Cell Signaling
1/45
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai | Chat |
|---|
No analytics yet
Send a link to your students to track their progress
46 Terms
Specific, Amplified, Tightly Regulated, and Coordinated
Cellular responses should have these four characteristics.
Gap Junctions
This type of junction allows small signaling molecules to directly diffuse from cytoplasm of one cell to adjacent cell. They allow fast communication between two cells. They are very important if you want highly coordinated actions from a group of cells.
Adhering & Tight Junctions
These junctions are important in the organization of tissues, especially during organ development and remodeling. They help cells know how they are organized in relation to other structures and cells beside it.
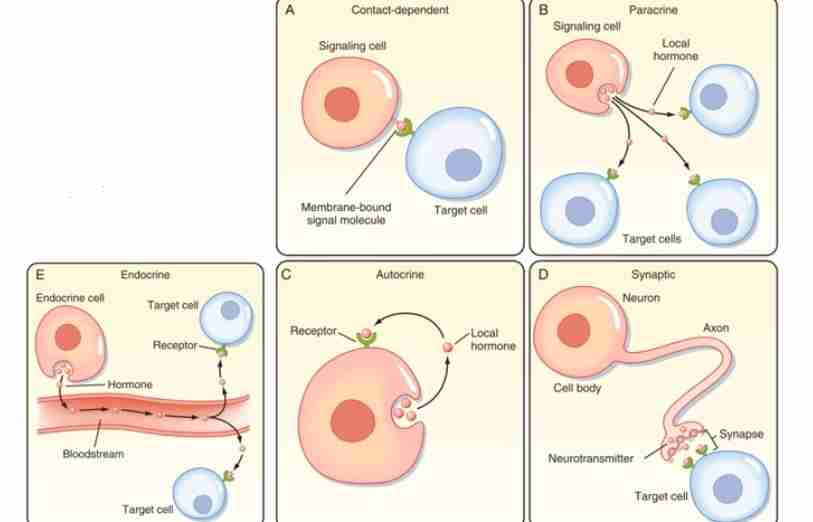
Contact-dependent Signaling
In this mechanism, the signal is found at the membrane or surface of the signaling cell. Here, it uses signaling molecules that is extracellular which is connected to signaling molecule. The signaling cell needs to come in contact with target cell for a cell response to happen. It can be mostly observed in immune cells.
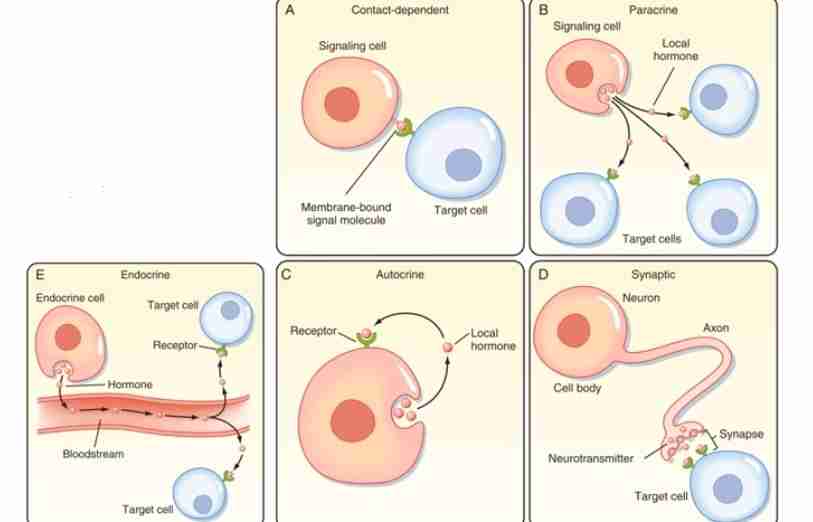
Paracrine Signaling
In this type of mechanism, the signaling cell sends hormones or signals to target close that are close by or nearby.
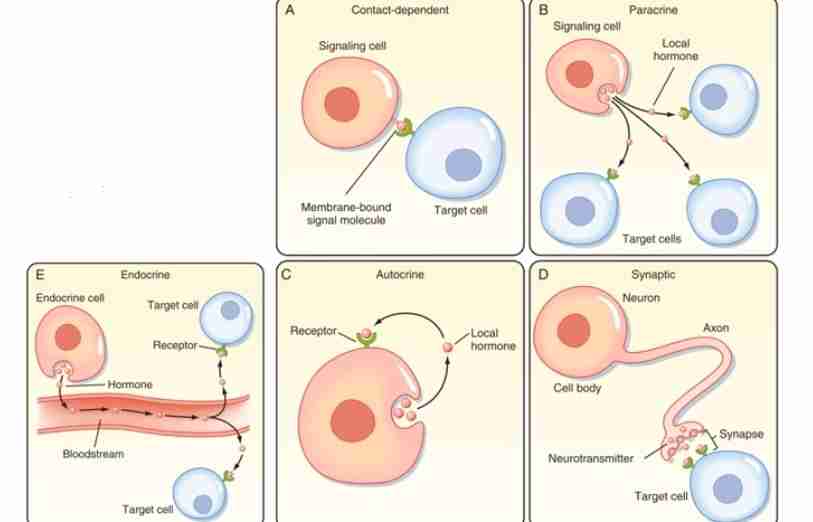
Endocrine Signaling
In this mechanism, the target cells are far away, thus hormone or signal travels via bloodstream. It usually produces a slower response but with a longer-lasting effect.
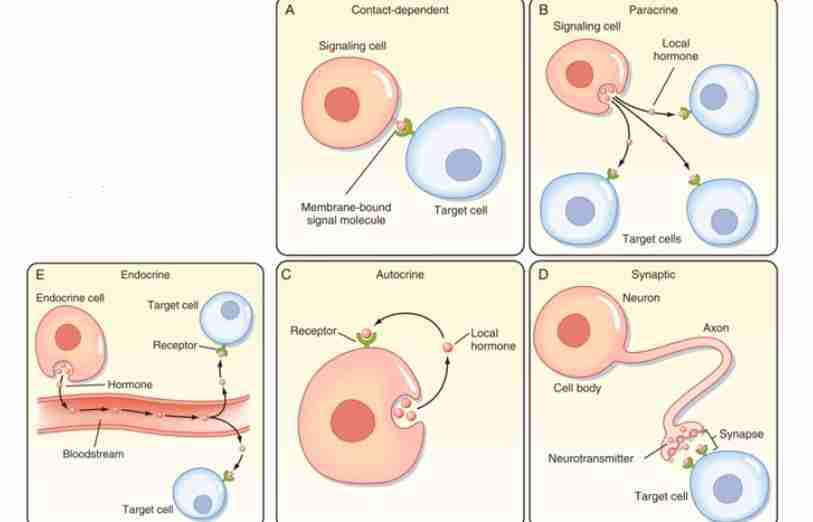
Autocrine Signaling
In this type of mechanism, a cell sends a signal or hormone that activates itself.
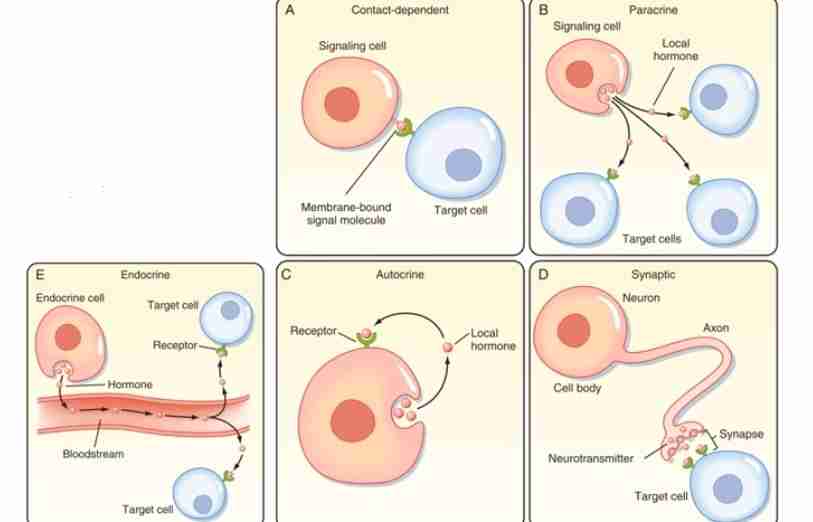
Synaptic Signaling
This type of mechanism is a highly specialized form of communication, which could be better understood through the study of nerves and the synapse.
Recognition
This step in the communication through signaling molecules posits that a specific signaling molecule will only attach to its specific receptor protein. If there's a signaling molecule but the cell lacks the receptor that responds to the signaling molecule, then the signal will be useless.
Transduction
This step in the communication through signaling molecules means that the signal will be converted from one form to another. The extracellular signal becomes an intracellular signal (second messenger).
Transmission
This step in the communication through signaling molecules is where the intracellular signals and the second messengers are transmitted to the effector or target proteins. It is an intracell step.
Modulation
This step in the communication through signaling molecules is where the target proteins or effector proteins are enhanced/inhibited by intracellular signaling proteins, usually by kinases or phosphotases (which attach or remove phosphate from proteins, which affects its functions).
Response
This step in the communication through signaling molecules is the summation and integration of input from multiple pathways.
Termination
This step in the communication through signaling molecules involves feedback mechanisms that may act at any level of the signaling pathway.
Ligand-gated Ion Channels
These receptors transduces a chemical signal into an electrical signal. When ligands open or close ion channels, ionic permeability changes. They are involved in synaptic signaling between electrically excitable cells such as the nerves and muscles.
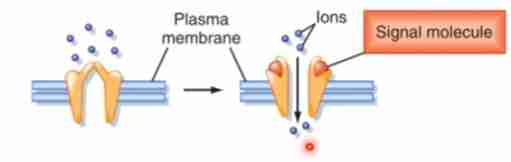
Ionotropic Receptor
This is another name for ligand-gated ion channel.
G protein-coupled receptors (GPCR)
This is the largest family of cell-surface receptors (>1000 members) which respond to a diverse array of signaling molecules. They are coupled or closely associated with heterotrimeric G proteins (GTP-binding protein).
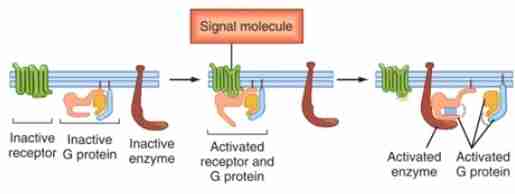
Seven-transmembrane receptors or Sepertine receptors
These are alternative names for the GPCR.
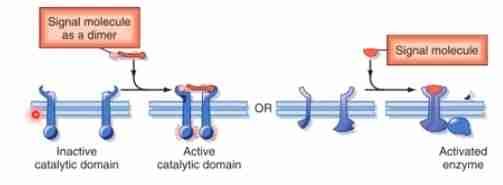
Enzyme-linked Receptors
These receptors that function as enzymes or are directly associated with enzymes.
Nuclear receptors
These receptors are found inside the cell; however, despite its name, some are located in the cytoplasm and the others in the nucleus. They are acted on mainly by hydrophobic molecules, and the activated receptors will bind to DNA to alter gene expression.
Second messenger
This is an alternative name for intracellular signaling molecules.
Scaffold protein
This type of protein helps keep the enzymes together in one place.
Bifurcate protein
This type of protein allows a signal to go to different pathways.
Kinases
This adds phosphate to protein, resulting to phosphorylation.
Phosphatases
These remove phosphate from protein, resulting to dephosphorylation.
GTPases
These enzymes can help alter the function of GTP. They are also responsible for GTP hydrolisis.
GTPase-activating proteins (GAP)
These activate GTPases, resulting to an inactive G protein.
Guanine nucleotide exchange factors (GEF)
These deactivate GTPase, resulting to an active G protein.
Monomeric G-proteins
These are small g-proteins as they are made up of only one subunit. They are central to many enzyme-linked receptor pathways. They also regulate multiple cell functions such as cytoskeletal organization, vesicular transport, gene expression, and apoptosis.
Heterotrimeric G-proteins
They are coupled to the GPCR. These are composed of three subunits: α, β, and γ.
Ras
This family of monomeric g-proteins is involved in pathways that control cell division, cell proliferation, and cell death.
Rho
This family of monomeric g-proteins regulate actin cytoskeletal organization.
Rab
This family of monomeric g-proteins regulates intravesicular transport and trafficking of proteins between organelles.
Ran
This family of monomeric g-proteins regulate nucleocytoplasmic transport of RNA and proteins.
Arf
This family of monomeric g-proteins regulates vesicular transport.
G Protein and Phospholipase
When G protein alpha activates phospholipase C, it breaks down phosphatidylinositol biphosphate (PIP2) into inositol triphosphate (IP3) and diacylglycerol (DAG).
Diacylglycerol (DAG)
This activates Protein Kinase C (PKC) and may further be converted to arachidonic acid.
Arachidonic Acid
They may act as a secondary messenger or may be further converted to eicosanoids.
Eicosanoids
These are a large class of a certain type of secondary messenger molecule. Some examples include prostaglandins (immune function), thromboxanes (platelet function), leukotrienes (inflammation).
G proteins and phosphodiesterases
Phosphodiesterases break down cAMP and cGMP to AMP and GMP. They help reverse or terminate the action of cAMP and cGMP.
Receptor Guanylyl Cyclases
Some examples of this enzyme receptor-linked pathways are the atrial natriuretic peptide (ANP) and nitric oxide (NO). These catalyze generation of cGMP from GTP.
Receptor Serine/Threonine Kinases
An example of this is the transforming growth factor beta (TGF-[beta sign]). Through this pathway, the ligand binds to the Type II subunit, which will then phosphorylate the Type I subunit. Serine/Threonine Kinase in Type I is activated and will continue to phosphorylate other effector proteins, thus eliciting cellular responses.
Autophosphorylation
This process is when the phosphorylation of each other domains occurs.
Receptor Tyrosine Kinase
Some examples of this enzyme receptor-linked pathway include nerve growth factor (NGF) receptors, and insulin receptors. Not all of these are dimers as some of these receptors are tetrameric.
Tyrosine Kinase-Associated Receptors
These receptors have no intirinsic tyrosine kinase activity, but are only associated with tyrosine kinases. These receptors can help alter gene transcription, as JAKs will phosphorylate STAT proteins, therefore affecting gene transcription. Some examples include the cytokine receptor and the growth hormone receptor.
Receptor Tyrosine Phosphatases
This receptor dephosphorylates the tyrosine residues phosphorylated by tyrosine kinase. An example of this is the CD45 protein, which dephosphorylates and activates some tyrosine kinase-associated receptors in lymphocytes.