Vertebrate Zoology Exam 2
1/79
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced |
|---|
No study sessions yet.
80 Terms
Chondrichthyes Synapomorphies
Tesserate prismatic calcified cartilage in endoskeleton
Globular or stellate deposits of crystalline calcium in the superficial layers of the cartilage matrix
Placoid scales
Composed of a vascular (supplied with blood) inner core of pulp, a middle layer of dentine and a hard enamel-like outer layer of vitrodentine
Pelvic claspers
Present in males, used for internal fertilization
Holocephali
Ratfishes and Chimeras
Swim by gently flapping pectoral fins
Synapomorphies:
Gill openings covered by soft tissues
Teeth in the form of toothplates that are slowly replaced
First dorsal spine erectile
Clasping organ on head of males (tentaculum)
3 families, around 30 species, mostly deepwater marine
Elasmobranchii
Sharks, skates, rays
Synapomorphy:
Gill openings separate and uncovered
Ancient vs. Modern Sharks
Snout characters
Ancestral condition
Short, rounded snout
Long jaws
Located at front of head (terminal)
Modern condition
Long pointy snout
Short jaws
Located underneath head (subterminal)
Significance - long jaws are structurally weaker than short jaws; results in less powerful bites
Jaws
Upper and lower components
Palatoquadrate (upper)
Meckel’s cartilage (lower)
Hyomandibula supports the palatoquadrate
Jaw suspension
Ancestral condition:
Amphistylic - palatoquadrate is fixed to the braincase in front and back
Modern condition:
Hyostylic - palatoquadrate is not fixed to braincase; entirely supported by hyomandibula.
Result – protrusible upper jaw
Teeth
Ancient condition - cladodont teeth
Smooth-edged
Multi-cusped
Large central blade
Best suited for grasping prey and swallowing whole
Modern condition - serrated teeth
Serrated edges
Single-cusped
Enables gouging pieces from prey too large to be swallowed and slicing through tough skin
Neoselachii Characteristics
Characteristics:
Underslung or ventral mouth
Polyphodont dentition – can undergo serial tooth replacement
Ancient sharks – spiral replacement
Ampullae of Lorenzini - electroreception for detection of prey, possibly for navigation
Jelly-filled canals
Galeomorphi
Sharks with an anal fin
Hexacanthiformes
Sixgills, sevengills, and frilled sharks
Heterodontiformes
Bullhead and horned sharks
Lamniformes
Sand, tiger, goblin, thresher, basking, mackerel, mako, and great white sharks
Orectolobiformes
Carpet, blind, nurse, zebra, whale, and wobbegong sharks
Carcharhiniformes
Cat, hound, leopard, soupfin, tiger, gray, blue, lemon, and hammerhead sharks
Squalomorphi
Sharks without an anal fin
Pristiophoriformes
Sawsharks
Squaliformes
Spiny dogfish, bramble, and sleeper sharks
Squatiniformes
Angel sharks
Batoidea
Skates and rays
Synapomorphies:
Dorsoventrally-flattened bottom dwellers large pectoral fins; dorsal fins reduced to absent; caudal fin reduced
Large dorsal spiracle
External gill openings are on the ventral side of the body; water (for breathing) is taken through the large spiracle on the dorsal side
Teeth usually flattened and united to form a pavement for crushing mollusks; also feed on crustaceans, and occasionally, fish
Rajiformes
Sawfishes, guitarfishes, rays, and skates
General biology of elasmobranchs
Large bodied
Feeding
Many sharks, but not the rays and skates, are specialized as piscivorous predators
In these sharks, the lower teeth are usually spiky, the upper teeth blade-like
A few filter feeders (megamouth, whale shark, basking shark, manta ray)
The rays and skates are usually hard shelled invertebrate feeders: teeth from multiple rows form pavement
Habitat
Marine (5% found in freshwater, as casual visitors or lifelong residents)
Bull sharks in Lake Nicaragua
Amazon stingray
Sensory abilities
Have relatively large brains
Very good olfactory capabilities
Good vision (particularly night vision), hearing
Electroreception
Location of prey, in near range
Possibly for orientation in migration
Physiology
Resting metabolic rate 1/3-1/2 that of bony fish of comparable size
Active metabolic rate of dogfish only 3X resting, vs. 10X for bony fish
Lamnidae (e.g., great whites) have regional endothermy - able to warm their muscles, stomach, viscera, brain and eyes to a temperature above that of the ambient sea water; rete mirable
Slow growth, late age at maturity, long lifespan (spiny dogfish mature at 35 years off of British Columbia)
Reproduction
Internal fertilization
Low fecundity
Direct development, development slow
Three reproductive modes
Ancestrally, oviparity (lay eggs) (30%-40% of extant spp)
Young retained in uterus, nourished by yolk sac: ovoviviparity (when yolk sac is depleted, mother gives birth) (in some, oophagy (young eats unfertilized eggs in the uterus) or embryophagy (young eat other young in the uterus) - a.k.a. intrauterine cannibalism)
Viviparity (mother gives birth to live sharks and feeds them) - maternal nourishment via placenta or uterine milk (secretion in wall of uterus)
These features make the elasmobranchs very susceptible to over-fishing, because the populations have low values for rmax (intrinsic rate of natural increase)
Osteicthyes Synapomorphies
Original dermal bone jaws are covered with marginal mouth bones that have rooted teeth
Upper jaw = maxilla, premaxilla
Lower jaw = dentary
Bony operculum – gill cover
Branchiostegal rays – fanlike series of dermal bones that form the floor of the gill chamber and aid in rapid expansion of the mouth for suction feeding and respiration
Lung or swim bladder
Sarcopterygii vs. Actinopterygii
Fins
In ancestral osteichthyans, individual rays are associated with a bone
Result: each individual fin ray can move independently (fine movement in the water)
Actinopterygii lost many of the bones
Better for propulsion; paddles; not fine movements
Sarcopterygii elaborated and gained many bones
Not good for propulsion; good at rotating (like a ball and socket joint)
Tetrapodomorph Sarcopterygii
Further elaborate ball and socket joint
Bones shorter and heavier, better for crawling and terrestrial locomotion
Skull
Actinopterygians: reduced the dermal bones in skull; greater mobility of premaxilla and maxilla (important for feeding and respiration)
Sarcopterygians: retained dermal bones in skull; little to no upper jaw mobility
Polypteriformes
Bichirs and reedfish
Breathe atmospheric air
Ancient lineage
Acipenseriformes
Sturgeons and Paddlefishes
Ancient lineage
Larger upper lobe in caudal fin, generating lift
Large pectoral fins stabilize
Neopterygii
First appeared 345 mya
Most extensive radiation was 80-65 mya
Leedsicthys: probably the only actinopterygian ever mistaken for a stegosaur; around 100 ft in length; supposedly the largest actinopterygian of all time
Lepisosteiformes
Gars
Now restricted to North America, but once Pangean in distribution
Specialized fish eaters; covered with heavy bony scales (ganoid scales; ganoine covering gives the scales an enamel-like cover
Amiiformes
Amiidae
Now restricted to eastern North America
Retain several primitive characters:
Spiral valve intestine
Ganoine covering scales
Solid jaws
Large, lung-like gas bladder (can breath air similar to gar and survive in low oxygen environments)
Gular plate to help crush prey (effective predators on fish and crayfish)
Teleostei Synapomorphies
Stiffened tail (uroneural bones)
Homocercal tail (internally asymmetrical (slightly) and externally symmetrical)
Stiffer, provides greater thrust
Earlier fishes were heterocercal (internally and externally asymmetrical)
Mobile Premaxilla
Maxilla and premaxilla move down and forward
Great for suction feeding
Mobile pharyngeal jaws
E.g. moray eels (can protrude pharyngeal jaws to grasp prey)
Lost basal fin bones (greater rigidity)
Osteoglossomorpha
Live in backwater areas in South America, Africa, Asia, some in North America
Have an elaborated bone on the floor of their mouth
Elopomorpha
Eels, tarpons, bonefish
Synapomorphy:
All elopomorphs share a specialized larval stage call a leptocephalus larva
Only a few cells thick
No GI tract
Absorb nutrients directly from the seawater
Teeth present → serve as a calcium reservoir to later form the skeleton
Otocephala
Herrings, sardines, anchovies, minnows, suckers, catfishes, loaches, characins
Otophysi
Have a special connection between their “ears” and their swim bladder that gives them enhanced hearing
Herrings, shad, sardines = otophysic connection
Minnows, suckers, catfishes, characins = Weberian apparatus
Series of bones connecting swim bladder to hearing organs
Euteleostei
Characteristics:
Large, toothed maxilla
Adipose fin (an additional, often small and fatty fin between dorsal and caudal fins without separate skeletal support)
Tubercles develop on breeding males (nuptial tubercles) - pattern timing and location of tubercles often diagnostic
Ca. 24,000 species
Neoteleostei
Retractor dorsalis muscle: greatly enhanced pharyngeal jaw mobility
Superorder Acanthopterygii (Acanthomorpha)
True spiny-rayed fishes
Monophyletic group, but relationships uncertain
Ca. 14,000 species
Percomorpha
Very diverse group
What made teleosts so successful?
Caudal fin locomotory complex
Homocercal caudal fin designed for thrust
Two advantages
Increased efficiency in horizontal swimming because all thrust is horizontal
Increased versatility because paired fins are now free to evolve other locomotory functions
Results in pectorals moving up the body and pelvics moving forward; both providing greater maneuverability
Feeding mechanisms
Greater jaw mobility (esp. upper jaw) opened up new trophic possibilities; allowed for specialization of mouth parts, which lead exploitation of specialized food resources
Most significant evolutionary change begins with freeing posterior end of maxillary bone from bones in the cheek
Mobile pharyngeal jaws
Retractor dorsalis in Neoteleosts
How did derived vertebrate jaws evolve?
Palatoquadrate cartilage evolves to form part of suspensorium, palatine bone, and part of quadrate. In primitive tetrapods, it forms part of the roof of the mouth (palate); in mammals, part of it forms the incus (one of the three inner ear bones)
Mandibular cartilage in derived fishes forms the articular. In mammals, part of it becomes the malleus (another inner ear bone)
In teleosts
A ball and socket joint developed between head of maxilla and palatine bone
In highly evolved teleosts, anterior end of premaxilla develops an ascending process that extends upward and backward to overlap the snout
Exclusion of maxilla in the gape
In addition, bony connection between premaxilla and snout in lower teleosts is replaced with a more flexible cartilaginous and connective tissue hinge
Categories of fish feeding
Hit and run
Used by mostly fast-swimming open water fishes
Rim of jaws used for biting and grasping; requires a firm jaw construction and large, powerful muscles to shut jaws quickly and firmly
Filter feeders
Open their mouths and hold them open while swimming
Gape and suck
Depends on ability to create sufficient negative pressure to suck-in individual food items
Effectiveness of this method depend on:
Degree to which the mouth cavity can be expanded
Suddenness with which the mouth cavity can be expanded
Sarcopterygii Synapomorphy
Monobasal fin
Compare a shark fin, with three supports attached to the limb girdle (tribasic fin) to a lungfish fin, which is attached to the pectoral girdle and supported along its length by a single row of bones
Gives more rotation: ball-and-socket joint
Actinistia
Coelacanths
Thought extinct since Cretaceous (ca. 65 mya)
Two species were discovered in the 1900s
Dipnoi
Synapomorphy:
Separate pulmonary circulation
Very little change in over 400 my of evolution
Their "lung" is a modified swim bladder, absorbs oxygen and removes wastes.
Respiration using the lungs is critical for survival - lungfish can drown if they can’t breathe air
Obligate air breathers
African and S. Am. lungfish survive dry season by estivation
Burrow into the mud and seal themselves within a mucous-lined burrow
Fossilized lungfish burrows found in Permian age rocks, with the lungfish still inside; older (empty) burrows are known from the Carboniferous and Devonian.
Osteolepiformes
Extinct group of lobe-finned fish
Synapomorphy
Labryinthodont teeth - teeth in cross-section that show complex infoldings
Loss of postaxial elements in fins
Panderichthyidae
Extinct group of lobe-finned fish; most are known only from fossil fragments, but there are a few complete specimens
Have more tetrapod-like characters (synapomorphies):
Flattened skull with snout
Eyes on top of head
No dorsal or anal fin
Reduced tail fin
Late Devonian Sarcopterygii Characters:
Development of free digits
Increased mobility of limb joints
Pelvic girdle attaching to spinal column
Pectoral girdle becoming free from its attachment to the skull
Vertebrae develop stronger interlinkages (zygapophyses)
Tiktaalik roseae
Symplesiomorphies w/ more primitive Sarcopterygians
Similar body scales, fin rays, lower jaw and palate bones
Autapomorphies
Shortened skull roof, dorsally placed eyes, a modified ear region, a mobile neck, imbricate ribs, a pectoral girdle and fore-fin capable of complex movements and substrate support, and a functional wrist joint
Advantages and Obstacles with Transition to Land
Advantages to invasion of land
New food resources
Avoidance of aquatic predators and competitors
Oxygen abundant
Disadvantages (Obstacles)
Water becomes limiting factor in distribution (desiccation, respiration, reproduction)
Gravity - necessitates new morphological designs
Water provides buoyancy and allows for good support even though it is more difficult to move through than air
Water has high heat capacity
Most aquatic animals, especially marine species do not have problems with the drastic temperature changes that occur on land
Locomotion in the transition to land
Modified axial skeleton with zygapophyses
Strengthened shoulder and pelvic girdles
Loss of skull bones, freeing the shoulder from the skull allowing a flexible neck; shoulder girdle supported by serratus musculature
Attachment of ilium to sacral rib
Evolution of paired limbs
Proximal limb elements are homologous with bones of rhipidistian (early sarcopterygian ‘fishes’) fins
Fins are divided into axial, preaxial and postaxial elements
To make a limb from a fin, the axis of the limb has to curve around; need an elongate, laterally directed humerus and femur
Some preaxials were incorporated into the arm and wrist, or fused with other bones
Some postaxials end up as digits
Respiration in the transition to land
Air versus water
Air has higher O2 content - 20 X more per volume; faster diffusion (500,000 X)
Less energy for ventilation; up to 25% of total metabolism for fish, 1-2% for air breathers
Air is not hyper- or hypotonic (no salt gain or loss)
Air does not remove as much heat
Air causes problems with desiccation; structures located deep within body; long passageways moisturize air
Structural adaptations
Absence of internal gills
Reduction and loss of operculum
Better developed lungs than were present in sarcopterygians
Three chambered heart with discrete systemic and pulmonary circulation
Three chambered heart with separation of blood in the ventricle
Loss of scales to allow for cutaneous respiration
Water balance in the transition to land
Excretion
Urea is the principal nitrogenous waste; Latimeria and the living lungfish can synthesize urea in their livers; lungfish use urea for water retention during estivation
Dehydration
Still a problem for most amphibians, so they have to live near water
Sense organs in the transition to land
Middle ear
Derived from spiracle
Hyomandibular (freed from jaw support) modified to columella (stapes); initially the stapes is large and heavy and when it became solely involved in aerial sound transmission is questioned
Evolution of otic notch - supports a tympanum
Reproduction in the transition to land
Usually external fertilization, eggs laid in water (e.g., spermatophore in salamanders)
Stegocephalia
Digit-bearing vertebrates plus Tetrapoda
Synapomorphies
Loss of several cranial bones
Allows the head to remain relatively stable while walking.
Loss of the opercular bones that cover the gill chamber in bony fishes.
The operculum was no longer needed in early choanates because they had lost the internal gills of their early ancestors.
A reduction of the notochord and a rigid spine.
Centra are thick and they constrict the notochord. Special articulatory surfaces (zygapophyses) link the neural arches to each other.
A shorter notochord that does not extend into the braincase.
The notochord of osteolepiforms extended up to the vicinity of the pituitary.
Four muscular limbs with discrete digits (fingers and toes).
A sacral rib connecting the axial skeleton (the spine) to the pelvic girdle (the hip).
This allows the weight of the body of tetrapods to be transmitted to the hind limb
The loss of dermal fin rays (the modified scales that support the fins).
This simply represents the elimination of a structure that was no longer needed and may even have been harmful on land
Acanthostega
First complete tetrapod with free digits (eight on each hand)
Based on recent reports, the limbs and gross morphology of the basal form Acanthostega were unsuitable for overland travel. Instead, Acanthostega probably propelled itself along the bottom with its limbs or held itself motionless in wait for an ambush.
Retained some “fish-like” characters
Internal gills, tail fin, partial connection between skull and pectoral girdle, labyrinthodont teeth, lateral line, ulna shorter than radius
Ichthyostega
Similar to Acanthostega, but…
Stronger limbs; radius and ulna of equal length
No gills in adult
Reduced number of skull bones
Retains skull like Panderichthyidae, tail fin, labyrinthodont teeth, lateral line
First tetrapod capable of life on land
Important characters
Pectoral girdle no longer attached to skull
Pelvic girdle is attached to vertebrae
Seven toes on hindlimb
Reduced number of bones in skull
Theories of why fishes moved onto dry land
Old theory – fish were living in habitats that dried up; survivors were the ones that crawled to new ponds
Natural selection for limbs
But a problem with this scenario is that the Devonian is no longer viewed as having been seasonally dry
Plus, early tetrapod limbs couldn’t support the animal on land
New theory – Acanthostega and other early tetrapods had fully formed limbs, but never left the water
Limbs were useful “after the fact” for moving onto land, but originally had a different function
Analogous living species:
Frogfishes use their modified pectoral fins for support and to move themselves around on the bottom. Frogfish can even “gallop” in short bursts.
Properties of water
Water is much more dense and viscous than air
Sound travels faster in water than air
Water has a much higher heat capacity than air
Water contains less O2 per unit volume than air
Solubility decreases as temperature increases
Warm water has less O2 than cold water
Solubility decreases as salt concentration increases
Salt water has less O2 than freshwater
Gill arch physiology
Each gill arch bears a number of gill filaments (holobranchs), each of which is made up of two halves (hemibranchs)
Fine subdivisions on hemibranch are gill lamellae (major respiratory portion of gills)
Total surface area of the gill lamellae averages about 5 cm2 per gram of body weight
Increased surface area → increased gas exchange
Deoxygenated blood reaches the gills by way of the afferent branchial arteries.
Oxygenated blood from the gills passes into the efferent branchial arteries and into the body
Blood flows through the lamellae in the opposite direction of water flow across the lamellae (counter-current)
Blood with higher O2 content meets water with highest O2 content so that O2 diffuses into the blood along the entire length of the lamellae
The effect is an extremely efficient interchange of O2 and CO2 between water and blood
O2 enters blood, CO2 and protons leave blood
Respiration in Agnathans
Non-feeding mode
Intake is through the nostril (hagfishes)
Ventilation pump = velum
1-16 gill sacs with countercurrent setup
When the hagfish is buried inside prey, water comes in and out through the gill opening behind the last gill pouch
Lampreys expand and contract the branchial area, causing water to flow in/out; practical when head buried in prey
Respiration in elasmobranchs
Water intake is through mouth and spiracle
Ventilation can be either ram (mouth) or pump (mouth and spiracle)
≥ 5 individual gill slits (vs single operculum)
Structure
Gill septum supports gill filaments
Respiration in teleosts
Ventilation ram (mandatory in tunas) and/or pump (buccal, opercular cavity pumps)
Structure
Surface area of gills is correlated to activity levels
Oxygen uptake and release
The tendency of blood to take up and release oxygen is described with an oxygen dissociation curve
The amount of oxygen taken up by hemoglobin (Hb) increases with oxygen tension (partial pressure)
The half-saturation point (P50) is defined as the oxygen tension at which blood is half-saturated.
The dissociation curve is typically sigmoid in shape; this is because of tetrameric molecular structure
Change in pH influences affinity of Hb for oxygen
Blood pH drops where there is higher carbon dioxide (areas of metabolic activity)
Conformation of Hb changes, and reduces its affinity for oxygen (lower half-saturation point). This is called the Bohr Effect; occurs in all animal hemoglobins
In some fishes, there is an additional effect: low pH lowers the oxygen capacity of Hb (reduction of asymptotic saturation). This is called the Root Effect
As a result, Hb unloads oxygen in tissues where there is metabolic activity (where there is lactic acid production)
Diversity among fishes in dissociation curves and Bohr effect
Relative to Hb in an active fish such as mackerel, the oxygen capacity of Hb in a fish living in lower oxygen waters (toadfish) is lower and the Bohr effect is lower; the toadfish is less active and there isn’t as much oxygen unloading at peripheral tissues
Elimination of CO2
In metabolizing tissues
Carbon dioxide hydration produces protons (CO2 + H2O => HCO3 + H+)
This creates an acidic environment, which facilitates unloading of oxygen from hemoglobin
There is Hb uptake of some CO2
Hb also binds protons, thereby providing some buffering
Most carbon dioxide is transported to the gills in the form of dissolved plasma bicarbonate; this is created in the red blood cells but then diffuses out of the cell.
At the gills
The hydration runs backwards as CO2 leaves gills quickly (because the environment is low in it, the molecule simply runs down gradient)
There is a rise in pH, and uptake of oxygen by Hb
The point here is to emphasize the linkages between CO2 and O2 exchange, and the roles of blood cells and plasma.
Gas bladder
Gas filled sac located between the alimentary canal and the kidneys
Filled with CO2, O2, and N2
Functions primarily in hydrostatic balance, respiration; secondarily in sound production and sound reception
When used in respiration, gas bladder is compartmentalized and highly vascularized
Types of gas bladders
Physostomous - retain connection between the esophagus and gas bladder through a pneumatic duct
Physoclistous - lose connection
Structures associated with Physoclistous fishes
Antroventral secretory region
Gas gland - secretes lactic acid, lowers blood pH, and reduces solubility of dissolved gasses
A change of 1 pH unit releases 50% of O2 bound to hemoglobin; raises partial pressure of blood O2 by the Bohr and Root Effects
Rete mirabile - countercurrent exchange system for gas; composed of a looping bundle of arterial and venous capillaries
Posterodorsal resorption region
Oval - thin, highly vascularized area in the swim bladder; circular muscles contract and close the oval, preventing gas outflow; longitudinal muscles contract and expose the oval, permitting gas escape
How did early freshwater vertebrates osmoregulate?
Glomerular kidney
Kidneys in freshwater fishes
Glomerulus: a typical kidney of a freshwater fish has tens of thousands of large glomeruli. Large amounts of water pass through them. Provides a filtrate that can be modified selectively by the kidney tubule
Neck Region: lined with cilia; ciliary action aids movement of materials into tubule. Important in the low-pressure filtration systems of fishes
First Proximal Segment (PCT I): location of reabsorption of many macromolecules (e.g., glucose, proteins); also excretion of organic acids
Second Proximal Segment (PCT II): largest region of tubule; has high metabolic activity (i.e., active transport mechanisms that are responsible for reabsorption of many salts, e.g., Mg2+, SO4-, Ca2+, P, Na+, Cl-, and HCO3)
Intermediate Segment: highly ciliated portion that assists in moving fluids through the tubule. In freshwater fish it is important to move the fluid through the length of the tubule as fast as possible to minimize reabsorption of water
Distal Segment (DCT): participates in active reabsorption of Na+ and some Cl-
Longitudinal Collecting Duct (CT): reabsorbs monovalent ions, again mostly Na+ and some Cl
Chloride cells
Special cells in the gills and oral membranes to absorb ions by active transport mechanisms
Absorbs acid phosphate, bromine, calcium, chloride, lithium, sodium, sulfate ions, etc.
Kidneys in marine fishes
Glomerulus: glomeruli in marine teleosts are small, poorly vascularized, and blood pressure is low. May be lost (aglomerular)
Neck Region: may be lost altogether, especially in aglomerular species
First Proximal Segment (PCT I): location of reabsorption of many macromolecules (e.g., glucose, proteins); also excretion of organic acids
Second Proximal Segment (PCT II): instead of active reabsorption of salts, as in freshwater teleosts, this is the site of active secretion of salts (e.g., Mg2+, SO4-, Ca2+, P, Na+, Cl- , and HCO3). Also responsible for active secretion of nitrogenous wastes (urea, creatine, creatinine)
Intermediate Segment: absent in marine fish; because the need here is to slow the movement of fluid to maximize the amount water passively diffusing back into the blood
Distal Segment (DS): participates in some reabsorption of Na+ and Cl-
Longitudinal Collecting Duct (CT): some reabsorption of Na+ and some Cl-
Osmoregulation in marine elasmobranchs
Marine elasmobranchs osmoregulate in a very different way from teleosts
Evolved a specialized segment of the nephron that reabsorbs urea and returns it to the blood
Influx of urea and TMAO (trimethylamine oxide) raises the osmotic pressure of the blood to a level just above that of seawater so that water flows into the body of the shark (similar to freshwater fish)
Have numerous well-developed glomeruli and excrete large amounts of dilute urine
Evolved a specialized segment of the nephron that reabsorbs urea and returns it to the blood
Have numerous well-developed glomeruli and excrete large amounts of dilute urine
Rectal gland - an organ with chloride cells, for excreting monovalent ions
This type of osmoregulation is a physiologically much less costly system than the marine teleost approach of salt excretion
Thermoregulation terms
Terms associated with the environment
Poikilothermy: A condition in which an organism’s body temperature relies on and varies with the temperature of the environment
Homeothermy: A condition in which an organism maintains a constant internal body temperature
Problem: the body temperatures of most fishes will change with their environment, but the ambient external temperature is often quite stable due to the thermal stability of water
Terms associated with the source of an animal’s body heat
Ectothermy: A condition in which an organism does not generate its own body heat but must use an external source of heat to warm itself
Advantage: low metabolic costs
Disadvantage: cannot live or function well in extreme thermal environments, especially the cold
Endothermy: A condition in which an organism is capable of generating its own body heat and maintaining a constant body temperature
Advantage: biochemical reactions become more efficient, fish can utilize wider thermal ranges
Disadvantage: high metabolic costs
Thermoregulation mechanisms
Regional Endothermy: different temperatures in different parts of an animal’s body
Rete mirabile near large swimming muscles (often found in tunas and some sharks)
Fine counter current network of veins and capillaries that exchange both oxygen and heat
Rete mirable on the liver
Functions in maintaining increased gut temperatures (aids in digestive efficiency; also found in tunas and sharks)
Warm parts of the central nervous system (especially brain and eyes). Allows fish to use deeper, colder, more biological productive habitats without a decrease in brain and visual function.
Modify the circulatory system by having retes near the eyes and brain (better vision at cold temperatures)
Thermogenic tissues - special heat-producing tissues
Billfish have specialized eye muscles without contractile filaments that produce heat without muscle contractions when stimulated by nerves.
Batrachomorphs vs. Reptilomorphs
Batrachomorphs (“similar to a frog”)
Skull roof attached to braincase via the exoccipitals
Loss of skull kinesis
Only four fingers in hand
Reptilomorphs (“similar to a reptile”)
Skull roof attached to braincase via the basioccipital
Increased skull kinesis
Temnospondylii
Temnospondyls are a very large and widespread extinct clade of stegocephalians.
Known from the Lower Carboniferous (ca. 340 mya) to the Lower Cretaceous (ca. 120 mya)
Found on all continents from Greenland to Antarctica.
More than twelve families are known, with about 90 genera.
Display a great diversity of forms, resembling large salamanders, crocodiles or gavials, with brevirostral (short-snouted) to longirostral (longsnouted) skulls.
Their size ranges from about 20 cm to 3 m in length
Lissamphibia Lineages
Anura (Salienta)
Caudata (Urodela)
Gymnophiona (Apoda)
Anura (Salienta) synapomorphies/characters
Frogs and toads have a body that is highly modified from the basic amphibian design:
Body is specialized for jumping (and/or swimming)
Body & vertebral column stiff and inflexible
Hind legs greatly elongated with additional limb segment derived from tarsal bones
Pelvis and vertebral column fused, stiffened, elongated
Loss of tail
Frogs vs. Toads
Not natural groupings; rather distinction is based on morphological and life history characters
Frogs
Relatively longer hindlimbs for longer jumps
Aquatic to semi-aquatic
Well developed webbing on feet
Relatively smooth skin
Toads
More stout body with shorter hindlimbs
Less aquatic; webbing reduced or absent
Skins exhibit a rougher texture
Anuran feeding
Aquatic frogs use suction feeding
Terrestrial frogs typically flip out their tongue
Anuran larvae and metamorphosis
“Tadpoles” have short rounded body, laterally compressed tail, lack legs
Internal gills
Lateral line
Unique hard beak or denticles on mouth parts
Most tadpoles feed on algae, either by filter feeding or scraping algae off rocks
Stream-dwelling tadpoles have sucker-like mouths and muscular tails
Body forms and mouth structures of tadpoles reflect differences in habitat and diet
Some species of frogs have predatory tadpoles
Tadpole metamorphosis involves:
Degeneration and resorption of tail
Growth of limbs (bone, muscle, skin, etc.)
Reorganization of mouth, head
Calcification of skeleton
Formation of dermal glands
Loss of gills, operculum
Development of lungs
Changes to eye musculature and retinal pigments
Growth of portions of brain (cerebellum, etc.)
Alterations to kidneys, pancreas, intestines
During metamorphosis, tadpoles are more vulnerable to predators
Metamorphosis is very rapid (often just a few days)
Caudata (Urodela) generalities
Salamanders have retained the most generalized body plan
Body is somewhat to very elongated, with 4 limbs splayed out to sides
Walking gait probably very similar to that of the earliest tetrapods
Salamanders often live in burrows, crevices, or under debris
Often results in reduction in size of limbs (which get in way) and elongation of body
Some sirenians have lost their hind limbs
Caudata (Urodela) Reproduction
Salamander larva are aquatic
Larval stage has similar elongated body, but…
Broad tail fin for swimming
External gills (in many)
Lateral line, lack of eyelids
Some salamanders are paedomorphic = retain larval characteristics (including fully aquatic lifestyle) throughout their life (e.g. mudpuppies, Necturus)
Many Plethodontid salamanders have direct development: young hatch from eggs (laid in moist places on land) in miniature adult form
Caudata (Urodela) Feeding
Aquatic larvae and adults typically use buccal expansion for suction feeding
Terrestrial forms typically protrude their sticky tongue to pick up prey
Plethodontid (lungless) salamanders can project their tongue to capture prey more than half their body length away
Involves hyobranchial apparatus which therefore can’t be used for buccal pumping (breathing)
Gymnophiona (Apoda) Generalities
Caecilians; tropical and subtropical; limb-less worm-like shape for burrowing
Caecilians are amphibians that resemble large earthworms in shape, but are a different color (yellow or even purple)
Their mouths, however, are large, and they are predacious upon various small fish or invertebrates. They tend to burrow in both the wild and in captivity.
Most caecilians are fossorial and remain underground throughout most of their lives while other forms are aquatic
Gymnophiona (Apoda) Synapomorphies
Limbless and completely lack pectoral and pelvic girdles.
Eyes completely or partially covered by bone or flesh.
A tentacle is present on either side of a caecilian’s head between the eye and the nares. These tentacles are apparently significant in receiving chemosensory cues.
Eversible male copulatory organ (phallodeum) that is partially formed by the cloacal wall and by which internal fertilization is accomplished.
An absence of ear drums or inner ear cavities.
Annuli (rings) are found throughout the body of most caecilians.
Caecilians are the only amphibians with scales.
These dermal scales are located within the annuli and require very close inspection and the use of a good microscope to notice.
Gymnophiona Reproduction
Oviparous species are either direct developing (no aquatic larval stage) or have limbless larvae with external gills and tail fin
Viviparity evolved several times within caecilians
After the developing embryos use up the last of their yolk they still require nourishment while inside their mother’s oviduct. The embryos begin feeding upon a substance labeled uterine milk which is secreted by the oviducts.
The embryos are equipped with tiny uterine teeth that may be used to scrape away and consume the oviduct secretion. The teeth are then lost before or shortly after birth.
Lissamphibia Synapomorphies
Moist, permeable skin and substantial cutaneous gas exchange
Pedicellate, bicuspid teeth
Operculum-columella complex
Papilla amphibiorum
Green rods
Except in Caecilians, which were secondarily lost
Levator bulbi muscle
Causes the eyes to bulge outward
Lissamphibia hearing
Stapes (columella) and operculum are sound-conducting structures
Stapes is only in tetrapods
Operculum is connected via opercular muscle to the suprascapula
Caecilians lack opercular muscles (no shoulder girdle)
Papilla Amphibiorum sound conduction:
From ground through leg through suprascapular through opercularis muscle through operculum to papilla amphibiorum
Papilla basilaris sound conduction:
Tympanum to stapes to papilla basilaris
There are two sensory patches in the inner ear: the papilla basilaris is found in other tetrapods and the papilla amphibiorum is unique to living amphibians
These two sensory patches allow them to hear sounds both >1000 Hz (basilaris) and <1000 Hz (amphibiorum)
Sound can conduct through the legs to the papilla amphibiorum, allowing them to hear vibrations from the ground
Lissamphibia reproduction
External fertilization – females lays eggs that males then fertilize
A few salamanders have external fertilization
Most frogs use amplexus (male grasps female until she lays her eggs)
Internal fertilization in salamanders accomplished via spermatophores (sperm packet) that males produce
Spermatophores may be inserted by males or placed on ground, after which female picks them up
Viviparity
Common in caecilians; possibly up to 75% of species; appears to have evolved several times
Embryos obtain nourishment by scraping nutrients off lining of oviduct
Caecilian embryos are large (up to 3 - 2 mother’s body length) at birth
Rare in salamanders and frogs; involves production of few, large young
Requires internal fertilization, but little is known about how it works in these species
Oviparity
Most common breeding method for both salamanders and frogs
Eggs are laid in water for most species, but numerous exceptions occur
Direct development may avoid competition or predators in the aquatic environment
Eggs are vulnerable and survivorship low, so fecundity must be high to compensate
Lissamphibia Parental Care
Egg guarding
Various salamanders, caecilians and frogs guard eggs (esp. terrestrial eggs)
Some frogs build nests to protect their eggs. Nests may be made of foam or scooped out pools that fill with seepage or rainwater
Egg transport
Surinam toad (Pipa pipa) eggs are pushed into thickened skin on female’s back with complex aquatic mating dance
Egg transport & brooding
Hemiphractus tree frogs – these frogs care for the developing eggs by carrying them on their back in a shallow basin or in pouches modified from the skin. The developing embryos hatch as froglets in some species or as tadpoles in others
Rhinoderma – the males carry the tadpoles in their vocal sacs, and development is completed as froglets.
Tadpole transport & care
Dendrobatid (dart-poison) frogs lay eggs in bromeliads or on the ground. In some species, females lay infertile eggs as food for the tadpoles
When eggs hatch, either male or female (depending on species) picks up tadpoles which stick onto back and carries them around to new locations
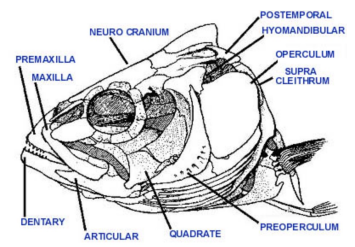
Hyoid Coupling
Emerged in primitive Actinopterygians
Lower jaw depression is initiated by contractions of epaxial and hypaxial muscles
Contraction of epaxial muscles causes head to rotate upward relative to bodies axis
Contraction of hypaxial muscles (especially those on the cleithrum) causes a backward and downward rotation of the pectoral girdle
Anteriorly, the cleithra are attached to the hyoid apparatus by the sternohyoideus muscle
This muscle also contracts so that the backward and downwards movement is transmitted to the hyoid
Because the hyoid lies between and is attached to the lower jaw, the downwards and backwards pull on the hyoid apparatus is transmitted to the lower jaw
Opercular Coupling
Emerged in bowfins and teleosts
Opening of mouth initiated by contraction of the levator operculi muscle
Causes the opercle to swing up and backward
Because the sub- and inter- opercle are attached, this up-and-backwards movement is transmitted ventrally throughout all elements of the opercular apparatus
The interopercle pulls back on lower jaw by means of a strong ligament
Advantage
Necessary pre-adaptation for those fish that feed by nipping at rocks and coral, pick-up small prey, bite-off and crush coral (e.g., surgeonfishes, parrotfishes, wrasses, triggerfishes, etc.)
Derived teleosts possess both gape-and-suck feeding and the opercular coupling mechanism
What muscle closes the mouth and where does it attach?
Adductor Mandibulae
Originates on the suspensorium and inserts along the length of the primordial ligament