Auditory and Vestibular System
1/29
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No study sessions yet.
30 Terms
Hair cells- motion detecting mechanoreceptors
A hair cell is a specialised mechanoreceptor that detects mechanical forces. Different groups of hair cells detect:
• Movement of surrounding water
• Self-movement in water
• Sound waves of different frequencies
• Lateral (side-to-side) movement of the head
• Rotational movement of the head
• The direction of gravity
Hair cells convert physical displacement of stereocilia into electrical signals (mechanotransduction). These signals then activate afferent nerves that carry information to the brain.
What is the structure of a generic hair cell?
• Cell body sitting on supporting cells
• Stereocilia bundle arranged in rows of increasing height
• Tip links connecting stereocilia
• Synapse with an afferent dendrite at the base
The apical surface faces the endolymph and contains mechanosensitive ion channels.
Hair cell and Stereocilia mechanism
Endolymph is the fluid that surrounds the stereocilia of hair cells in the inner ear
It is found in the cochlear duct, semicircular canals, utricle, and saccule
It has an unusual ionic composition:
Very high K⁺
Very low Na⁺
It is electrically positive relative to the inside of the hair cell
This environment is what makes hair-cell mechanotransduction possible.
Where the ion channels are
Mechanically gated ion channels are located at the tips of stereocilia
These channels are physically attached to tip links
Tip links connect shorter stereocilia to taller ones
So movement of the bundle directly pulls on the channels.
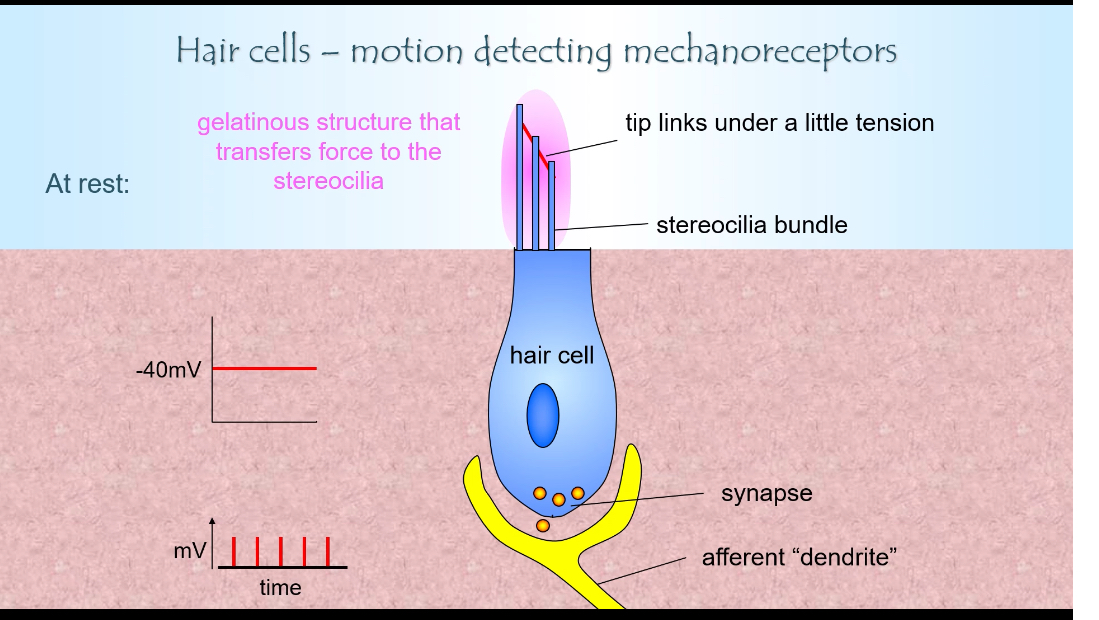
What happens at rest
Tip links are under slight tension
Some ion channels are partially open
K⁺ enters from the endolymph
Hair cell sits at about −40 mV
There is continuous low-level glutamate release
The afferent neuron fires at a baseline rate
This baseline allows signals to increase or decrease.
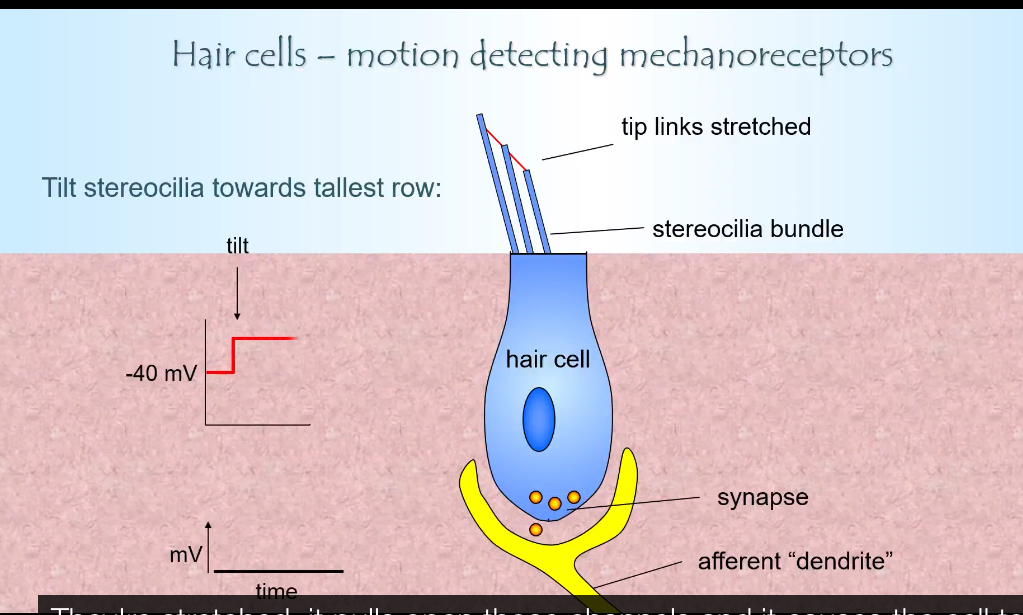
Movement toward the tallest stereocilia (depolarisation)
Stereocilia bend toward the tallest row
Tip links are stretched
More mechanosensitive channels open
K⁺ flows from the endolymph into the stereocilia
K⁺ then travels down the stereocilia bundle into the hair cell body
Hair cell depolarises
Voltage-gated Ca²⁺ channels open at the base
More glutamate is released
Afferent firing rate increases
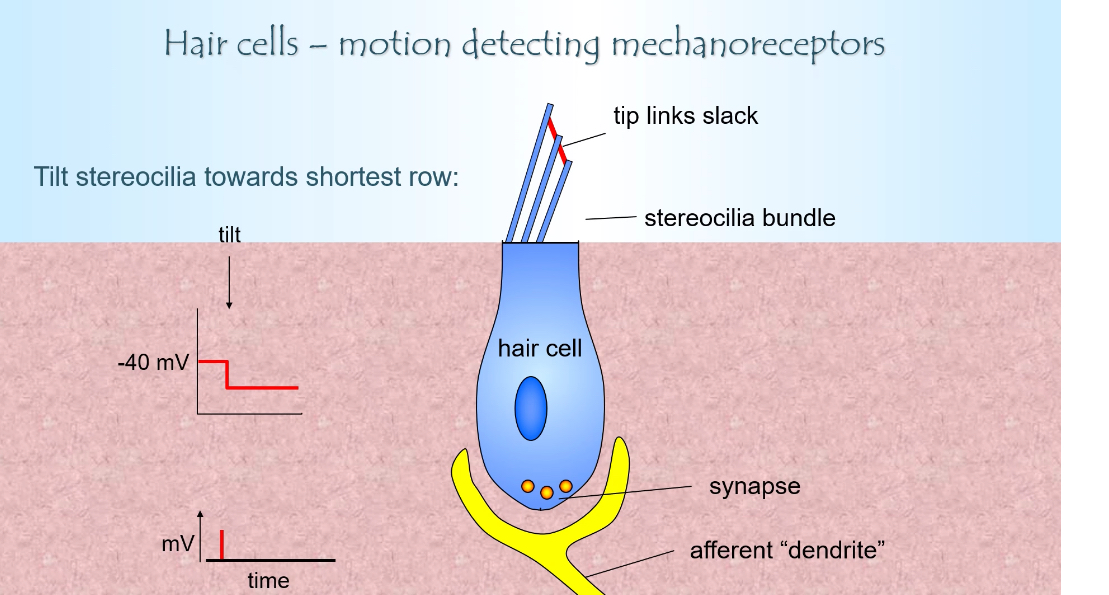
Movement toward the shortest stereocilia (hyperpolarisation)
Stereocilia bend away from the tallest row
Tip links become slack
Ion channels close
Less K⁺ enters from endolymph
Hair cell hyperpolarises
Less Ca²⁺ entry
Less glutamate released
Afferent firing rate decreases
Endolymph surrounds only the stereocilia (apical end) of the hair cell
The cell body and synaptic base are bathed in perilymph (normal extracellular fluid, low K⁺)
So functionally:
Apical side (stereocilia) → endolymph → K⁺ influx
Basal side → perilymph → Ca²⁺ entry + neurotransmitter release
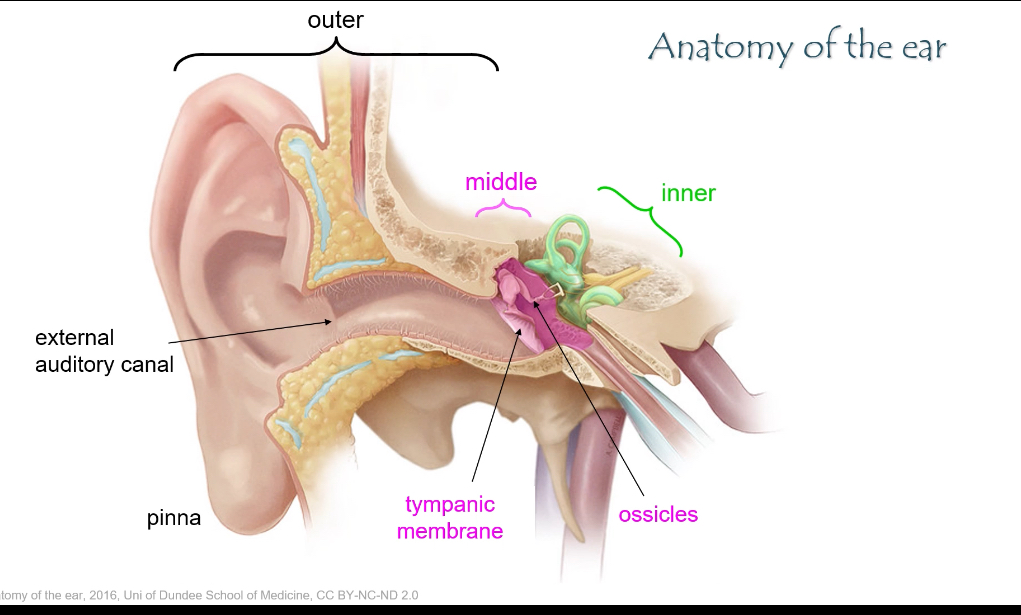
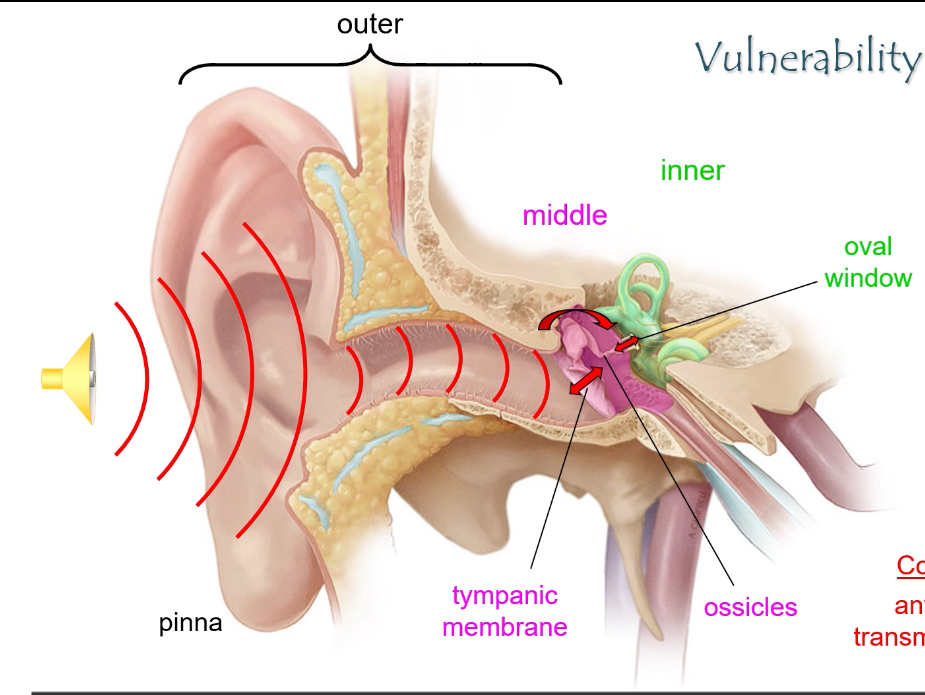
What are the three main parts of the ear and their roles?
• Outer ear: pinna + external auditory canal → captures sound.
• Middle ear: tympanic membrane + ossicles → amplifies sound.
• Inner ear: cochlea + vestibular system → sound detection and balance.
Inner ear- auditory
Cochlea
Spiral-shaped part of the inner ear
Does hearing
Turns sound vibrations into nerve signals
Oval window
Where sound enters the cochlea
Vibrates when the stapes pushes on it
Starts fluid movement inside the cochlea
Round window
Pressure release
Moves so the inner-ear fluid can vibrate properly
Perilymph
Fluid in the outer chambers of the cochlea
Low K⁺, high Na⁺
Carries sound vibrations through the cochlea
Cochlear duct (scala media)
Middle chamber of the cochlea
Contains endolymph
Holds the organ of Corti
Endolymph
Fluid around the stereocilia
High K⁺
Allows K⁺ to enter hair cells when they move
Organ of Corti
Where sound is detected
Sits on the basilar membrane
Contains hair cells
Hair cells
Sensory cells for hearing
Movement → electrical signal
Release glutamate to the auditory
Stereo-cilia
Hair-like projections on hair cells
Bend with sound-induced movement
Control K⁺ channel opening
Basilar membrane
Vibrates with sound
Different areas respond to different frequencies
Auditory (cochlear) nerve
Carries signals to the brain
Brain interprets them as sound
Flowchart
Sound waves enter the ear canal
Sound hits the tympanic membrane (eardrum) → it vibrates
Vibrations pass through the ossicles(3 small bones) (malleus → incus → stapes)
Stapes pushes on the oval window
This creates fluid waves in the cochlea (perilymph)
Fluid movement causes the basilar membrane to vibrate
This bends stereocilia on hair cells in the organ of Corti(in cochlear duct)
Bending opens mechanically gated K⁺ channels
K⁺ enters from endolymph → hair cell depolarises
Depolarisation opens Ca²⁺ channels at the base
Glutamate is released onto the auditory nerve fibre
Auditory (cochlear) nerve carries the signal to the brain

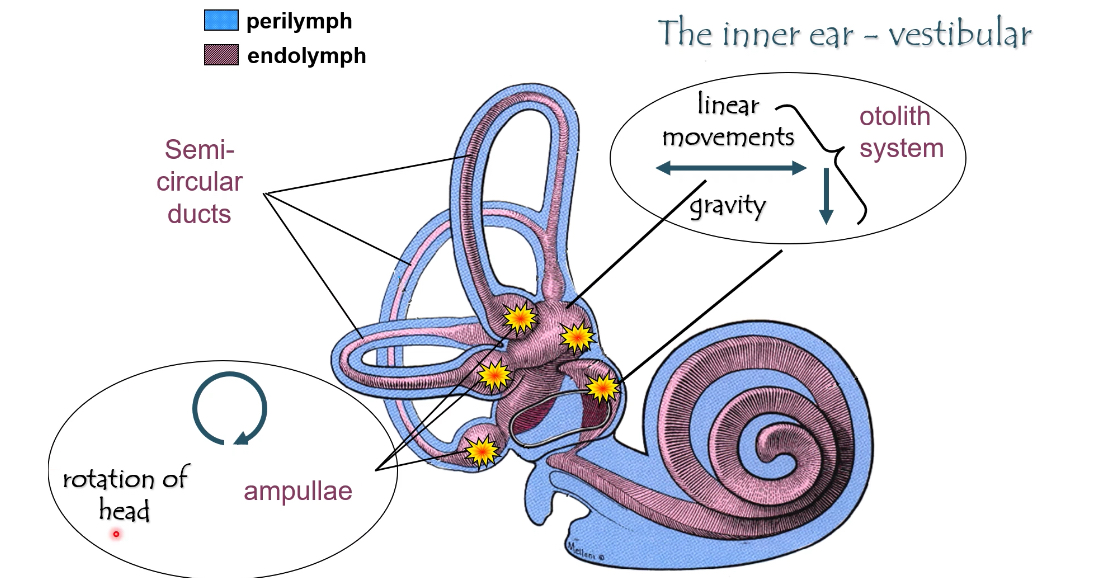
Inner ear: Vestibucular
The vestibular system detects head movement and head position
It tells the brain:
Are you rotating?
Are you moving in a straight line?
Which way is gravity (tilt)?
It uses hair cells, just like hearing, but for movement, not sound
Fluids (same idea as hearing)
Perilymph (blue)
Surrounds the membranous structures
Normal extracellular fluid
Endolymph (purple)
Inside the vestibular ducts
High K⁺
Bathes the stereocilia
Hair-cell depolarisation still depends on K⁺ entering from endolymph.
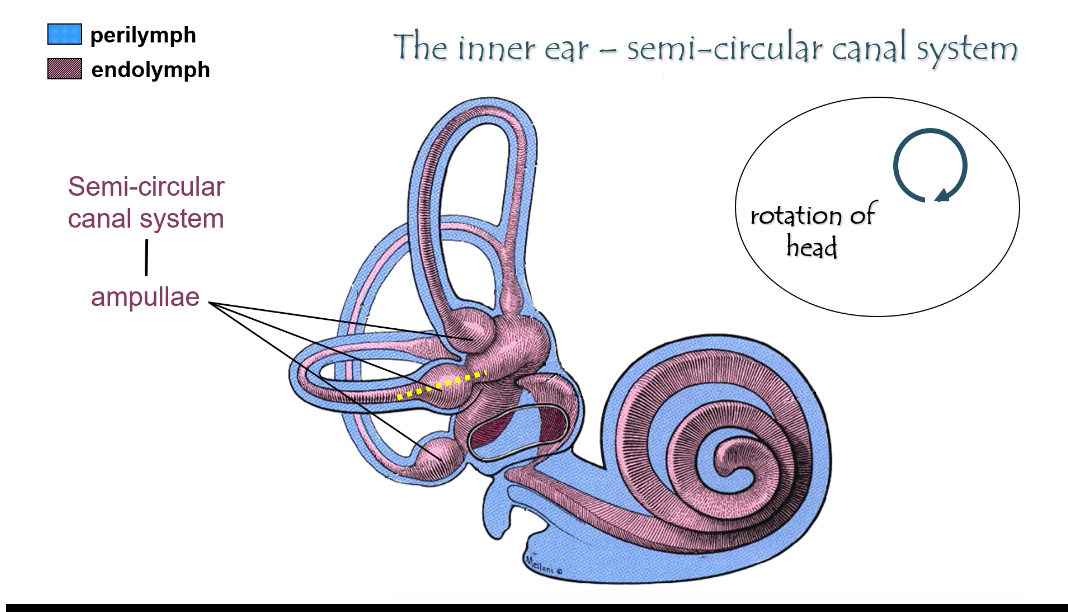
Two vestibular subsystems
1) Semicircular canals → rotation
What they detect
Rotational (angular) movement of the head
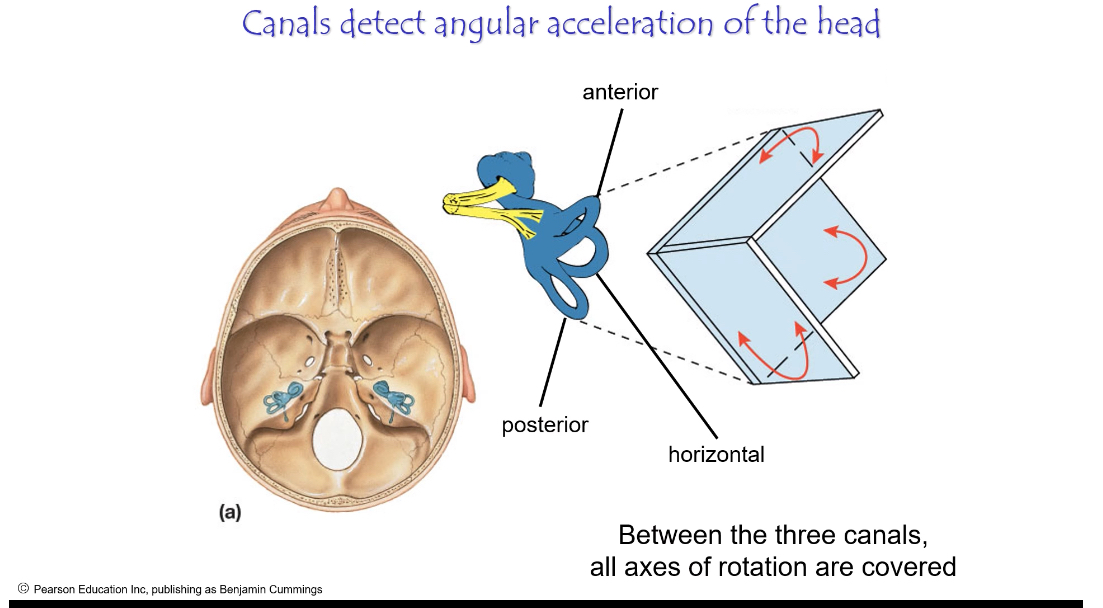
Turning your head left/right, nodding, tilting sideways
Structure
Three canals (horizontal, anterior, posterior)
Each canal ends in an ampulla
Ampulla
Enlarged region at the base of each canal
Contains the crista ampullaris (sensory organ)
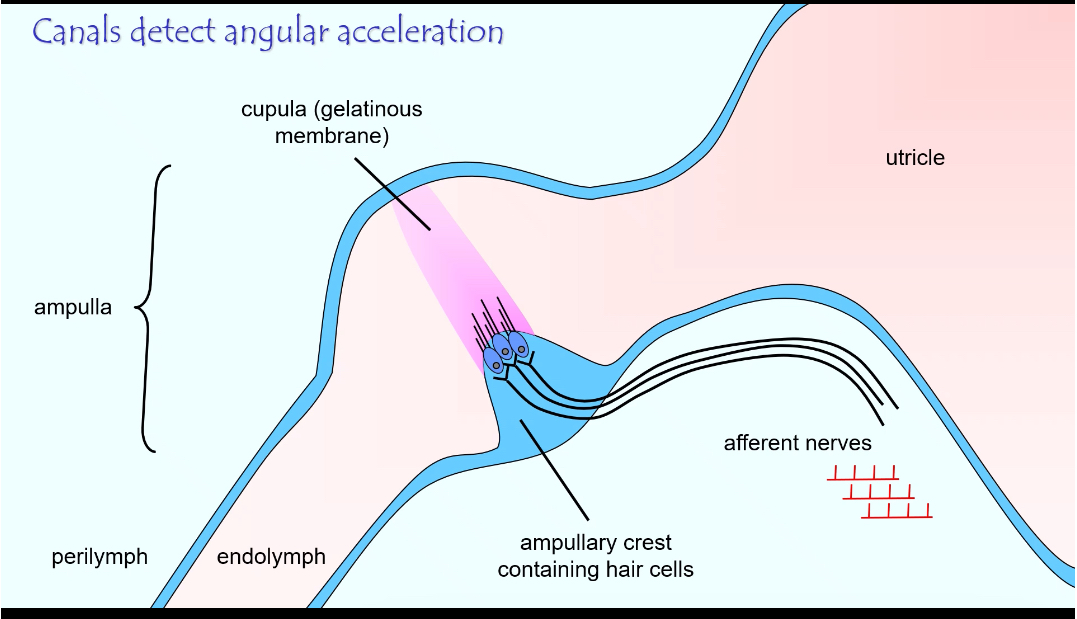
How it works
Head rotates
Endolymph lags behind due to inertia
This bends the cupula in the ampulla
Stereocilia bend
Hair cells change firing rate
Direction of rotation is encoded by increase vs decrease from baseline
Key idea
Semicircular canals = rotation
2) Otolith system (utricle + saccule) → linear movement & gravity
What they detect
Linear acceleration (forward/back, up/down)
Head tilt relative to gravity
Structures
Utricle
Saccule
Together called the otolith organs.
Sensory region
Macula
Contains hair cells
Otoliths
Tiny calcium carbonate crystals
Sit on a gelatinous layer above stereocilia
How it works
Head tilts or moves linearly
Otoliths shift due to gravity or acceleration
This bends stereocilia
Hair cells depolarise or hyperpolarise depending on direction
Key idea
Utricle & saccule = straight-line movement + gravity
What the yellow stars mean on the slides
They mark where the sensory receptors (hair cells) are
In the:
Ampullae of semicircular canals
Maculae of utricle and saccule
Compare hearing vs vestibular hair cells
Same basic mechanism:
Stereocilia bend
Tip links open K⁺ channels
K⁺ enters from endolymph
Glutamate released
Different stimulus:
Hearing → sound vibration
Vestibular → head movement
Hair cell: Receptor Systems
1. Cochlear system: sound frequency, loudness, timing.
2. Otolith system: linear movement and gravity.
3. Semi-circular canal system: angular acceleration.(Rotation)
How are sound waves transmitted from the outer ear to the inner ear?
1. Sound enters the pinna and travels through the external auditory canal.
2. It vibrates the tympanic membrane.
3. Vibrations are amplified by the ossicles (malleus, incus, stapes).
4. The stapes footplate pushes on the oval window, transmitting vibrations into the cochlea.
Ossicles provide impedance matching: without them, sound waves would reflect off the fluid-filled cochlea.
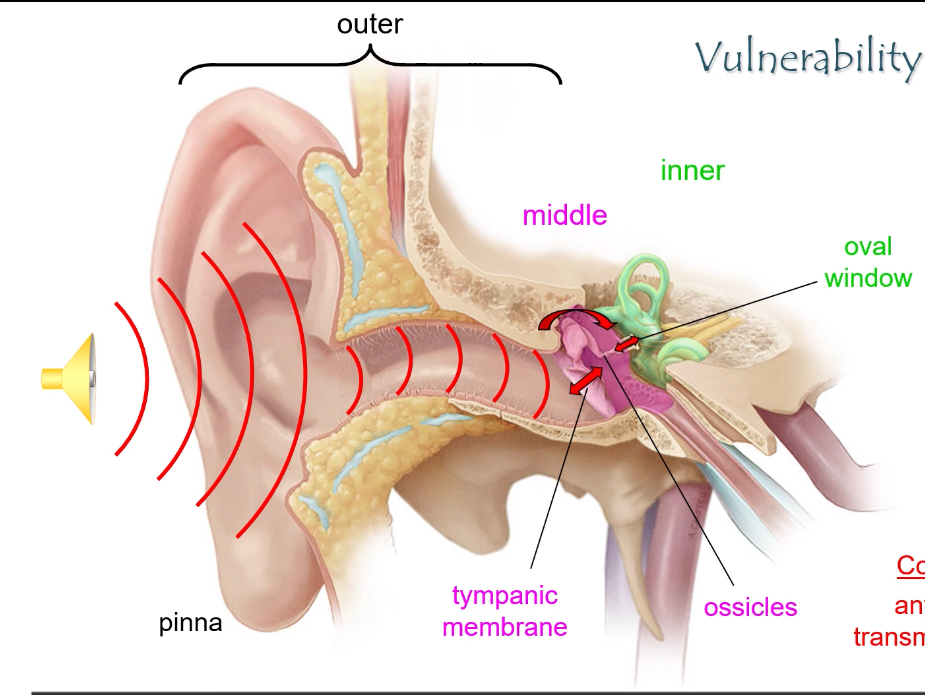
What causes conductive hearing loss?
Any blockage that prevents sound from reaching the oval window, e.g.:
• Ear canal obstruction
• Perforated tympanic membrane
• Otitis media
• Fixation or damage of ossicles
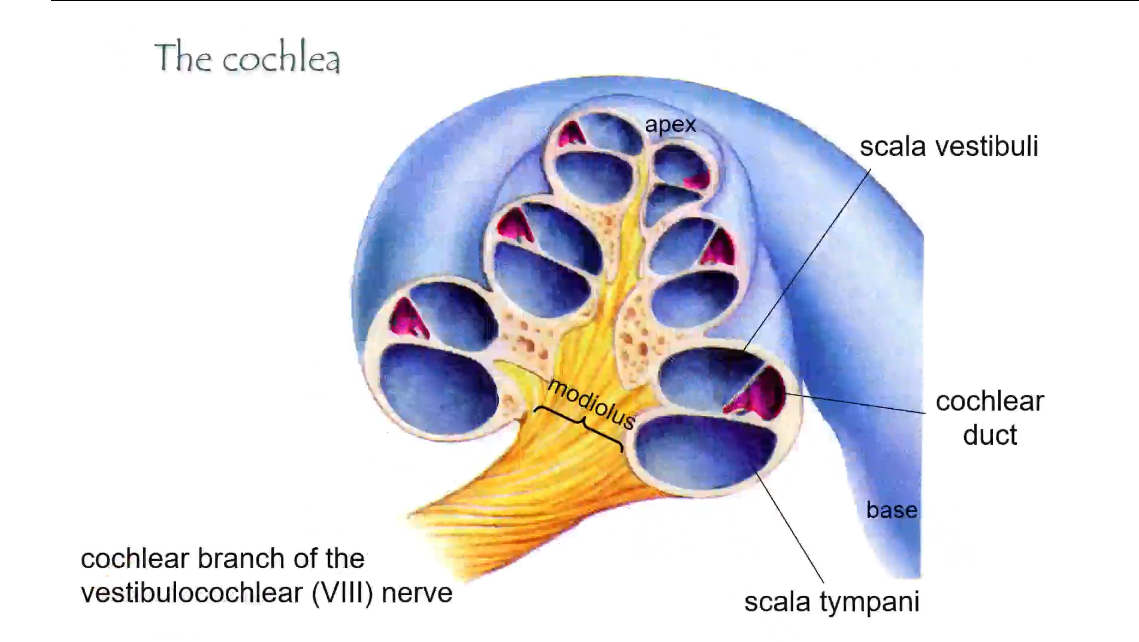
Cochlea: 3 Components and fluid types
• Scala vestibuli – perilymph
• Scala media (cochlear duct) – endolymph
• Scala tympani – perilymph
These compartments are separated by the vestibular membrane (top) and basilar membrane (bottom).
Sound enters at the base and travels toward the apex
Fluids (colour-coded in the diagrams)
• Perilymph (blue)
• Fills the outer chambers
• Normal extracellular fluid (high Na⁺, low K⁺)
• Carries pressure waves
• Endolymph (purple)
• Fills the cochlear duct
• High K⁺
• Surrounds hair-cell stereocilia
The three chambers of the cochlea (cross-section)
Each “slice” of the cochlea has three tubes:
Scala vestibuli
• Top chamber
• Filled with perilymph
• Receives vibrations from the oval window
Cochlear duct (scala media)
• Middle chamber
• Filled with endolymph
• Contains the organ of Corti
• Where hearing actually happens
Scala tympani
• Bottom chamber
• Filled with perilymph
• Ends at the round window (pressure release)
Organ of Corti (inside the cochlear duct)
• The sensory organ of hearing
• Contains hair cells
• Hair-cell stereocilia project into endolymph
• Base of hair cells releases glutamate onto the auditory nerve
Oval window and round window
• Oval window
• Where the stapes pushes
• Starts fluid movement in perilymph
• Round window
• Moves to allow fluid displacement
• Prevents pressure build-up
Base vs apex (important)
Base
• Near oval window
• Detects high-frequency sounds
Apex
• Tip of the spiral
• Detects low-frequency sounds
Modiolus and nerve
• Modiolus
• Central core of the cochlea
• Cochlear branch of CN VIII
• Carries signals from hair cells to the brain
Cochlear duct: Structures
Basilar membrane
A flexible membrane forming the floor of the cochlear duct
Vibrates in response to sound-induced fluid movement
Different parts vibrate best at different frequencies:
Base → stiff → high frequency
Apex → floppy → low frequency
Movement of this membrane is what drives hair-cell stimulation
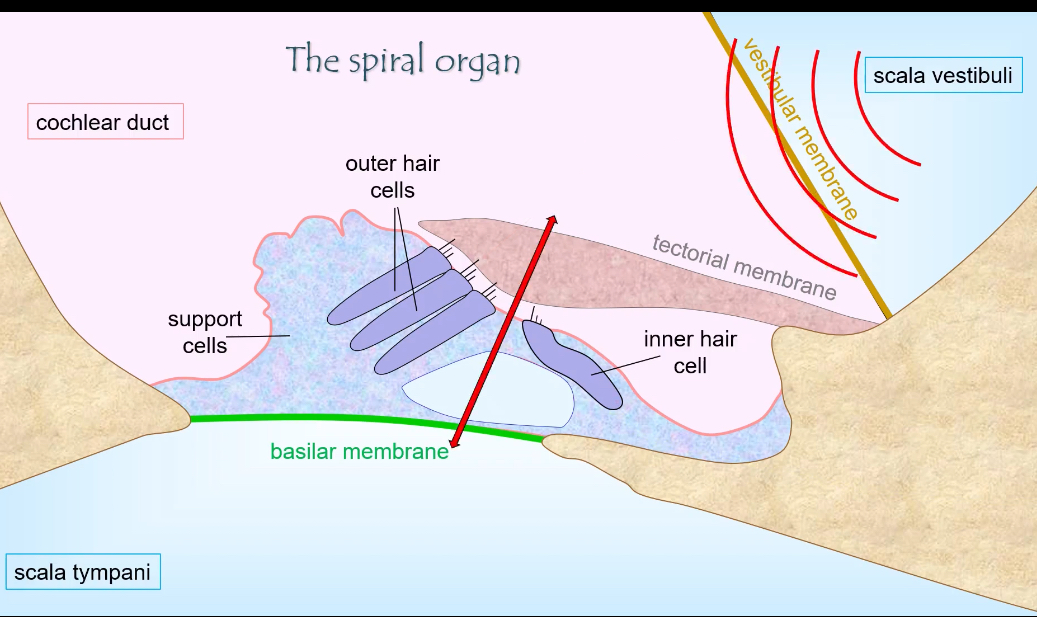
Spiral organ (organ of Corti)
The sensory organ of hearing
Sits on top of the basilar membrane
Located inside the cochlear duct
Contains:
Inner hair cells
Outer hair cells
Supporting cells
Its job is to convert basilar membrane movement into neural signals
Tectorial membrane
A gelatinous membrane that lies above the hair cells
Outer hair-cell stereocilia are embedded in or contact it
When the basilar membrane moves:
The tectorial membrane moves slightly differently
This creates a shearing force
That force bends stereocilia
Key role: turns vibration into stereocilia bending
Hair cells
Mechanoreceptors that detect movement
Two types:
Inner hair cells
Main sensory receptors
Send most signals to the brain
Outer hair cells
Act as amplifiers
Increase sensitivity and frequency tuning
Stereocilia bending:
Opens mechanically gated K⁺ channels
K⁺ enters from endolymph
Hair cell depolarises
Ca²⁺ enters at the base
Glutamate is released onto auditory nerve fibres
Hair cells themselves do not fire action potentials
How they work together (simple flow)
Sound → fluid movement
Fluid movement → basilar membrane vibration
Basilar membrane movement → shearing against tectorial membrane
Shearing → stereocilia bend
Bending → hair-cell depolarisation
Depolarisation → nerve signal to brain
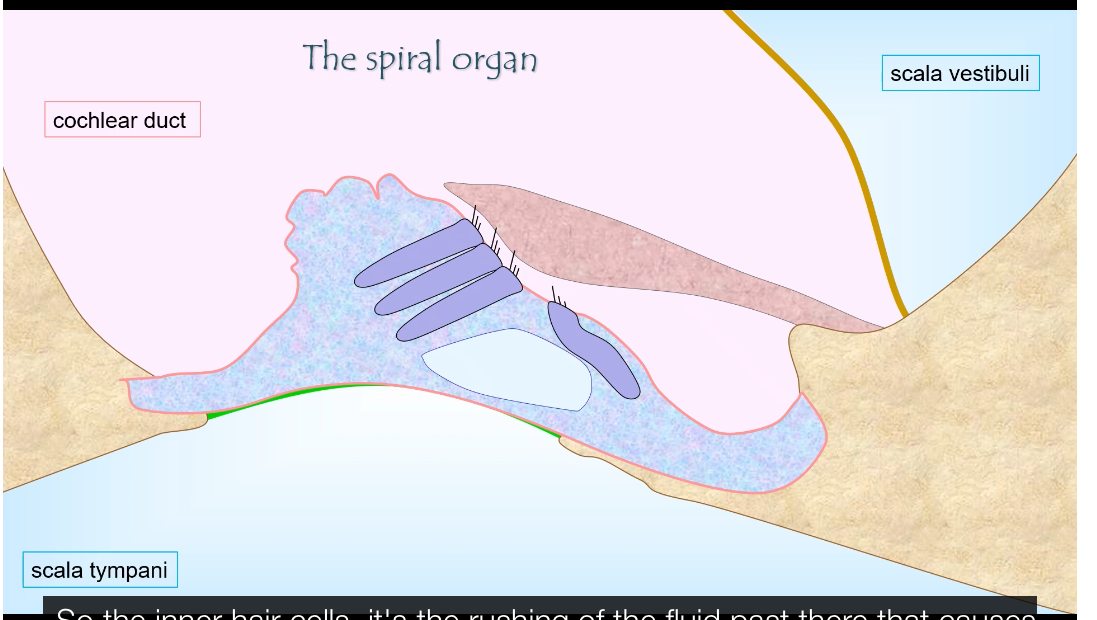
How does the auditory system encode sound frequency?
Through a place code:
• Base of cochlea: narrow, stiff → responds to high frequencies (20 kHz).
• Apex: wide, flexible → responds to low frequencies (20 Hz).
The brain determines pitch by which neurons fire.
How does the auditory system encode loudness and timing?
The louder the sound:
• Larger basilar membrane vibration
• Larger receptor potentials
• More transmitter released
• Higher action potential firing rate
Timing:
Preserved by fast axons and powerful synapses
Primary auditory pathway
This shows how sound information travels from the ear to the brain
It is the primary auditory pathway used for discriminative hearing (pitch, loudness, sound identity)
Step 1: Cochlear nerve (CN VIII)
Hair cells in the organ of Corti release glutamate
Signals travel along the cochlear nerve
This nerve is part of the vestibulocochlear nerve (cranial nerve VIII)
Step 2: Cochlear nuclei (brainstem)
First synapse in the auditory pathway
Located in the medulla
From this point, auditory information is sent to both sides of the brain
This bilateral projection explains why damage on one side rarely causes total deafness
Step 3: Superior olivary nuclei (brainstem)
First site of binaural comparison
Important for sound localisation
Compares:
Timing differences (low frequencies)
Loudness differences (high frequencies)
Step 4: Inferior colliculus (midbrain)
Major integration centre for sound
Combines information about:
Frequency
Intensity
Timing
Involved in sound localisation and auditory reflexes
Step 5: Medial geniculate nucleus (thalamus)
The auditory relay nucleus of the thalamus
Filters and organises auditory input
Sends processed signals to the auditory cortex
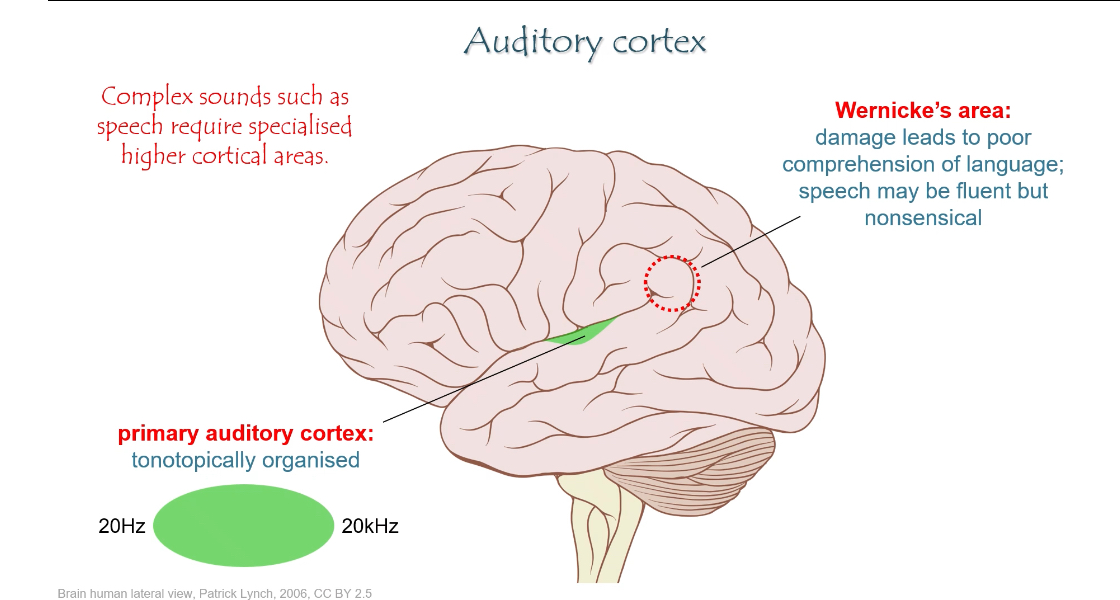
Step 6: Primary auditory cortex (A1)
Located in the temporal lobe
Tonotopically organised:
Low frequencies map to one region
High frequencies map to another
This is where sound becomes consciously perceived
Higher auditory cortical areas
Surround the primary auditory cortex
Process complex sounds, especially speech
Include Wernicke’s area:
Responsible for language comprehension
Damage → fluent but meaningless speech (poor understanding)
Key exam points to remember
Pathway is mostly bilateral after the cochlear nuclei
Thalamic relay = medial geniculate nucleus
Primary auditory cortex is tonotopic
Wernicke’s area = understanding speech, not producing it
Auditory System: Origin of sound
The brain compares what each ear hears
This first happens in the superior olivary complex (brainstem)
Two different nuclei do two different comparisons:
Timing differences
Loudness differences
Medial Superior Olive (MSO) — timing
Compares when sound arrives at the left vs right ear
Uses interaural time difference (ITD)
Best for low-frequency sounds
Why only low frequency:
Low-frequency waves are long
The phase of the wave is clear and comparable between ears
The brain can tell which ear was stimulated first
What it tells you:
Sound arriving earlier at the left ear → sound is on the left
Sound arriving earlier at the right ear → sound is on the right
Key phrase to remember:
MSO = timing = low frequency
Lateral Superior Olive (LSO) — loudness
Compares how loud the sound is in each ear
Uses interaural level difference (ILD)
Best for high-frequency sounds
Why only high frequency:
High-frequency sounds are short wavelength
The head blocks them → head shadow effect
One ear gets a quieter signal than the other
What it tells you:
Louder in left ear → sound is on the left
Louder in right ear → sound is on the right
Key phrase to remember:
LSO = loudness = high frequency
What is the primary auditory pathway?
1. Cochlear nerve
2. Cochlear nuclei
3. Superior olivary nuclei
4. Inferior colliculus
5. Medial geniculate nucleus (thalamus)
6. Primary auditory cortex (A1)
A1 is arranged tonotopically, preserving the frequency map.
Outer hair cells
OHCs actively amplify vibrations:
• They contract/elongate in response to sound (electromotility).
• This boosts basilar membrane motion.
• It increases sensitivity and frequency resolution.
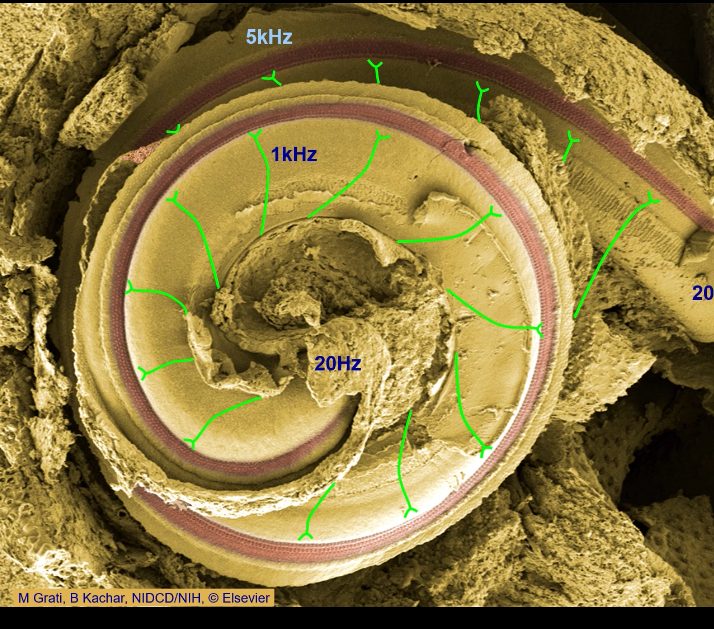
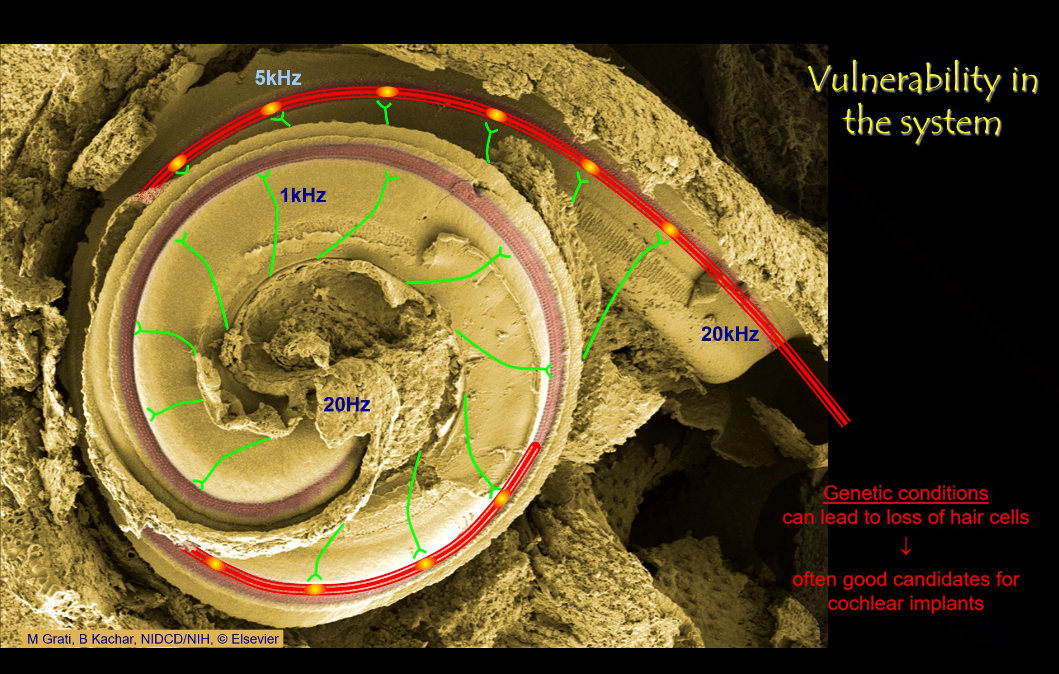
Why is the auditory system vulnerable to damage?
• Only ~3,500 inner hair cells per ear.
• Loud sounds cause excessive vibration → mechanical destruction.
• Excess glutamate at synapses can destroy afferents.
• Genetic conditions can eliminate hair cells.
How do cochlear implants help?
They bypass lost hair cells by directly stimulating the auditory nerve with electrical signals, preserving tonotopic coding along the cochlea.
What are the two major vestibular receptor systems and what do they detect?
1. Otolith system (utricle & saccule): detects linear acceleration and gravity.
2. Semi-circular canals: detect angular acceleration (rotational movement).
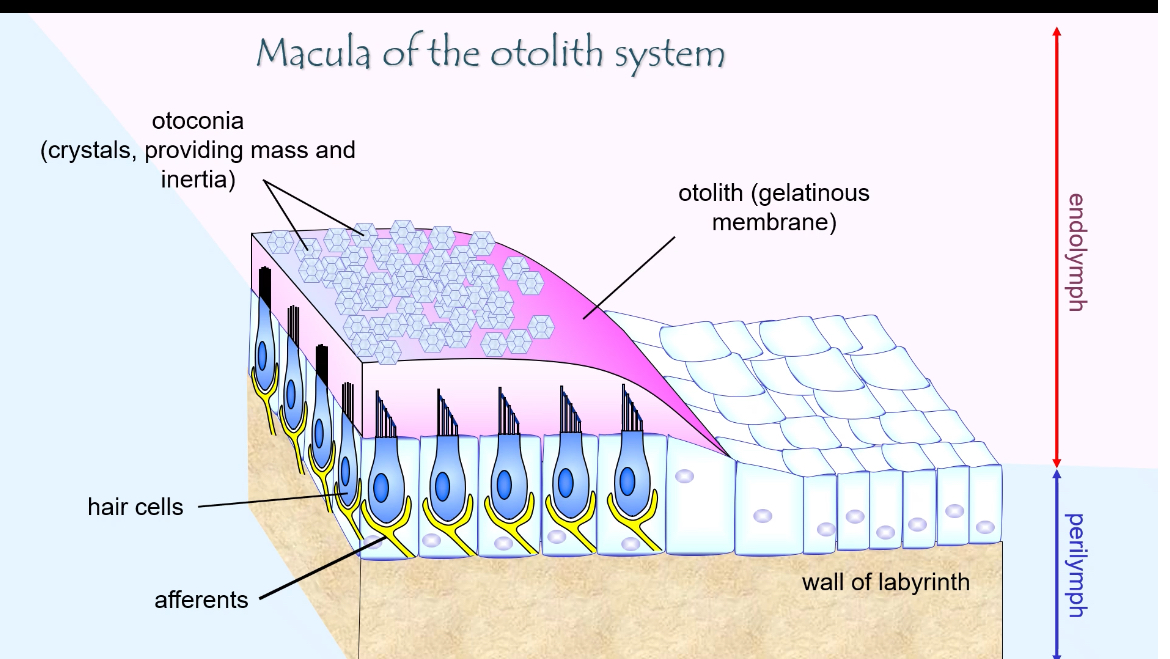
Otolith Macula: Structure
What the macula is
The macula is the sensory epithelium of the otolith organs (utricle and saccule)
Its job is to detect linear acceleration and head tilt (gravity)
Hair cells
Sensory mechanoreceptors
Each hair cell has a bundle of stereocilia (and one kinocilium)
The direction the bundle bends determines whether the cell depolarises or hyperpolarises
The apical ends of hair cells face endolymph
The basal ends synapse with afferent vestibular nerve fibres
Otolithic (gelatinous) membrane
A gelatinous layer sitting on top of the hair-cell stereocilia
Stereocilia are embedded in this membrane
It moves relative to the hair cells during head movement or tilt
Otoconia (otoliths)
Tiny calcium carbonate crystals
Sit on top of the otolithic membrane
Add mass and inertia
This extra weight makes the membrane shift when:
You accelerate
You tilt your head relative to gravity
This shift is what bends the stereocilia.
Fluids around the macula
Endolymph
Surrounds the stereocilia
High K⁺
Enables depolarisation when channels open
Perilymph
Surrounds the macula outside the membranous labyrinth
Normal extracellular fluid
Afferent nerve fibres
Carry signals from hair cells to the brain
Firing rate changes depending on stereocilia deflection
How the structure works together
Head tilt or linear movement → otoconia shift
Otolithic membrane moves
Stereocilia bend
K⁺ channels open or close
Hair-cell transmitter release changes
Vestibular nerve firing changes
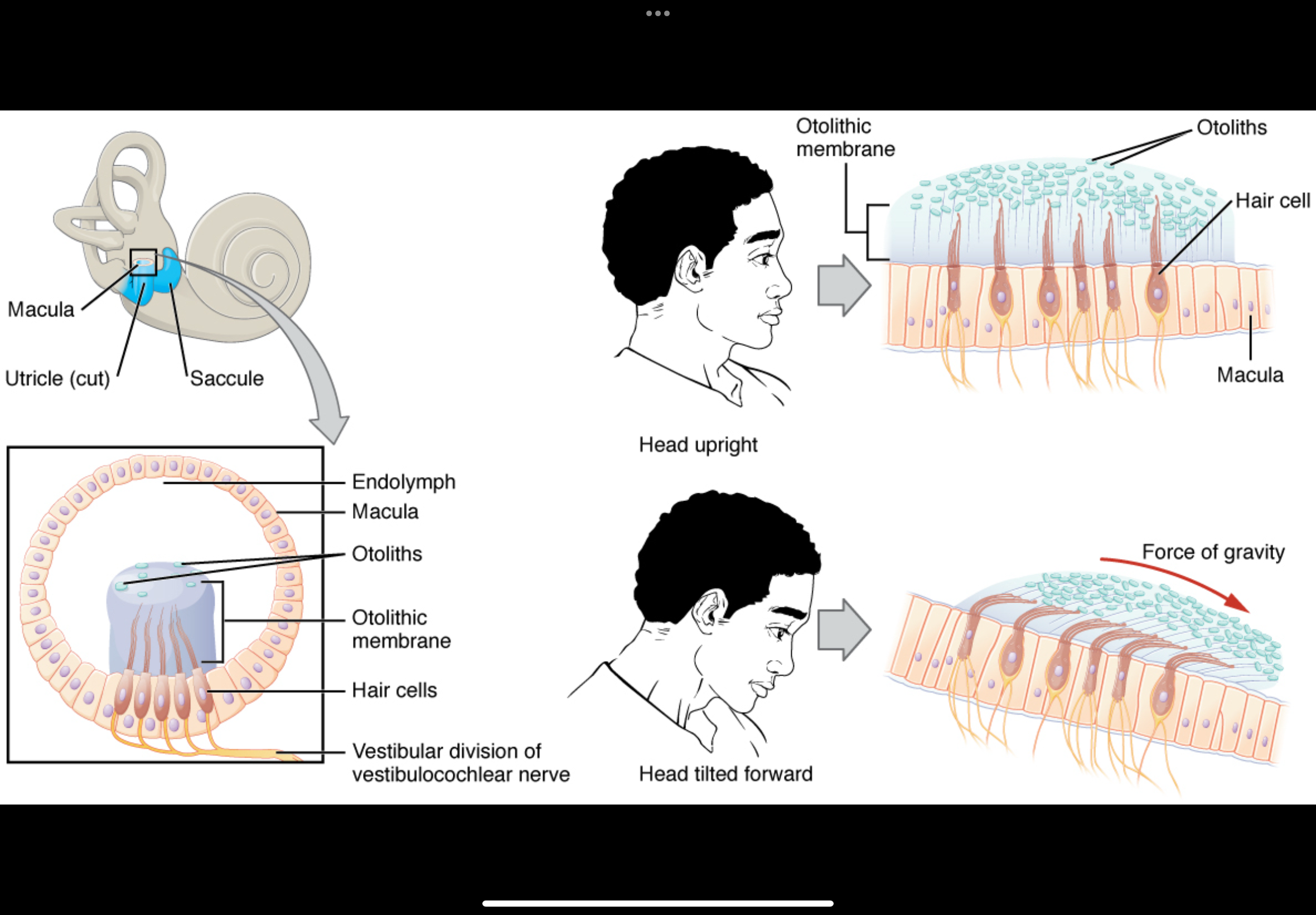
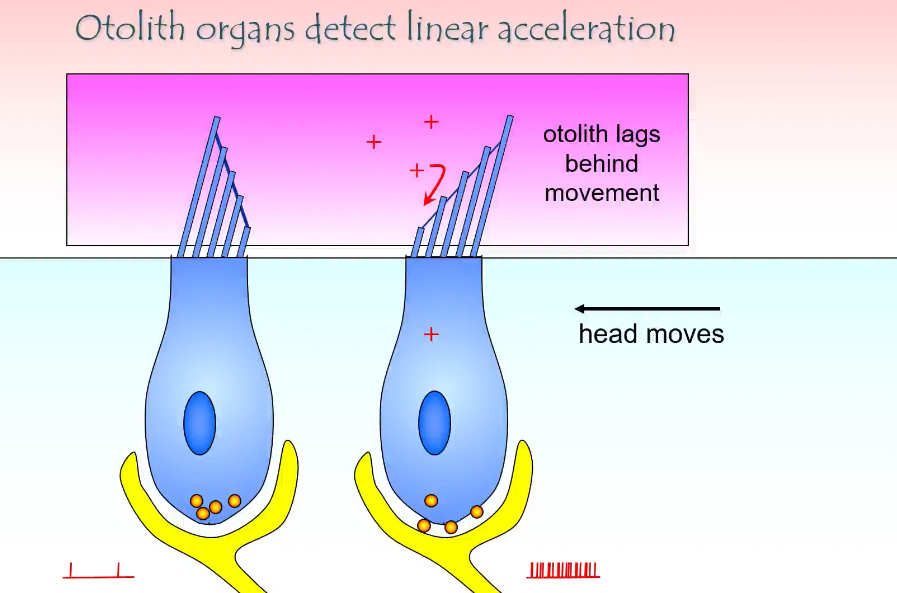
How do otolith organs detect linear acceleration?
This is the otolith system (utricle + saccule)
It shows how the ear detects linear acceleration (moving in a straight line) and head tilt
Key structures involved
Hair cells with stereocilia
Otolithic membrane (gel-like layer)
Otoconia (otoliths) on top of the membrane
Endolymph around the stereocilia
Afferent vestibular nerve fibres at the base
What happens when the head starts moving
The head moves (arrow in the diagram)
The otolithic membrane + otoconia lag behind
This lag happens because otoconia have mass and inertia
As a result, the otolithic membrane shifts relative to the hair cells
How hair cells are activated
Movement of the otolithic membrane bends the stereocilia
If stereocilia bend towards the tallest cilium:
Mechanically gated K⁺ channels open
K⁺ enters from endolymph
Hair cell depolarises
More neurotransmitter released
Afferent firing increases
If stereocilia bend away from the tallest cilium:
Channels close
Hair cell hyperpolarises
Less neurotransmitter released
Afferent firing decreases
What the + signs and spikes mean
+ signs = depolarisation of the hair cell
More spikes = increased firing in the vestibular nerve
Fewer spikes would indicate inhibition
Why “lag” is essential
If the otolithic membrane moved exactly with the head, nothing would bend
The lag is what converts motion into a signal
This allows detection of:
Starting to move
Stopping
Speed changes
Direction of movement
Otolith Organs: Hair Cells form an orderly pattern
Otolith organs detect head tilt and gravity by using many hair cells arranged in different directions
The brain works out direction from which hair cells are activated and which are inhibited
Orderly pattern of hair cells
Hair cells in the macula are not all aligned the same way
Each hair cell has a preferred direction (the direction that depolarises it most)
Different groups of hair cells are oriented at different angles
This creates a direction map.
What gravity does
Gravity pulls the otoconia in a constant downward direction
This shifts the otolithic membrane
The shift bends stereocilia, but:
Some hair cells bend towards their preferred direction → depolarise
Others bend away → hyperpolarise
What happens when you tilt your head
Tilting changes the direction of otoconia movement
A different set of hair cells is now excited
Another set is inhibited
So head tilt is encoded by a pattern of activity, not a single cell.
Role of utricle and saccule
Utricle: mainly covers horizontal directions
Saccule: mainly covers vertical directions
Together, they cover all possible head tilt directions
How the brain reads this
The brain compares:
Which afferent fibres increase firing
Which decrease firing
From this population pattern, it determines:
Direction of gravity
Direction of linear movement
Main outputs of the Vestibular System
Otolith afferents project via the vestibulospinal tract.
Targets:
Anti-gravity muscles of legs and trunk
Functions:
• Maintaining upright posture
• Preventing falls
• Compensation for linear disturbances
Semi Circular Canal System: Structure
Contains:
• Ampullary crest with hair cells
• Gelatinous cupula
• Endolymph inside the canal
• Afferent nerve fibres
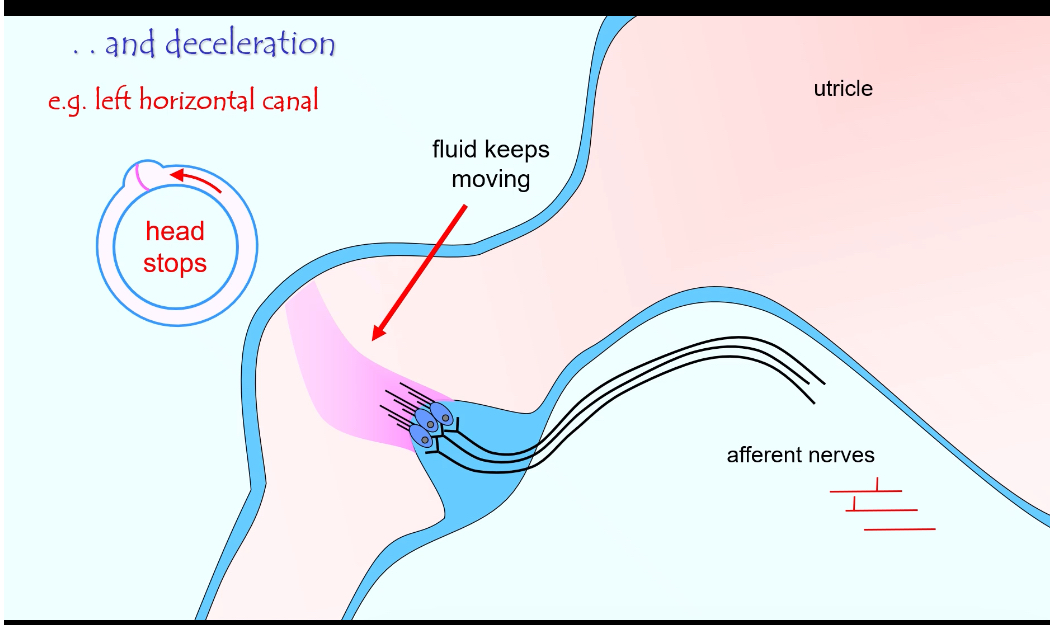
Semi Circular Canal: eg, left horizontal canal
What semicircular canals do
They detect head rotation (angular acceleration)
They tell you when you start, change, or stop turning your head
How it works
When the head starts to turn:
The canal moves with the head
The fluid (endolymph) lags behind
This bends the cupula
Hair cells bend → signal sent to the brain
When the head keeps turning:
Fluid catches up
Bending stops
Signal reduces
When the head stops:
Fluid keeps moving briefly
Cupula bends the opposite way
Brain senses deceleration
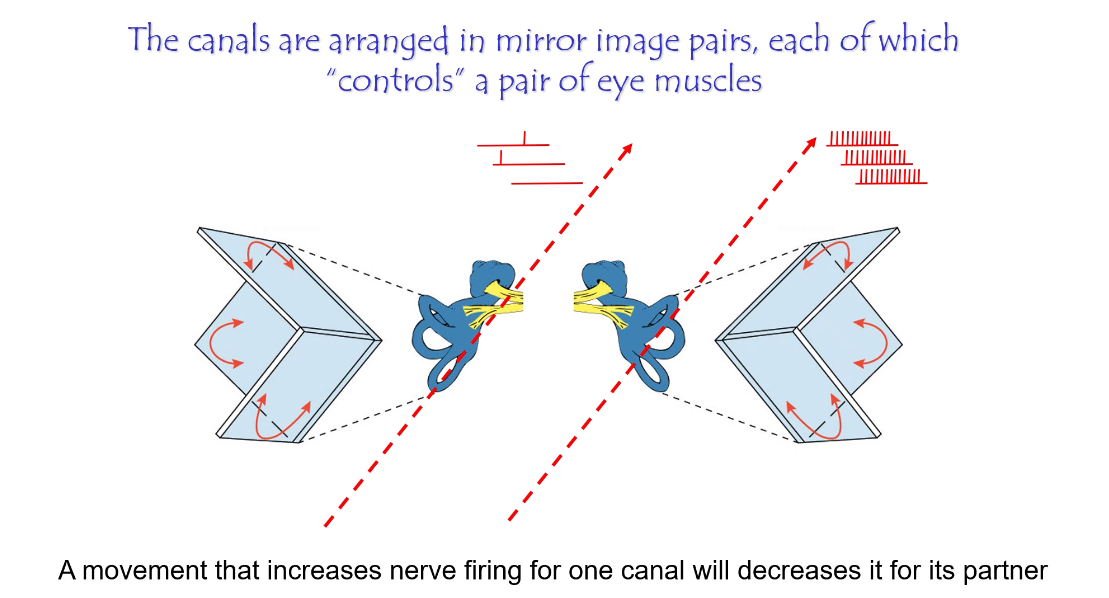
Why do we have three semi-circular canals?
They are arranged at right angles so that together they detect rotation in all three axes.
Each canal has a partner on the opposite side of the head.
Semicircular canals work in mirror-image pairs
When one canal is excited, its partner is inhibited
This push–pull system makes head rotation signals precise and fast
How the canals are paired
Each canal on one side of the head is paired with a canal on the opposite side that lies in the same plane
Examples:
Left horizontal ↔ Right horizontal
Left anterior ↔ Right posterior
Right anterior ↔ Left posterior
These pairs detect rotation in opposite directions.
What happens when you turn your head
Head turns to one side
Endolymph movement:
Excites one canal → firing rate increases
Inhibits the partner canal → firing rate decreases
The brain compares increase vs decrease, not absolute firing
This contrast tells the brain:
Direction of rotation
Speed of rotation
Neck and Shoulder muscles
Ampullary afferents → vestibulospinal tract → neck and shoulder muscles.
Role:
• Stabilising head position during movement
• Counteracting unwanted rotational disturbances
What is the vestibulo-ocular reflex (VOR)?
What the vestibulo-ocular reflex (VOR) is
A brainstem reflex that keeps vision stable during head movement
It moves the eyes in the opposite direction to the head
It works without needing the cortex, so it is extremely fast
Example:
Head turns left → eyes move right → image stays on the fovea
Where the signal starts
Semicircular canals detect head rotation
Hair cells change firing rate
Signal travels to the vestibular nuclei in the brainstem
Core pathway (horizontal canal example)
Horizontal semicircular canal activates
→ Vestibular nuclei
→ Abducens nucleus (CN VI)
Activates lateral rectus of one eye
→ via medial longitudinal fasciculus (MLF)
→ Oculomotor nucleus (CN III)
Activates medial rectus of the opposite eye
Result:
Both eyes move together in the opposite direction to head movement
Why the MLF is important
The medial longitudinal fasciculus links eye movement nuclei
It ensures both eyes move together
It must conduct signals very fast
Why the VOR must be extremely fast
Head movements are rapid
Visual feedback would be too slow
Delay would cause:
Blurred vision
Loss of fixation
So the pathway is:
Short
Heavily myelinated
Brainstem-based

What are conjugate eye movements and their cortical control?
Smooth pursuit movements that keep a moving object in focus.
Controlled primarily by the visual cortex projecting to brainstem eye movement centres.