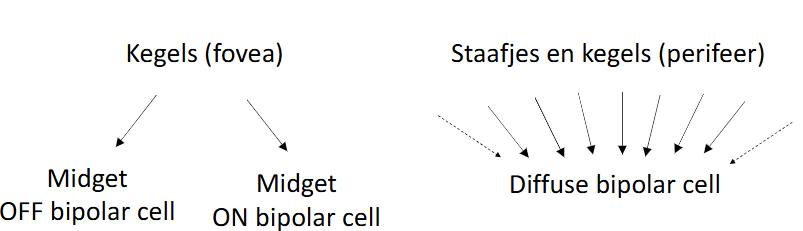
in perifere zicht:
bipolaire cel ontvangt input van wel 50 fotoreceptoren → bundelt info dan & geeft deze door aan ganglioncel
→ convergentie van informatie van vele fotoreceptoren naar 1 diffuse bipolaire cel
kenmerkend voor staafjesroute
convergentie ook optredend in kegeltjesroute in perifere netvlies
→ bundelen van informatie van vele fotoreceptoren: zeer belangrijk mechanisme om visuele sensitiviteit te verhogen
meeste staafjes communiceren met ganglioncellen via diffuse bipolaire cellen → verklaart waarom staafjessysteem goed kan functioneren in schemerige lichtomstandigheden:
diffuse bipolaire cel kan even snel reageren op 1 punt met fel licht of op meerdere vlekken met schemerig licht, omdat meerdere fotoreceptoren synaps nemen op elke diffuse bipolaire cel & ganglioncel die naar diffuse bipolaire cel luistert, niet in staat is te onderscheiden welk lichtpatroon aanwezig is
hoge graad van convergentie in perifere zicht: belangrijke gevolgen voor gezichtsscherpte (want verlies van detail)
gezichtsscherpte neemt snel af met excentriciteit
in de fovea:
kleine midget bipolaire cellen ontvangen input van afzonderlijke kegeltjes → geven informatie door aan afzonderlijke ganglioncellen
1-op-1 pathway tussen kegeltjes & ganglioncellen alleen bestaand in fovea
daarom beelden hier duidelijkst te zien wanneer ze op dit deel van netvlies vallen
→ hoge convergentiegraad in periferie van netvlies: zorgt voor hoge lichtgevoeligheid, maar lage gezichtsscherpte
→ lage convergentiegraad in fovea: zorgt voor hoge gezichtsscherpte, maar lage lichtgevoeligheid
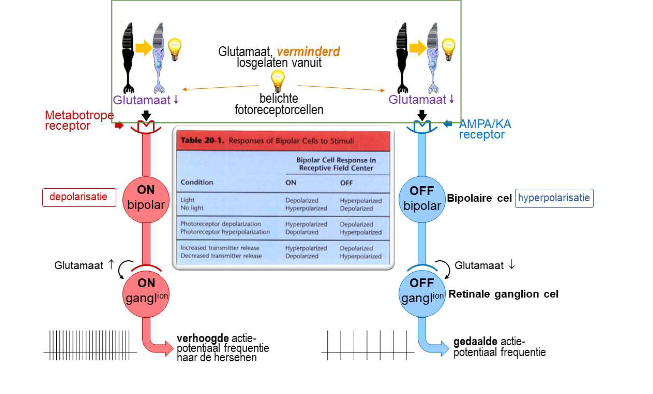
convergentie & divergentie van informatie via bipolaire cellen → divergentie van informatie via foveale ON & OFF bipolaire cellen
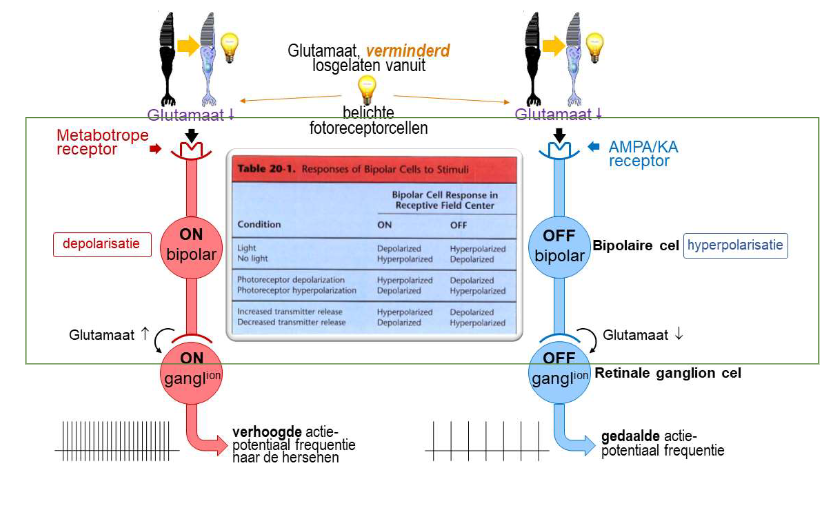
elke foveale kegel → maakt contact met 2 bipolaire cellen (= divergentie van informatie)
ON-bipolaire cel
depolariseert als reactie op toename van licht dat door kegel wordt opgevangen
met metabotrope receptoren
OFF-bipolaire cel
hyperpolariseert als reactie op toename van licht dat door kegel wordt opgevangen
met AMPA/KA receptoren
→ ON- & OFF-bipolaire cellen reageren verschillend op dezelfde fotoreceptorinput
want: brengen verschillende soorten postsynaptische glutamaatreceptoren tot expressie → zorgt voor veranderingen in membraanpotentiaal in tegengestelde richtingen
→ ON- & OFF-bipolaire cellen:
geeft informatie of belichting van netvlies toenam of afnam
ON/OFF-onderscheid → op veel niveaus van visuele pathway aanwezig
convergentie & divergentie van informatie via bipolaire cellen → ON & OFF ganglioncellen
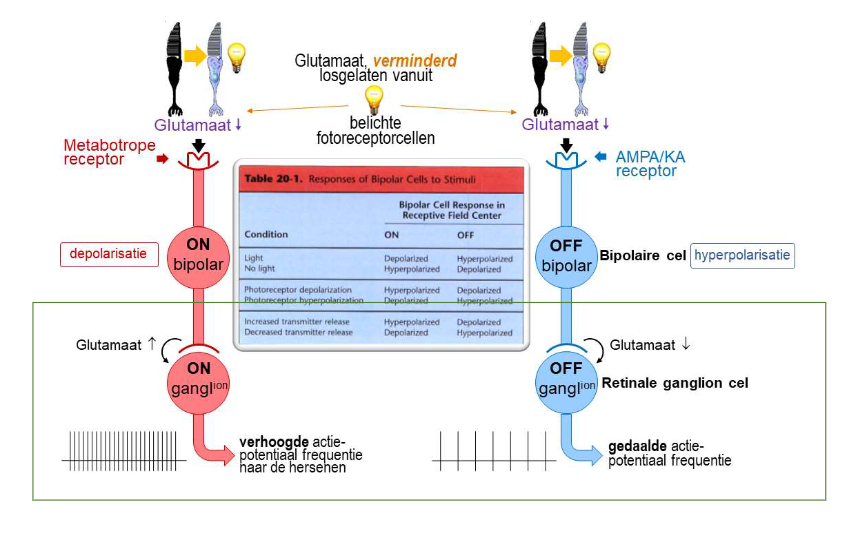
communicatie met hersenen via ganglioncellen
ganglioncellen → laatste laag van netvlies
tegen tijd dat signalen hier toekomen heeft er al veel informatieverwerking plaatsgevonden
sommige informatie → gebundeld door convergentie
andere informatie is versterkt of geïnhibeerd door laterale pathways
menselijk netvlies: ± 1.250.000 ganglioncellen = 1% van aantal fotoreceptoren
dus er is een serieuze convergentie gebeurd onderweg!
P-ganglioncellen
= kleine ganglioncellen
krijgen informatie van midget bipolaire cellen
projecteren naar parvocellulaire (kleine cel) laag van nucleus geniculatus lateralis (LGN)
vormen ± 70% van ganglioncellen in menselijk netvlies
M-ganglioncellen
= grote ganglioncellen
krijgen informatie van diffuse bipolaire cellen
projecteren naar magnocellulaire (grote cel) laag van LGN
dendrieten van M-ganglioncellen spreiden zich veel meer uit dan die van P-ganglioncellen → hebben parapluvormig uiterlijk
± 8-10% van ganglioncellen in menselijk netvlies
→ M- & P-ganglioncellen vormen samen ± 80% van alle ganglioncellen
andere ganglioncellen = koniocellulaire cellen
projecteren naar koniocellulaire lagen in LGN
sommige van deze (met input van S-kegeltjes) maken deel uit van ‘primordiale’ blauw-geel route
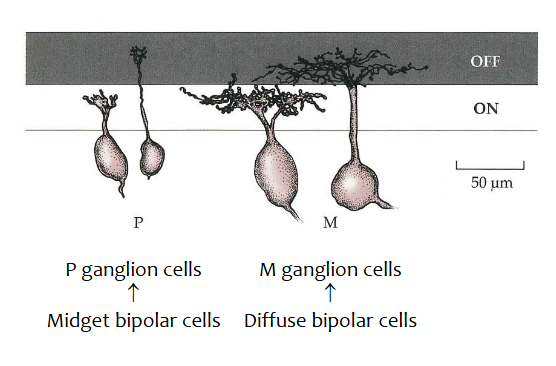
center-surround receptieve velden → experiment van Kuffler
kleine elektroden gebruikt om elektrische veranderingen in individuele ganglioncellen te bestuderen
ganglioncellen: vuren spontaan actiepotentialen af (± 1 spike per seconde), zelfs bij afwezigheid van visuele stimulatie
elke ganglioncel: klein venster op de wereld = receptieve veld
receptieve veld
= gebied op netvlies (en corresponderende gebied in visuele ruimte) waarin visuele stimuli de vuurfrequentie van neuronen beïnvloeden
invloed kan exciterend zijn of inhiberend
exciterend: vuurfrequentie van ganglion verhogen
inhiberend: vuurfrequentie van ganglion verlagen
experiment van Kuffler:
visuele stimulus: klein lichtpuntje op projectiescherm, dat over netvlies bewoog en aan & uit ging
terwijl impulsen van enkele retinale ganglioncel geregistreerd
wanneer vlek op specifiek klein gebied van netvlies werd geplaatst → ganglioncel verhoogde vuursnelheid wanneer licht werd aangezet
gebied = center van het receptieve veld van de ganglioncel
wanneer vlek naar aangrenzend gebied van netvlies werd verplaatst → ganglioncel verlaagde vuursnelheid wanneer licht werd aangezet
→ is het geval voor een ON-center cel
ON-center cel:
vuursnelheid neemt toe wanneer licht in midden van receptieve veld wordt aangezet
vuursnelheid neemt af wanneer licht in surround wordt aangezet
OFF-center cel:
vuursnelheid neemt af wanneer licht op plek in center van receptieve veld word aangezet
vuursnelheid neemt toe wanneer licht op plek in surround wordt aangezet
→ meeste retinale ganglioncellen hebben 1 van de 2 typen concentrische center-surround organisatie
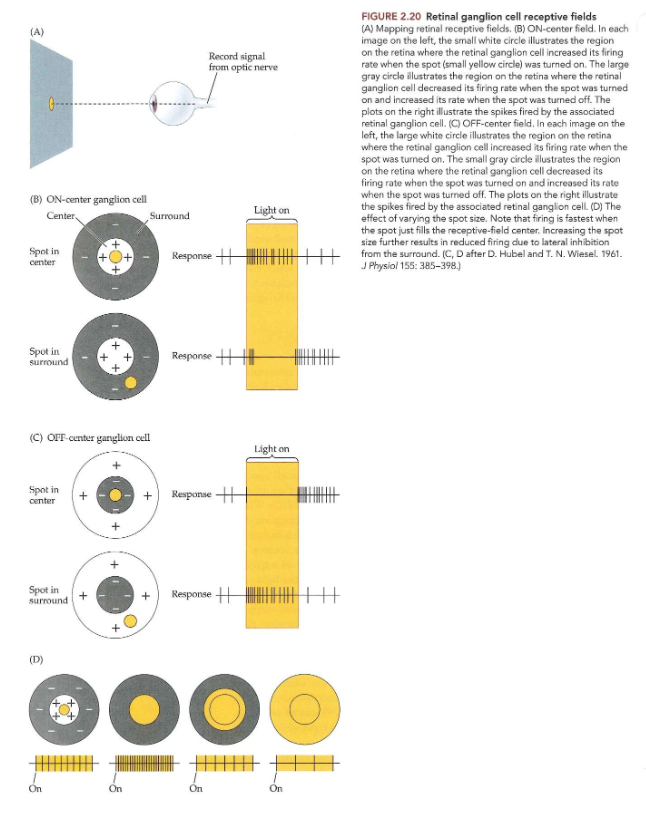
center-surround receptieve velden → ruimtelijke indeling van receptieve veld van ganglioncel is concentrisch
klein cirkelvormig gebied in center reageert op toename van belichting
omringende ring reageert op afname van belichting
→ ganglioncel vuurt het snelst wanneer grootte van lichtvlek overeenkomt met grootte van exciterende ‘center’
→ ganglioncel vermindert vuursnelheid wanneer lichtvlek zijn remmende ‘surround’ begint te bereiken
antagonistische interactie tussen ‘center’ & ‘surround’
= laterale inhibitie
deels gemedieerd door horizontale cellen
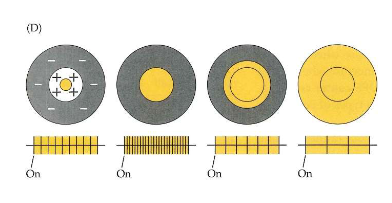
center-surround receptieve velden → 2 belangrijke functionele gevolgen
elke ganglioncel reageert het best op vlekken van een bepaalde grootte (& minder op vlekken die groter of kleiner zijn)
retinale ganglioncellen werken zo als filter:
best reageren op stimuli die precies juiste grootte hebben
minder reageren op stimuli die groter of kleiner zijn
ganglioncellen zijn meest gevoelig voor verschillen in intensiteit van licht in center & surround, en minder beïnvloed door gemiddelde intensiteit van het licht
nuttige eigenschap want: gemiddelde intensiteit van licht dat op netvlies valt varieert sterk, afhankelijk of men binnen of buiten is, of het dag of nacht is, enzovoort
MAAR: contrast (verschil in luminantie/ helderheid tussen aangrenzend delen van scène) zal ongeveer hetzelfde zijn, ongeacht lichtomstandigheden
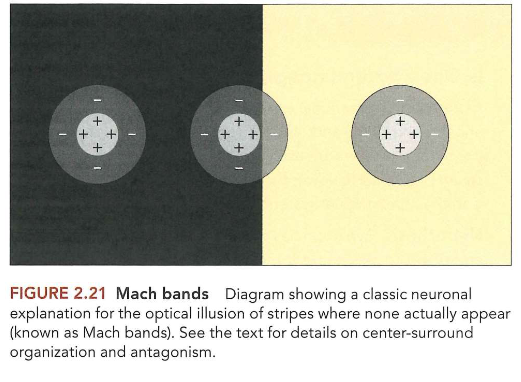
center-surround receptieve velden → Mach banden
center-surround antagonisme = laterale inhibitie → perceptie-effecten wat resulteert in illusie van strepen & vlekken
wat we denken: ogen werken als videocamera’s & leggen getrouwe momentopnamen vast van wereld om ons heen
MAAR: rest van visuele systeem ziet alleen wat retinale ganglioncellen het laten zien en ganglioncellen zijn niet tevreden met simpelweg doorgeven van ruwe beelden die door fotoreceptoren gecodeerd zijn
→ ganglioncellen fungeren, samen met bipolaire, amacriene & horizontale cellen, als beeldfilter
transformeren ruwe beeld tot nieuwe representatie
nieuwe representatie benadrukt bepaalde belangrijke informatie (bv. contrast) & negeert grotendeels andere soorten minder bruikbare informatie (bv. intensiteit van omgevingslicht)
→ visuele systeem: te beschouwen als lange reeks filters, waarbij elke fase in systeem verantwoordelijk is voor extraheren van bepaald aspect van visuele wereld & doorgeven hiervan aan volgende fase
P- & M-ganglioncellen herbekeken
→ receptieve velden verschillen op aantal belangrijke punten:
P-cellen kleinere receptieve velden dan M-ganglioncellen
reden: grootte van receptieve veld wordt bepaald door grootte van dendritische veld & type bipolaire cellen waarmee het verbonden is
M-cellen luisteren naar meer fotoreceptoren dan P-cellen → M-cellen reageren op groter deel van gezichtsveld
M-cellen veel gevoeliger onder omstandigheden met weinig licht
beter in staat om visuele stimuli te detecteren
P-cellen fijnere resolutie (mits voldoende licht)
P- & M-ganglioncellen verschillen ook in hun temporele reacties
P-cellen reageren met veranderingen in aanhoudende activiteit terwijl licht op exciterende gebieden schijnt (contrast)
geven info door over contrast in netvliesbeeld
M-cellen reageren meer tijdelijk (verandering beeld)
M-cel reageert met korte impulsstoot wanneer lichtbron wordt ingeschakeld, en keert dan snel terug naar spontane frequentie (zelfs al blijft lichtbron branden)
geven info door over hoe beeld in loop van de tijd verandert
P- & M-ganglioncellen verschillen in doorgeven kleurinformatie
intrinsiek fotogevoelige retinale ganglioncellen in ontwikkelende netvlies
melanopsin-bevattende retinale ganglioncellen reageren op licht, maar ontvangen geen input van staafjes of kegeltjes
1e fotoreceptoren die rijpen in netvlies & daarom eerste signalen obv licht naar zich ontwikkelende hersenen sturen (reeds in 2e trimester)
→ baby’s in baarmoeder kunnen dus licht detecteren, lang voordat ze beelden kunnen zien
van stippen naar strepen:
lagen van netvlies vertalen het ruwe licht dat door fotoreceptoren wordt opgevangen naar patronen van lichtstippen omgeven door duister, of omgekeerd, die worden gedetecteerd door ganglioncellen
netvlies laat ons toe om patroon van lichte & donkere gebieden in gezichtsveld waar te nemen, ongeacht algehele lichtniveau (bv. in schemering net zo goed kunnen zien als op middaguur)
beeldverwerking van oogbol naar hersenen
neuronen in primaire visuele hersenschors geven voorkeur aan lijnen, randen & gestreepte patronen
visuele cortex georganiseerd in duizenden minicomputers → elk verantwoordelijk voor bepalen van oriëntatie, breedte, kleur & andere kenmerken van strepen in klein deel van gezichtsveld
gezichtsscherpte
nucleus geniculatus lateralis
primaire visuele cortex
receptieve velden in primair visuele cortex
kolommen & hyperkolommen
van stippen naar strepen: gezichtsscherpte & contrast
contrast
= verschil in luminantie tussen object & achtergrond, of tussen lichtere & donkerdere delen van hetzelfde object
gezichtsscherpte
= kleinste ruimtelijke detail dat kan worden waargenomen bij 100% contrast
→ op bepaald moment: strepen niet meer afhankelijk van elkaar te zien (staat in relatie tot gezichtsscherpte)
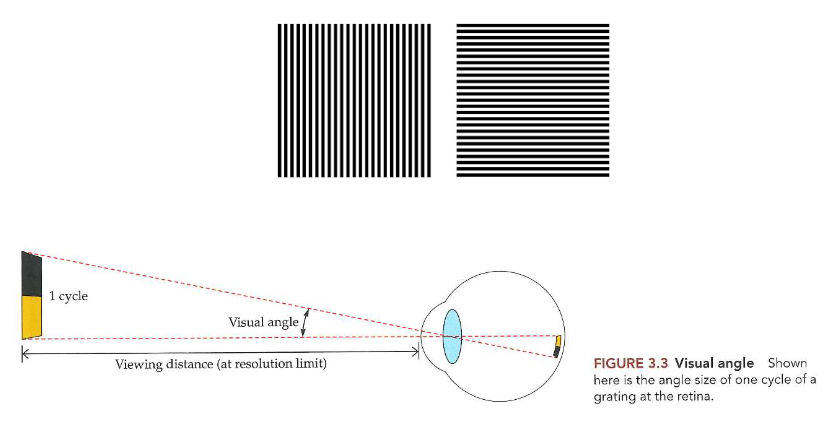
kleinste visuele hoek waarmee we 1 cyclus kunnen waarnemen
1 cyclus: 1 herhaling van een zwarte & witte streep
visuele hoek: hoek die gevormd wordt door lijnen gaande van boven naar onder (of van links naar rechts, afhankelijk van oriëntatie van strepen) van 1 cyclus op de pagina, die door midden van lens gaan & eindigen op netvlies
onder ideale omstandigheden: onderscheiden wanneer 1 cyclus een hoek van ongeveer 0,017 graden (1 boogminuut) beslaat
→ resolutiescherpte vertegenwoordigt een fundamentele spatiale beperking van het zicht:
fijnste details met hoog contrast die kunnen worden waargenomen
limiet primair bepaald door afstand tussen fotoreceptoren in het netvlies
van stippen naar strepen: gezichtsscherpte & contrast → rasterpatronen
sinusgolfroosters projecteren op netvlies (A)
lichtintensiteit varieert geleidelijk & continu gedurende elke cyclus
bij B: intensiteit verandert abrupt van zwart naar wit & terug naar zwart
→ receptoren zo verdeeld dat witste & zwartste delen van rooster op afzonderlijke kegels vallen (C): we zien strepen
→ als hele cyclus op 1 kegel valt (D): we zien een grijs veld

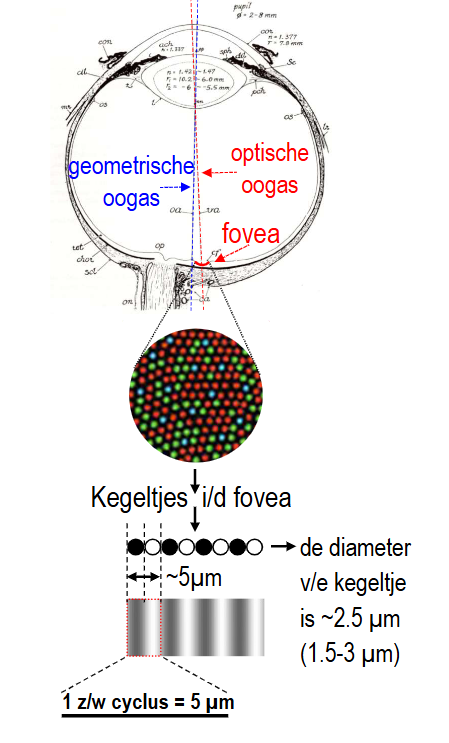
van stippen naar strepen: gezichtsscherpte & contrast → optische vs. geometrische oogas
geometrische as (blauw)
optische as (rood)
iets schuiner, door de fovea
functioneel belangrijker als referentie
fovea:
enkel kegeltjes (rood-groen-blauw)
diameter van kegeltje thv fovea: ± 2,5 micrometer
→ aanbieden van zwart-wit patroon:
zwart-wit cyclus moet 5 micrometer zijn om die nog te kunnen onderscheiden (donker deel valt dan op ‘zwarte’ kegeltje & bleke deel op ‘witte’ kegeltje)
center van kegeltjes in fovea: ligt op afstand van ± 0,008 graden (= halve boogminuut) → past goed bij waargenomen gezichtsscherptelimiet van 0,017 graden (want 2 kegeltjes per cyclus nodig om strepen nauwkeurig te kunnen waarnemen)
in periferie: staafjes dichter op elkaar (denser) dan kegeltjes, & hoge graad van convergentie waardoor gezichtsscherpte in periferie veel slechter is dan in fovea
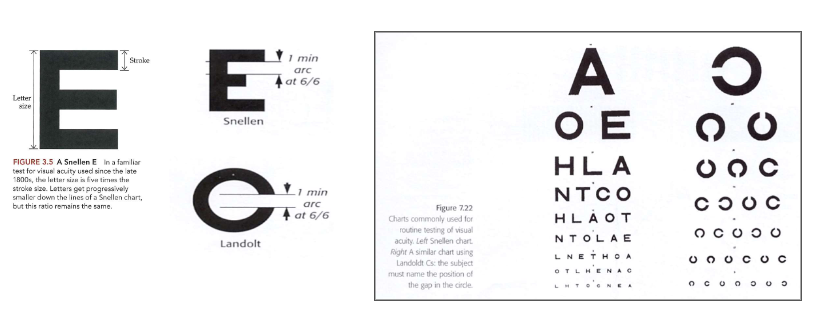
van stippen naar strepen: gezichtsscherpte & contrast → visuskaarten: Snellen & Landolt C’s
beschrijving van gezichtsscherpte bij oogartsen
letters lezen → steeds kleiner worden tot u meerdere fouten maakt
gezichtsscherpte 6/6 als u goed ziet
methode:
set blokletters waarbij letter als geheel 5x zo groot is als componenten waaruit letter bestaat
gebruik maken van charts bij testen van visuele resolutie
kijker bevindt zich op constante afstand van 6 meter, dan letters van afnemende grootte weergegeven
gezichtsscherpte uitgedrukt in breuk
6/6 (= 100% visus)
persoon ziet op 6 meter afstand even goed als iemand met normaal zicht
visuele hoek → 6/6-letter: bestrijkt hoek van 5 boogminuten (0,083 graden) op oog
elke streep van een 6/6-letter: bestrijkt hoek van 1 boogminuut (= 0,017 graden)
dus 6/6-letter kunnen lezen → details te onderscheiden die 1 boogminuut bestrijken
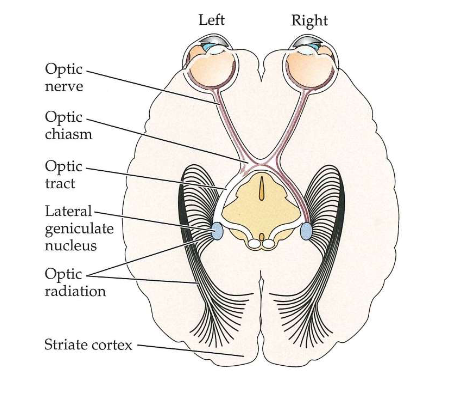
van stippen naar strepen: nucleus geniculatus lateralis (LGN)
LGN
structuur in thalamus (onderdeel van middenhersenen)
fungeert als verbindingsstation op weg van netvlies naar cortex
axonen van retinale ganglioncellen maken synapsen in 2 LGNs (1 in elke hersenhelft)
LGN heeft input- & outputverbindingen met visuele cortex
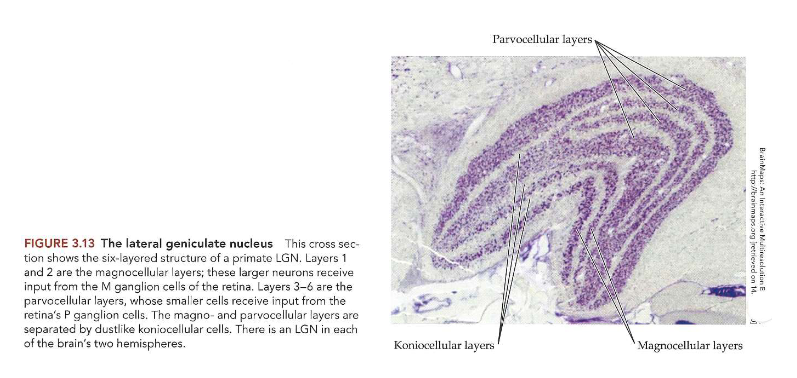
van stippen naar strepen: nucleus geniculatus lateralis (LGN) → structuur
6-lagige structuur, in het midden gebogen
neuronen in onderste 2 lagen zijn fysiek groter dan die in bovenste 4 lagen
onderste 2 lagen → magnocellulaire lagen
ontvangen input van M-ganglioncellen in netvlies
bovenste 4 lagen → parvocellulaire lagen
ontvangen input van P-ganglioncellen
magno- & parvocellulaire lagen: van elkaar gescheiden door koniocellulaire lagen
betrokken bij doorgeven van signalen van S-kegeltjes
kunnen deel uitmaken van primordiale blauw-gele pathway
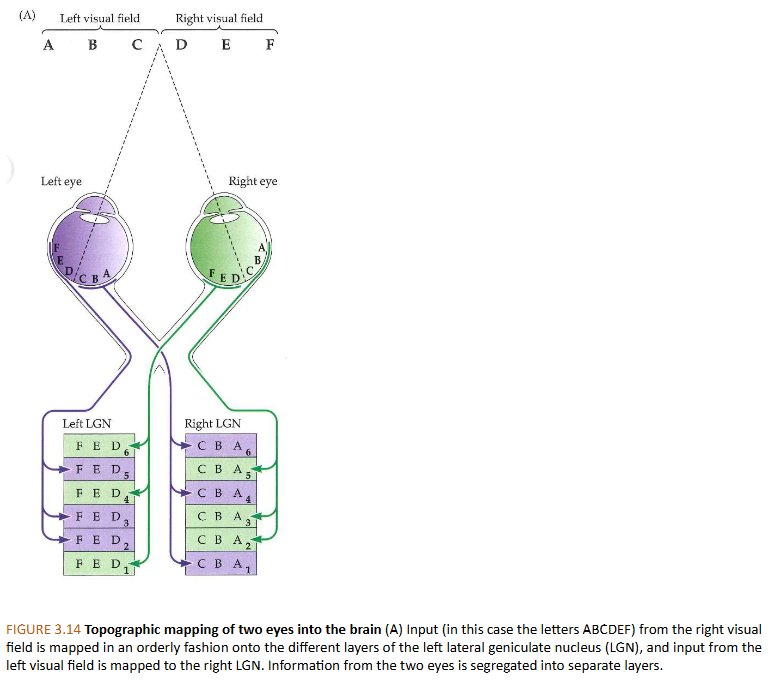
van stippen naar strepen: nucleus geniculatus lateralis (LGN) → organisatie van retinale input naar LGNs
linker LGN ontvangt projecties van linkerkant van netvlies in beide ogen
rechter LGN ontvangt projecties van rechterkant van beide netvliezen
elke laag van LGN input ontvangt van het ene of andere oog:
van onder naar boven:
lagen 1,4 & 6 van rechter LGN ontvangen input van linker (contralaterale) oog
lagen 2,3 & 5 van rechter LGN ontvangen input van rechter (ipsilaterale) oog
→ informatie van de 2 ogen worden dus gescheiden in verschillende lagen in LGN
elke LGN-laag: bevat zeer georganiseerde kaart van complete helft van het gezichtsveld
= topografische mapping
hierdoor hebben we een neurale basis om te weten waar dingen zich in ruimte bevinden
LGN-neuronen: concentrische receptieve velden
lijken sterk op die van retinale ganglioncellen
reageren op dezelfde patronen als ganglioncellen die hun input leveren
waarom lopen axonen van ganglioncellen niet gewoon rechtstreeks over naar de hersenschors?
LGN niet slechts een halte op lijn van netvlies naar cortex
MAAR: veel verbindingen tussen andere delen van hersenen en LGN → input vanuit ogen kan daardoor worden gemoduleerd
LGN → deel van thalamus
bij slapen: thalamus wordt geremd
dus: zelfs als oogleden open waren tijdens slaap → je zou niets zien in schemerige kamer omdat input van netvliezen nooit cortex zou bereiken
want neurale signalen zouden stoppen in LGNs
→ MAAR: thalamische inhibitie is niet volledig:
bij fel licht → wordt wel waargenomen → maakt je wakker
van stippen naar strepen: primaire visuele cortex
primaire visuele cortex = V1 = Brodmann area 17 = striate cortex
uit 6 hoofdlagen
sommige hoofdlagen hebben sublagen
vezels van LGN → projecteren voornamelijk naar laag IVC
magnocellulaire axonen: naar bovenste deel van laag IVC (= IVCα)
parvocellulaire axonen: naar onderste deel van laag IVC (= IVCβ)
→ door massale input vanuit LGN in laag IV in V1: bijzonder breed
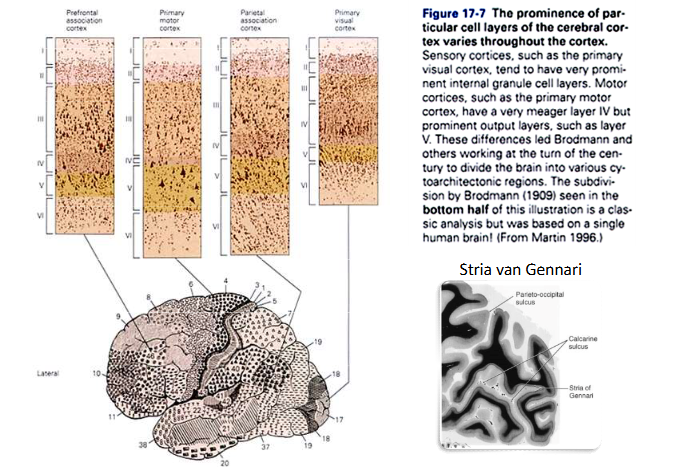
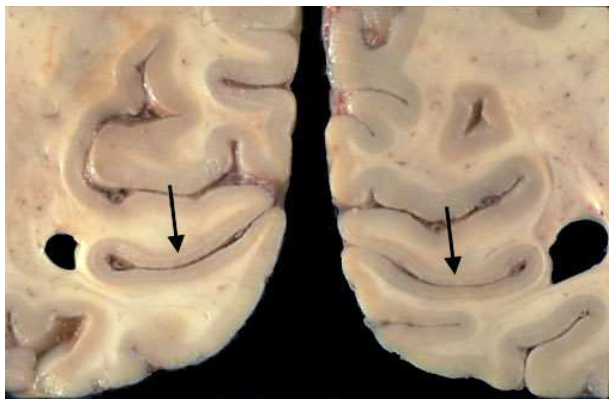
van stippen naar strepen: primaire visuele cortex → sulcus calcarinus
massale aankomst van gemyeliniseerde axonen vanuit LGN naar laag IV van primaire visuele cortex (langsheen sulcus calcarinus)
parallel aan oppervlakte van cortex, in occipitale kwab
→ streping met blote oog zichtbaar
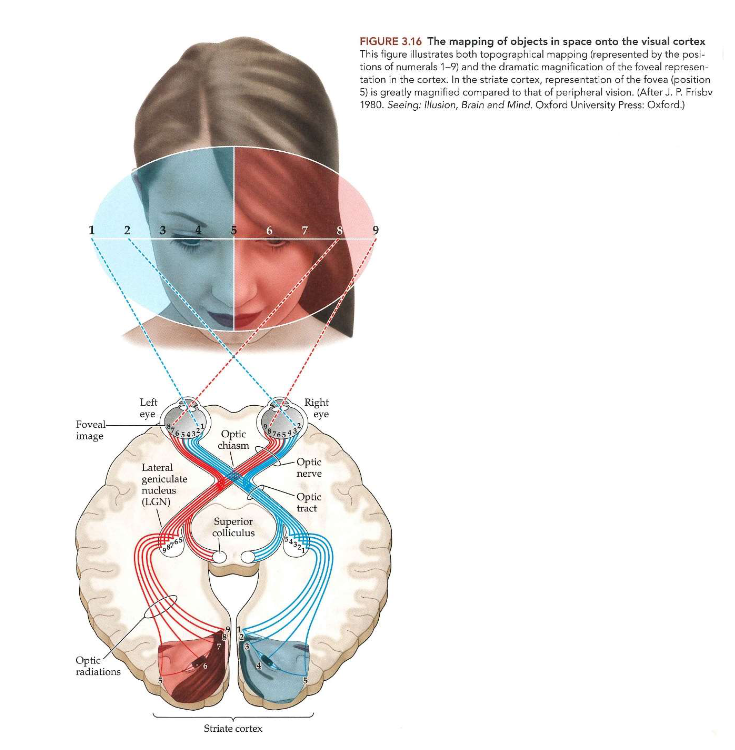
van stippen naar strepen: primaire visuele cortex → topografie van de primaire visuele cortex
thv van primaire visuele cortex → systematische topografische weergave van gezichtsveld & complexe transformatie van visuele informatie
V1 bevat meer dan 100x zoveel cellen als LGN
2 belangrijke kenmerken van visuele cortex:
topografie
topografische mapping
vergroting
informatie vanuit verschillende delen van gezichtsveld worden drastisch geschaald
objecten die worden afgebeeld op of nabij fovea → worden verwerkt door neuronen in groot deel van V1
objecten die worden afgebeeld in uiterst rechtse of linkse periferie → worden slechts toegewezen aan klein deel van V1
→ vervorming van visuele-veldkaart op cortex = corticale vergroting
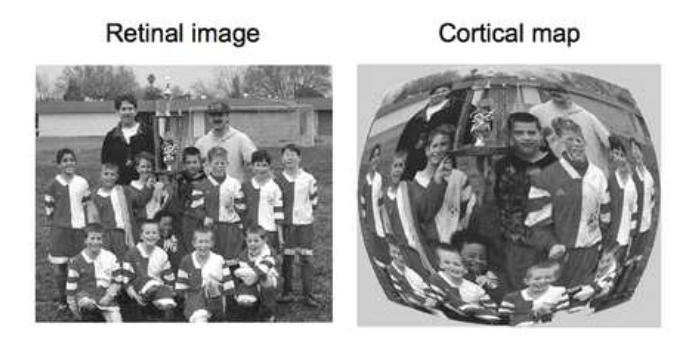
corticale vergroting thv primaire visuele cortex
→ gevolg:
gezichtsscherpte neemt af met excentriciteit (= hoekafstand tov fovea)
wanneer we details in perifeer gezichtsveld willen zien → we moeten ogen/ hoofd draaien zodat object op fovea valt
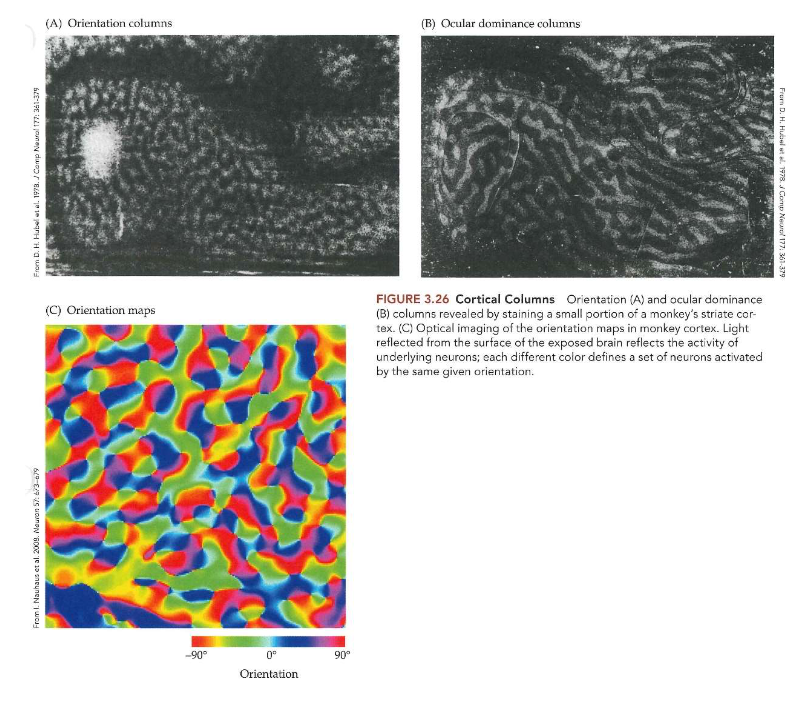
van stippen naar strepen: receptieve velden in primair visuele cortex → Hubel & Wiesel
ontdekten dat receptieve velden van neuronen in V1 (bij kat) niet circulair zijn (zoals die van retinale ganglioncellen), maar langwerpig
reageren op stimuli als lijn of rand, streep & raster
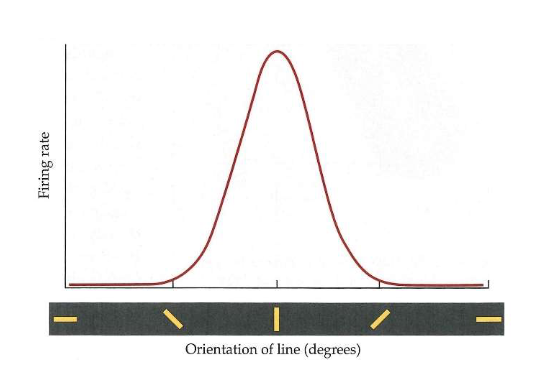
van stippen naar strepen: receptieve velden in primair visuele cortex → orientation tuning
individueel neuron reageert niet op willekeurige streep in receptief veld
beste reageren wanneer lijn of rand precies de juiste oriëntatie heeft
reageert nog nauwelijks wanneer lijn meer dan 30 graden van optimale oriëntatie afwijkt (verandering die gelijk is aan verplaatsing van wijzer van klok van 12 naar 1)
→ selectieve responsiviteit = orientation tuning
cel is afgestemd om lijnen in specifieke oriëntatie te detecteren
populatie neuronen als geheel afgestemd om alle mogelijke oriëntaties te detecteren
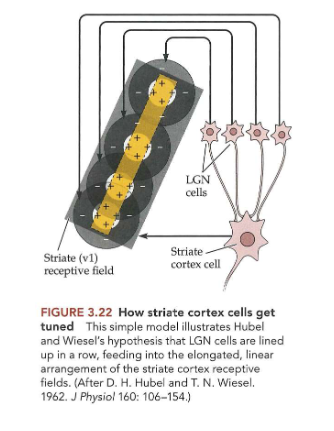
hoe worden circulaire receptieve velden in LGN getransformeerd tot langwerpige receptieve velden in gestreepte cortex?
→ LGN-cellen die input geven aan corticale cel: staan in een rij
circulaire receptieve velden zo samengevoegd tot lineair receptief veld
→ corticale cellen reageren ook op strepen/ rasters
individuele cellen afgestemd op streep/ raster met welbepaalde spatiale frequentie
→ zorgt dat elk van die neuronen in V1 functioneert als filter voor deel van beeld dat cel activeert
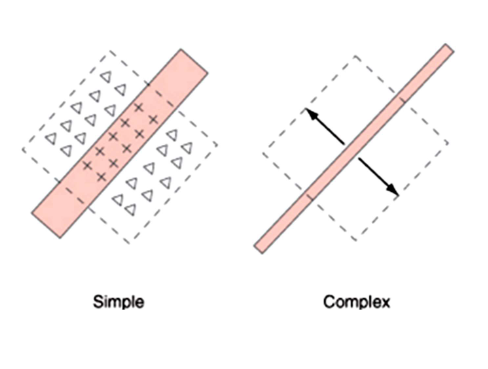
van stippen naar strepen: receptieve velden in primair visuele cortex → ‘simple cells’ & ‘complex cells’
simple cells:
corticale neuronen waarvan receptieve velden duidelijk gedefinieerde exciterende & inhiberende gebieden hebben
detector links op figuur: reageert best op lichtstreep (aan beide kanten omgeven door duisternis) met welbepaalde breedte & oriëntatie
complex cells:
vertonen reacties die niet eenvoudigweg kunnen worden voorspeld obv reacties op stationaire lichtstrepen
corticale neuronen waarvan receptieve velden geen duidelijk gedefinieerde exciterende & inhiberende gebieden hebben
elke complexe cel is afgestemd op bepaalde oriëntatie & spatiale frequentie (zoals simple cell)
MAAR: complexe cel reageert ongeacht waar streep wordt gepresenteerd, zolang deze ergens binnen receptieve veld van cel valt
anders dan simple cell: deze reageert alleen als streep in midden van zijn receptieve veld wordt gepresenteerd
van stippen naar strepen: kolommen & hyperkolommen → oriëntatievoorkeur (& oculaire dominantie)
elk van ± 200 miljoen neuronen in V1 reageert op reeks stimuluseigenschappen:
lijnen, randen, strepen en/of roosters
in een bepaalde hoek georiënteerd
met bepaalde breedte of ruimtelijke frequentie
op een bepaalde manier bewegen
→ niet lukraak verspreid over V1
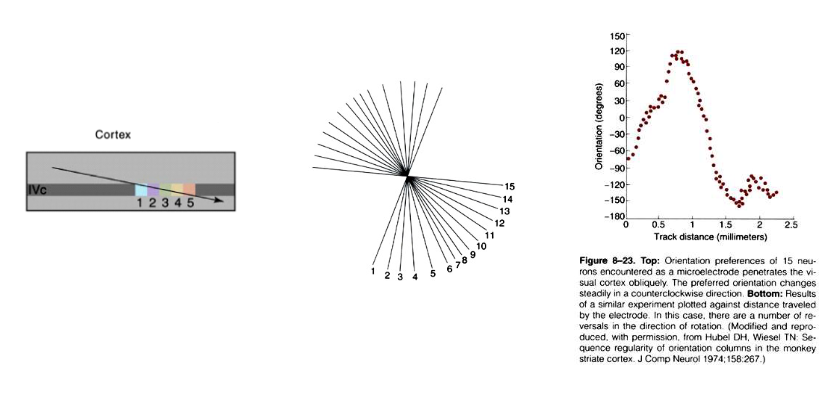
ontdekking:
registratie-elektrode door lagen van cortex duwen in richting loodrecht op corticale oppervlak → alle cellen die ze tegenkwamen vertoonden vergelijkbare oriëntatievoorkeuren
elektrodepositie over kleine afstand verschoven & opnieuw loodrecht doordringen → alle cellen reageerden beste op een iets andere oriëntatie (10 of 15 graden tov oorspronkelijke oriëntatie)
→ conclusie: neuronen met vergelijkbare oriëntatievoorkeuren zijn gerangschikt in kolommen die zich verticaal door cortex uitstrekken
→ ander experiment: tangentiële penetraties in gestreepte cortex:
systematische & progressieve verandering in voorkeursoriëntatie
vrijwel alle oriëntaties binnen afstand van ± 0,5mm tegengekomen
→ oriëntatie niet enige eigenschap die in kolommen in visuele cortex is gerangschikt:
informatie van 2 ogen zijn gescheiden gehouden in LGN (elke LGN-cel reageert op ene of andere oog, maar nooit op beide ogen) → indeling in V1 verandert:
meeste cellen: beïnvloed door input van zowel linkeroog als rechteroog
neuronen in V1 reageren iets sterker wanneer stimulus in ene oog wordt gepresenteerd dan wanneer deze in andere oog wordt gepresenteerd
= oculaire dominantie
neuronen die dezelfde oogvoorkeur delen (en oculaire dominantie vertonen) → kolomvormige rangschikking
oogvoorkeur verandert zowat elke 0,5mm
van stippen naar strepen: kolommen & hyperkolommen → hyperkolommen
→ architectuur van primaire visuele cortex
blok van 1mm V1 → ‘all the machinery necessary to look after everything the visual cortex is responsible for, in a certain small part of the visual world’
= hyperkolom
bevat ten minste 2 sets kolommen, die beide elke mogelijke oriëntatie (0-180 graden) bestrijken, waarbij 1 set de voorkeur geeft aan input van linkeroog & 1 set aan input van rechteroog
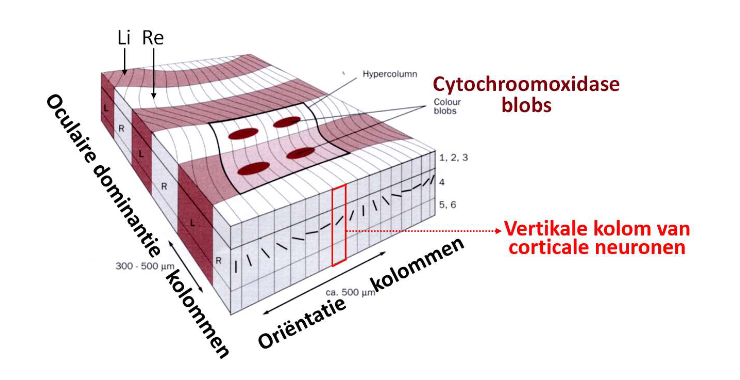
van stippen naar strepen: kolommen & hyperkolommen → hyperkolommen (realistischer beeld)
van stippen naar strepen: kolommen & hyperkolommen → cytochroom oxidase blobs
→ oriëntatie & oculaire dominantie: niet enige stimulusdimensie met systematische kolomvormige rangschikking in visuele cortex
kleuringstechniek → gebruikmakend van enzym cytochroomoxidase (CO)
toont regelmatige reeks CO-blobs, met afstand van ± 0,5mm
functionele rol: nog onduidelijk, maar CO-blobkolommen betrokken bij verwerking van kleur
interblobgebieden: verwerking van beweging & oriëntatie

van stippen naar strepen: gevolgen van mono-oculaire deprivatie
veranderingen in corticale fysiologie → resulteert in verwoestend & permanent verlies van ruimtelijk zicht
kritieke periode in vroege visuele ontwikkeling waarin normale binoculaire visuele stimulatie nodig is voor normale corticale ontwikkeling:
bij mensen: eerste 3-8 jaar
tijdens deze periode: corticale neuronen worden nog steeds verbonden met input van de 2 ogen
periode van neurale plasticiteit → abnormale visuele ervaring kan normale neurale bedradingsproces veranderen
indien 1 oog geen normale stimulatie ontvangt → neuronen die op dat oog zouden moeten reageren, worden niet goed geconnecteerd
bv. cataract onbehandeld tijdens kritieke periode → verkeerde corticale verbindingen kunnen nooit meer hersteld worden
amblyopie
= verminderde gezichtsscherpte in 1 oog door abnormale vroege visuele ervaring (lui oog) & gebrek aan binoculaire dieptewaarneming (stereopsis)
oorzaken:
strabisme (= scheelzien)
1 oog zo gedraaid dat het de wereld vanuit abnormale hoek bekijkt
anisometropie
2 ogen hebben zeer verschillende refractieafwijkingen (bv. ene oog is verziend & andere niet)
→ klinische behandeling: plakken van pleister over goede oog & dwingen van amblyope oog om te werken
deze behandeling enkel uitgevoerd bij jonge kinderen (meestal jonger dan 8 jaar)

objecten waarnemen & herkennen:
→ naar de wereld kijken: geen reeks lijnen, randen, strepen & rasters MAAR samenhangende objecten & uitgestrekte oppervlakken (+ specifieke objecten herkennen)
van eenvoudige lijnen & randen tot eigenschappen van objecten
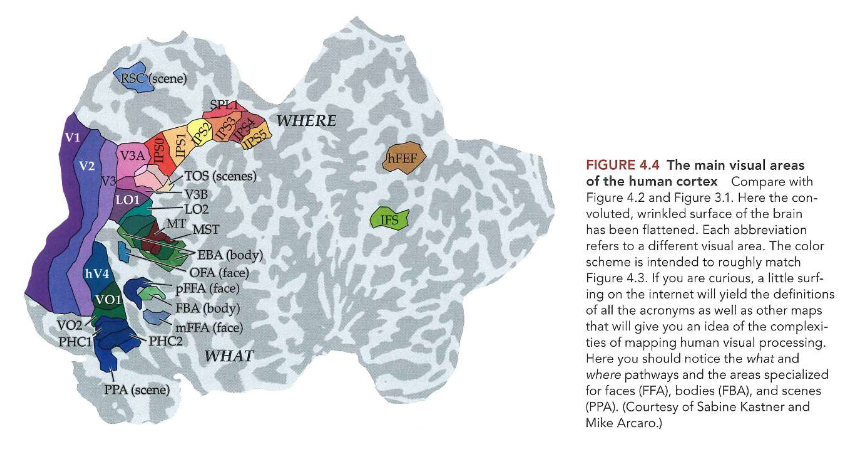
‘what’ & ‘where’ pathways
contouren & illusionaire contouren
object herkenning
objecten waarnemen & herkennen: van eenvoudige lijnen & randen tot eigenschappen van objecten
net voorbij V1: extrastriate cortex (bv. V2)
= verzameling visuele gebieden die net buiten primaire visuele cortex liggen
geen eenvoudige keten van verwerkingsgebieden
voor geavanceerde taken: bv. objectherkenning → verwerking nodig door groot aantal verschillende visuele verwerkingsgebieden
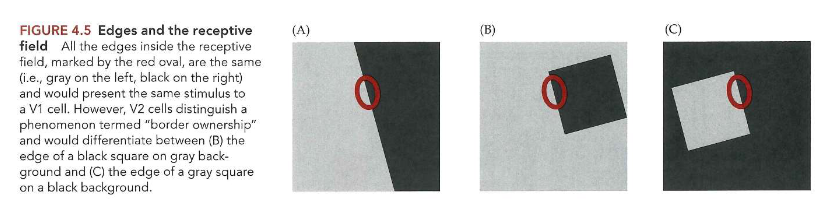
receptieve velden tonen interesse in eigenschappen die belangrijk zijn voor objectperceptie:
figuur: alles binnen receptieve veld (in rode ovaal) = hetzelfde (grijs links, zwart rechts) → zouden dus dezelfde stimulus aan V1-cel presenteren
MAAR: V2-cellen zorgen voor onderscheid tussen rand van zwart vierkant op grijze achtergrond & rand van grijs vierkant op zwarte achtergrond
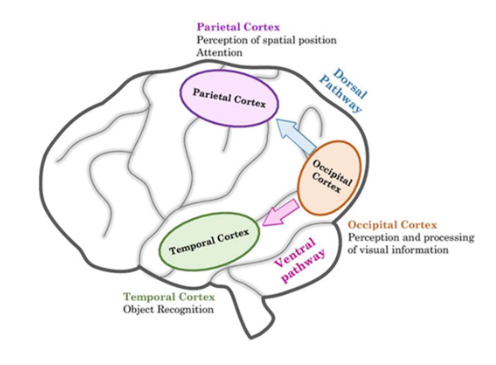
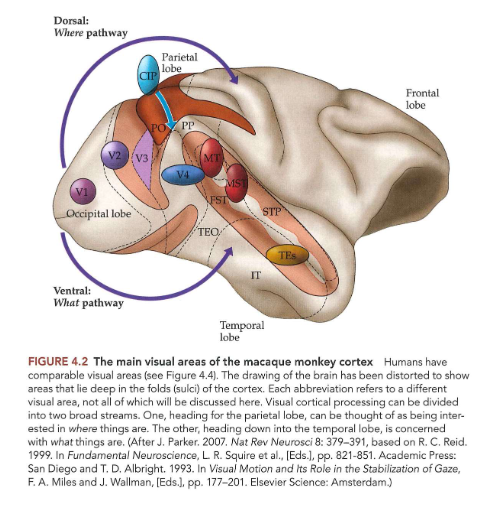
objecten waarnemen & herkennen: ‘what’ & ‘where’ pathways
vanuit extrastriate gebieden van occipitale kwab van hersenen → visuele informatie beweegt via 2 hoofroutes:
where-pathway = dorsale pathway
gaat omhoog (of dorsaal) → naar pariëtale kwab
voor verwerking van informatie met betrekking tot locatie van objecten in ruimte & handelingen die nodig zijn om ermee te interageren (bewegen van handen, ogen, enz.)
belangrijk bij instellen van aandacht
what-pathway = ventrale pathway
gaat omlaag (ventraal) → naar temporaalkwab
belangrijk voor objectherkenning
→ nuttig concept, MAAR: scheiding tussen pathways zijn verre van absoluut
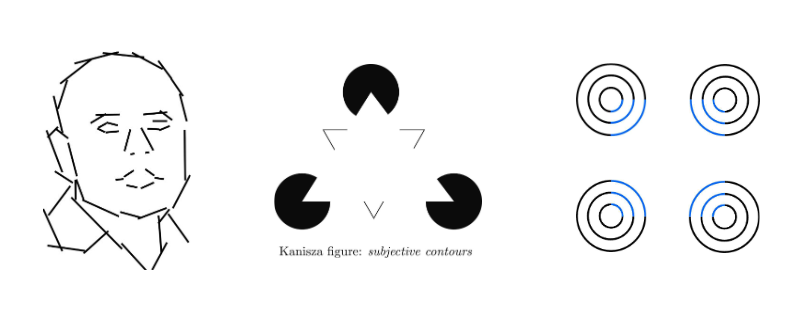
objecten waarnemen & herkennen: contouren & illusoire contouren
V1 kan informatie over object contouren doorsturen naar hogere visuele areas
ontbreken van stuk van rand van object lijkt visuele systeem niet te hinderen
verschillende stukjes informatie worden gecombineerd om visuele systeem inschatting te laten maken over aanwezigheid & betekenis van contour
inferentiële aard van contourperceptie → Kanizsa-figuur:
driehoek niet getekend, maar wordt toch gezien
wordt gesuggereerd door hoeken die uit cirkels weggenomen zijn, terwijl eigenlijk geen grens tussen witte figuur & witte achtergrond
→ illusoire contour vermoedelijk in V2 gegenereerd als beste schatting van visuele systeem over wat er op die locatie in wereld gebeurt
objecten waarnemen & herkennen: object herkenning
niet geweten wat individuele cellen optimaal activeert terwijl we via de what-pathway de temporaalkwab ingaan
fMRI (= functionele beeldvormingsstudies): toont dat verschillende delen van cortex beter worden geactiveerd door bepaalde categorieën stimuli dan door andere
bewijs voor verband tussen temporaalkwab & objectherkenning
studies: bij apen waar grote delen van temporaalkwab werden beschadigd (chirurgisch verwijderd)
apen gedroegen zich alsof ze konden zien, maar niet wisten wat ze zagen
= agnosie
later onderzoek → bewijs dat
deel van temporaalkwab (inferiotemporale (IT) cortex): belangrijk voor visuele problemen van die apen
registraties van afzonderlijke cellen in IT gebied:
cellen in IT-cortex: receptieve velden die zich over helft of meer van gezichtsveld van aap konden verspreiden
soorten stimuli die IT-cellen activeerden
grandmother cell
= elke cel die selectief lijkt te reageren op 1 specifiek object
IT-cortex: nauwe verbindingen met hippocampus (betrokken bij geheugenvorming) → belangrijk want IT-cellen moeten receptieve-veldeigenschappen leren
neuronen die reageren op grootmoeders → kunnen niet hardwired zijn
want ieders grootmoeder is anders
→ cellen in IT-cortex vertonen plasticiteit
anatomie van hersenen van mensen & makaken: niet identiek
gebieden met vergelijkbare rollen = homologe gebieden
menselijke cortex → gebieden die homoloog lijken te zijn met IT-cortex & hippocampus van apen
agnosie
= je kan zien, maar je weet niet wat je ziet
bv. door beroerte
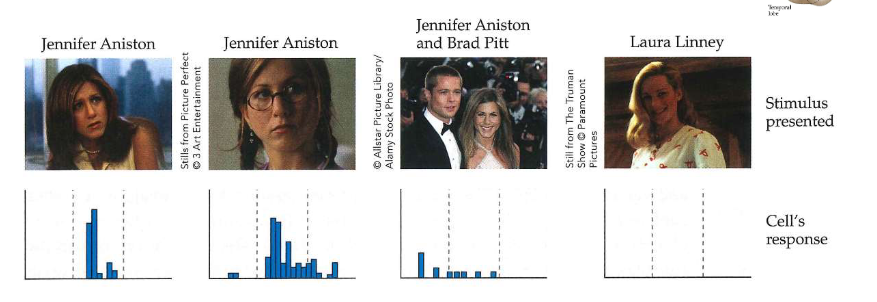
Jennifer Aniston cel
registraties van afzonderlijke cellen in temporaalkwab van menselijke waarnemers (implanteren van elektroden maakte deel uit van voorbereiding op epilepsiechirurgie)
experiment: waarnemer kijkt naar verzameling beeld terwijl activiteit van cel wordt gemonitord
→ foto: cel die alleen reageert op Jennifer Aniston
object herkenning: visuele verwerking bij de mens
niet veel systematische gegevens over reacties van individuele cellen in menselijke visuele systeem
wel groeiende hoeveelheid functionele beeldvormingsgegevens die gebieden in menselijke hersenen documenteren die gespecialiseerd lijken te zijn voor verschillende soorten stimuli
→ veel van die gebieden hebben naam gekregen voor specialisatie:
cellen in fusiforme face area (FFA)
voor gezichten
specifieke vorm van agnosie: prosopagnosie = onvermogen om gezichten te herkennen
cellen in fusiform body area (FBA)
voor andere lichaamsstructuren dan gezicht
parahippocampal place area (PPA)
cellen die reageren op ruimtes in wereld: bv. kamers met meubels erin
→ visuele systeem: modules die gespecialiseerd zijn in werken aan verschillende aspecten
visual word form area (WWFA)
gespecialiseerd in herkennen van geschreven woorden
pas wanneer iemand leert lezen
kleurperceptie
→ zicht werkt prima zonder kleur
→ vermogen om kleur te gebruiken: meerdere keren op verschillende manieren ontwikkeld in dierenrijk
basisprincipes van kleurwaarneming
kleurdetectie
kleurdiscriminatie
kleurwaarneming
individuele verschillen in kleurperceptie
van de kleur van licht naar een wereld van kleur
waar is kleurenzicht goed voor?

kleurperceptie: basisprincipes van kleurwaarneming
kleur: geen fysieke eigenschap van dingen in de wereld
kleur: resultaat van interactie van fysieke stimulus met bepaald zenuwstelsel
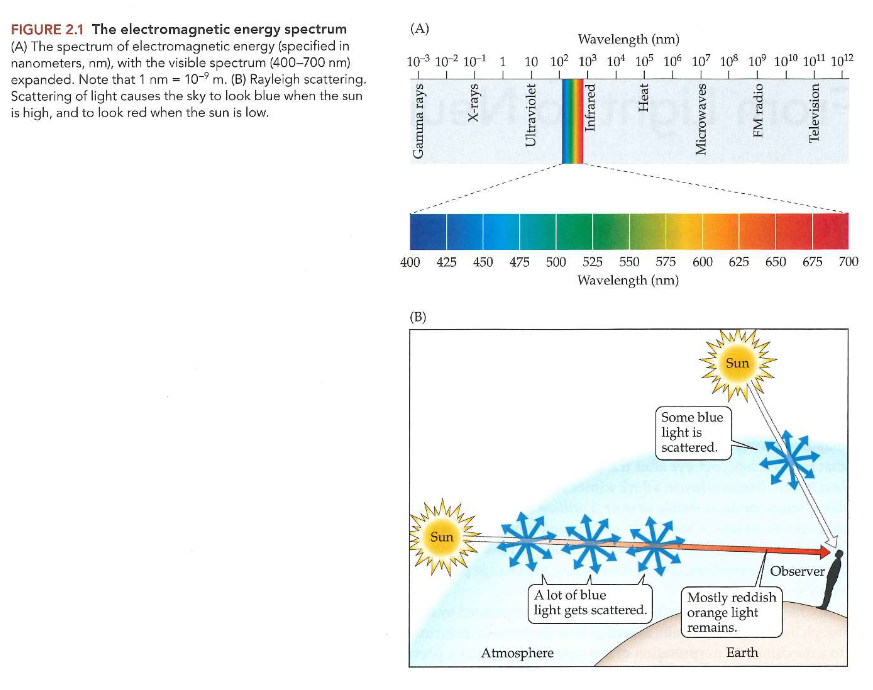
fout om te denken dat specifieke golflengten van licht specifieke kleuren zijn
smal bereik van elektromagnetische spectrum tussen golflengten te zien door mens
golflengten: van ± 400-700 nanometer (1nm = 10-9/ 0.000000001)
schijnbare kleur van deel van zichtbare wereld → gecorreleerd met golflengten van lichtstralen die oog bereiken vanuit deel van de wereld
meeste licht dat we zien = gereflecteerd licht
breed spectrum aan golflengten → raken oppervlakken in de wereld om ons heen:
uitgezonden door typische lichtbronnen: bv. zon of lamp
golflengten:
geabsorbeerd door oppervlakken die ze raken
gereflecteerd → deel van dat gereflecteerde licht bereikt ogen
kleur van oppervlak:
afhankelijk van combinatie van golflengten die oog bereiken vanaf oppervlak
kleurperceptie: kleurdetectie
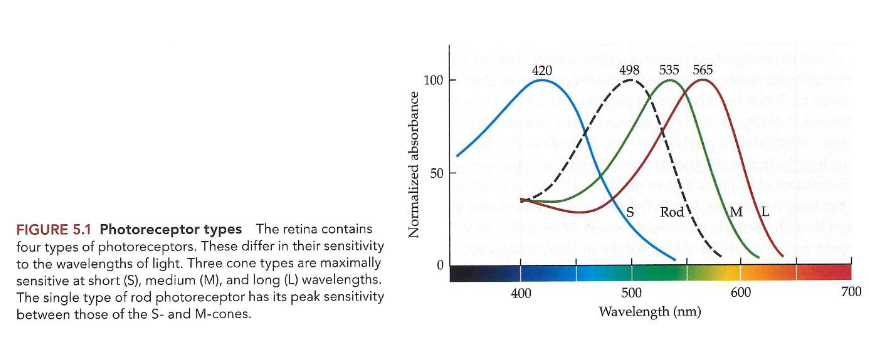
3 soorten kegeltjesfotoreceptoren → verschillen in fotopigment dat ze dragen
verschillen daarom in gevoeligheid voor licht van verschillende golflengten
elk kegeltype: vernoemd naar locatie van piek van gevoeligheid op spectrum
short-wavelength cones/ S-kegeltjes
piek bij ± 420nm
relatief zeldzaam
minder gevoelig dan M- & L-kegeltjes
medium-wavelength cones/ M-kegeltjes
piek bij ± 535nm
long-wavelength cones/ L-kegeltjes
piek bij ± 565nm
→ veel overlap tussen spectrale gevoeligheden van verschillende kegeltjes
→ combinatie van gevoeligheden van de 3 soorten kegels: geeft mogelijkheid om golflengtes van ± 400nm tot ± 700nm te detecteren
werking van kegels vs. staafjes
kegels → fotopische lichtniveaus (daglicht)
staafjes → scotopisch (zwakker) licht
1 type staafvormige fotoreceptor
iets ander gevoeligheidsprofiel
piek bij ± 500nm
kleurperceptie: kleurdiscriminatie → output van 1 enkele fotoreceptor
output van enkele fotoreceptor: volledig dubbelzinnig
output van enkele fotoreceptor op zichzelf → kan ons niets vertellen over golflengtes die hem stimuleren
menselijke visuele systeem heeft dat probleem opgelost, maar niet onder alle omstandigheden
1 type staafjesfotoreceptor → met hetzelfde type fotopigmentmolecuul: rodopsine
dus: allemaal dezelfde golflengtegevoeligheid
dus: enkel mogelijk om licht van donker te onderscheiden onder scotopische omstandigheden
MAAR: onmogelijk om ermee kleuren te onderscheiden
→ kleurenblindheid ‘s nachts: aanwijzing dat kleur psychofysisch is & niet fysiek
wereld die we zien onder heldere maan: niet fysiek ontdaan van kleur
dezelfde mix van golflengte die overdag kleurwaarneming produceert, blijft aanwezig op maanverlichte nacht → maar we zien geen kleuren onder zwakke lichtbronnen zoals maanlicht
want zwak licht stimuleert alleen staafjes & output van die fotoreceptorvariant maakt geen kleurenzicht mogelijk

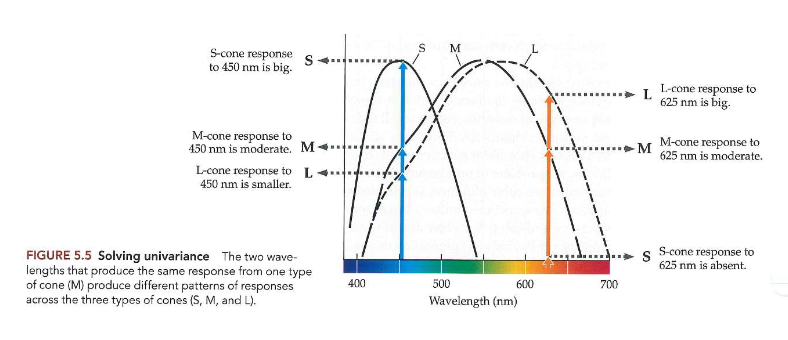
kleurperceptie: kleurdiscriminatie → oplossing van de univariance
verschillen tussen golflengten of combinaties van golflengten: precies detecteren → want we hebben meer dan 1 type kegelfotoreceptor
via 3 kegeltypes: verschil zien tussen licht van verschillende golflengten
reacties van 3 kegels op 2 golflengten: 450 & 625nm
zouden dezelfde reactie opleveren bij M-kegel (indien univariance)
MAAR: de 2 golflengten produceren verschillende outputs van L-kegels & S-kegels
= trichromatische theorie van kleurzicht
kleurperceptie: kleurdiscriminatie → R/Gr of B/Gl opponente kanalen
fovea: hoge densiteit aan kegels
belangrijk voor kleurenzicht
→ hersenen krijgen geen afzonderlijke signaal binnen (bv. rood of groen)
MAAR: vanuit retina wordt verschilsignaal gestuurd
kleuropponente kanalen;
rood-groen opponent kanaal
blauw-geel opponent kanaal
je hebt geen geelgevoelige fotoreceptoren/ kegels
luminantie kanaal: geeft info over lichtsterkte (M + L)
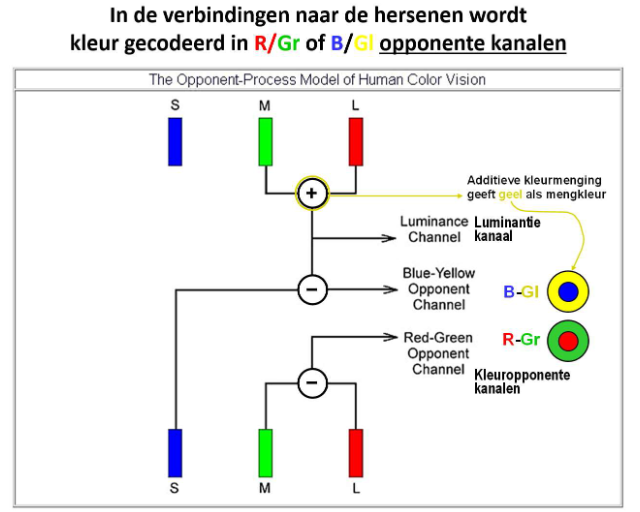
additieve kleurmenging
= mengen van gekleurd licht waarbij kleuren worden opgeteld
primaire kleuren:
rood (R)
groen (G)
blauw (B)
indien primaire kleuren licht in gelijke verhoudingen samenkomen → wit licht
geel = additieve kleur van groen & rood

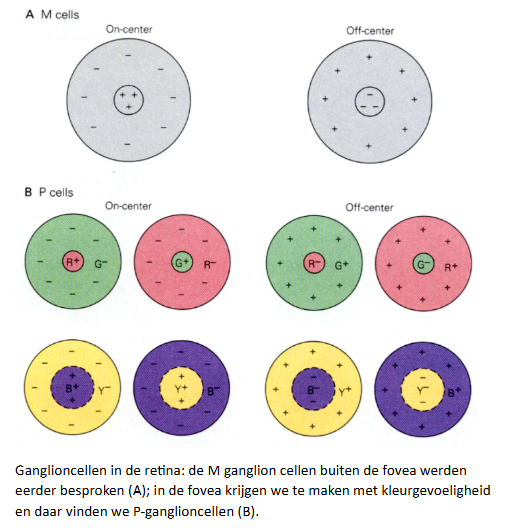
kleurperceptie: kleurdiscriminatie → ganglioncellen in de retina
veel ganglioncellen in netvlies & LGN van thalamus → maximaal gestimuleerd door lichtvlekken
ganglioncellen: receptieve velden met karakteristieke center-surround organisatie
M-ganglioncellen:
ON center:
geëxciteerd wanneer licht aangaat in centrale deel van receptieve velden
geïnhibeerd wanneer licht aangaat in surround
OFF center:
omgekeerde van ON center
→ vergelijkbare antagonistische relatie kenmerkt kleur
sommige van ON center retinale ganglioncellen & LGN cellen:
geëxciteerd door L-kegelaanzet in centrum
geïnhibeerd door M-kegelaanzet in surround
→ = 1 type kegel-opponente cel
maar ook:
M-L cellen
(M+L)-S cellen
S-(M+L) cellen
→ en geheel te herhalen voor OFF center retinale ganglioncellen
cellen die worden geëxciteerd door onset van licht:
(L+M)-cellen (~brightness)
→ feitelijke fysiologie = behoorlijk ingewikkeld
S-kegelsignalen gaan bv. door koniocellulaire lagen in LGN, terwijl M- & L-kegel-antagonistische signalen in parvocellulaire lagen worden aangetroffen
(we bespreken slechts 1 soort receptief veld ON P-ganglion cel met roodgevoelig center & groengevoelig surround)
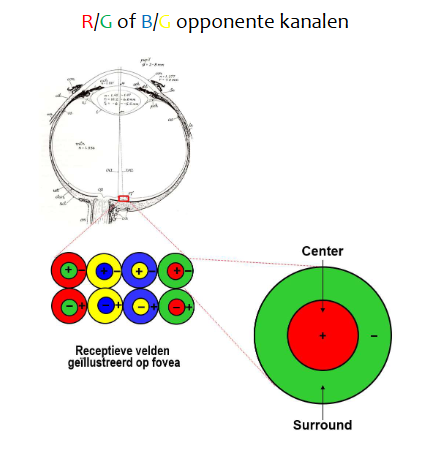
kleurperceptie: kleurdiscriminatie → schematische weergave van receptieve velden in fovea (overlap niet getekend voor duidelijkheid)
optische as: recht door fovea
thv fovea: P-type ganglioncellen
kegeltje connecteert met kleine bipolaire cel (midget bipolar cell) → connecteert dan met P-type ganglioncel
ON & OFF P-type ganglioncellen
organisatie in fovea: zorgt voor scherp zicht (kleinere receptieve velden, fijnere resolutie)
op center rood licht gericht → toename van actiepotentiaal frequentie
op surround groen licht gericht → afname van actiepotentiaalfrequentie
center met rood licht & surround met groen licht belichten → er gebeurt niets
hele receptieve veld met wit licht beschijnen → er gebeurt niets
want wit licht bevat zowel golflengte voor groen als golflengte voor rood
hele veld met rood licht beschijnen → toename van actiepotentiaalfrequentie
hele veld met groen licht beschijnen → afname van actiepotentiaalfrequentie
→ baan vanuit P-type ganglioncellen via thalamus (LGN) naar cortex stuurt kleurverschilsignaal (ROOD - GROEN)
(doorzenden van kleurinformatie zit vervat in actiepotentiaalfrequentie)
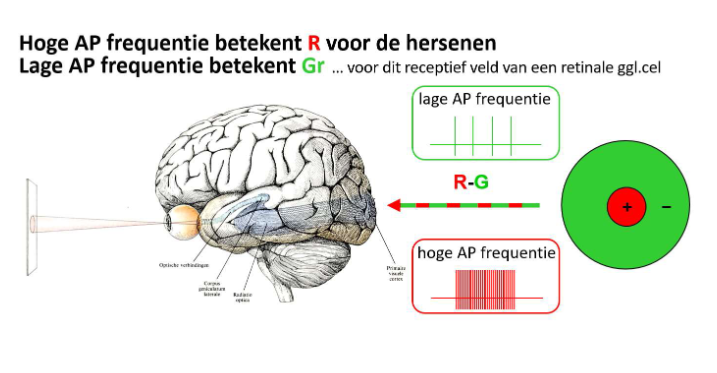
kleurperceptie: kleurwaarneming
3 kegeltjes detecteren reeks golflengten
netvlies & LGN bevatten cellen die 3 kegeltjessignalen omzetten in kegel-opponente verschilsignalen
→ verdere verwerking van signalen: creëren van opponente kleuren
transformaties zorgen voor productie van waargenomen kleuren → vinden plaats in visuele cortex
niet duidelijk hoe
‘blobs’ in V1:
in die blobs: cellen niet geïnteresseerd in oriëntatie, wel in kleur
blobs sturen output naar ‘thin stripe’ gebieden in V2 → van daaruit naar V4
achromatopsie
→ beste bewijs voor gespecialiseerde hersengebieden voor kleur
= verlies van kleurenzicht na hersenschade (centrale kleurenblindheid)
mogelijk grenzen tussen gebieden met verschillende kleuren wel vinden, maar niet aangeven welke kleuren dat zijn
kleurbeleving lijkt verstoord
(terwijl wel aanwijzingen dat golflengte-informatie in eerdere stadia in visuele route wordt verwerkt)
komt voor uit schade aan visuele cortex
laesies van specifieke delen van visuele cortex buiten de primaire visuele cortex
Lilac chaser

kleurperceptie: individuele verschillen in kleurperceptie
prevalentie kleurenblindheid:
8% van mannelijke bevolking
0,5% van vrouwelijke bevolking
→ meer bij mannen want genen die coderen voor M- & L-kegel fotopigmenten zitten op X-chromosoom
mannen: 1 X-chromosoom → dus 1 defect = probleem
vrouwen: 2 kopieën van X-chromosoom → dus 1 defect = normaal kleurenzicht
→ S-kegel fotopigment: elders gecodeerd, dus iedereen heeft 2 kopieën (dus S-kegel kleurendeficiënties zeldzaam)
perifere kleurenblindheid/ daltonisme:
verstoring in 1 of meer van genen die coderen voor de 3 kegel fotopigmenten
2 bepalende factoren:
factor 1: type kegeltje dat is aangetast
factor 2: type defect:
fotopigment voor kegeltje is afwijkend (anders dan normaal)
kegeltje ontbreekt volledig
mensen die 1 kegeltje missen → kleurenblind
MAAR: wereld nog steeds in kleur, maar vlakkere kleurervaring
dichromaat = 2 kegeltjes ipv 3
M- & L-kegeldefecten:
meest voorkomend → meeste kleurenblinden hebben moeite met onderscheiden van licht in midden tot lange golflengtebereik
bv. golflengten 560 & 610 nm
geen van beide lichtsoorten activeert S-kegels sterk
L-kegels vuren ± evenveel af voor beide golflengten
→ we onderscheiden deze lichtsoorten obv M-kegel signalen (hoger voor licht van 560nm dan voor licht van 610nm)
trichomaten zien kleuren: groen & oranjerood
deuteranoop (= iemand zonder M-kegeltjes)
fotoreceptoruitvoer naar deze 2 golflengtes: identiek
dus: 560 & 610 nm golflengtes hebben dezelfde kleur
genetische factoren kunnen ervoor zorgen dat mensen een afwijkende kleur waarnemen
kleurperceptie: individuele verschillen in kleurperceptie → soorten kleurenblindheid
deuteranoop
protanoop
tritanoop
color-anomalous
kegel monochromaat
staaf monochromaat
(centrale kleurenblindheid = achromatopsie)
deuteranoop
= zonder M-kegeltjes
protanoop
= zonder L-kegeltjes
tritanoop
= zonder S-kegeltjes
color-anomalous
= 3 kegel fotopigmenten, maar 2 daarvan lijken zo op elkaar dat deze mensen de wereld vrijwel hetzelfde ervaren als mensen met slechts 2 kegeltypes
→ shift in spectrale gevoeligheid
kegel monochromaat
= slechts 1 type kegeltje in netvlies, naast staafjes
→ dan echt volledig kleurenblind (zeer zeldzaam)
→ enkel grijstinten zien
staaf monochromaten
= geen kegeltjes, enkel staafjes
slecht gezichtsvermogen
geen kleuren onderscheiden
zeer slechte gezichtsscherpte (want staafjes niet in fovea)
ernstige problemen met zien onder normale daglichtomstandigheden
kleurperceptie: van de kleur van licht naar een wereld van kleur
beperkingen bij benadering van kleur:
bruine golflengte?
bruin alleen in relatie tot andere kleuren gezien
color constancy
= neiging om kleuren van objecten relatief onveranderd te zien, ondanks aanzienlijke veranderingen in lichtomstandigheden
bv. waarom ziet papier er zowel wit uit in een groezelige bibliotheek als buiten in het zonlicht?
kleurperceptie: nut van kleurenzicht
kleurenzicht: geen absolute noodzaak, wel een voordeel
in dierenrijk: minstens 2 gedragsgebieden waar kleurenzicht bijzonder nuttig is:
eten
makkelijker om voedsel te vinden & goed van slecht voedsel te onderscheiden
bv. vinden van rijpe frambozen makkelijker met kleurenzicht
voortplanting
zoeken & beoordelen van potentiële partners
kleurrijke vertoningen, van patronen op tropische vissen, tot staart van pauw & gezicht van mandril → seksuele signalen
algemeen: kleurenzicht nuttig bij visuele zoektaken

ruimteperceptie & binoculair zicht
visuele ervaring: reconstructie van wereld obv 2 vervormde signalen:
2 afzonderlijke beelden op netvlies → verschillen altijd, want beide oogbollen bevinden zich op iets andere plaatsen in hoofd
monoculaire cues voor driedimensionale ruimte
binoculaire cues
binoculair zicht & stereopsis
optische illusies
waarom hebben we 2 ogen?
evolutionaire voordeel
je kunt 1 oog verliezen & nog steeds zien
(zelfde als bij 2 longen of 2 nieren)
je ziet meer van de wereld
overlappende, frontale, binoculaire visuele velden geven roofdieren zoals mens een betere kans om kleine, snelbewegende objecten voor zich te zien die mogelijk als voedsel dienen (prooidieren zoals konijnen hebben vaak een zeer breed gezichtsveld, waardoor ze de hele omgeving kunnen observeren op zoek naar roofdieren)
met frontale ogen & overlappende gezichtsvelden → voordeel dat 2 detectoren naar hetzelfde kijken
evolutionair voordeel van overlappende visuele velden:
mogelijkheid om kleine verschillen tussen ogen te gebruiken als aanwijzing voor driedimensionale diepte (stereopsis)
stereopsis = geen noodzakelijke voorwaarde voor diepte- of ruimteperceptie
ruimteperceptie & binoculair zicht: monoculaire cues voor driedimensionale ruimte
occlusie: geeft informatie over relatieve positie van objecten
meest betrouwbare van alle depth cues
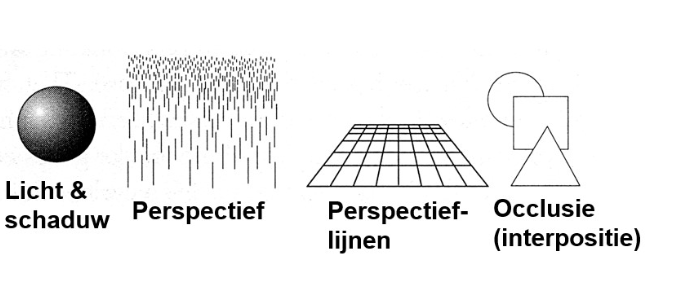
ruimteperceptie & binoculair zicht: monoculaire cues voor driedimensionale ruimte → grootte- & positiesignalen
visueel systeem weet dat, als alle andere zaken gelijk blijven, kleinere dingen verder weg zijn

ruimteperceptie & binoculair zicht: monoculaire cues voor driedimensionale ruimte → bewegingsparallax
in een trein, uit raam kijken naar landschap
beeld A → verandert dan naar beeld B
terwijl trein van links naar rechts ging → alle objecten zijn van recht naar links verschoven
MAAR: sommige objecten meer verschoven dan andere
→ bloem: bijna helemaal over netvlies bewogen, koe wat minder & boom nauwelijks veranderd
→ parallax = geometrische relatie
objecten dichter bij je verplaatsen zich meer dan objecten verder weg
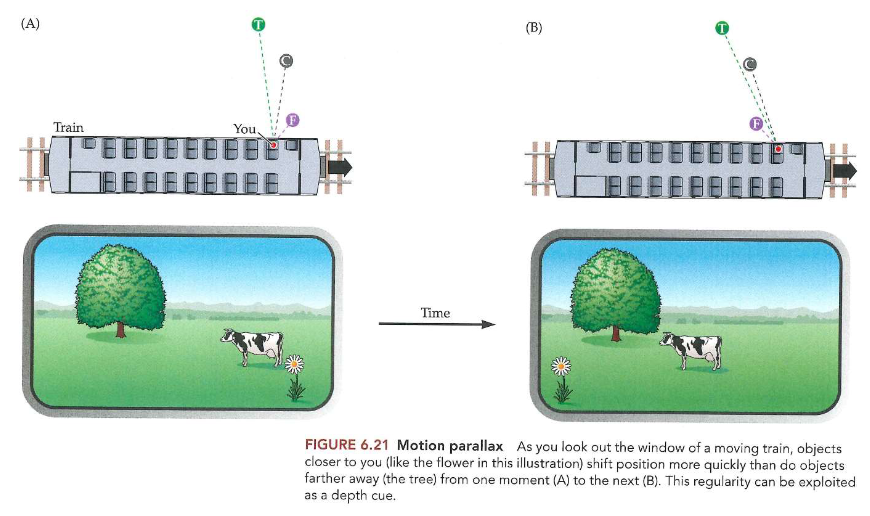
bewegingsparallax vs. binoculaire stereopsis
bewegingsparallax = geometrische informatie verkregen uit een oog in 2 verschillende posities op 2 verschillende tijdstippen
binoculaire stereopsis = informatie verkregen uit 2 ogen in verschillende posities in het hoofd op hetzelfde moment
→ beeld wel vergelijkbaar