Gastrulation and segmentation in vertebrate embryos
1/25
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced |
|---|
No study sessions yet.
26 Terms
Establishment of the basic body plan
Symmetry and segmentation of vertebrates
Vertebrates are bilaterians- animals with bilateral symmetry
anterior: posterior (AP) body axis
Serial repetition of parts along the body axis = segmentation
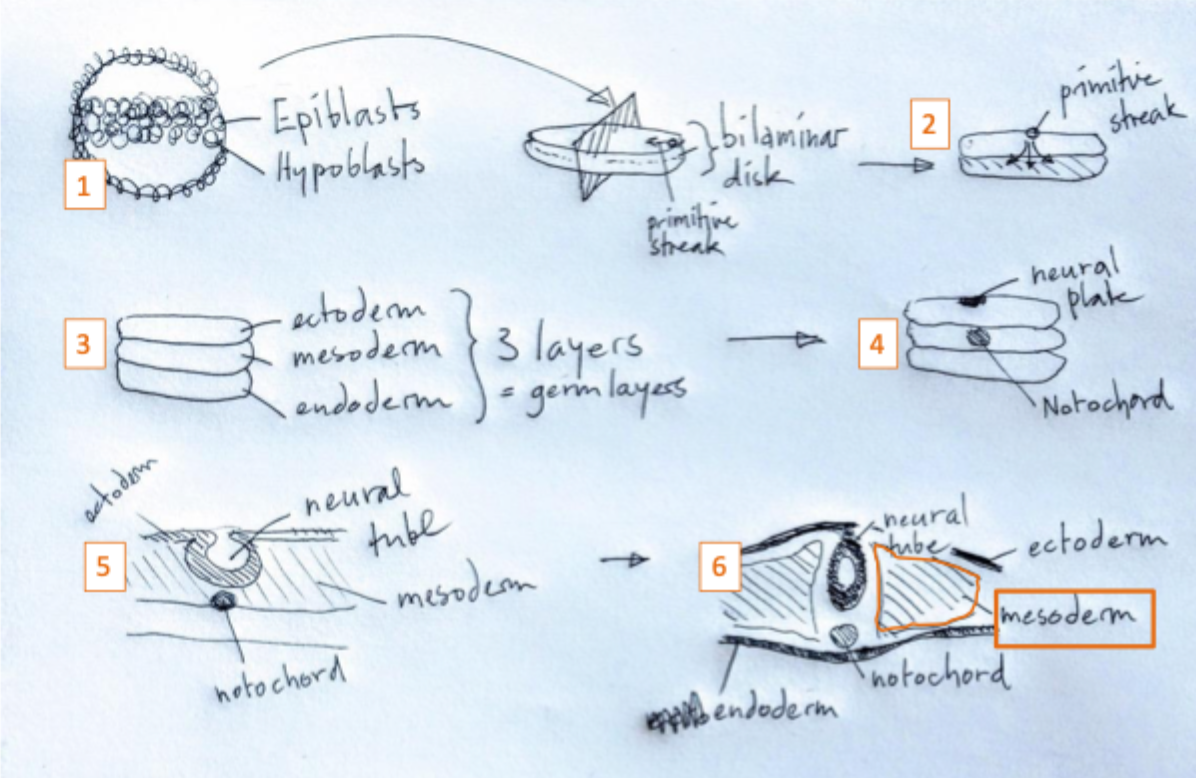
Gastrulation and the formation of the 3 germ layers
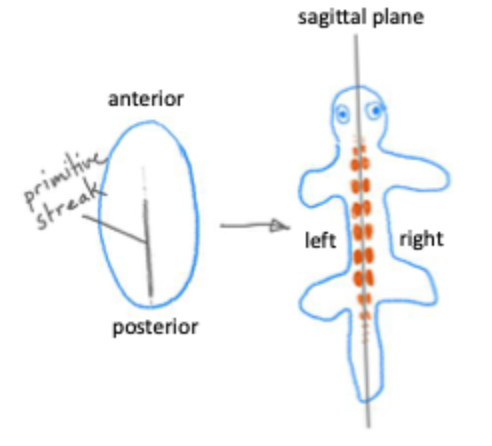
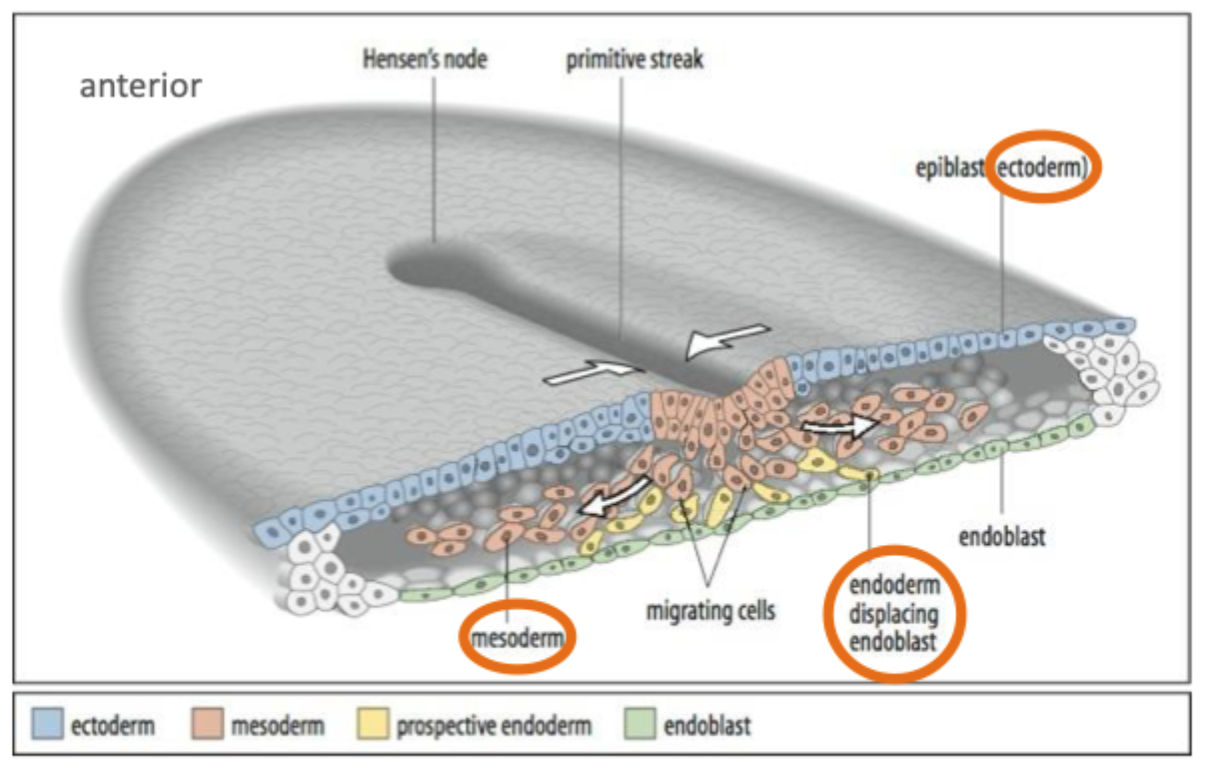
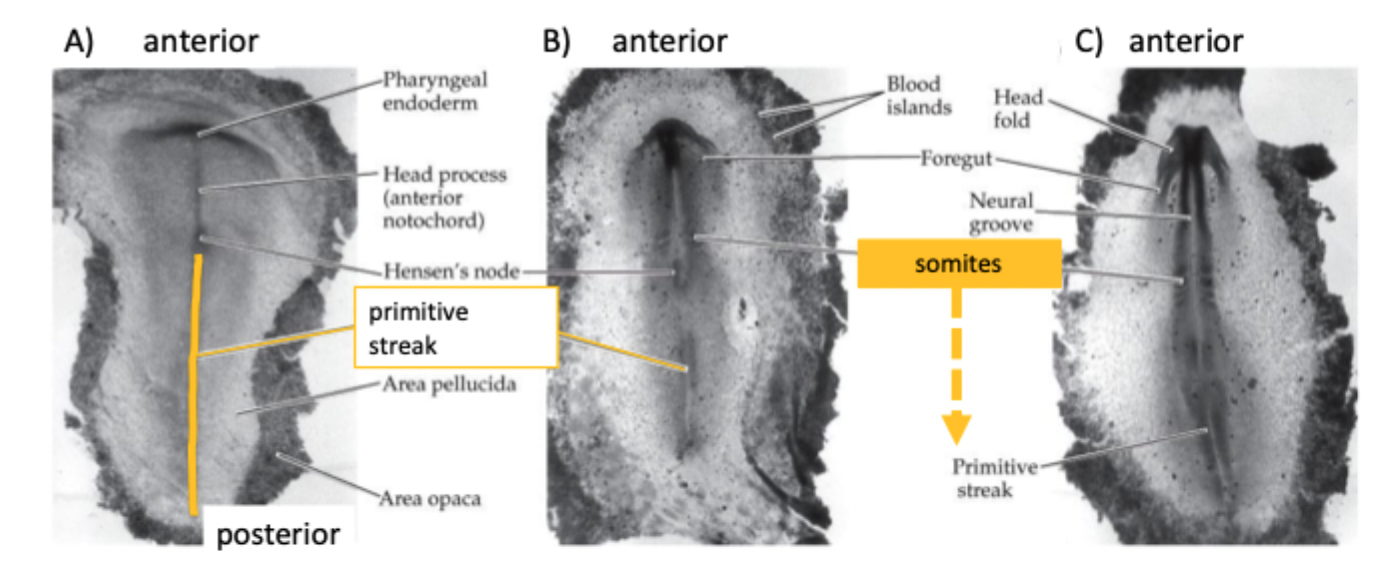
During gastrulation cells from the epiblast migrate into the primitive streak
Here they undergo an epithelial to mesenchymal transition and migrate under the epiblast
These cells give rise to the endoderm and the mesoderm
Gastrularion and the formation of the 3 germ layers
Germ layers
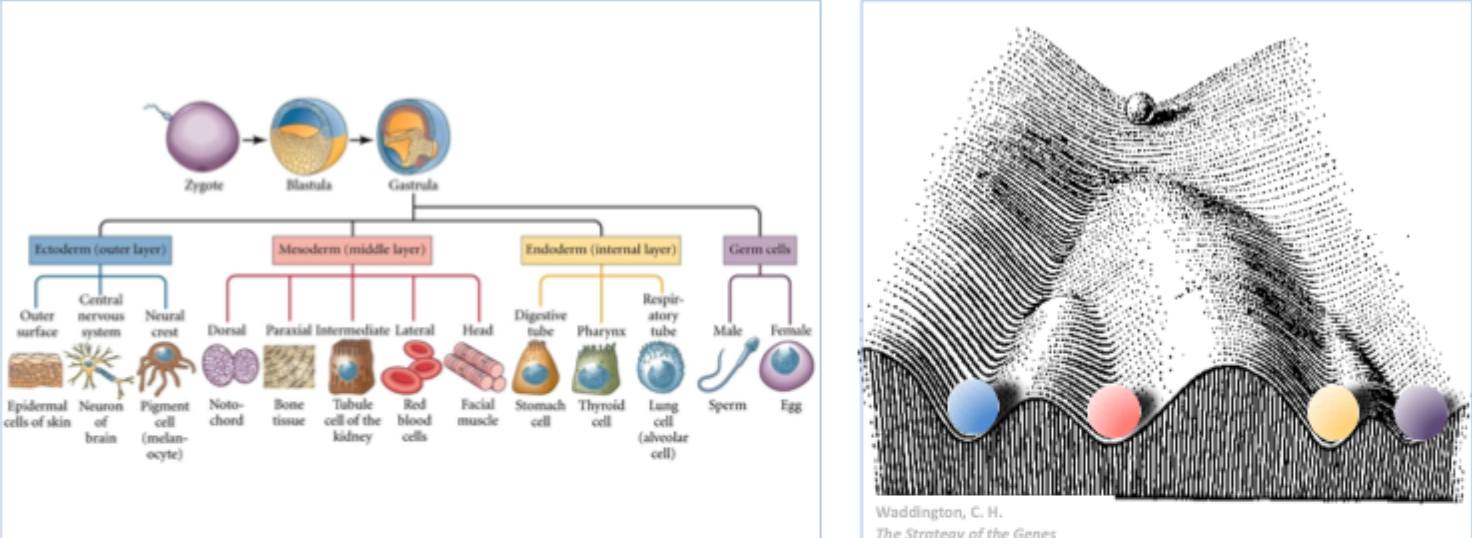
Structure of the embryo
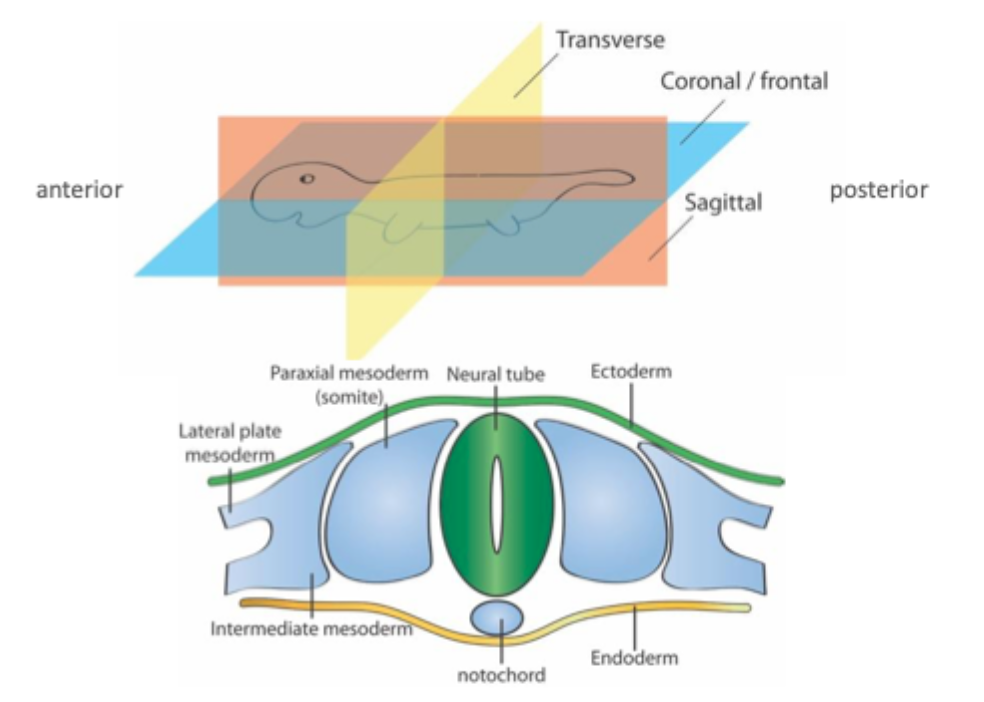
Segmentation
Segmental organisation is found in many diverse animal groups including vertebrates
Segmentation is an efficient way to generate diverse body parts from similar basic structures during development
Somites develop from unsegmented mesoderm
Somites are balls of epithelial tissue that bud off from the unsegmented paraxial mesoderm
They start forming at the anterior end and are added to the posterior
They form in a regular pattern with a set periodicity (1 every 90 mins in chickens, 1 every 2 hrs in mice)
Axis extension
The axial skeleton (with the exception of the skull) arises from somites
Somites start to form at the anterior end of the embryo and are then added in pairs:
sequentially
periodically
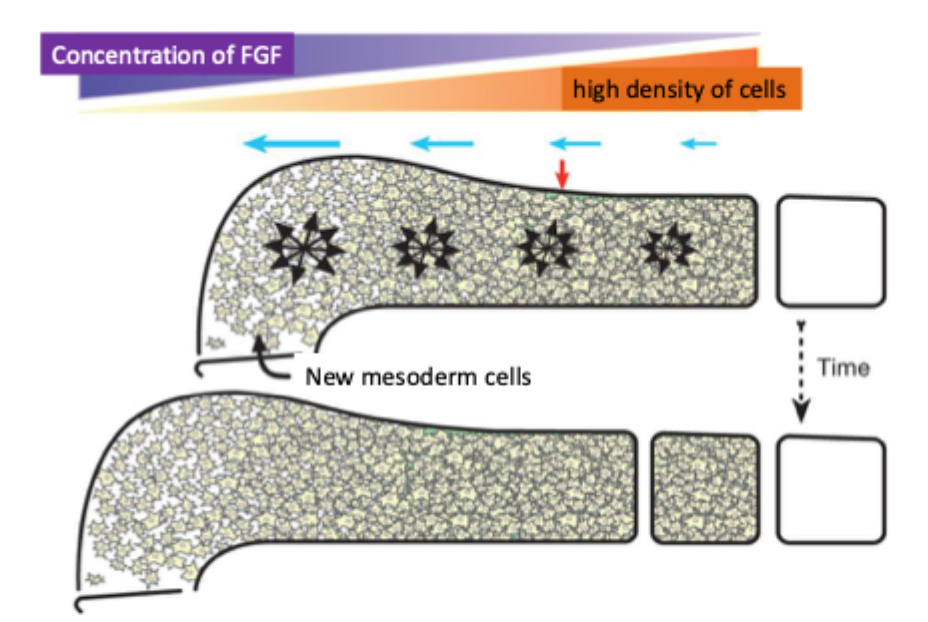
Fibroblast growth factors (FGFs) control cell motility and elongation of the embryo
Example
FGF8 is expressed in a gradient in paraxial mesoderm = PSM
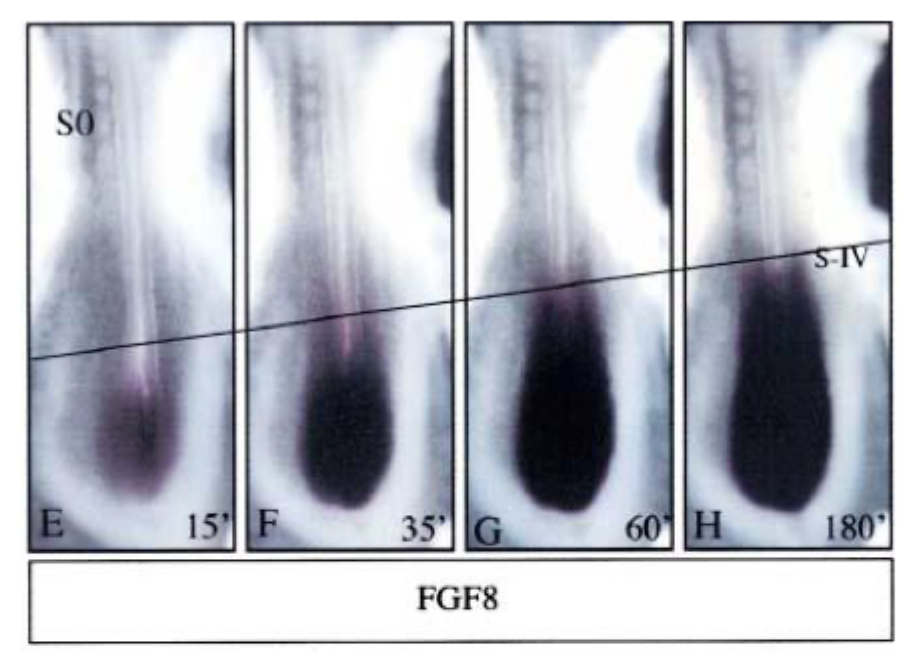
Experiment to show that FGFs play a role in normal somite formation
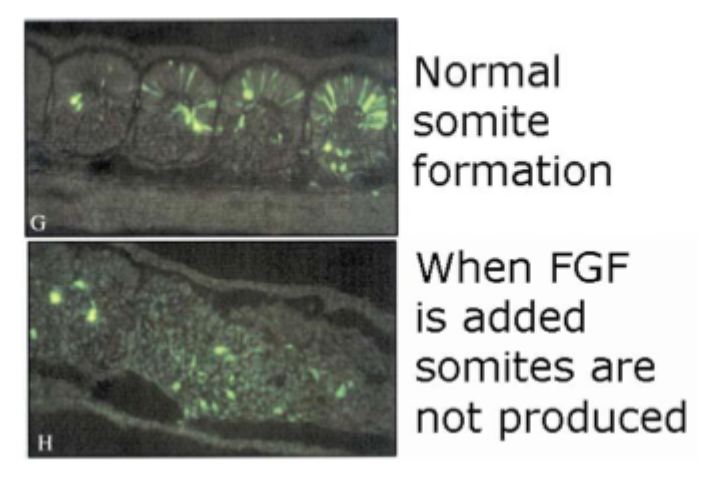
Experiment to show that FGFs play a role in normal somite formation pt2
High concentrations of FGF in the posterior inhibit the formation of somites
Decay of FGF mRNA produced a gradient of FGF
At low concentrations (anterior) somites can form
However, this does not explain the precise timing of somite formation
Another component is required to act as a ‘clock’ to generate this periodicity

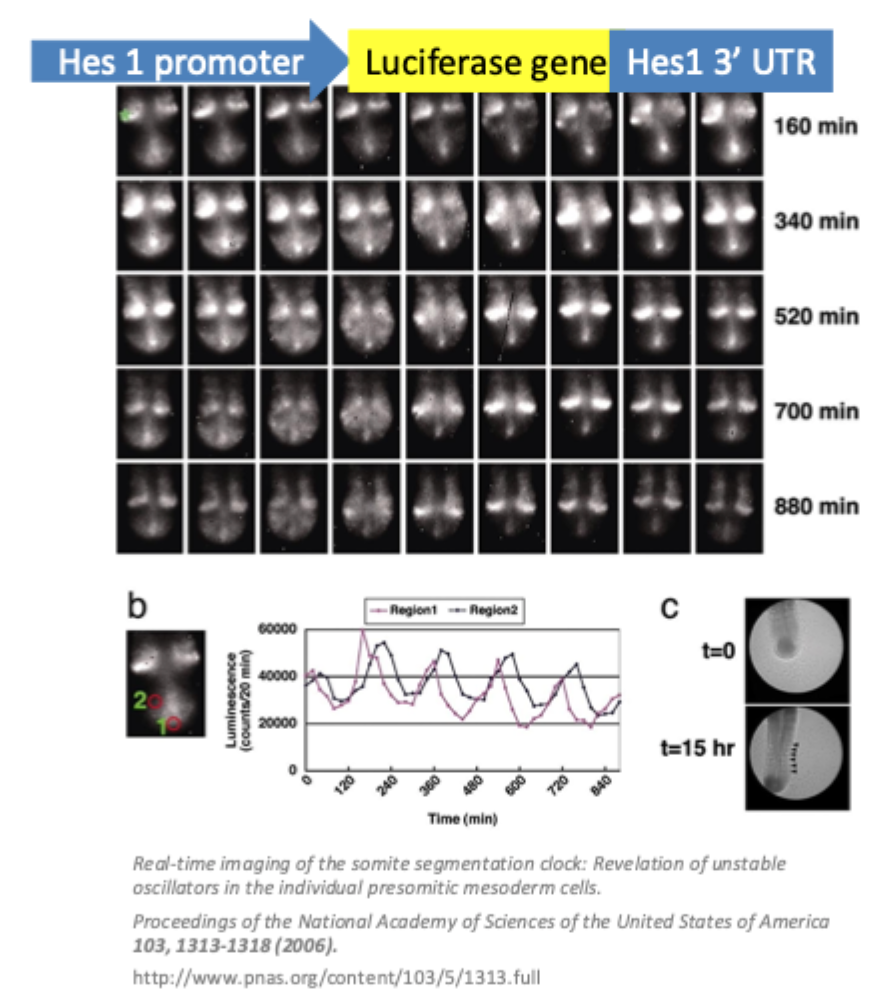
Component of the segmentation/ somite clock
Hes1 is cyclically expressed in presomitic mesoderm (PSM)
Hes1 is a transcription factor
It is a vertebrate homolog of the hairy gene in Drosophilia
This contributes the somite segmentation clock
Segmentation summary
Segmentation produces a repeated array of tissues along the embryonic axis
This is controlled by a ‘clock and wavefront’ process producing regular pulses of segmental activity- originally proposed as a theoretical model of segmentation
Segmental identity
Each segment has a specific indentity which determines the nature of the tissues being formed
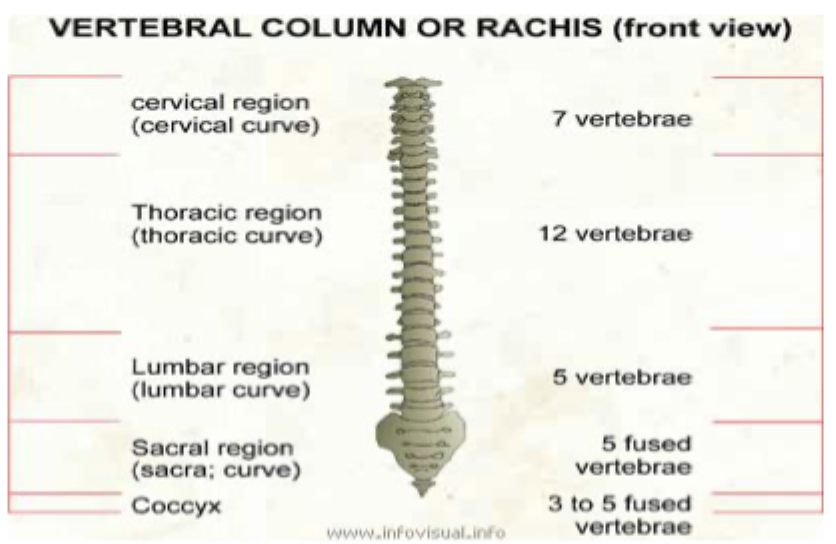
This is clearly seen in the vertebrae- cervical vertebrae lack rib attachments for example
In any species this pattern is highly conserved
Segmental identity is determined before segmentation
If pre- segmented mesoderm is grafted to a different position along the axis it retains its original identity
E.g. thoracic level mesoderm will form thoracic vertebrae with ribs
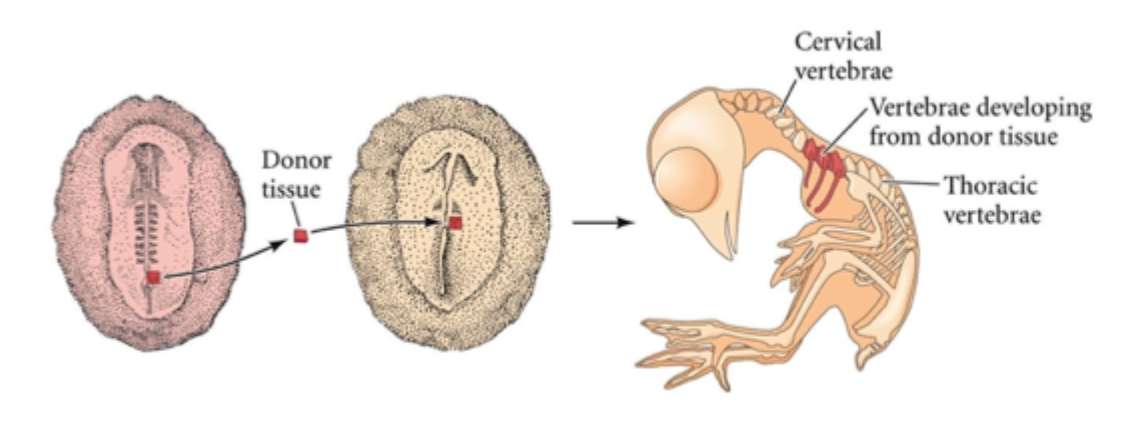
How genes play a key role in establishing of the morphological identity of somites and the vertebrae
How genes encode transcription factors
Regular aspects of morphogenesis in animals
Evolutionarily conserved
How gene → target gene
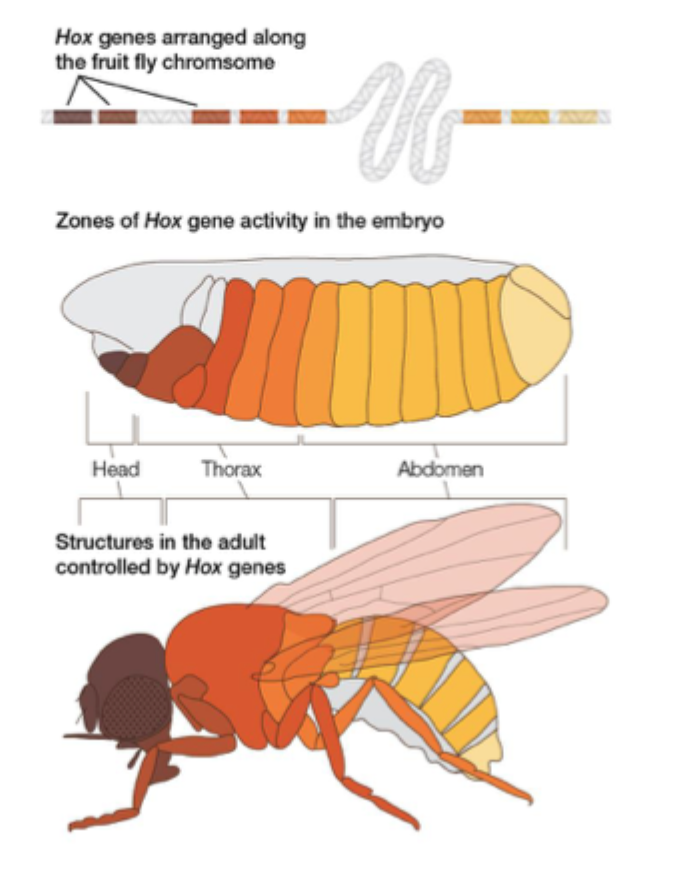
Hox genes in diverse animals
Comparison of hot genes in Drosophila and mice shows a remarkable degree of conservation of this mechanism
Hox genes are used across the animal kingdom to define region of the animal
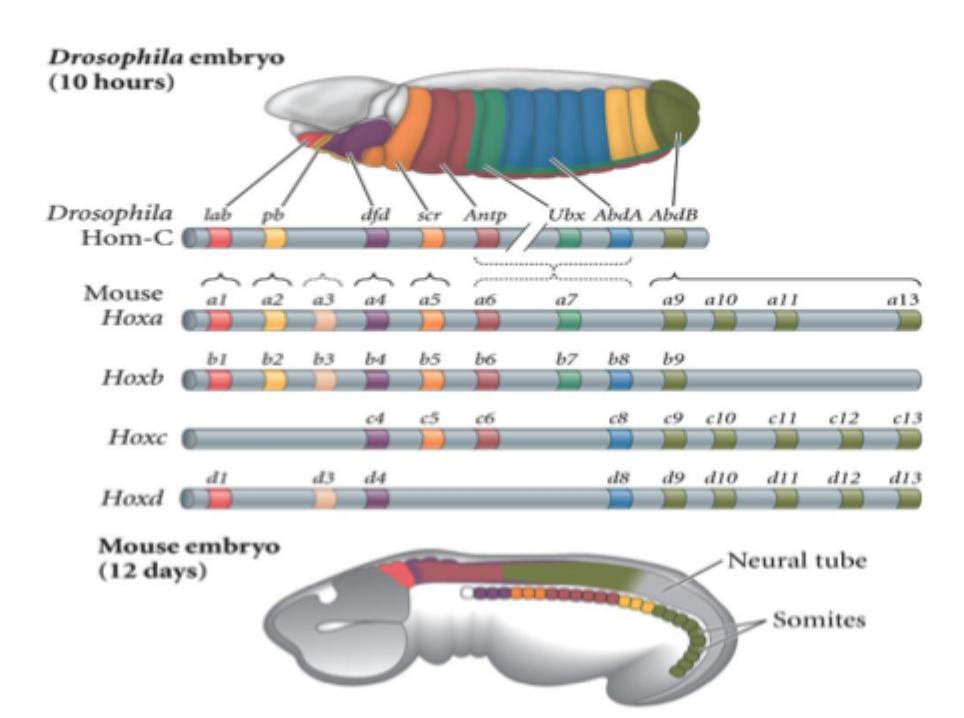
How gene expresion domains and vertebral formula of the mouse embryo
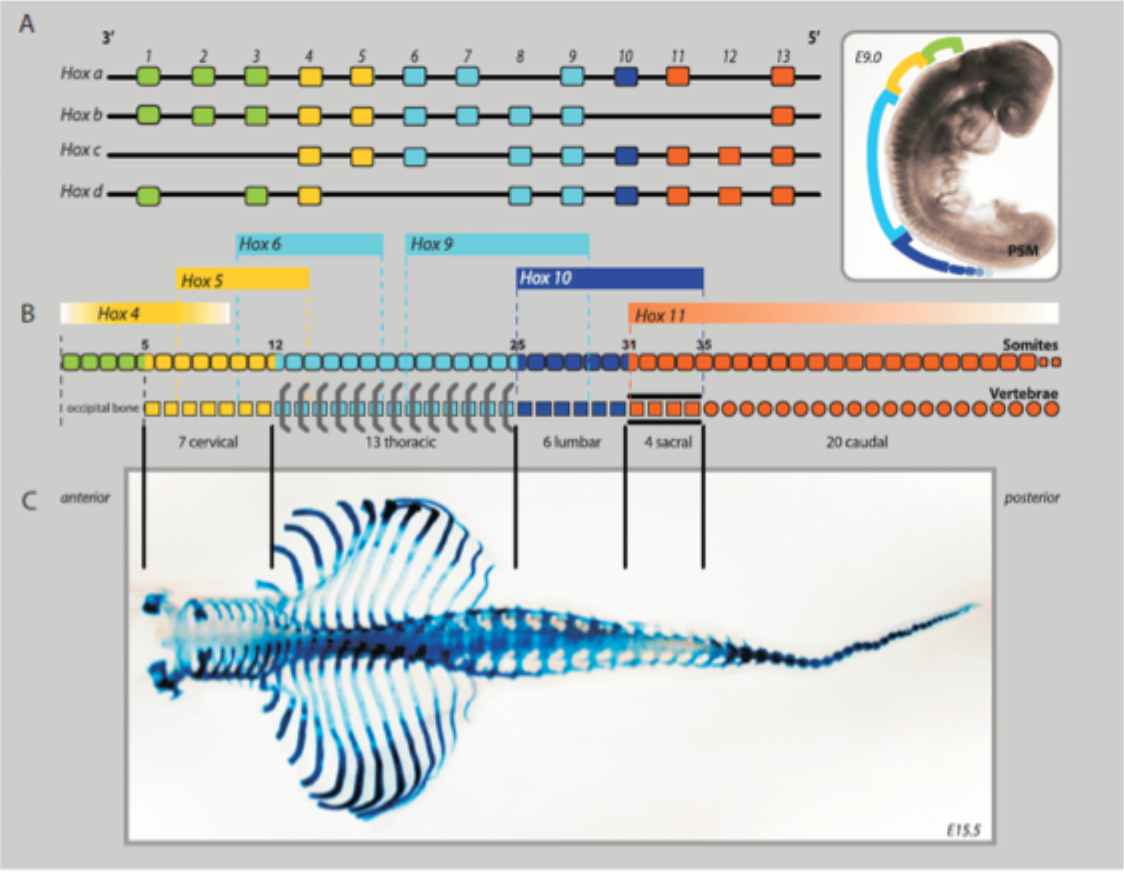
Segmental identity
Although related species show similar patterns there are differences
These differences are 1 way that evolution can generate differences between species
Mice (and all mammals except sloths and manatees) have 7 cervical vertebrae. Chickens have 14, swans 25 and elasmosaurus had 76
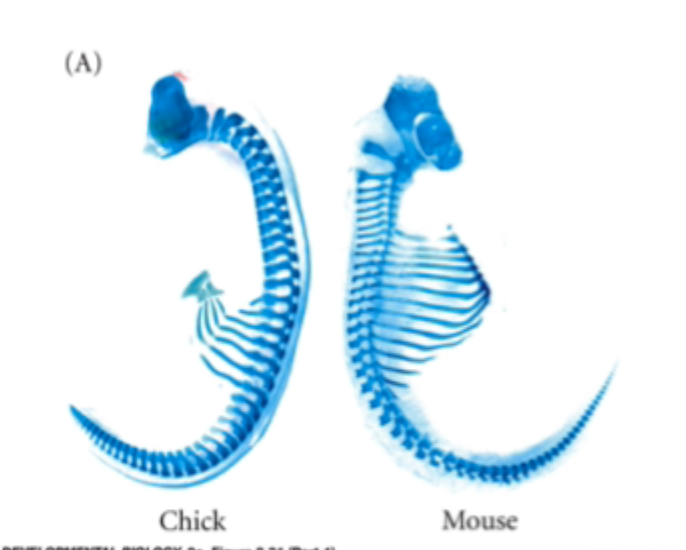
Segmental identity and gene expression patterns
Changes in veterbral identity correlate with boundaries of expression of homeobox (Hox) genes
This can explain changes in vertebral patterning e.g. more cervical vertebrae in birds correlate with longer expression domains of anterior hot genes
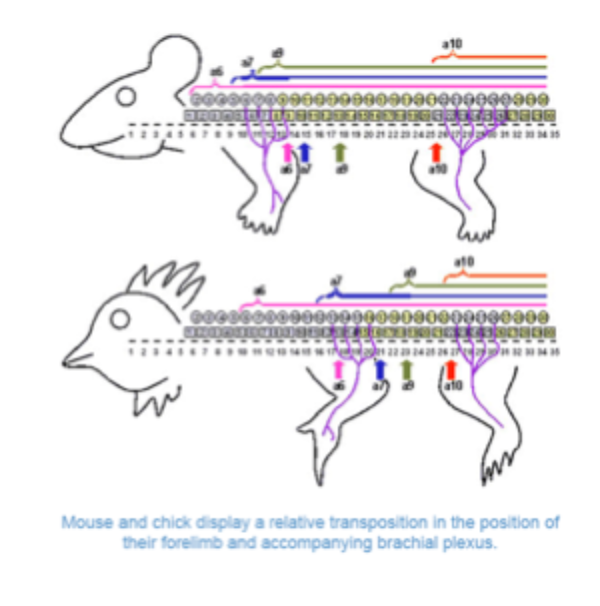
Mutations in how genes
Further evidence for the role of hot genes has come from mouse knockouts
In mice where hot genes have been mutated changes in the identity of vertebrae are seen
Mice lacking the hox 10 genes show lumbar to thoracic transformations, mice lacking hot 11 genes show sacral to lumbar changes
Linking gastrulation and axis patterning
Hox genes are differentially expressed in the primitive streak
Cells expressing anterior hot genes are only found in the early streak and the anterior mesoderm
More posterior hox genes are expressed in the streak for longer but are not seen in the anterior mesoderm even though cells in the streak express these genes
Linking gastrulation and axis patterning
The control of axial identity is linked to gastrulation
Cells expressing more posterior hot genes leave the streak later and form more posterior somites
Summary
The anterior- posterior axis in vertebrates is organised segmentally
These segments are generated by sequential addition of somites
This identity is determined during gastrulation as cells emerge from the primitive streak