9. Germ layer formation and gastrulation
1/16
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced |
|---|
No study sessions yet.
17 Terms
Cell Derivation
From what do ALMOST all cells of the body derive?
Which cells are an exception?
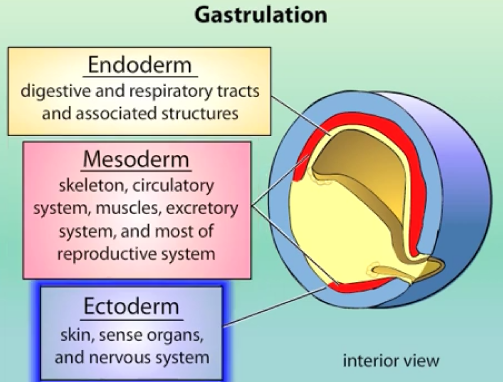
ALMOST all cells of the body derive from 3 primary germ layers:
ectoderm (external layer) - skin, sense organs & nervous system.
mesoderm (middle layer) - skeleton, circulatory system, muscles, excratory system, and most of reproductive system.
endoderm (internal layer) - digestive and respiratory tracts & associated structures.
Germ cells (sperm and egg) are an exception, arising separately.
Why is Xenopus (African clawed frog) a common model organism for studying germ layer formation and gastrulation?
Xenopus is a good model because it has:
large and abundant eggs
external development (allowing easy observation)
ease of injection (for manipulating gene expression or introducing tracers)
amenability to microsurgery and transplants
it serves as a good general model for vertebrate development.
Many mechanisms are conserved in other vertebrates, including humans. The same molecules in Xenopus are thought to operate in chicks/humans (i.e the genes and mechanisms are conserved).

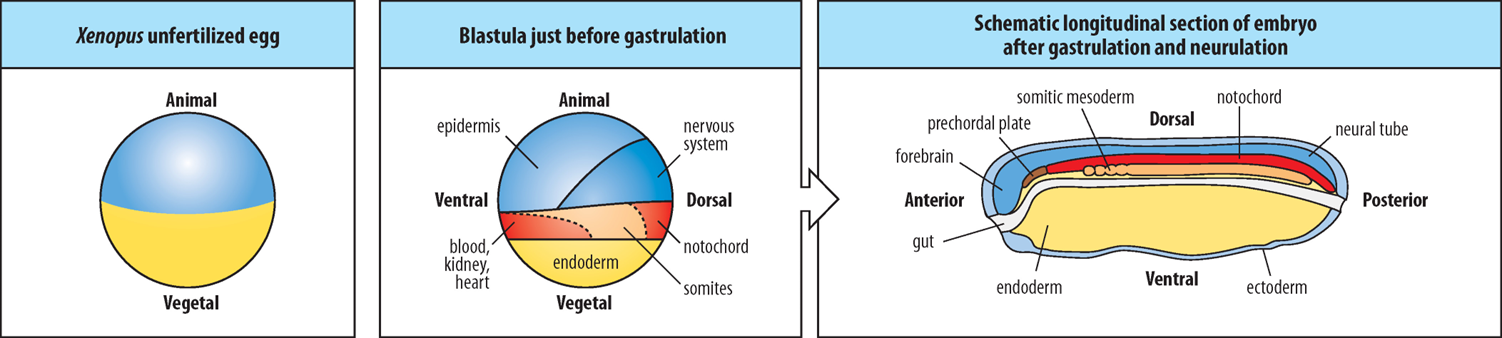
What is the fundamental polarity of an unfertilized Xenopus egg, and how does this influence germ layer formation?
A Xenopus egg is polarised with an animal pole (pigmented, less yolky, top) and a vegetal pole (yolky, bottom). This polarity is established durng oogenesis through the differential localisation of maternal mRNAs and proteins (maternal determinants) (e.g VegT maternal factor is localised to the vegetal pole).
This polarity triggers germ layer formation:
Specific maternal mRNAs (cytoplasmic determinants) are already localized in the vegetal hemisphere of the egg.
These mRNAs produce proteins (like VegT) that signal to animal hemisphere cells, inducing them to differentiate into the 3 germ layers.

How are the 3 primary germ layers (ectoderm, mesoderm, endoderm) initially formed in relation to the animal and vegetal hemispheres in Xenopus?
How do cells arising from the Vegetal and Animal Hemispheres of the early Xenopus blastula differ from one another?
What is the crucial interaction that occurs between cells derived from the Vegetal and Animal Hemispheres?
Which germ layer does the Vegetal Hemisphere primarily contribute to?
In the Animal Hemisphere, what determines whether cells become mesoderm or ectoderm?
Where are the germ layers localised in the embryo after gastrulation and neurulation, and what does each go on to form?
Cells arising from within the Vegetal and Animal Hemispheres of the early blastula are different to one another in terms of their developmental potential and the signals they produce or respond to.
Cells derived from the Vegetal (lower) Hemisphere SIGNAL to cells derived from the Animal Hemisphere, influencing their developmental fate.
Vegetal Hemisphere cells primarily become endoderm.
Proximity to the Vegetal Hemisphere determines the fate of Animal Hemisphere cells. Much of the Equatorial Animal Hemisphere cells (in the Marginal Zone) close to the Vegetal Hemisphere become mesoderm, while Animal Hemisphere cells further away from the Vegetal Hemisphere become ectoderm.
The layers:
Ectoderm is the outermost layer, forming the epidermis and nervous system (neural tube, forebrain).
Mesoderm is the middle layer, forming structures like the notochord, prechordal plate, somitic mesoderm, blood, kidney, and heart.
Endoderm is the innermost layer, forming the gut.
How do rapid cleavage divisions produce the blastula and contribute to the formation of embryonic germ layers from the animal and vegetal hemispheres?
Following fertilization, the zygote undergoes rapid cleavage divisions (mitotic, without growth), parcelling the large cell into smaller blastomeres to form the blastula with a blastocoel. This process, driven by maternal factors:
Subdivides the egg into manageable cells.
Unevenly distributes maternal determinants (e.g., in animal vs. vegetal hemispheres).
Establishes spatial relationships critical for later cell signaling via the blastula with its blastocoel.
This differential distribution, alongside subsequent zygotic gene expression and cell interactions, leads to the specification of distinct germ layers from the previously different animal and vegetal hemisphere cells.
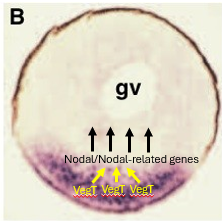
Vegetally-localised determinant - What is the role of VegT in Xenopus germ layer formation?
What is VegT?
Where is VegT’s mRNA localised?
What happens when VegT protein is produced after fertilisation?
What are Nodal signal important for?
What is Nodal?
VegT is a T-box transcription factor.
VegT is encoded by mRNA that is maternally localised to the vegetal hemisphere of the Xenopus egg.
After fertilization, VegT protein is synthesised and binds to the promoters of genes encoding proteins related to the secreted morphogen Nodal - (Xnr genes - Xenopus nodal-related) activating their transcription.
These Nodal signals are crucial for inducing mesoderm and patterning the endoderm.
Nodal is part of the Transforming Growth Factor Beta (TGF-β) protein family.
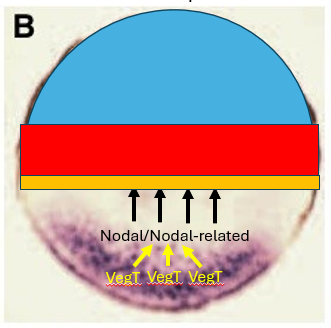
How does the Nodal morphogen gradient specify different germ layers in Xenopus?
Nodal-related proteins are secreted from the vegetal cells (where their expression is activated by VegT) and diffuse into the overlying animal hemisphere cells, forming a concentration gradient.
Cells respond differently based on the Nodal concentration:
Cells closest to the source (highest Nodal) become endoderm.
Cells at intermediate distances (intermediate Nodal) are induced to become mesoderm.
Cells furthest away (no Nodal) become ectoderm.
Morphogen
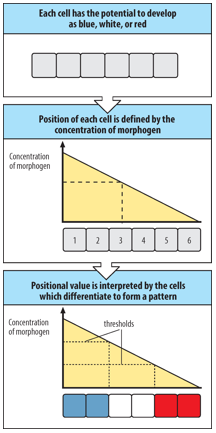
What is a morphogen? (3)
Give examples relevant to early development.
A morphogen is a:
molecule that induces concentration-dependent changes in the fates of responding cells.
molecule distributed in a gradient within a tissue that directs cell differentiation in a position-dependent manner.
molecule that elicits different cell differentiation responses at different threshold concentrations.
Relevant examples include Nodal, Wnt, and Sonic hedgehog (Shh) proteins.
Symmetry-breaking event
What is the "symmetry-breaking event" in Xenopus, and what key structures does it lead to?
Gastrulation is prefigured by a symmetry-breaking event that occurs at the time of fertilization, leading to the formation of the Nieuwkoop Centre in the blastula. The Nieuwkoop Centre induces the formation of the Spemann-Mangold Organizer, which then initiates gastrulation.
Describe the 5 key steps from sperm entry to the formation of the Spemann-Mangold Organizer in Xenopus.
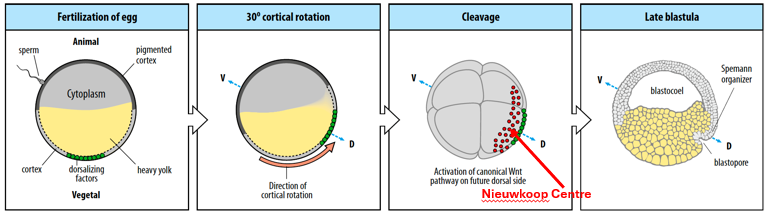
The future dorsal (back) side of the embryo develops opposite the site of sperm entry (which is typically in the animal hemisphere).
Cortical rotation occurs, where the cortical layer of the egg rotates by 30°, towards the future dorsal region which is the point opposite the sperm entry site.
This rotation relocates maternal dorsalising factors (e.g., Wnt11, Dishevelled, β-catenin) along microtubules to the future dorsal side, inducing the formation of the organiser, the Nieuwkoop Centre (in the dorsal-vegetal region of the blastula).
The Nieuwkoop Centre (a signaling center rich in β-catenin and Nodal/Vg1 signals) then induces the overlying dorsal marginal zone cells to form the Spemann-Mangold Organizer.
The Spemann-Mangold Organizer will pattern the future A-P and D-V axes and initiate gastrulation.
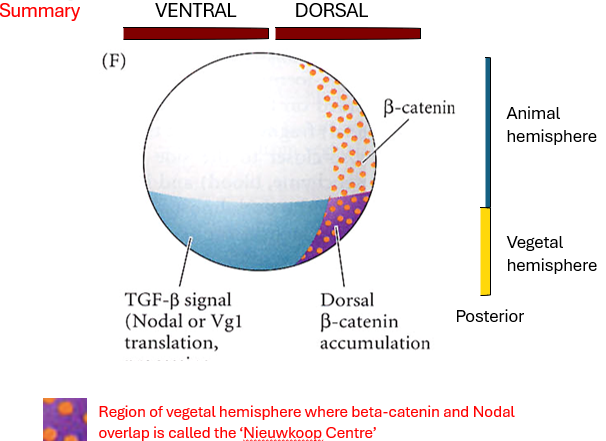
How do β-catenin and Nodal-related signalling cooperate to establish the Nieuwkoop Centre and the Spemann-Mangold Organizer?
What is the role of VegT in early Xenopus development, and where is it located?
How does β-catenin become asymmetrically distributed in the early embryo, and what is its significance?
Define the "Nieuwkoop Centre" and explain how it forms.
What is the function of the Nieuwkoop Centre?
What is the Spemann-Mangold Organizer, and how is it induced?
How does the concentration of Nodal-related signals influence mesodermal fates in the marginal zone?
VegT (vegetally localized) specifies endoderm and activates Nodal-related gene expression throughout the vegetal hemisphere.
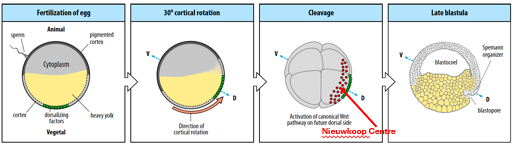
Cortical rotation (triggered by fertilisation) leads to the accumulation β-catenin on the future dorsal side of the embryo (specifically in the dorsal animal and dorsal vegetal regions). This dorsal accumulation is crucial for establishing the dorsal-ventral axis.
The Nieuwkoop Centre is a signalling region that forms in the dorsal-vegetal region of the embryo. It arises where high Nodal signaling (activated by VegT) and high β-catenin activity overlap. This combination results in the highest concentration of Nodal-related proteins.
The Nieuwkoop Centre acts as the primary organiser of the embryonic axis. Its main function is to induce the formation of the Spemann-Mangold Organizer in the overlying dorsal marginal zone.
The Spemann-Mangold Organiser is a critical signalling center in the dorsal marginal zone. It is induced by the high Nodal signaling and β-catenin activity originating from the Nieuwkoop Centre. The Organiser later patterns the future A-P and D-V axes and initiates gastrulation.
Cells with lower Nodal (ventral and lateral marginal zone) become ventral mesoderm.
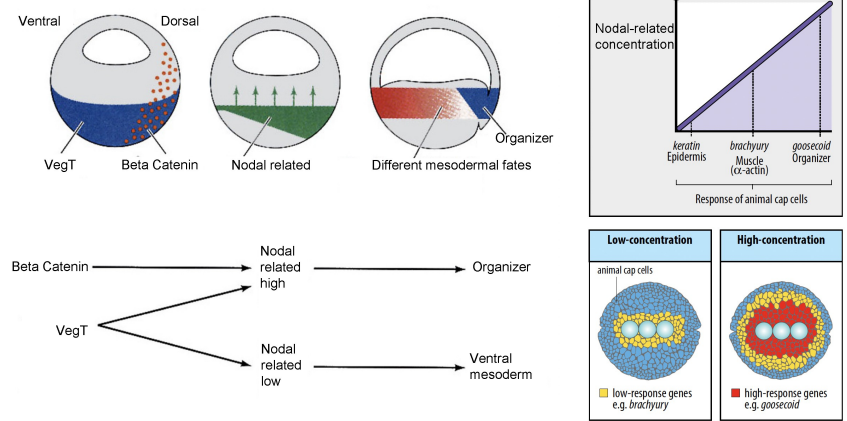
How does the concentration of Nodal-related signals influence mesodermal fates in the marginal zone of the embryo?
A gradient of Nodal-related signals (created by Beta Catenin activity) in the marginal zone specifies different mesodermal fates as the Nodal concentration decreases laterally and ventrally:
High Nodal-related concentration driven by the Nieuwkoop Centre, in the Organiser activates genes such as Goosecoid, which later results in inducing dorsal mesoderm (e.g., notochord, prechordal plate). Although Wnt/β-catenin is also required.
Intermediate concentrations induce muscle (e.g., brachyury, α-actin).
Lower Nodal-related concentrations in the ventral and lateral marginal zone cells leads to their differentiation into ventral mesoderm (e.g., blood, kidney, heart).
Animal cap cells not exposed to Nodal become ectoderm (e.g., keratin Epidermis).
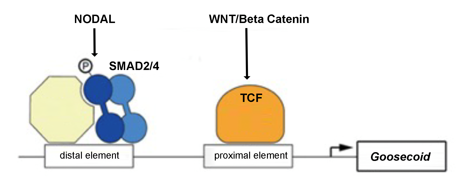
What are the key transcription factors expressed in the Spemann-Mangold Organizer, and how are they induced?
What are organiser specific genes like Goosecoid transcriptionally activated by?
What does this mean the Organiser can express?
Where are Brachyury transcription factors expressed?
What is created that initiates the gastrulation process, and how was this created?
What is the equivalent structure of the Organiser in chicks and humans?
Organiser specific genes like Goosecoid (Gsc) are only transcriptionally activated via BOTH:
A Nodal downstream effector (Smad2/4)
A Wnt/β-catenin downstream effector (activating TCF (T-cell factor) downstream effectors).
The Spemann-Mangold Organiser expresses a combination of β-catenin-induced T-cell factor & Nodal-related-activated Smad2 transcription factors, which induce other transcription factors such as Goosecoid, Not1, and Lim1 in the Organiser tissue.
Brachyury transcription factor is expressed throughout the mesoderm, including in the Organiser.
The targets of Organiser-specific transcription factors in combination with Brachyury targets create dorsal axial mesoderm which initiates the gastrulation process.
In chicks and humans, the equivalent structure of the Organiser is “Hensen's Node”.
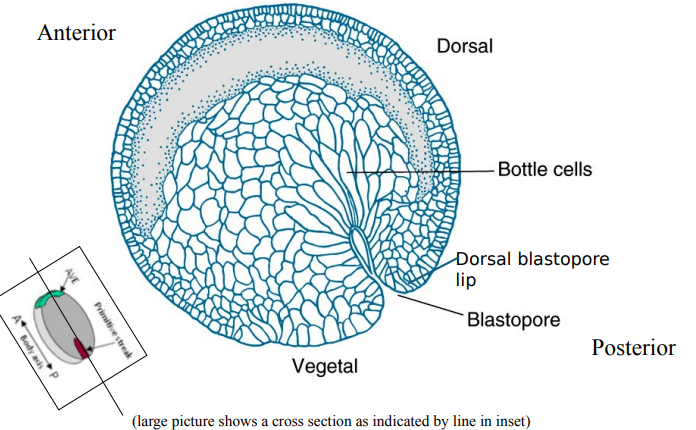
What are the main cell movements during Xenopus gastrulation, and how is the anterior-posterior axis established?
What is gastrulation driven by and where does it begin?
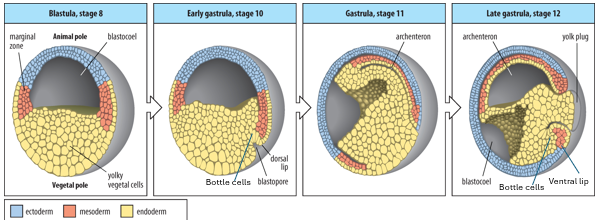
Describe the 3 main movements by which gastrulation occurs?
1: Involution
2: Convergent Extension
3: Epiboly
So how is the A-P axis established?
Gastrulation is driven by coordinated cell movements. It begins at the dorsal lip of the blastopore, formed by the Organiser.
Involution: Sheets of cells (mesoderm and endoderm) move from the exterior into the interior of the embryo through the blastopore lip. Bottle cells at the dorsal lip initiate this movement.
Convergent extension: Cells of the dorsal mesoderm (Organiser region) intercalate mediolaterally, causing the tissue to converge (narrow) and extend (lengthen) along the A-P axis. This elongates the notochord and forms the primary A-P axis.
Epiboly: Ectoderm cells spread to cover the entire surface of the embryo. The archenteron (primitive gut) forms, replacing the blastocoel.
Convergent extension is when a tissue narrows along one axis and extends along another. It begins at one end, defining the posterior (blastopore), and then moves anteriorly.
The Organiser drives the tissue movements that create the A-P axis during gastrulation through its differentiation & convergent extension.
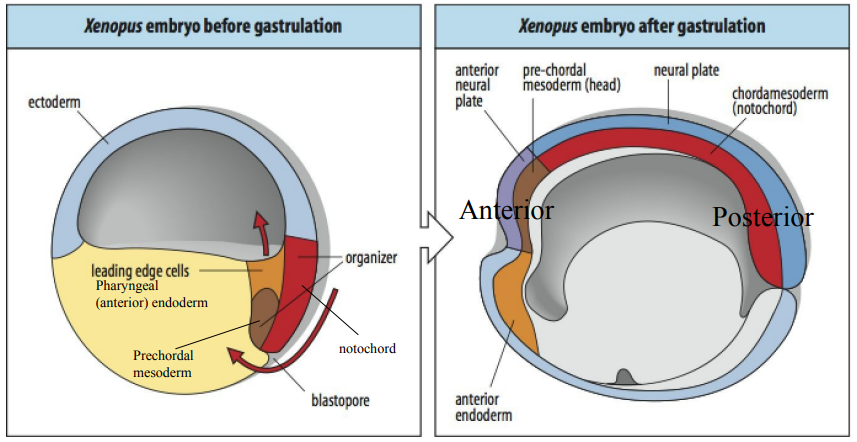
Describe the key events of Xenopus gastrulation and the resulting germ layer arrangement after. (5)
Gastrulation begins as bottle cells initiate involution, where a sheet of cells moves inward into the interior of the gastrula through the dorsal lip of the blastopore. This is indicated by the spreading of mesoderm cells. Mesoderm cells at different A-P and D-V positions become prechordal mesoderm, notochord, paraxial mesoderm (somitic), intermediate mesoderm, and lateral plate mesoderm.
Simultaneously, ectoderm cells spread around the embryo's surface via epiboly.
As gastrulation progresses, the blastocoel shrinks, and a new cavity, the archenteron (the future gut), forms and is completely lined by endoderm.
A 2nd population of bottle cells forms the ventral lip of the blastopore , where more mesoderm involutes (producing some of the reproductive tissues of the embryo) as epiboly has pushed it towards the dorsal lip.
Finally, the embryo is organized into 3 distinct germ layers: the ectoderm externally surrounds the embryo, the endoderm lines the future gut (archenteron), and the mesoderm is positioned in between the ectoderm and endoderm.
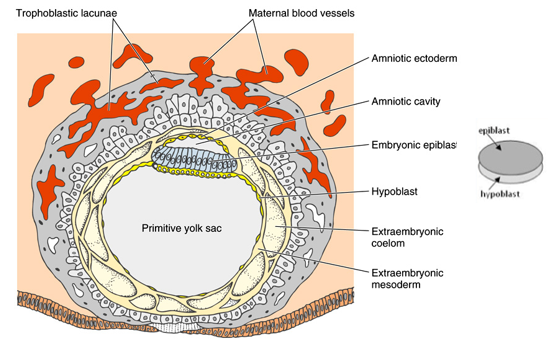
How does early human embryonic development (e.g., 12-day old embryo) compare to early Xenopus development in terms of establishing basic layers?
By 12 days, the embryo has implanted into the uterine wall.
The implanted human embryo has formed a bilaminar disc consisting of a top epiblast layer and a bottom hypoblast layer.
This is analogous to the animal (epiblast) and vegetal (hypoblast) hemispheres of an early Xenopus embryo.
The epiblast will give rise to all 3 germ layers of the embryo proper (ectoderm, mesoderm, endoderm) during gastrulation, similar to how the animal hemisphere cells in Xenopus differentiate.
The blastopore lip in Xenopus is analogous to the primitive streak in mammals and birds, which is the site of gastrulation.
Summarize the key steps in Xenopus germ layer formation and early patterning.
Egg Polarisation: Vegetal hemisphere contains maternal determinants (e.g., VegT mRNA).
Nodal Induction: VegT activates Nodal-related (Xnr) genes in vegetal cells.
Mesoderm Induction: Nodal proteins diffuse, inducing overlying equatorial cells to become mesoderm.
Dorsal Axis Formation: Fertilisation breaks symmetry; cortical rotation relocates Wnt components, forming the Nieuwkoop Centre dorsally.
Organiser Induction: Combined VegT and β-catenin (from Wnt signaling) create a Nodal gradient; highest Nodal in Nieuwkoop Centre induces the Spemann-Mangold Organiser.
Mesodermal Fates: Different Nodal thresholds specify different mesodermal fates (e.g., Organiser vs. ventral mesoderm).
Organiser Gene Expression: Specific transcription factors (e.g., Goosecoid) are activated in the Organiser by combined Nodal and Wnt/β-catenin pathways.
Gastrulation: The Organiser drives cell movements (convergent extension, involution) establishing the A-P axis. These mechanisms are highly conserved in vertebrates.