Cellebiologi
1/200
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
201 Terms
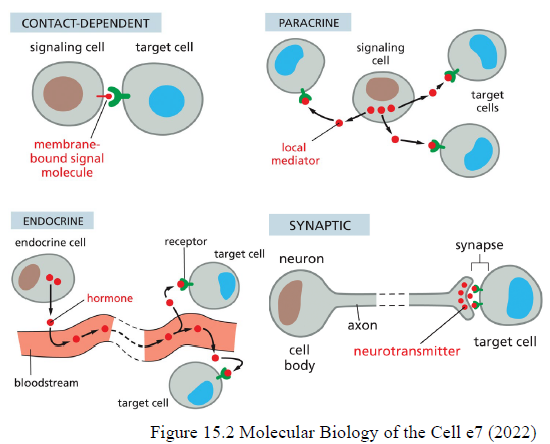
Hvad er de fire hovedtyper af cell signaling, og hvordan fungerer de?
Contact-dependent: signalmolekyle sidder i membranen og kræver direkte cellekontakt
Paracrine: lokale mediatorer diffunderer til nærliggende celler
Endocrine: hormoner transporteres gennem blodet til fjerne celler
Synaptic: neuroner frigiver neurotransmittere i synapser
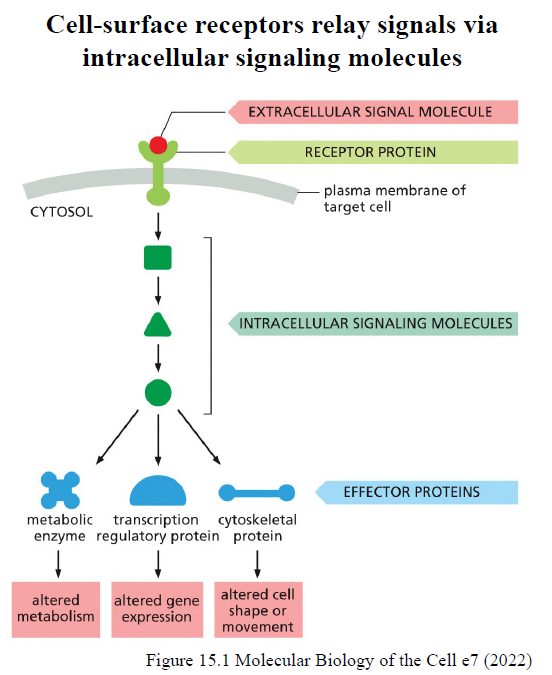
Beskriv det generelle flow i cell signaling.
Et extracellulært signal binder en receptor → intracellulære signaling molecules aktiveres → effector proteins påvirkes → cellulært respons opstår.
Effector proteins kan ændre:
metabolisme
genekspression
cytoskelet
cellebevægelse/form
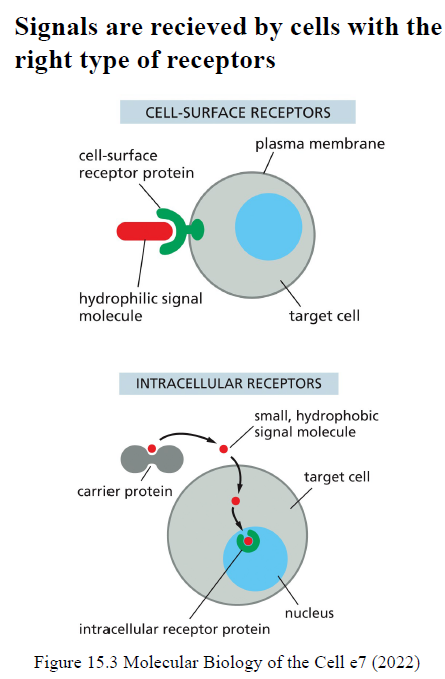
Hvad er forskellen mellem cell-surface receptors og intracellular receptors?
Cell-surface receptors binder typisk hydrofile/store signalmolekyler, som ikke kan krydse membranen.
Intracellular receptors binder små hydrofobe signalmolekyler, som kan diffundere gennem membranen og ofte regulerer genekspression direkte.
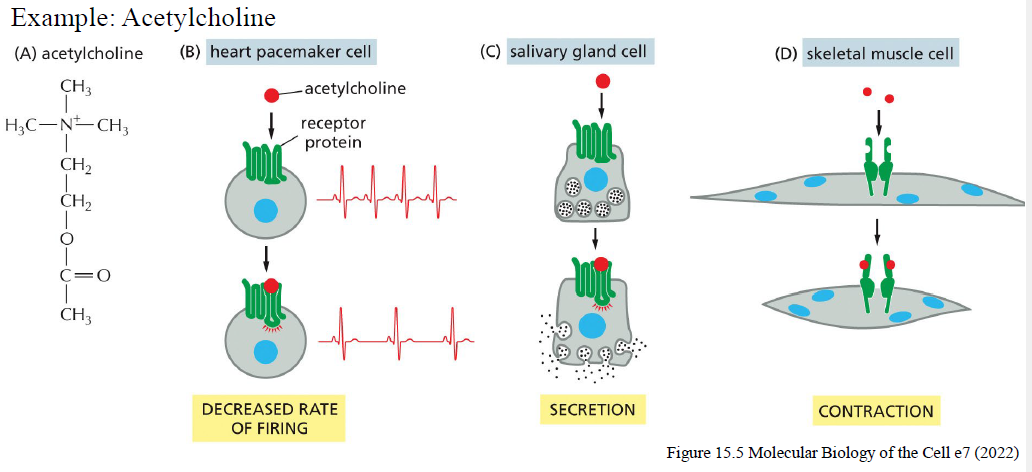
Hvorfor kan samme signalingmolekyle give forskellige effekter i forskellige celletyper?
Fordi forskellige celletyper udtrykker forskellige:
receptorer
intracellulære signaling proteins
effector proteins
Eksempel: acetylcholine kan:
sænke firing rate i hjertet
stimulere sekretion i kirtler
inducere kontraktion i skeletmuskel
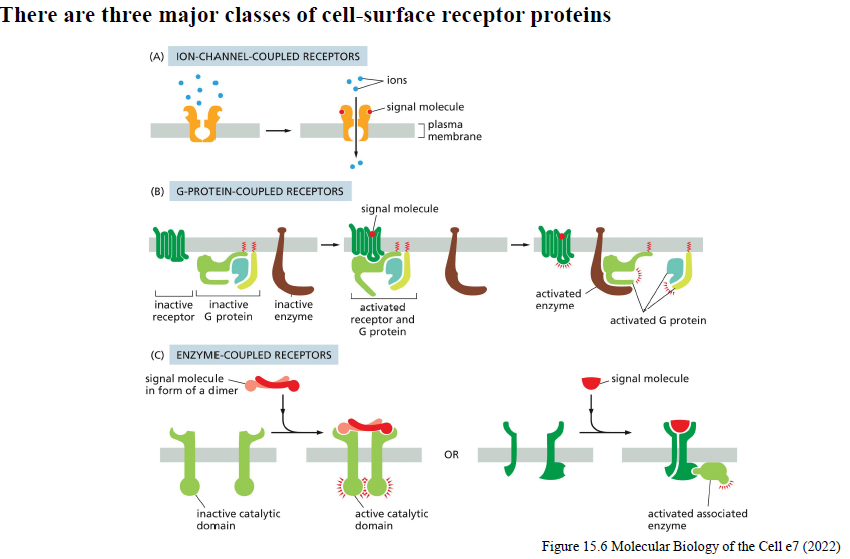
Hvad er de tre hovedtyper af cell-surface receptors, og hvordan fungerer de?
1. Ion-channel-coupled receptors
Ligandbinding åbner/lukker ionkanaler → hurtige responser.
2. G-protein-coupled receptors (GPCRs)
Ligandbinding aktiverer G-proteiner, som regulerer enzymer eller ionkanaler.
3. Enzyme-coupled receptors
Ligandbinding aktiverer receptorens enzymaktivitet eller associerede enzymer, ofte via dimerisering.
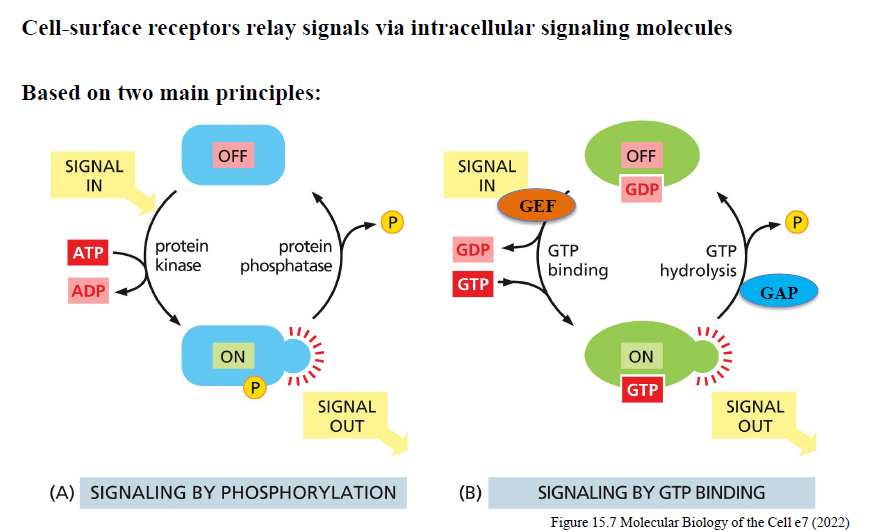
Hvordan fungerer signaling via phosphorylation?
Protein kinases overfører fosfat fra ATP til målproteiner og ændrer deres aktivitet.
Protein phosphatases fjerner fosfatgrupper igen.
Balancen mellem kinase- og phosphataseaktivitet bestemmer signaling-outputtet.
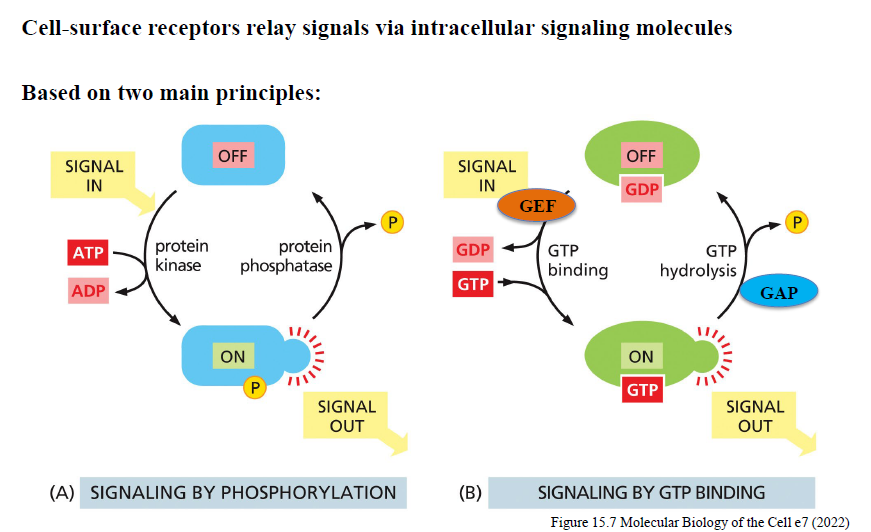
Hvordan fungerer signaling via GTP-binding proteins?
GTP-bindende proteiner fungerer som molekylære switches:
GDP-bundet = OFF
GTP-bundet = ON
GEFs aktiverer ved at udskifte GDP med GTP.
GAPs inaktiverer ved at stimulere GTP-hydrolyse.
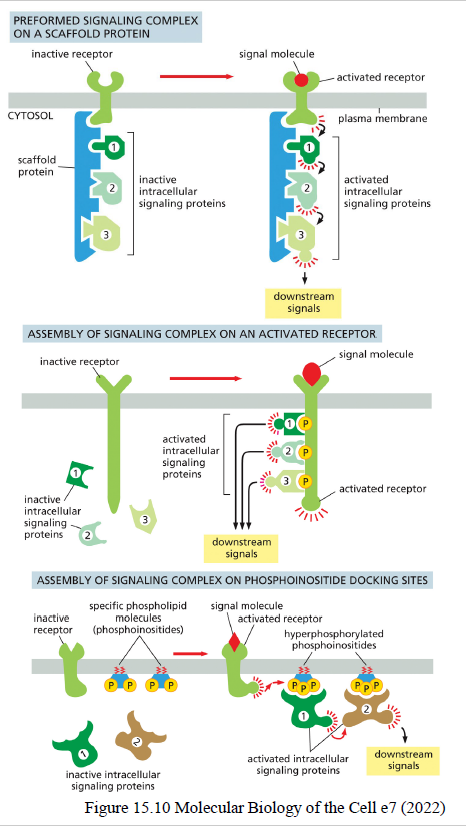
Hvordan sikrer cellen specificitet i signaling pathways?
Cellen bruger flere mekanismer:
scaffold proteins samler signalingproteiner i komplekser
modular interaction domains binder specifikke motiver/fosforyleringer
lokal koncentration af signalingmolekyler i bestemte områder af cellen
Dette gør signaling hurtigere og mere præcis i det “støjende” cytoplasma.
Hvad betyder integration af multiple signaler?
Et cellulært respons kan kræve flere samtidige signaler (“coincidence detection”).
Eksempel:
Anchorage-dependent proliferation kræver både:
growth factor signaling
integrin signaling
Dette fungerer som en kontrolmekanisme, så celler ikke deler sig forkert.
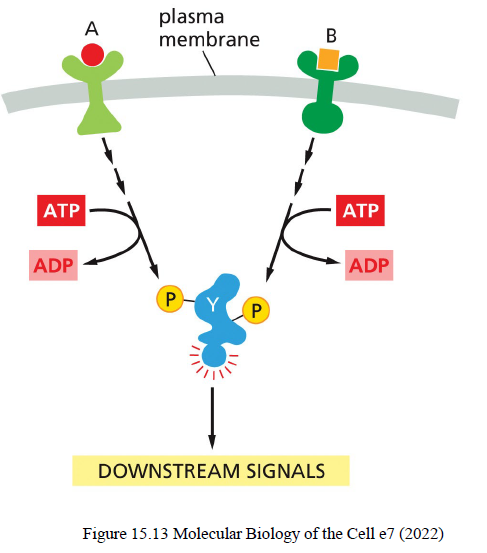
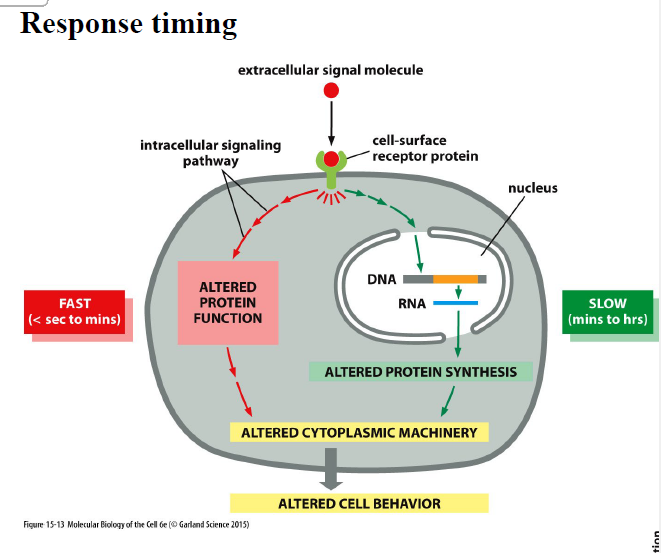
Hvad er forskellen på hurtige og langsomme signaling-responser?
Hurtige responser
sekunder til minutter
ændrer aktivitet af eksisterende proteiner
kræver ikke ny proteinsyntese
Langsomme responser
minutter til timer
ændrer genekspression
kræver ny proteinsyntese
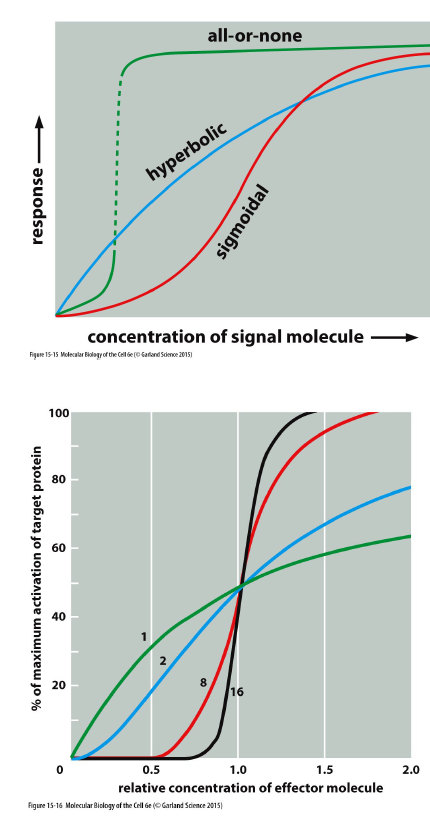
Hvordan kan forholdet mellem signal og respons variere mellem signaling pathways?
Pathways kan variere i:
sensitivitet
dynamisk range
persistence
signal processing
integration
coordination
Responskurver kan være:
hyperbolske → gradvis respons
sigmoidale → tærskel-lignende respons
all-or-none → næsten ingen respons indtil tærskel nås
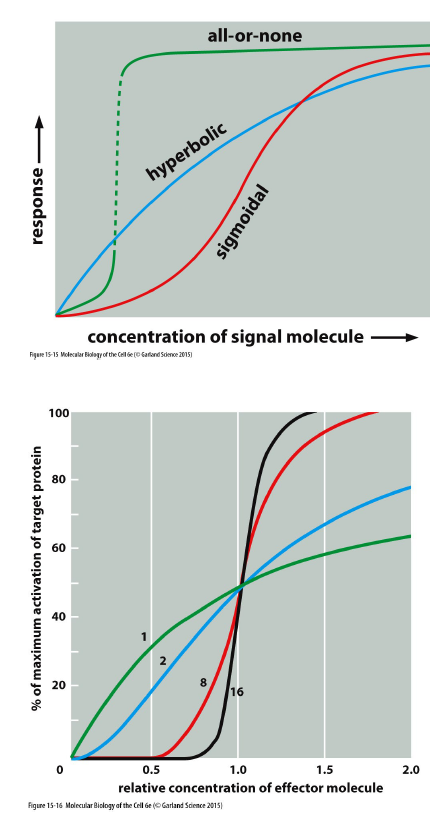
Hvilke mekanismer kan ændre formen på en signaling-responskurve?
aktivering af allosteriske proteiner
feedback-mekanismer
Disse kan gøre responset mere gradvist, skarpere eller switch-lignende.
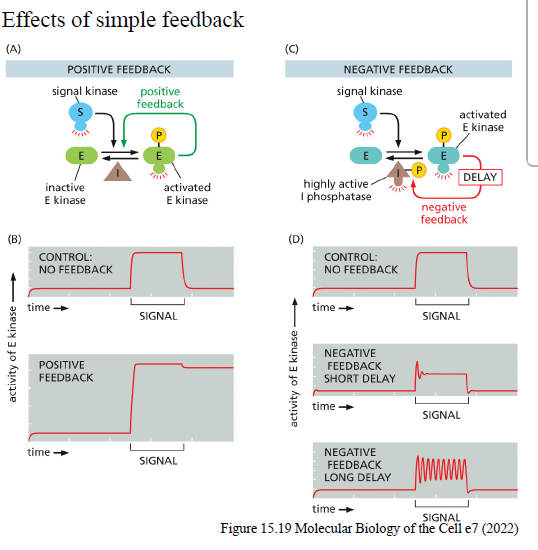
Hvad er forskellen mellem positiv og negativ feedback i signaling pathways?
Positiv feedback
Et aktivt komponent stimulerer yderligere sin egen aktivering.
Kan give:
hurtig forstærkning
switch-lignende responser
“memory”-effekter
Negativ feedback
Et aktivt komponent hæmmer pathwayet.
Kan give:
stabilisering
dæmpning af signal
oscillationer hvis feedback er forsinket
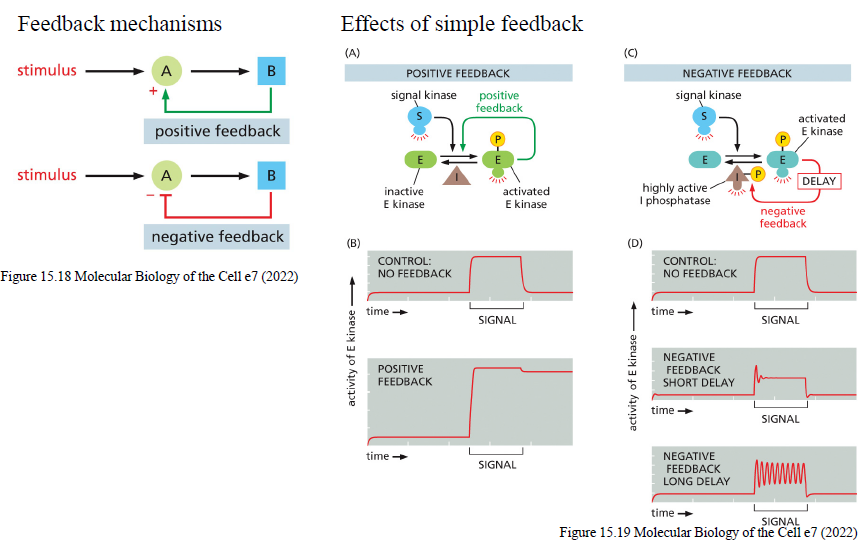
Hvordan kan negativ feedback skabe oscillationer i signaling pathways?
Hvis negativ feedback er forsinket:
pathway aktiveres
inhibitor opbygges langsomt
pathway hæmmes
inhibitor forsvinder igen
pathway kan reaktiveres
Dette kan skabe gentagne oscillationer.
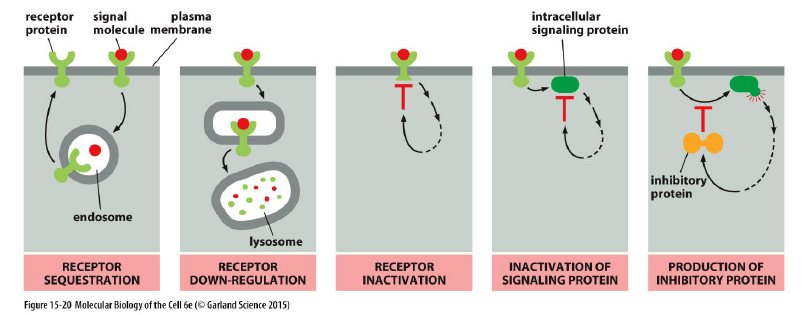
Hvordan kan celler justere deres sensitivitet til et signal?
Ved adaptation/desensitization.
Sensitiviteten afhænger af:
antal receptorer
receptorernes affinitet for ligand
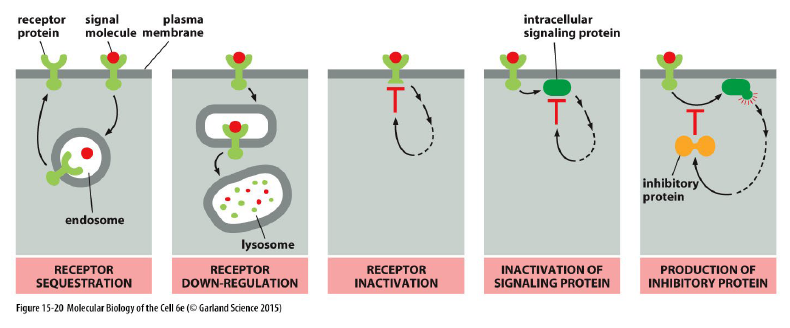
Hvilke mekanismer kan reducere en celles sensitivitet til et signal?
Receptor sequestration
Receptorer fjernes midlertidigt fra plasmamembranen.Receptor down-regulation
Receptorer nedbrydes i lysosomer.Receptor inactivation
Receptoren inaktiveres direkte.Inactivation of signaling proteins
Intracellulære signalingproteiner hæmmes.Production of inhibitory proteins
Cellen producerer inhibitoriske proteiner.
Hvad karakteriserer G-protein-coupled receptors (GPCRs)?
GPCRs er celleoverfladereceptorer, som aktiverer trimere G-proteiner.
Mennesker har >800 GPCRs.
Ligander inkluderer:
proteiner
peptider
aminosyrederivater
fedtsyrer
lys/fotoner
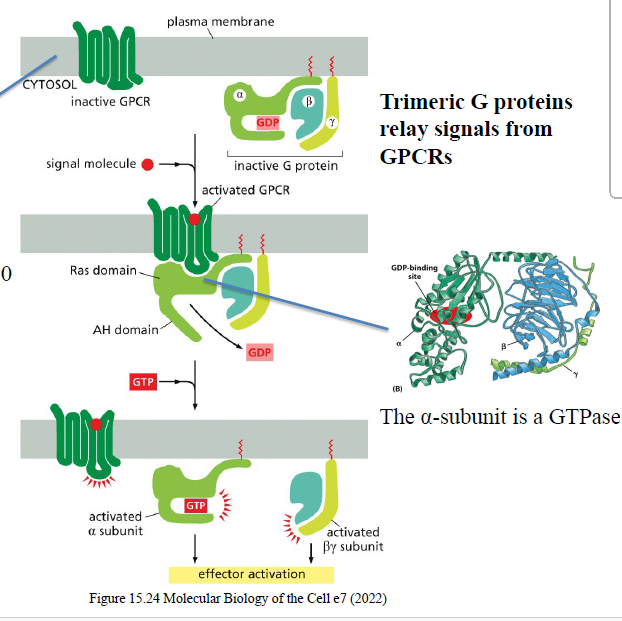
Beskriv aktiveringen af trimere G-proteiner via GPCRs.
Ligand binder GPCR
GPCR ændrer konformation
GDP på Gα udskiftes med GTP
Gα dissocierer fra βγ
Gα-GTP og βγ kan aktivere effektorproteiner
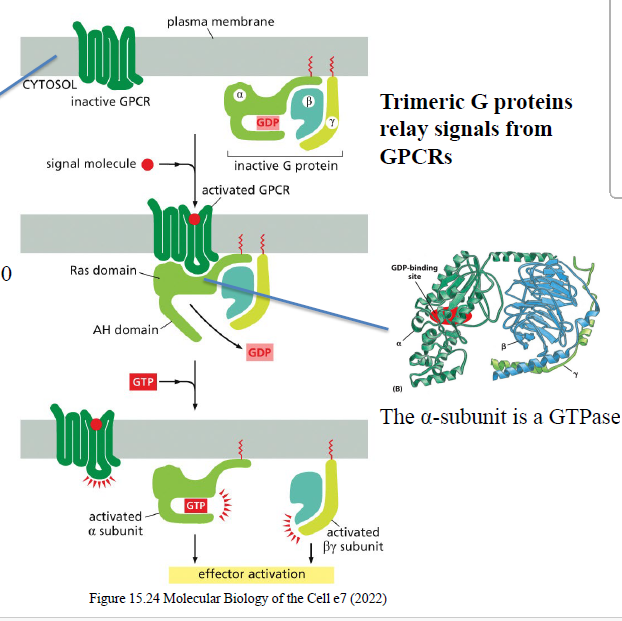
Hvilken vigtig enzymaktivitet har Gα-subunit?
Gα-subunit er en GTPase og kan hydrolysere GTP → GDP for at slukke signalet.
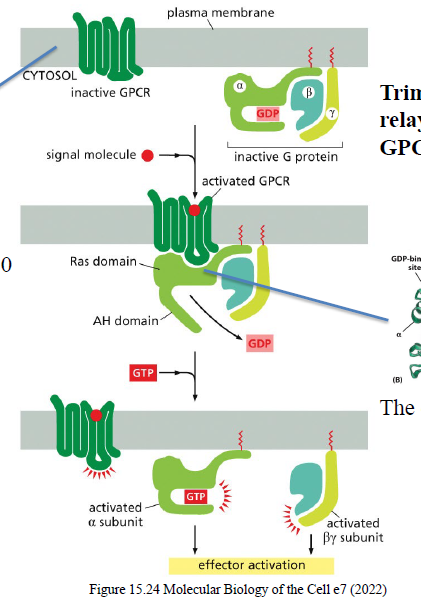
Hvordan klassificeres trimere G-proteiner?
De klassificeres efter deres Gα-subunit.
Fire hovedfamilier:
Gs
Gi
Gq
G12/13
Hvordan påvirker Gs og Gi adenylyl cyclase?
Gs aktiverer adenylyl cyclase
Gi inhiberer adenylyl cyclase
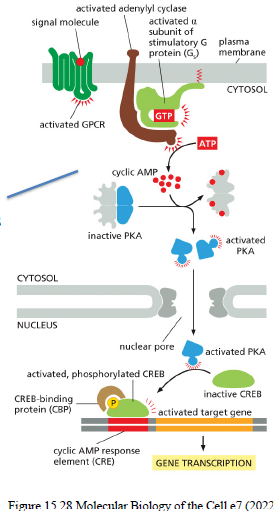
Beskriv den klassiske cAMP signaling pathway downstream for GPCRs.
Ligand aktiverer GPCR
Gs aktiveres
Gαs aktiverer adenylyl cyclase
ATP omdannes til cAMP
cAMP aktiverer PKA
PKA fosforylerer målproteiner og transcription factors (fx CREB)
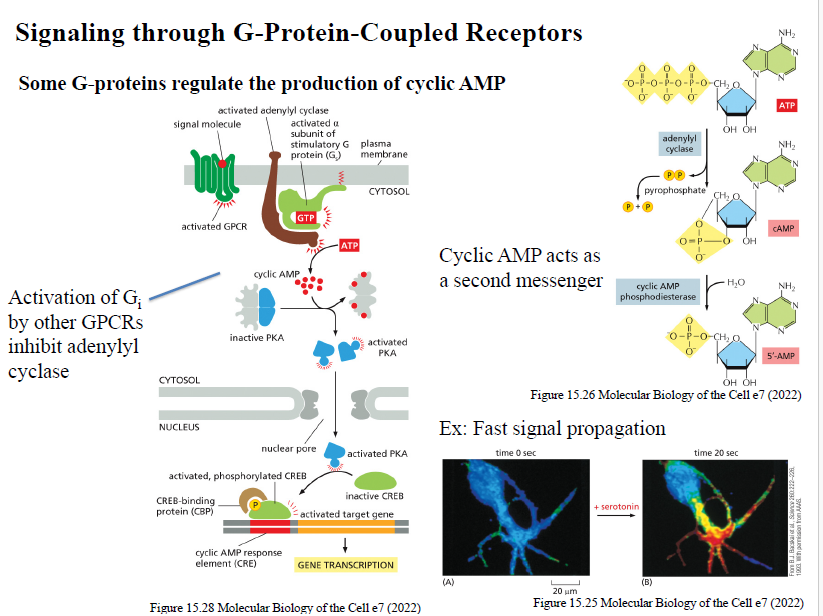
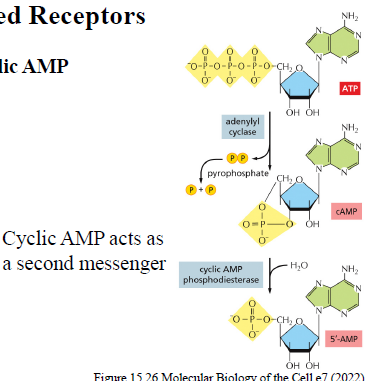
Hvad er cAMP, og hvordan dannes det? Hvordan nedbrydes cAMP?
cAMP er en second messenger.
Det dannes fra ATP af enzymet adenylyl cyclase.
cAMP phosphodiesterase omdanner cAMP til 5’-AMP.
Hvad sker der når cAMP aktiverer PKA?
PKA fosforylerer intracellulære målproteiner og kan aktivere transcription factors som CREB, hvilket ændrer genekspression.
Beskriv signaling via Gq og phospholipase C-β.
GPCR aktiverer Gq
Gαq aktiverer phospholipase C-β (PLCβ)
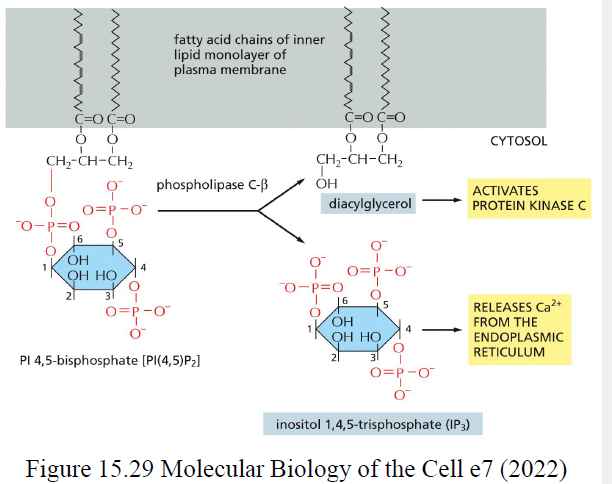
PLCβ kløver PIP2 til:
DAG
IP3
IP3 åbner Ca²⁺-kanaler i ER
Cytosolisk Ca²⁺ stiger
DAG + Ca²⁺ aktiverer protein kinase C (PKC)
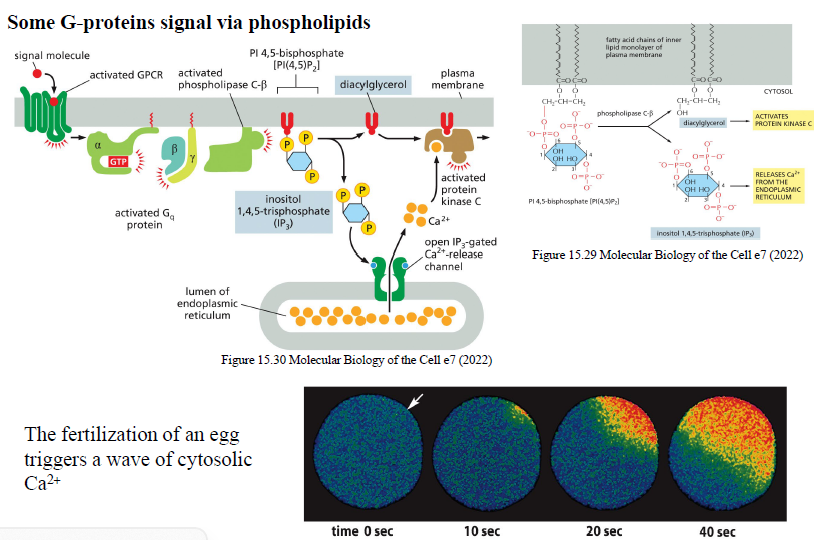
Hvad er funktionerne af IP3 og DAG?
IP3 frigiver Ca²⁺ fra ER
DAG aktiverer protein kinase C
Hvorfor bruges Ca²⁺ ofte som second messenger?
Ca²⁺ kan hurtigt ændre koncentration i cytosol og regulere mange proteiner og cellulære responser.
Hvordan kan feedback skabe Ca²⁺-waves og oscillationer?
Frigivet Ca²⁺ kan stimulere yderligere Ca²⁺-frigivelse fra ER via positiv feedback, hvilket skaber bølger og oscillationer.
Hvad er calmodulin?
Calmodulin er et Ca²⁺-bindende regulatorisk protein, som ændrer konformation når det binder Ca²⁺ og derefter aktiverer målproteiner.
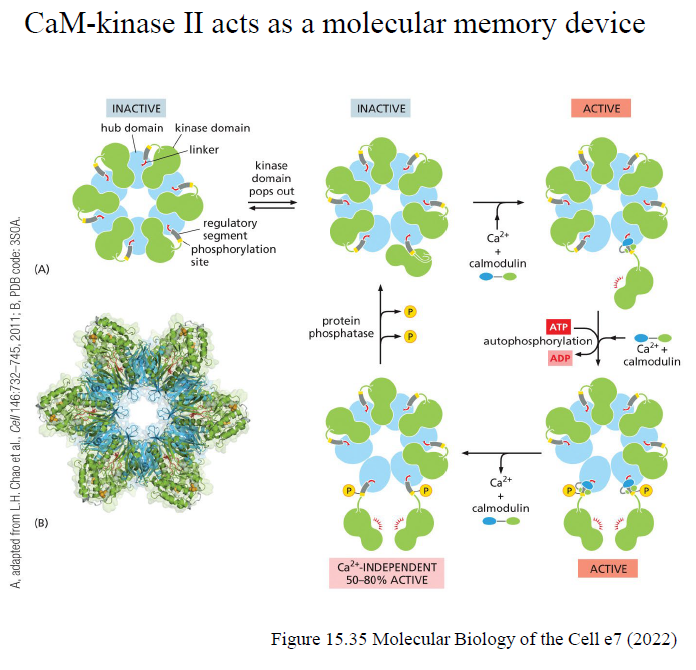
Hvordan aktiveres CaM-kinase II, og hvorfor kaldes det et “molecular memory device”?
Ca²⁺/calmodulin aktiverer CaM-kinase II.
Kinase kan derefter autophosphorylere sig selv og forblive delvist aktiv selv efter Ca²⁺-niveauet falder.
Dette giver en form for cellulær “hukommelse”.
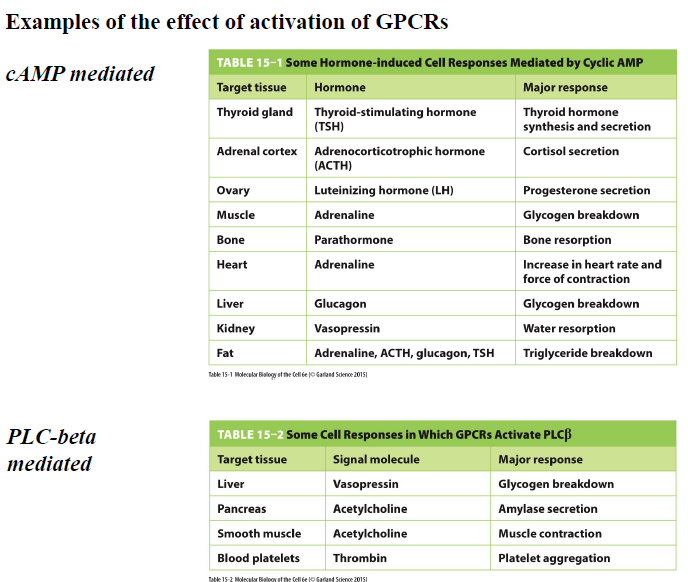
Hvilke typer cellulære responser kan GPCR pathways mediere via cAMP eller PLCβ?
GPCR signaling kan regulere mange forskellige responser afhængigt af væv og pathway.
cAMP-medierede responser:
glykogennedbrydning
øget hjertefrekvens
hormonsekretion
triglyceridnedbrydning
PLCβ-medierede responser:
muskelkontraktion
sekretion
platelet aggregation
Ca²⁺-signaling
Hvordan kan GPCRs signalere ud over cAMP og PLCβ pathways?
GPCRs kan:
direkte aktivere ionkanaler
indirekte regulere ionkanaler via cyclic nucleotides eller PKA
aktivere small GTPases som Rho-familien og dermed regulere actin-cytoskelettet
Hvordan forstærkes signaler i GPCR signaling pathways?
GPCR pathways bruger enzymatiske kaskader og second messengers til signal amplification.
Et enkelt aktiveret receptorprotein kan aktivere mange G-proteiner, som aktiverer mange downstream molekyler og giver et stort cellulært respons.
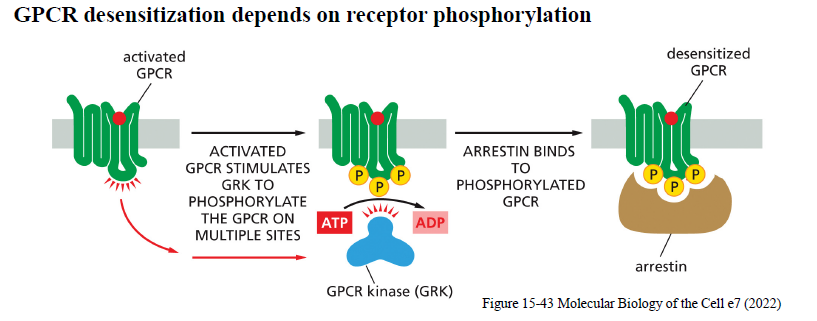
Hvordan desensitiseres GPCRs?
Aktiverede GPCRs fosforyleres af GPCR kinases (GRKs).
Arrestin binder derefter til den fosforylerede receptor:
forhindrer interaktion med G-proteiner
fremmer receptor-endocytose
Dette reducerer signaling.
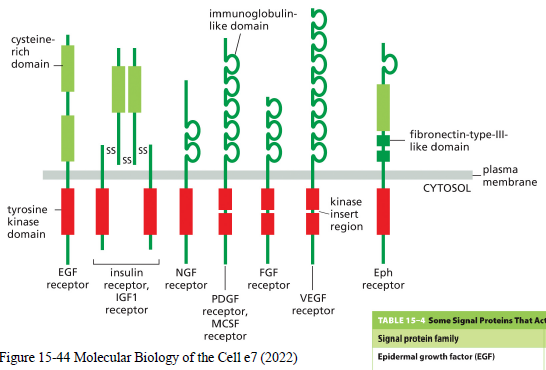
Hvad karakteriserer receptor tyrosine kinases (RTKs)?
RTKs er enzyme-coupled receptorer med en intracellulær tyrosine kinase domain.
Ligandbinding aktiverer receptoren og fører ofte til:
cellevækst
proliferation
survival
differentiering
Eksempler:
EGF receptor
insulin receptor
VEGF receptor
PDGF receptor
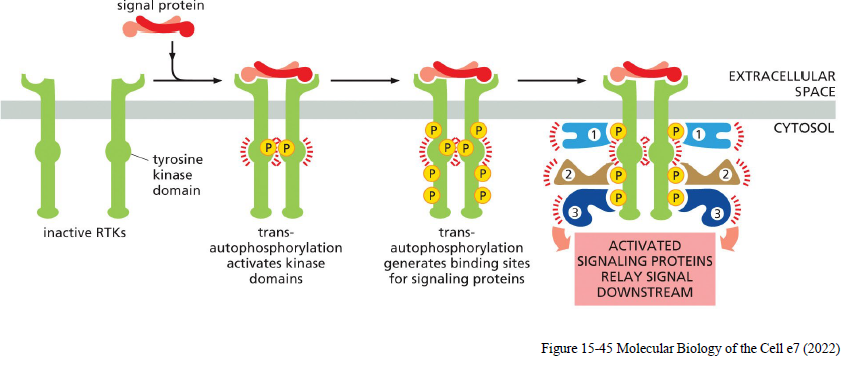
Hvordan aktiveres receptor tyrosine kinases (RTKs)?
Ligand binder receptorer
RTKs dimeriserer eller rearrangeres
Receptorerne trans-autophosphorylerer hinanden på tyrosiner
Phosphotyrosiner fungerer som docking sites for signaling proteins
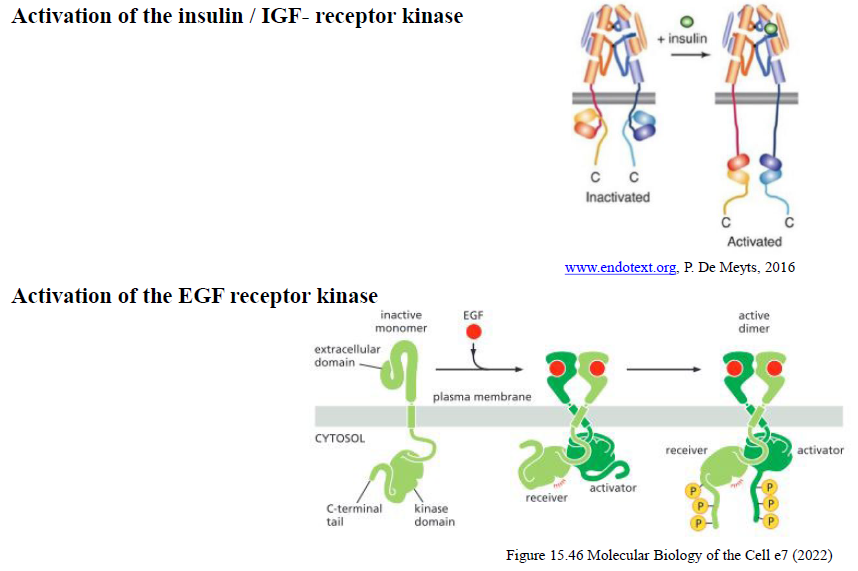
Hvordan adskiller aktivering af insulinreceptoren sig fra EGF-receptoren?
Insulinreceptoren eksisterer allerede som en dimer før ligandbinding.
EGF-receptoren findes som monomer og dimeriserer først efter ligandbinding.
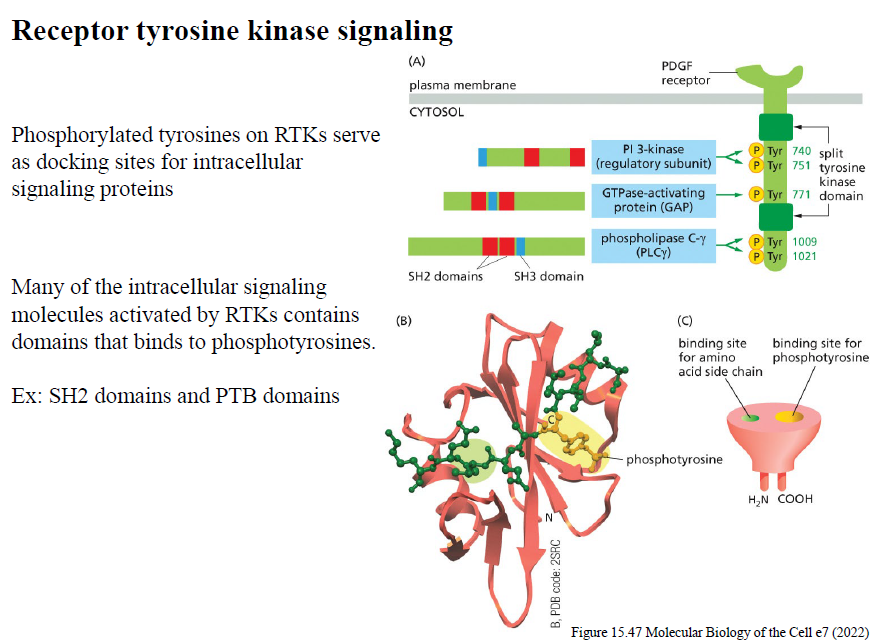
Hvad er funktionen af SH2- og PTB-domæner i RTK signaling?
SH2- og PTB-domæner binder phosphotyrosiner på aktiverede RTKs og rekrutterer intracellular signaling proteins til receptoren.
Hvordan aktiveres Ras downstream for RTKs?
Adapterproteinet Grb2 binder phosphotyrosiner på RTK via SH2-domain
Grb2 rekrutterer Sos via SH3-domain
Sos fungerer som Ras-GEF
Ras udskifter GDP med GTP og aktiveres
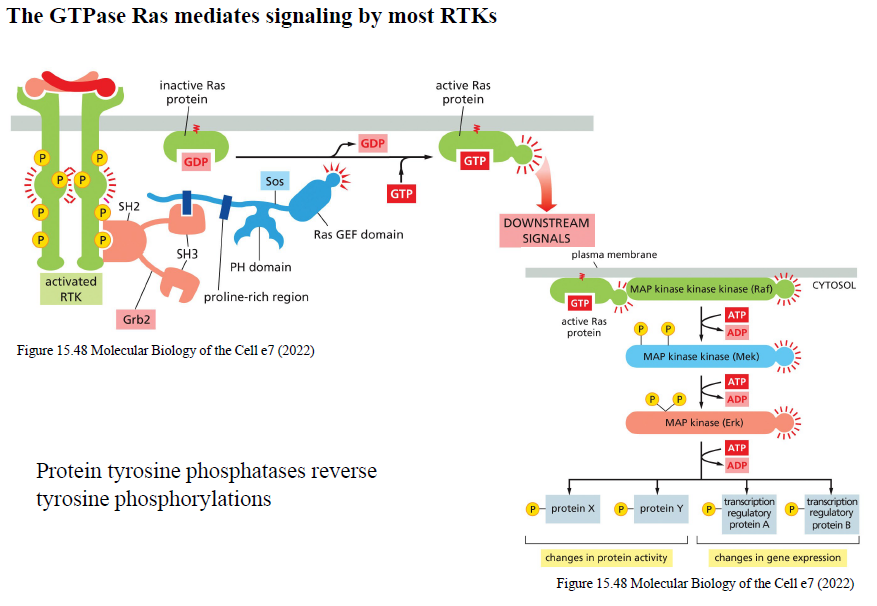
Beskriv Ras/MAPK signaling pathway downstream for RTKs.
Aktivt Ras-GTP aktiverer Raf (MAPKKK) → Raf aktiverer MEK (MAPKK) → MEK aktiverer ERK (MAPK).
ERK fosforylerer proteiner og transcription factors og ændrer:
proteinaktivitet
genekspression
Hvad er funktionen af protein tyrosine phosphatases?
De fjerner fosfatgrupper fra phosphotyrosiner og modvirker RTK signaling.
Hvilke funktioner har forskellige familier i Ras-superfamilien af monomere GTPaser?
Ras: RTK signaling
Rho: regulering af cytoskelet
Rab: vesikeltrafik
ARF: coat assembly på vesikler
Ran: nuklear transport og mitose
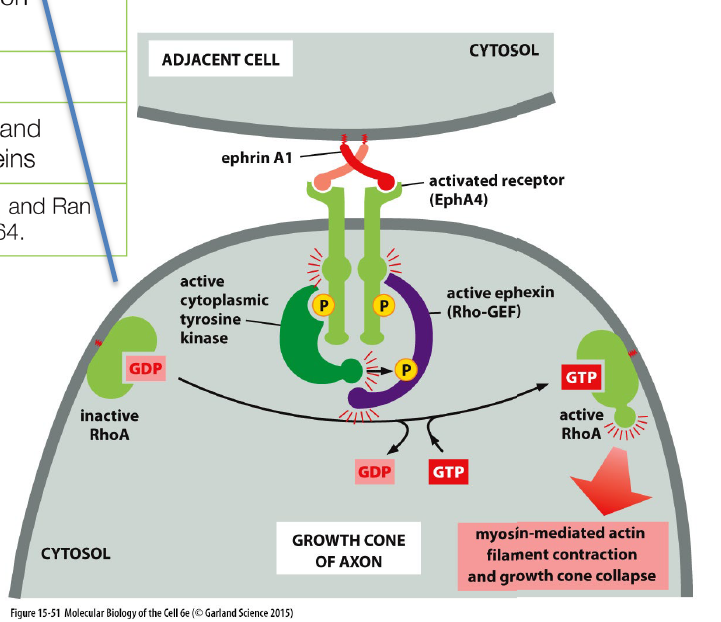
Hvordan regulerer Rho-familien cytoskelettet?
Receptorer kan aktivere Rho-GEFs, som aktiverer Rho GTPases (fx RhoA).
Aktivt Rho regulerer actin-cytoskelettet og påvirker:
cellebevægelse
kontraktion
growth cones
celleform
Hvordan kan signal duration, intensity og localization påvirke cellulære responser downstream for RTKs?
Det samme signaling pathway kan give forskellige responser afhængigt af:
hvor stærkt signalet er
hvor længe det varer
hvor i cellen signalet findes
Eksempel:
transient ERK-aktivering → proliferation
sustained ERK-aktivering → differentiering
Hvad er problemet ved at måle signaling i pooled cellepopulationer?
Average-signaler kan skjule store forskelle mellem individuelle celler.
En gradvis respons i en population kan faktisk skyldes:
nogle celler er fuldt aktiverede
andre er helt inaktive
Hvordan bruges FRET og BiFC til at studere signaling?
FRET og BiFC bruges til at måle:
protein-protein interaktioner
konformationsændringer
signaling dynamics i levende celler
De fungerer via ændringer i fluorescens.
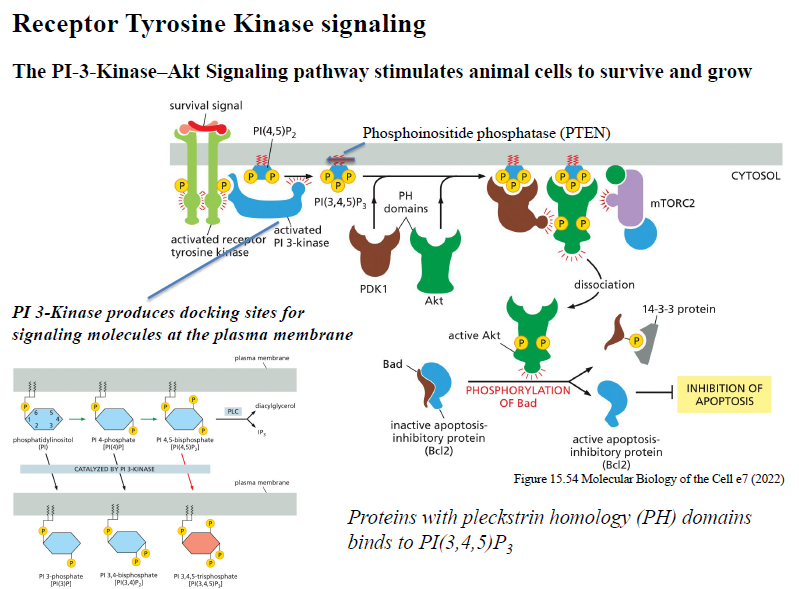
Beskriv PI3K-Akt signaling pathway downstream for RTKs.
RTK aktiverer PI3K
PI3K omdanner PIP2 → PIP3
PH-domain proteiner som Akt rekrutteres til membranen
Akt aktiveres af fosforylering
Akt fremmer survival og vækst ved at hæmme apoptose og stimulere growth pathways
PTEN modvirker pathwayet ved at fjerne fosfat fra PIP3.
Hvordan stimulerer Akt cellevækst via mTOR?
Akt inhiberer Tsc2, som normalt hæmmer Rheb.
Aktivt Rheb-GTP aktiverer mTORC1, som stimulerer:
proteinsyntese
ribosomproduktion
cellevækst
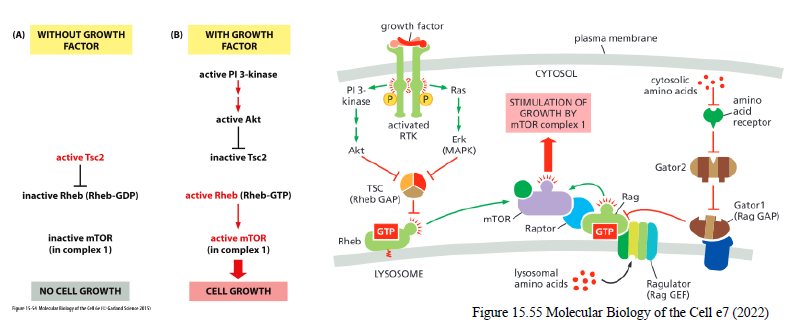
Hvordan inaktiveres receptor tyrosine kinases (RTKs)?
RTKs kan inaktiveres via:
endocytose + dephosphorylering/recycling
endocytose efterfulgt af lysosomal nedbrydning
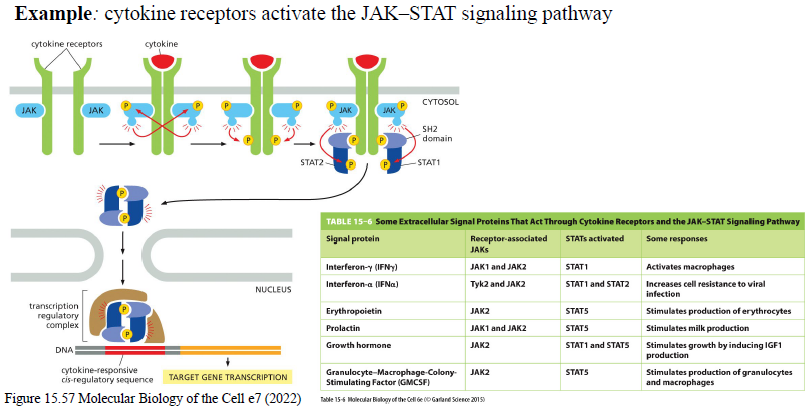
Beskriv JAK-STAT signaling pathway.
Cytokin binder receptor
Receptor-associerede JAKs aktiveres
JAKs fosforylerer receptoren
STATs binder phosphotyrosiner og fosforyleres
STATs dimeriserer
STAT-dimer går til nucleus og regulerer genekspression
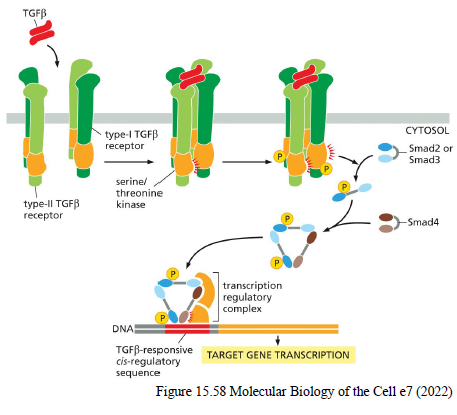
Beskriv TGFβ/Smad signaling pathway.
GFβ binder receptor serine/threonine kinases
Receptorer fosforylerer Smad2/3
Smads binder Smad4
Smad-komplekset går til nucleus
Genekspression ændres
Inhibitory Smads (Smad6/7) hæmmer pathwayet via negativ feedback.
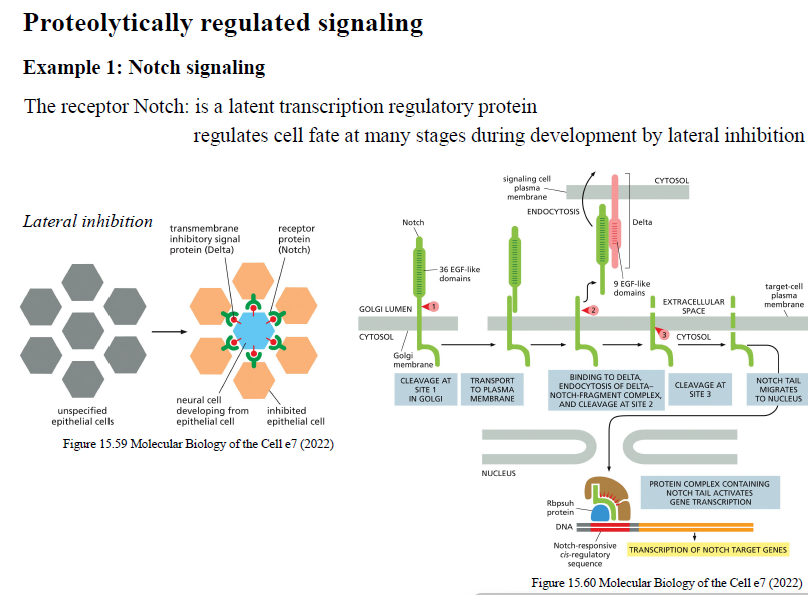
Hvordan fungerer Notch signaling?
Notch aktiveres ved kontakt mellem naboceller via Delta.
Ligandbinding fører til proteolytisk kløvning af Notch, så receptorens intracellulære del frigives og går til nucleus, hvor den regulerer target genes.
Notch signaling er vigtig for:
celle fate determination
lateral inhibition under udvikling.
Hvad er forskellen mellem canonical og non-canonical Wnt signaling?
Canonical Wnt pathway
Regulerer stabiliteten af β-catenin.
Uden Wnt nedbrydes β-catenin.
Med Wnt stabiliseres β-catenin og aktiverer target gene transcription.
Non-canonical Wnt pathway
Regulerer især:
cellepolaritet
actin-cytoskelet
cellebevægelse
og involverer ofte Rho-familie GTPaser og Ca²⁺-signaling.
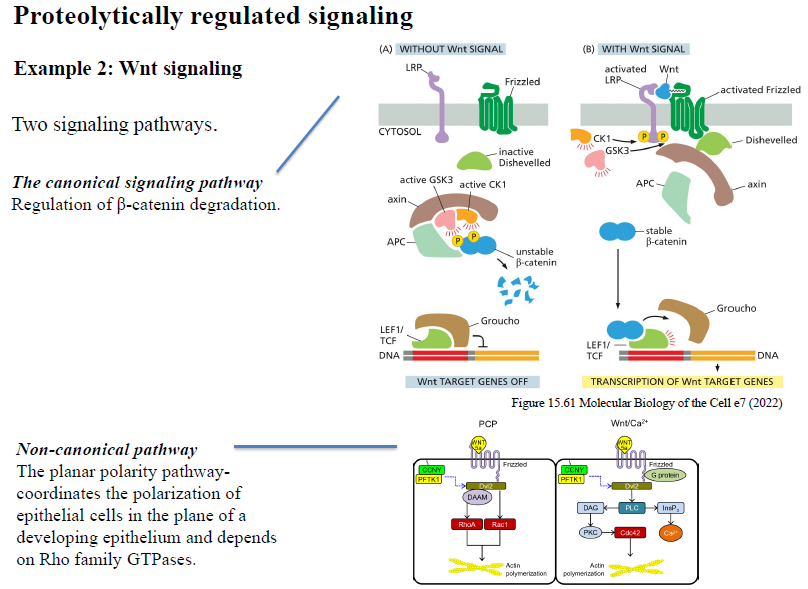
Hvordan kan Wnt signaling reguleres extracellulært?
Wnt signaling kan reguleres ved:
soluble Wnt binders
co-receptor binders
fjernelse af lipid/fedtsyre fra Wnt
regulering af receptor-nedbrydning
Disse mekanismer kan enten hæmme eller forstærke signaling.
Beskriv NFκB signaling pathway.
NFκB holdes normalt inaktivt i cytosol bundet til IκB.
Ved signalering:
IKK-komplekset fosforylerer IκB
IκB ubiquitinyleres og nedbrydes
NFκB frigives
NFκB translokerer til nucleus
NFκB aktiverer target genes
Negativ feedback kan skabe oscillationer i NFκB signaling.
Hvordan fungerer nuclear receptors?
Nuclear receptors er ligand-aktiverede transcription regulators for små hydrofobe molekyler som:
steroidhormoner
thyroidhormoner
retinoider
vitamin D
Ligandbinding aktiverer receptoren, som binder DNA og regulerer genekspression.
Hvordan overlapper signaling pathways fra GPCRs og RTKs?
GPCRs og RTKs kan aktivere mange af de samme downstream pathways, fx:
MAPK pathway
PI3K/Akt pathway
PLC/Ca²⁺ pathway
Signal integration mellem pathways er vigtig for at bestemme det endelige cellulære respons.
Hvad er cytoskelettets overordnede funktioner, og hvilke tre hovedtyper af filamenter består det af?
Cytoskelettet fungerer som cellens “skelet og muskler” og bruges til at:
opretholde struktur og mekanisk styrke
ændre celleform
celledeling
bevægelse af celler og organeller
De tre hovedtyper af filamenter er:
aktinfilamenter (mikrofilamenter)
mikrotubuli
intermediære filamenter
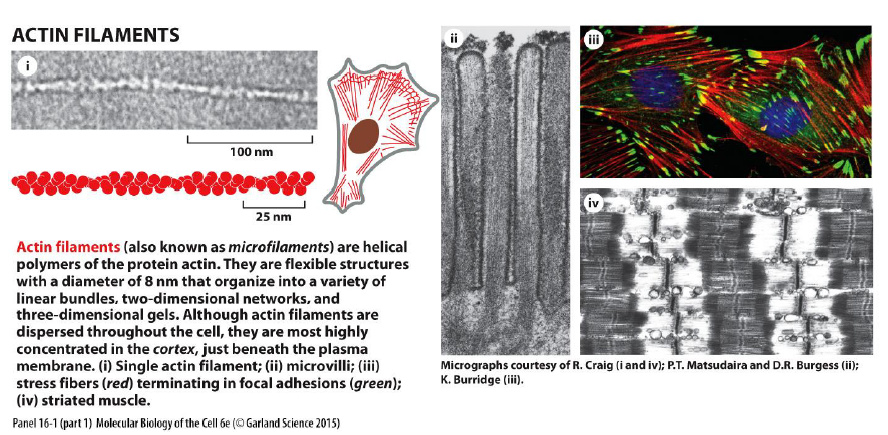
Hvad karakteriserer aktinfilamenter?
Aktinfilamenter er fleksible, helikale polymerer af aktin (~8 nm). De findes især i cortex lige under plasmamembranen.
Figuren viser at aktin:
kan danne bundter og netværk
findes i mikrovilli
danner stressfibre ved fokale adhæsioner
indgår i filopodia og lamellipodia
Aktin er derfor særligt vigtigt for cellebevægelse og dynamiske formændringer.
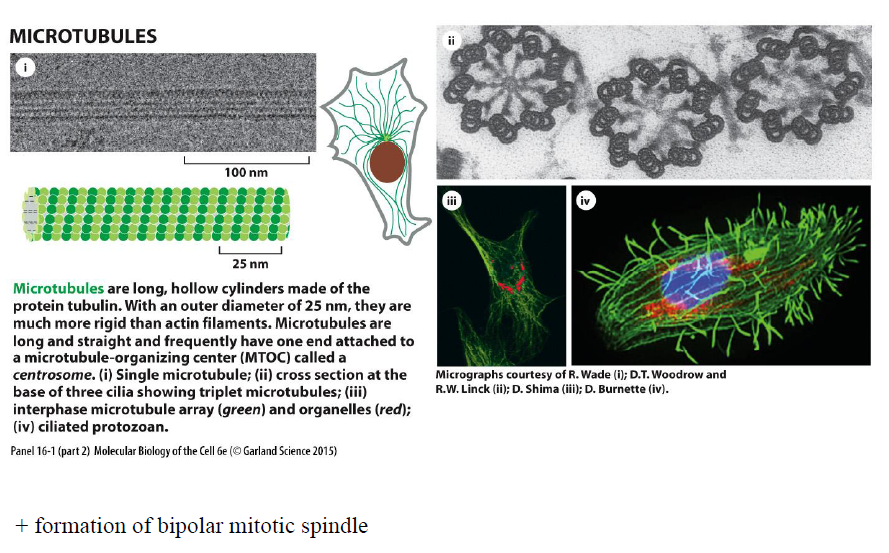
Hvad karakteriserer mikrotubuli?
Mikrotubuli er lange, hule tubulinrør (~25 nm) som er mere stive end aktinfilamenter.
Figuren viser at de:
organiseres fra et centrosom/MTOC
findes i cilier
hjælper med intracellulær transport
danner den bipolære mitotiske spindel ved celledeling
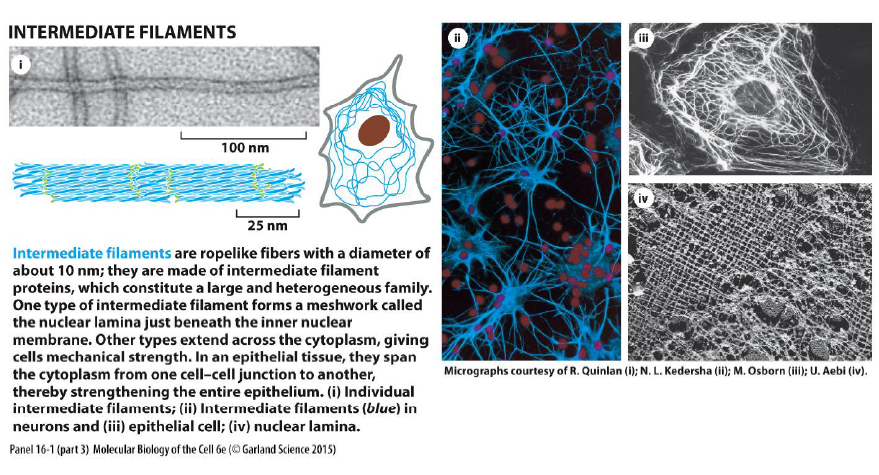
Hvad karakteriserer intermediære filamenter?
Intermediære filamenter er reb-lignende fibre (~10 nm), som giver mekanisk styrke.
Figuren viser at de:
spænder gennem cytoplasmaet
forbinder celle-celle junctions
styrker epithelvæv
danner nuclear lamina under kernemembranen
De er mindre dynamiske end aktin og mikrotubuli.
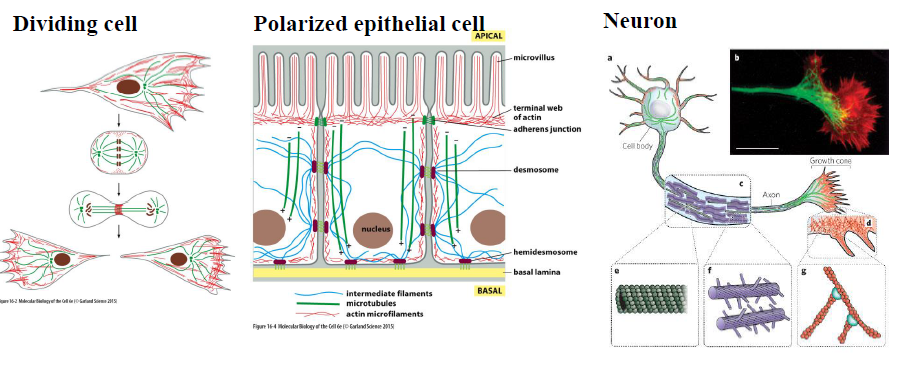
Hvordan er cytoskelettet organiseret forskelligt i forskellige celletyper?
Cytoskelettet tilpasses cellens funktion:
Delende celler: mikrotubuli organiseres i den mitotiske spindel
Polariserede epitelceller: aktin findes apikalt ved mikrovilli, intermediære filamenter forbinder desmosomer, og mikrotubuli hjælper med polaritet
Neuroner: mikrotubuli organiseres i axonet, mens aktin findes i growth cones
Figuren understreger, at cytoskelettet bestemmer cellulær organisering og polaritet.
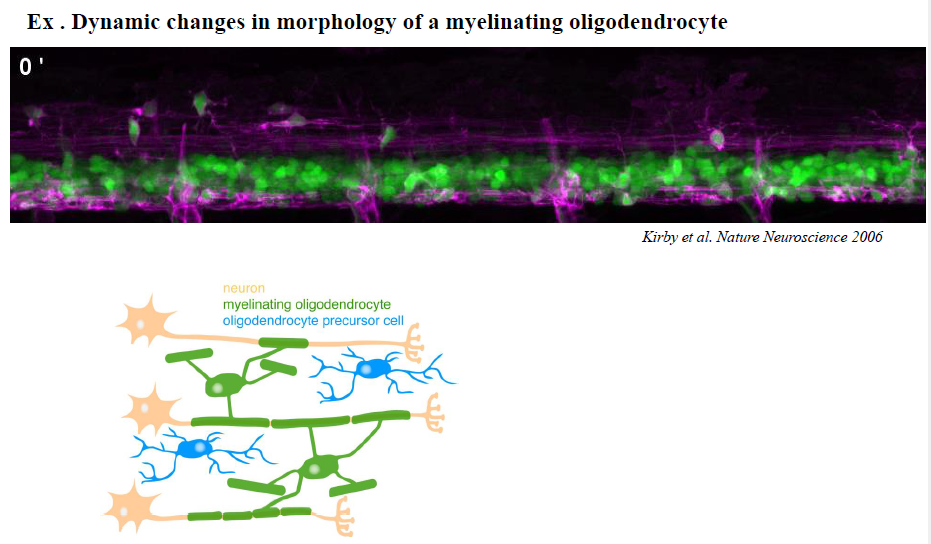
Hvordan bidrager aktindynamik til udvikling og myelinisering af oligodendrocytter?
Under modning ændrer oligodendrocytter form dramatisk for at kunne danne myelin omkring axoner.
Figurerne viser:
process extension og branching
overgang fra simple til komplekse lamellare strukturer
reorganisering af aktin under differentiering
Aktinpolymerisering er nødvendig i de tidlige stadier for udvækst og branching.
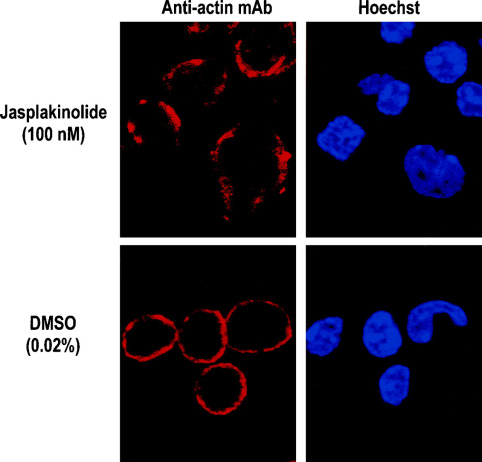
Hvad viser jasplakinolide-eksperimentet om aktin?
Jasplakinolide stabiliserer aktinfilamenter og hæmmer normal aktindynamik.
Eksperimentet viser derfor, at dynamisk reorganisering af aktin er nødvendig for korrekt modning af oligodendrocytter og normale morfologiske ændringer. (man kan ikke se billedet på slides, så det her er fra nettet)
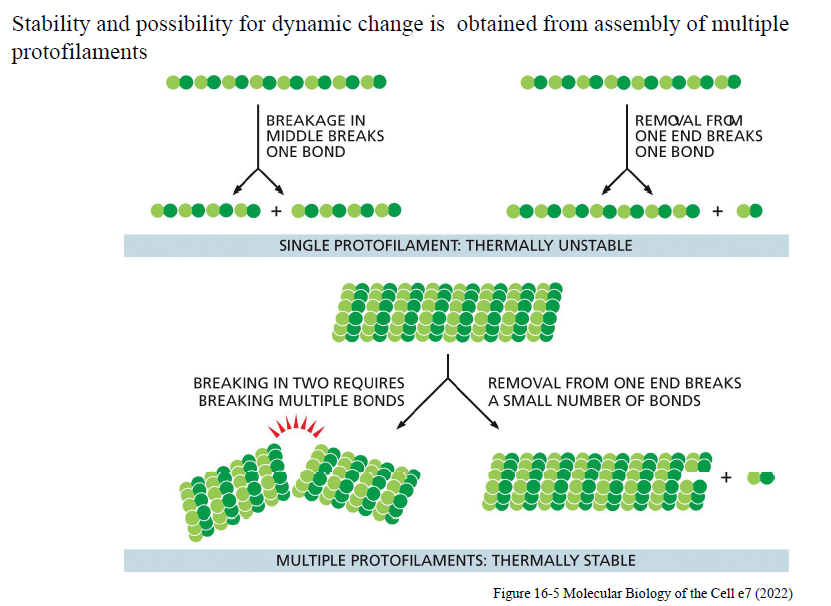
Hvorfor giver multiple protofilamenter både stabilitet og dynamik?
En enkelt protofilament er termisk ustabil, fordi et brud kun kræver at én binding brydes.
Når flere protofilamenter samles:
kræver brud mange bindinger → høj stabilitet
men enderne kan stadig vokse og krympe relativt let → dynamik
Figuren forklarer derfor, hvordan cytoskelet-filamenter både kan være stabile og samtidig hurtigt reorganiseres.
Hvordan er aktinfilamenter opbygget, og hvad betyder deres polaritet?
Aktinfilamenter dannes ved head-to-tail samling af aktinmonomerer, hvilket giver filamentet polaritet med:
en plus-end (+)
en minus-end (−)
Plus-enden vokser hurtigere end minus-enden. Filamentet består af ATP-bindende aktin, hvor ATP senere hydrolyseres til ADP efter inkorporering i filamentet.
Hos vertebrater findes α-actin især i muskelceller, mens β- og γ-actin findes i andre celletyper.
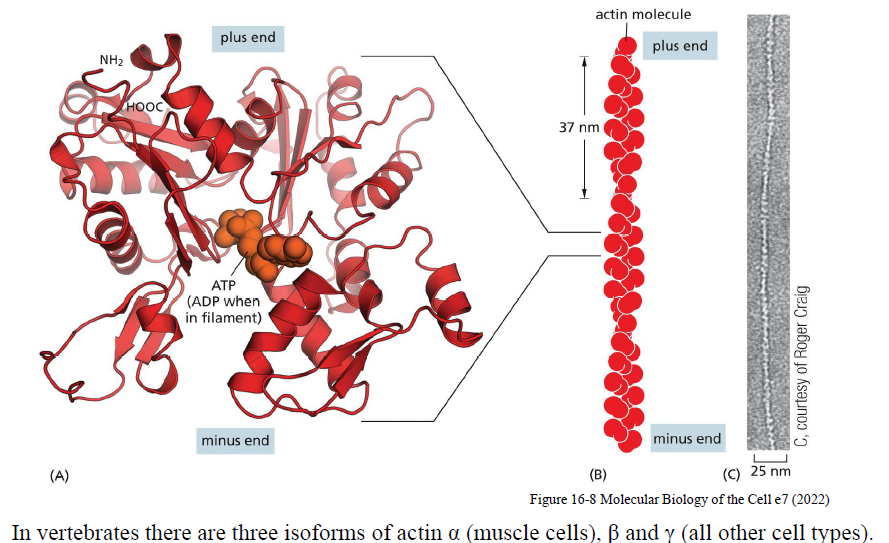
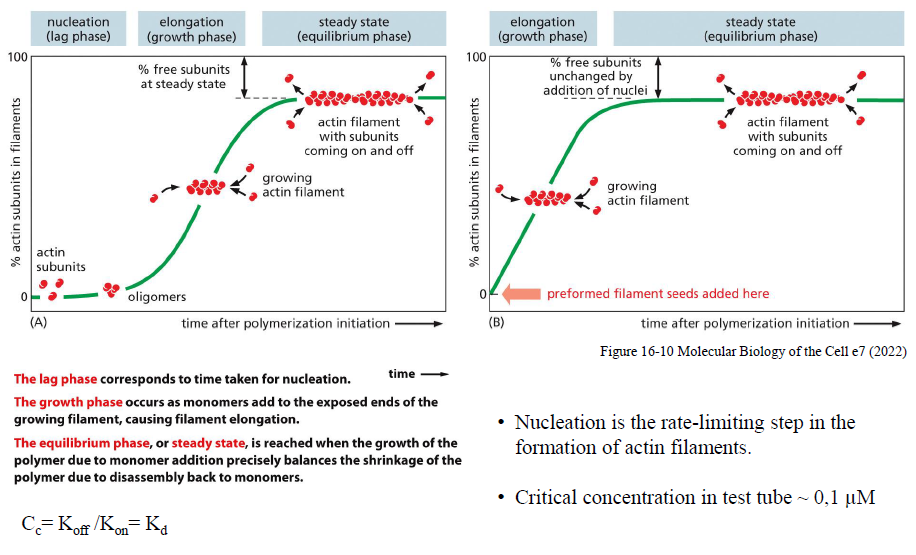
Hvilke faser indgår i aktinpolymerisering?
Aktinpolymerisering foregår i tre faser:
Nukleation (lag phase)
Små oligomerer dannes langsomt. Dette er den rate-limiterende fase.Elongation (growth phase)
Monomerer tilføjes hurtigt til filamentenderne, især plus-enden.Steady state (equilibrium phase)
Tilføjelse og tab af subunits balancerer hinanden.
Figuren viser også, at tilsætning af “preformed filament seeds” fjerner lag-fasen ved at omgå nukleation.
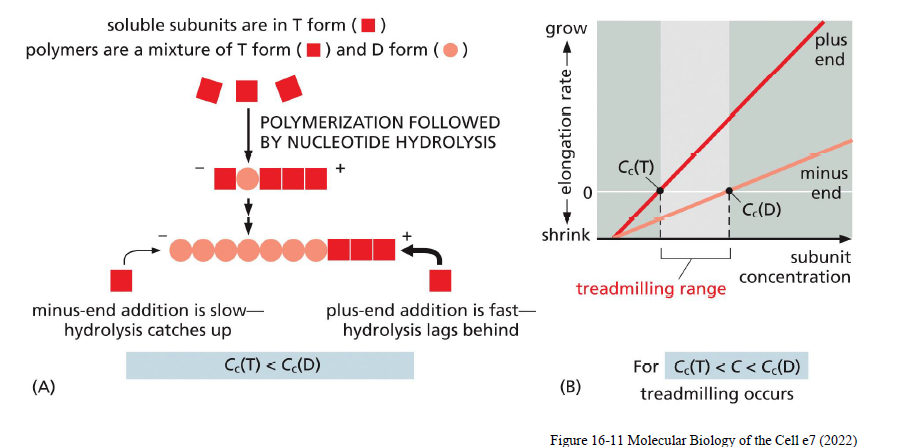
Hvad er treadmilling i aktinfilamenter?
Treadmilling sker når:
subunits tilføjes ved plus-enden
subunits fjernes ved minus-enden
samtidig med at filamentets samlede længde forbliver omtrent konstant.
Dette skyldes:
ATP-actin binder stærkere og favoriserer vækst
ATP hydrolyseres efter inkorporering
ADP-actin er mindre stabilt og dissocierer lettere
Treadmilling opstår når monomerkoncentrationen ligger mellem den kritiske koncentration for plus- og minus-end:
Cc(T) < C < Cc (D)
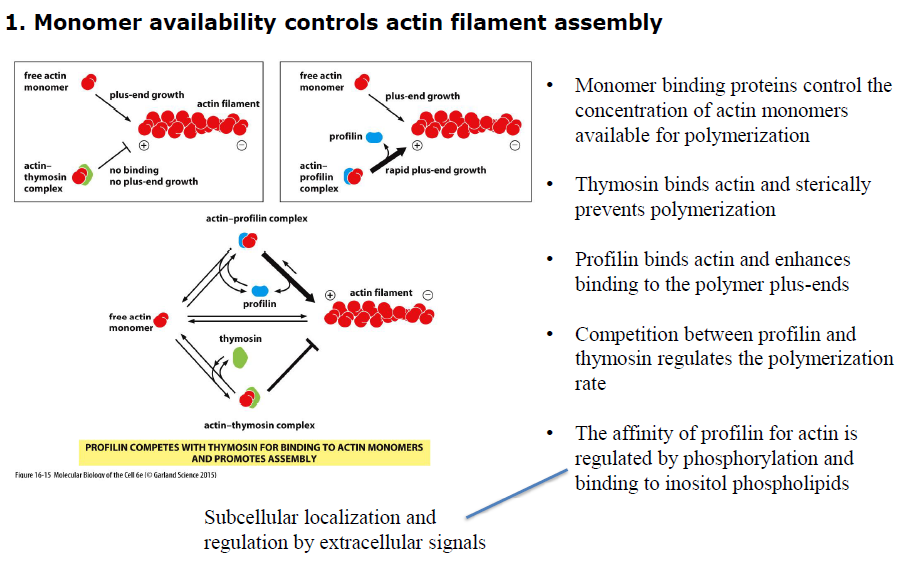
Hvordan regulerer thymosin og profilin aktinpolymerisering?
Begge proteiner binder aktinmonomerer, men har modsatte effekter:
Thymosin binder aktin og forhindrer polymerisering
Profilin binder aktin og fremmer binding til plus-enden
Konkurrencen mellem profilin og thymosin regulerer derfor hvor meget frit aktin der er tilgængeligt for filamentvækst.
Hvordan accelererer Arp2/3 og formin aktinpolymerisering?
Begge fungerer som nukleeringsfaktorer:
Arp2/3-komplekset danner forgrenede aktin-netværk
Formin danner lange, uforgrenede filamenter
Arp2/3 er vigtigt i lamellipodia, mens formin ofte bruges til stressfibre og filopodia.
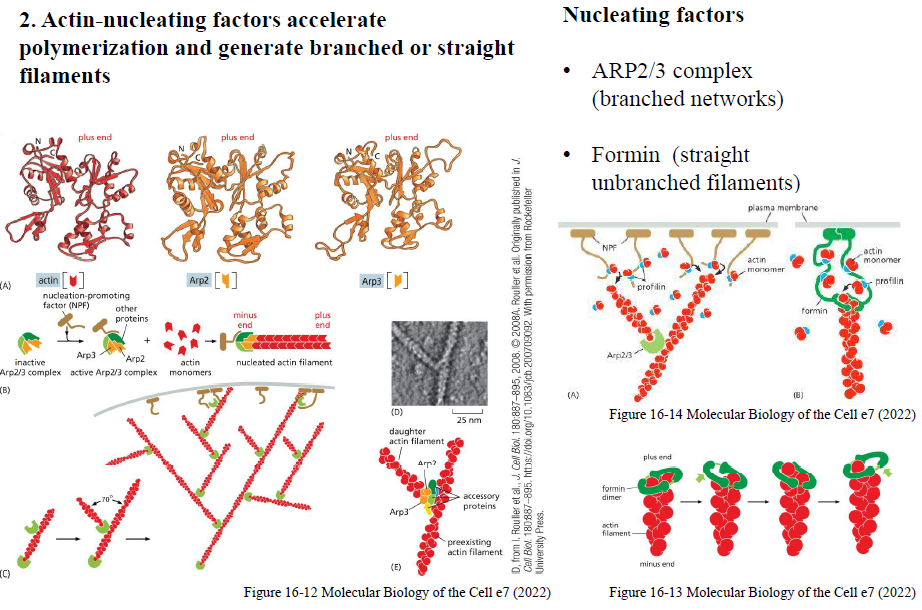
Hvordan påvirker capping- og severing-proteiner aktindynamik?
Capping-proteiner binder filamentender og regulerer vækst:
CapZ binder plus-enden
Tropomodulin binder minus-enden
Tropomyosin stabiliserer filamenter
Severing-proteiner fremmer depolymerisering:
Gelsolin kløver filamenter (aktiveres af højt Ca²⁺)
Cofilin binder især ADP-actin og destabiliserer filamenter
Balancen mellem stabiliserende og destabiliserende proteiner bestemmer filamentdynamikken.
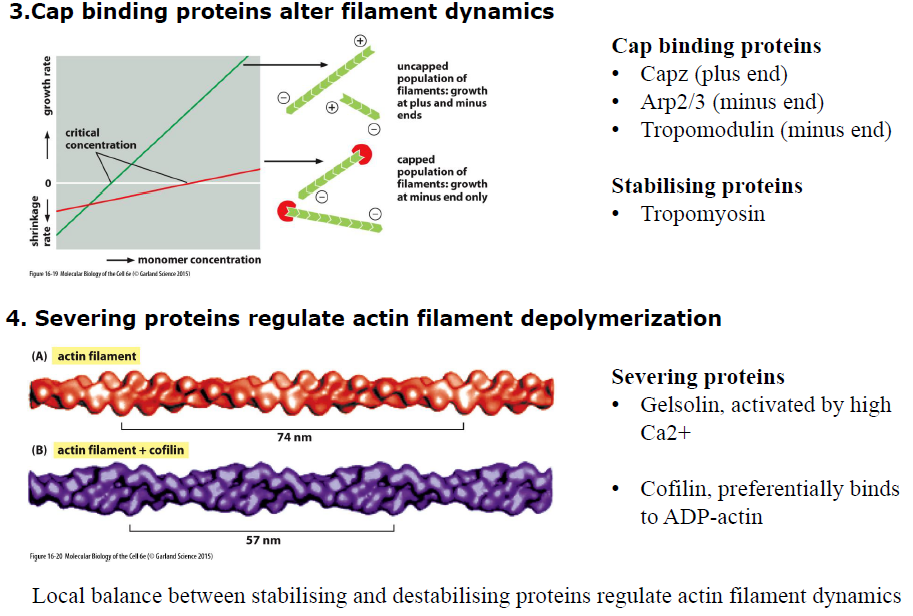
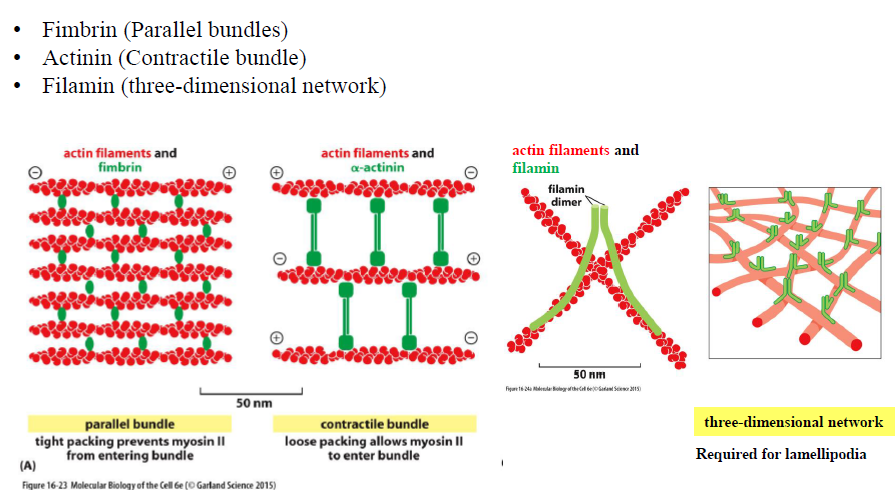
Hvordan bestemmer filament-bindende proteiner typen af aktin-netværk?
Forskellige crosslinking-proteiner giver forskellige netværk:
Fimbrin → tætte parallelle bundter
α-actinin → kontraktile bundter
Filamin → tredimensionelle netværk
Figuren viser at netværkets struktur bestemmer dets funktion.
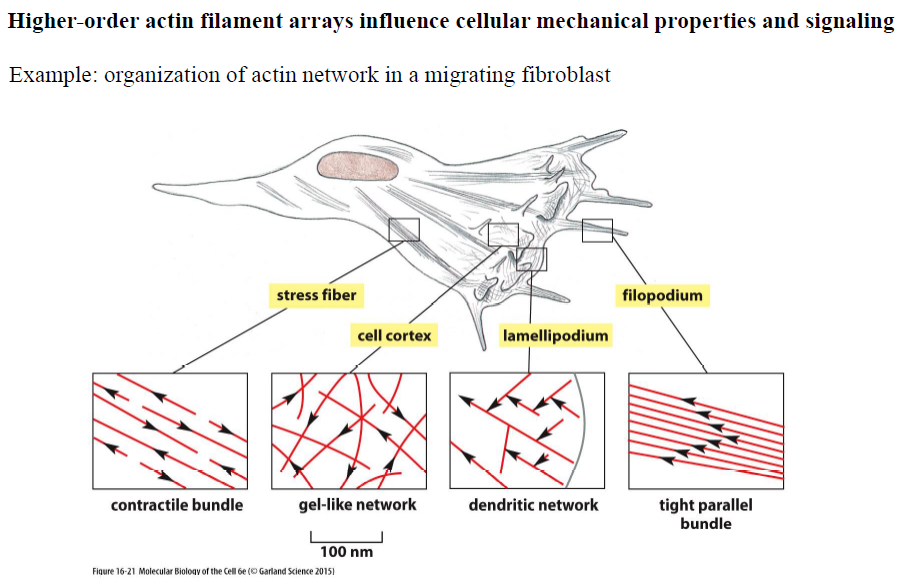
Hvordan er aktin organiseret i en migrerende fibroblast?
Forskellige områder af cellen indeholder forskellige aktin-strukturer:
Stress fibers → kontraktile bundter
Cell cortex → gel-lignende netværk
Lamellipodium → dendritisk/netværksstruktur
Filopodium → tætte parallelle bundter
Denne organisering gør det muligt for cellen både at bevæge sig og generere mekanisk kraft.
Hvilke hovedmekanismer bruger aktin-bindende proteiner til at regulere filamentdynamik?
De vigtigste mekanismer er:
Kontrol af monomer-tilgængelighed
Acceleration af polymerisering via nukleeringsfaktorer
Regulering af filamentender via capping-proteiner
Depolymerisering via severing-proteiner
Organisering af filamentnetværk via crosslinking-proteiner
Figuren samler de vigtigste aktin-regulatorer i ét overblik.
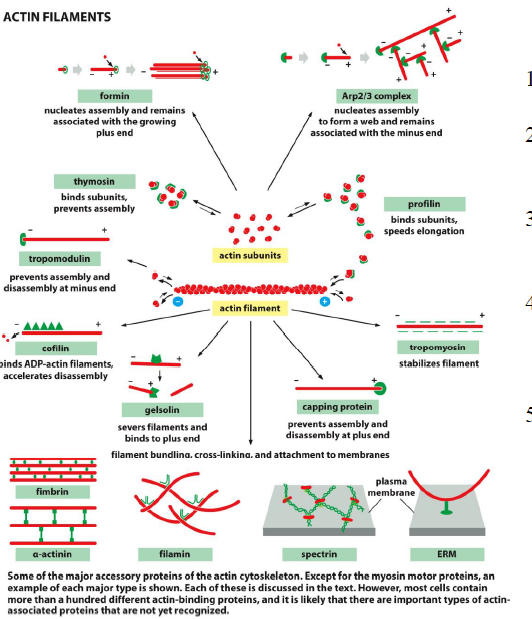
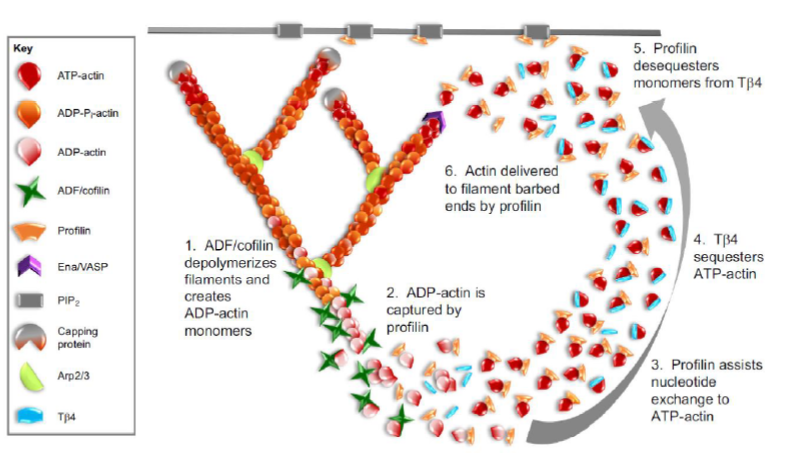
Hvordan samarbejder aktin-bindende proteiner om aktin turnover?
Figuren viser et cyklisk system:
ADF/cofilin depolymeriserer ADP-actin filamenter
Profilin binder ADP-actin monomerer
Profilin fremmer nucleotide exchange til ATP-actin
Thymosin β4 kan sequestrere ATP-actin
Profilin frigør monomerer fra thymosin
Profilin leverer ATP-actin til filamentets plus-end
Sammen gør disse proteiner hurtig og kontrolleret reorganisering af aktin-cytoskelettet mulig.
Hvilke værktøjer bruges til at studere aktindynamik?
Forskellige stoffer kan manipulere aktinfilamenter:
Phalloidin → binder og stabiliserer filamenter
Cytochalasin → capper plus-enden
Swinholide → kløver filamenter
Latrunculin → binder aktinmonomerer og forhindrer polymerisering
Visualisering af aktin kan ske med:
fluorescensmærket phalloidin
GFP-fusionsproteiner
LifeAct eller F-actin probes i levende celler
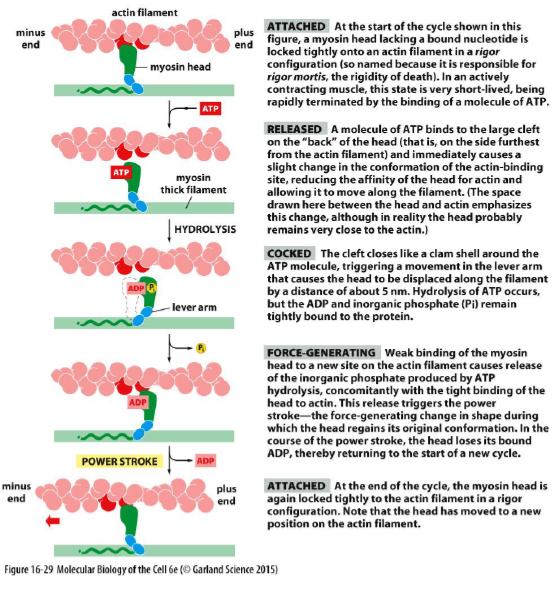
Hvordan genererer myosin kraft på aktinfilamenter?
Myosin er ATP-drevne motorproteiner der bevæger sig langs aktinfilamenter.
Kraft genereres gennem en cyklus:
ATP binder myosin → myosin slipper aktin
ATP hydrolyseres → myosinhovedet “cockes”
Myosin binder aktin igen
Frigivelse af Pi udløser power stroke
ADP frigives → ny ATP kan binde
Denne cyklus omdanner kemisk energi fra ATP til mekanisk bevægelse.
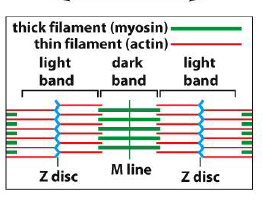
Hvordan er aktin og myosin organiseret i muskelceller?
Muskelfibre består af gentagne sarcomerer:
Tynde filamenter = aktin
Tykke filamenter = myosin
Aktin er forankret i Z-discs, mens myosin ligger centralt omkring M-line.
Når myosin glider langs aktin, forkortes sarcomeren og musklen kontraherer.
Figuren viser også:
mørke bånd = områder med myosin
lyse bånd = områder med kun aktin
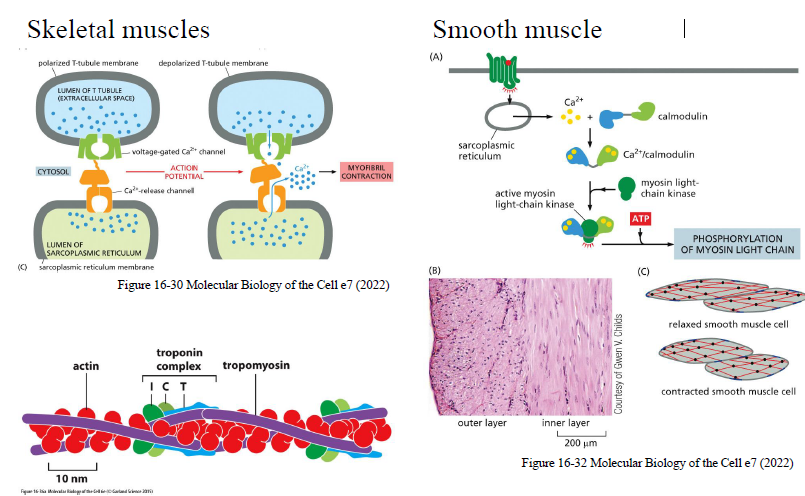
Hvordan initierer Ca²⁺ muskelkontraktion?
I skeletmuskel:
et aktionspotentiale åbner Ca²⁺-kanaler i SR
Ca²⁺ frigives til cytosolen
Ca²⁺ binder troponin
tropomyosin flyttes væk fra aktin
myosin kan nu binde aktin → kontraktion
I glat muskulatur:
Ca²⁺ binder calmodulin
Ca²⁺/calmodulin aktiverer MLCK
MLCK fosforylerer myosin light chain
myosin aktiveres og kontraktion starter
Hvilke funktioner har actin-myosin interaktioner i ikke-muskelceller?
I ikke-muskelceller bruges actin-myosin til:
cellemigration
kontraktion under cytokinese
intracellulær transport
Myosinaktivitet reguleres ofte ved fosforylering af myosin light chains.
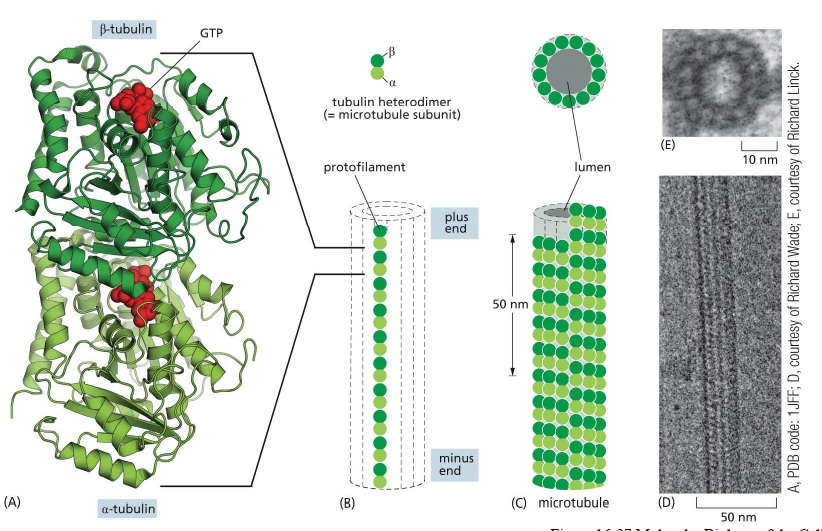
Hvordan er mikrotubuli opbygget?
Mikrotubuli er hule rør opbygget af protofilamenter.
Hvert protofilament består af α/β-tubulin heterodimere arrangeret head-to-tail, hvilket giver polaritet:
plus-end
minus-end
13 protofilamenter danner tilsammen et mikrotubulus-rør.
Tubulin binder GTP, som er vigtigt for polymerisering og stabilitet.
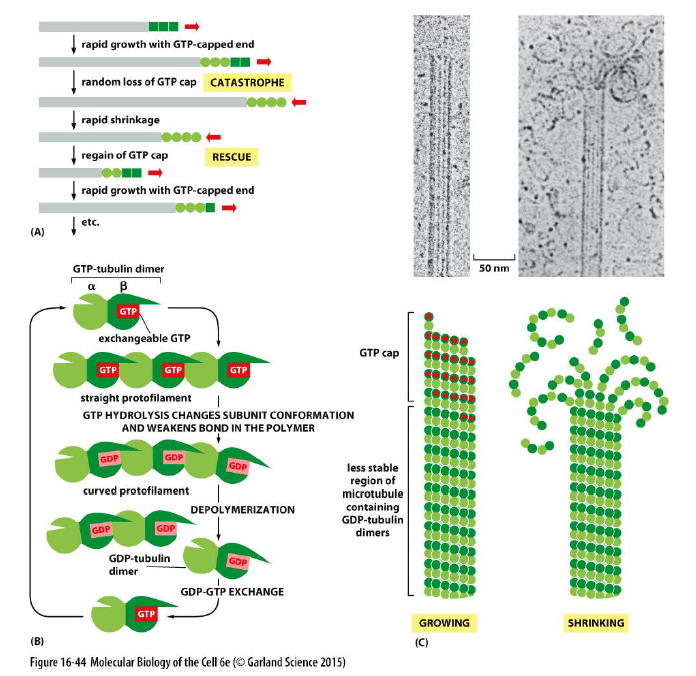
Hvad er dynamisk instabilitet i mikrotubuli?
Mikrotubuli skifter mellem:
vækst
hurtig depolymerisering
Dette kaldes dynamisk instabilitet.
En GTP-cap stabiliserer den voksende ende. Hvis GTP-cappen mistes:
GDP-tubulin destabiliserer filamentet
protofilamenter bøjer udad
hurtig depolymerisering (“catastrophe”) sker
Genetablering af GTP-cap kaldes “rescue”.
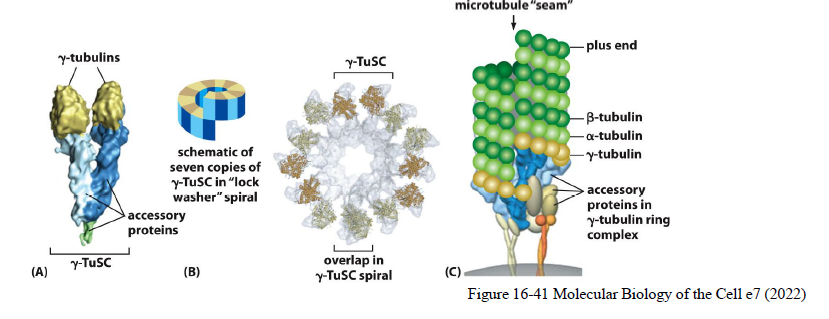
Hvordan nukleeres mikrotubuli?
γ-tubulin ring complexes (γ-TuRC) fungerer som nukleeringsskabeloner for mikrotubuli.
De findes ofte i centrosomet, hvorfra mikrotubuli vokser ud:
minus-end forankres ved centrosomet
plus-end vokser ud mod cytoplasmaet
Figuren viser også centrosomet som et microtubule-organizing center (MTOC).
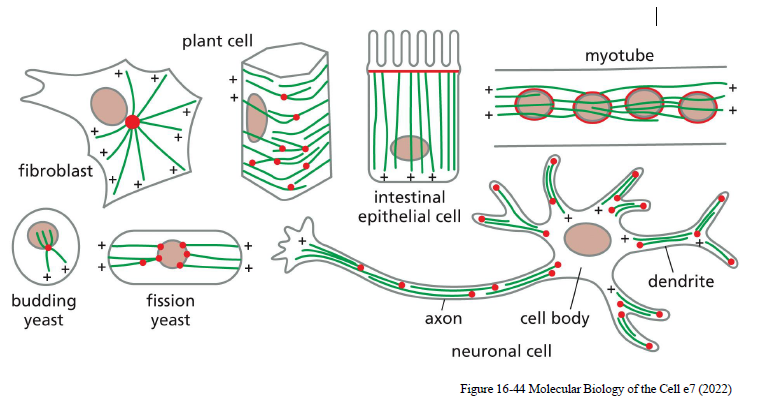
Hvordan er mikrotubuli organiseret forskelligt i forskellige celletyper?
Mikrotubuli-orientering afhænger af celletypen:
Fibroblaster: radial organisation fra centrosomet
Epithelceller: organiseret langs apikal-basal aksen
Axoner: plus-ender peger distalt
Dendritter: blandet orientering
Muskelceller: parallelle arrays langs cellens længdeakse
Denne organisering hjælper med:
intracellulær transport
polaritet
celledeling
formopretholdelse.
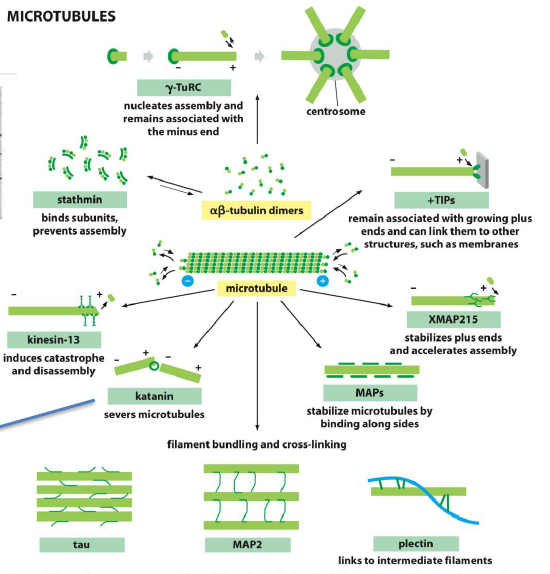
Hvordan regulerer mikrotubuli-bindende proteiner mikrotubuli-dynamik?
Plus-end-bindende proteiner kan enten stabilisere eller destabiliserer mikrotubuli.
Eksempler fra figuren:
Kinesin-13 → øger catastrophe og depolymerisering
XMAP215 → øger polymerisering og vækst ved plus-enden
Andre proteiner:
MAP2 og tau binder langs siden af mikrotubuli og stabiliserer dem
Stathmin binder tubulin-dimere og forhindrer assembly
Mikrotubuli-dynamik reguleres altså både ved:
ændring af væksthastighed
stabilisering
severing
sequestrering af tubulin.
Hvilke typer mikrotubuli-associerede proteiner findes der?
Forskellige MAPs har forskellige funktioner:
γ-TuRC → nukleerer mikrotubuli
+TIPs → binder voksende plus-ender
XMAP215 → fremmer assembly
Kinesin-13 → inducerer catastrophe
Katanin → severer mikrotubuli
Tau og MAP2 → stabiliserer og bundler mikrotubuli
Plectin → linker mikrotubuli til intermediate filaments
Stathmin → binder frie tubulin-dimere og hæmmer polymerisering
Figuren viser også, at severing kan skabe nye mikrotubuli-ender og dermed ændre netværksorganisationen hurtigt.
Hvordan påvirker post-translationelle modifikationer mikrotubuli?
Tubulin kan modificeres kemisk efter translation, hvilket påvirker mikrotubuli-egenskaber.
Eksempler:
acetylering
glutamylering
glycylation
tyrosinering
fosforylering
Disse modifikationer påvirker:
stabilitet
depolymerisering
hvilke motorproteiner der binder
motorproteiners hastighed
Hvordan kan man eksperimentelt undersøge mikrotubuli-dynamik?
Kemisk manipulation
Forskellige stoffer kan stabilisere eller nedbryde mikrotubuli:
Stoff | Effekt | Mekanisme |
|---|---|---|
Taxol (paclitaxel) | Stabiliserer mikrotubuli | Binder langs filamentet |
Nocodazole | Depolymeriserer | Binder tubulin-subunits |
Colchicine | Depolymeriserer | Capper filament-ender |
Taxol bruges derfor ofte til at forhindre dynamik, mens nocodazole og colchicine bruges til at nedbryde mikrotubuli-netværket.
Visualisering
Mikrotubuli kan visualiseres med:
GFP-tubulin
GFP-EB1 (+TIP protein)
EB1-markerede “comets” viser voksende plus-ender og bruges til at måle væksthastighed og dynamik.
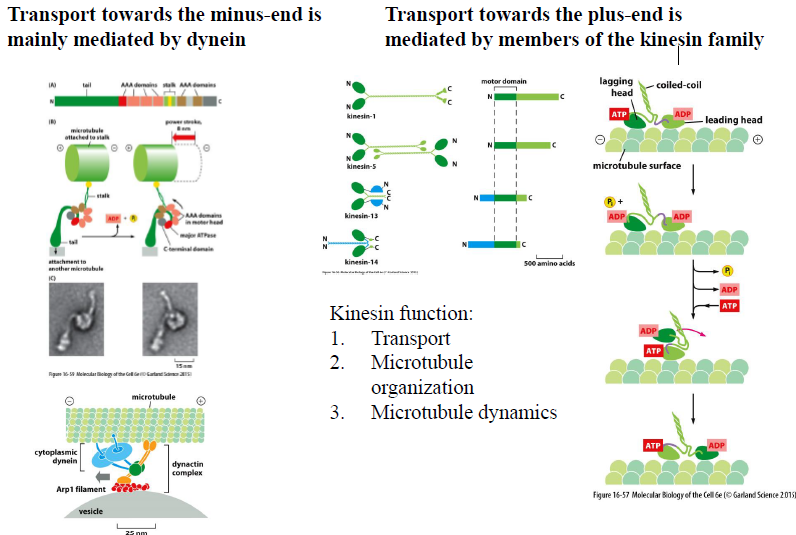
Hvordan fungerer motorproteiner på mikrotubuli?
Der findes to hovedtyper:
Motorprotein | Bevægelsesretning |
|---|---|
Dynein | Mod minus-end |
Kinesin | Mod plus-end |
Motorproteiner bruger ATP-hydrolyse til at “gå” langs mikrotubuli.
Kinesin
Kinesin bevæger sig typisk mod plus-enden via en “hand-over-hand” mekanisme:
ATP binder til leading head
Hals-regionen ændrer konformation
Det bageste hoved svinger frem
ATP hydrolyseres og cyklussen fortsætter
Funktioner
Kinesiner bruges til:
transport af vesikler og organeller
organisering af mikrotubuli
regulering af mikrotubuli-dynamik
Dynein samarbejder ofte med dynactin-komplekset for effektiv cargo-transport.
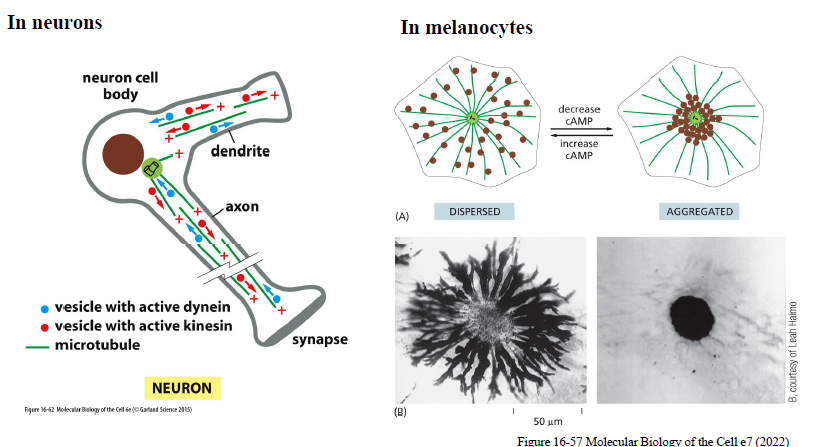
Hvordan bruges mikrotubuli og motorproteiner til intracellulær transport?
I neuroner
Axoner indeholder lange mikrotubuli-spor:
kinesin transporterer cargo mod synapsen (+ ende)
dynein transporterer cargo tilbage mod cellekroppen (− ende)
Dette er vigtigt, fordi neuroner er meget lange celler og kræver effektiv transport.
I melanocytter
Melanosomer kan enten:
være dispergerede i hele cellen
samles centralt
cAMP regulerer dette:
↑ cAMP → dispergering
↓ cAMP → aggregering
Transporten sker via motorproteiner på mikrotubuli.
Hvad er intermediate filaments?
Intermediate filaments er stærke cytoskelet-filamenter, der giver mekanisk styrke.
Der findes ca. 70 IF-gener opdelt i 5 klasser:
Type | Protein | Lokalisation |
|---|---|---|
I | Lamins | Nuklear lamina |
II | Vimentin | Mesenkymale celler |
III | Desmin, GFAP m.fl. | Muskel og glia |
IV | Keratiner | Epitel |
V | Neurofilamenter | Neuroner |
De er mere stabile end aktin og mikrotubuli.
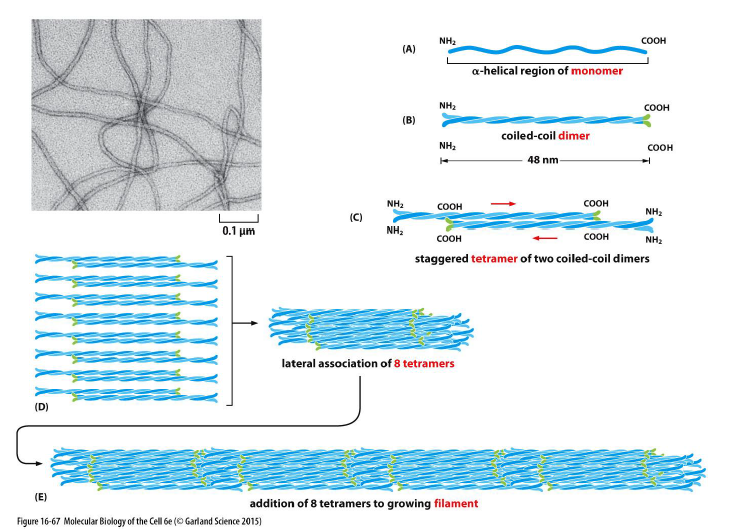
Hvordan er intermediate filaments opbygget?
Assembly foregår trinvis:
Monomer med α-helikal region
Coiled-coil dimer
To dimere danner antiparallelt tetramer
Tetramere pakkes lateralt
Lange filamenter dannes
Vigtige egenskaber:
ingen polaritet
binder ikke ATP eller GTP
meget fleksible og stærke
Manglende polaritet betyder også, at motorproteiner ikke går på IFs.
Hvad er intermediate filaments funktion?
Intermediate filaments giver mekanisk styrke til væv.
Eksempel:
keratin-filamenter kobles til desmosomer og hemidesmosomer i epithel
Dette hjælper celler med at modstå stræk og mekanisk stress.
Epidermolysis bullosa
Mutationer i keratiner eller linkerproteiner giver skrøbelig hud, hvor epidermis let løsner sig og danner blærer.
Hvordan forbindes cytoskelettet med kernen?
Plectin
Plectin forbinder:
intermediate filaments
aktin
mikrotubuli
adhæsionskomplekser
SUN-KASH komplekset
SUN-KASH proteiner spænder over kernemembranen og forbinder:
cytoskelettet i cytoplasma
nuklear lamina og kromosomer i kernen
Dette skaber mekanisk kobling mellem celleoverflade og nucleus.
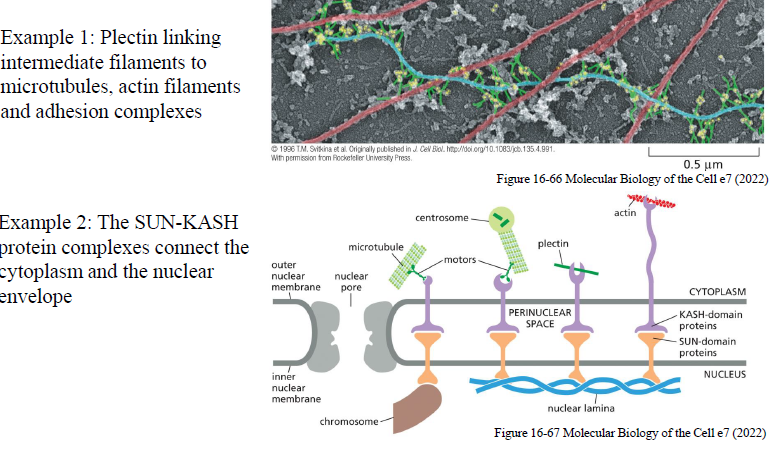
Hvorfor er koblingen mellem cytoskelet og nucleus vigtig?
Koblingen bruges til:
1. Nuclear positioning
Kernen placeres korrekt i:
migrerende celler
muskelceller
epithel
neuroner
2. Mechanosensing
Mekaniske signaler fra ECM kan overføres til nucleus og ændre genekspression.
Det gør, at celler kan reagere på:
stræk
stivhed
mekanisk belastning
Hvad er septiner?
Septiner er GTP-bindende filamentproteiner.
De:
danner hetero-hexamere eller octamere
organiserer sig til nonpolære filamenter
Funktioner:
compartmentalization af membraner
interaktion med aktin og mikrotubuli
vigtig for asymmetri i celler
Septiner er især vigtige ved:
cytokinese
budding i gær
polariserede celler
Hvordan etableres cellepolaritet?
C. elegans embryo
Sperm-entry bryder symmetrien → PAR-proteiner reorganiseres:
anterior PAR-proteiner samles i én ende
posterior PAR-proteiner i den anden
Dette etablerer anterior-posterior polaritet.
Epitelceller
Epitelceller opdeles i:
apikal domæne
basolateral domæne
Proteinkomplekser:
PAR
Crumbs
Scribble
hjælper med at etablere og vedligeholde denne polaritet.
Polaritet er vigtig for:
transportretning
vævsorganisation
cellefunktion.
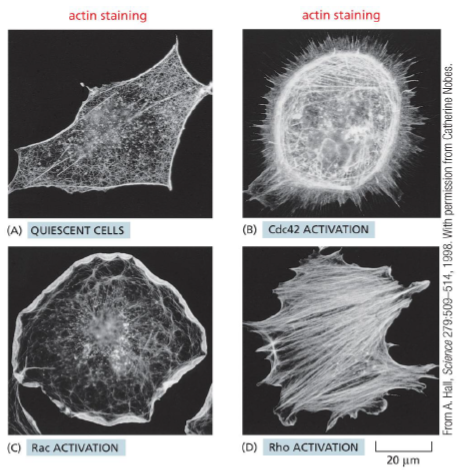
Hvordan regulerer Rho-familien aktin-cytoskelettet?
Rho-familiens små GTPaser fungerer som molekylære switches:
GDP-bundet = inaktiv
GTP-bundet = aktiv
Reguleres af:
GEFs → aktiverer (GDP → GTP)
GAPs → inaktiverer
Rho-GDI → holder Rho i cytosol og hæmmer aktivering
Tre centrale medlemmer
Protein | Primær effekt |
|---|---|
Cdc42 | Filopodia |
Rac | Lamellipodia / branched actin |
RhoA | Stress fibres og kontraktion |
Cdc42
Aktiverer:
WASP/WAVE
Arp2/3
→ danner forgrenede aktin-netværk og protrusioner.
Rac
Stimulerer protrusion og cellebevægelse via Arp2/3-medieret aktinpolymerisering.
RhoA
Aktiverer ROCK:
øger myosin aktivitet
hæmmer myosin phosphatase
stabiliserer stress fibres og focal adhesions
→ mere kontraktilitet.
Hvordan bevæger celler sig?
Cellemigration foregår i tre trin:
Trin | Cytoskelet-funktion |
|---|---|
1. Protrusion | Aktinpolymerisering skubber membranen frem |
2. Attachment | Integriner danner focal adhesions |
3. Traction | Actin-myosin kontraktion trækker cellen frem |
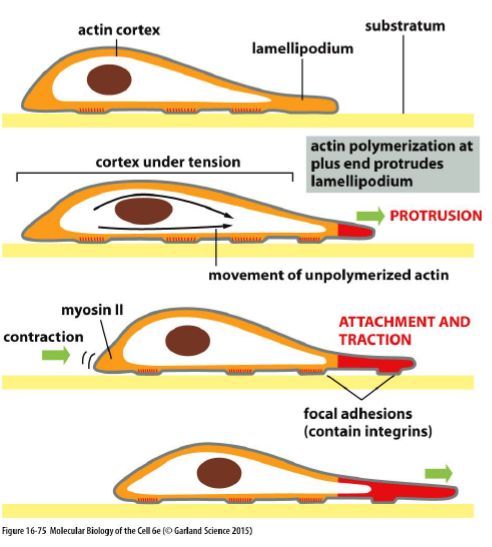
Hvordan styrer Rac og Rho rettet migration?
Ved kemotaksi i neutrofiler:
Front af cellen
Rac dominerer:
aktinpolymerisering
protrusion
lamellipodia
Bagende af cellen
Rho dominerer:
actin-myosin kontraktion
retraktion af bagenden
Signalgradienter fra chemoattractants skaber asymmetrisk aktivering af Rac og Rho.
Det giver:
en protruderende front
en kontraherende bagende
→ retningsbestemt bevægelse.