M2L2 Transcriptional/translational control
1/29
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
30 Terms
What is the role of AP-1?
TF that regulates genes related to proliferation, differentiation, apoptosis, inflammation
Target of MAPK cascade
Made of of products of two JUN and FOS family proteins
Both contain basic leucine zipper domains that facilitate their binding as dimers to response element in target genes
Activated in response to GFs, ROS and radiation, with the specific combination of dimers influencing the biological response
What is the role of AP-1 oncogenes in cancer?
AP-1 oncogenes c-JUN and c-FOS cause aberrant expression and inappropriate increase in expression of AP-1 regulated genes
Associated with dysfunction and onset, development, invasion, and migration of cancer
Involved in drug/radiation resistance
What is the function of Myc and Max?
Belongs to family of helix-loop-helix TFs that act as heterodimers to modulate the transcription of target genes with E-box sequences (5’-CACGTG-3’)
Myc-Max complexes act as enhancers also also promote elongation of nascent transcripts by releasing RNA pol II tcomplexes from pause sites immediately downstream of TSS, allowing it to proceed with transcription and synthesise full length pre-mRNA transcripts
As cells differentiate the Mad protein levels increase, displacing Myc and reducing the expression of Myc-Max
Mad-Max complex prevents RNA pol II release, losing its ability to stimulate transcription, perimitting cells to enter post-mitotic differentiated state
Under normal conditions Myc is ubiquitinated and degraded - short 15-20 min half life
Myc-Max heterodimeric TF complex —> expression of 1,000+ genes which can also affect the cell cycle, favouring proliferation over differentiation
In healthy cells the Myc proto-oncogene is regulated downstream of many signal transduction pathways including Wnt, Hedgehog, Notch, TGF-β and many RTKs
What is the role of Myc-Max in cancer?
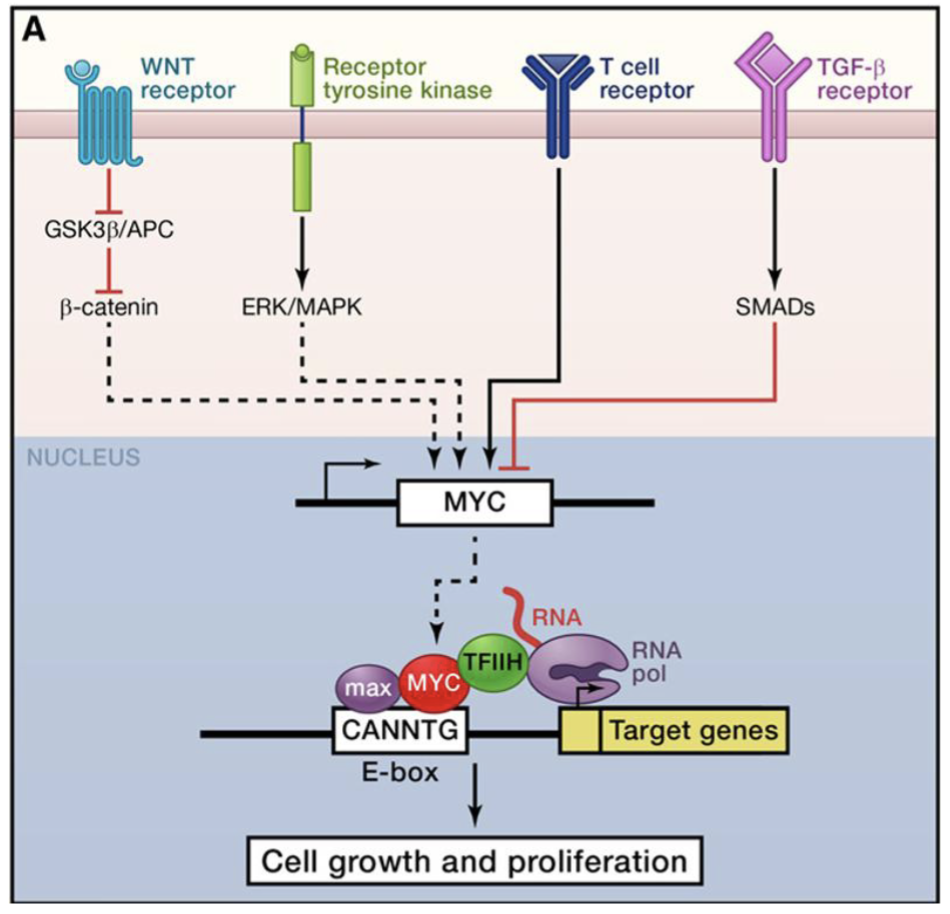
Myc activation in cancer can result from loss of this regulation, eg mutations in Wnt signalling APC, or direct alterations of Myc gene, eg. amplification or chromosomal transclocation
Acute sustained Myc expression —> inappropriate increase in transcription of Myc-regulated genes and E-box driven genes, unregulated cell cycle checkpoint progression, genomic instability due to Myc induction of ROS via mitochondrial biogenesis, and increased metabolism
Myc represses genes involved in cell-cell binding while cells undergo mitosis, linked to EMT and metastasis
What is the chromosomal translocation in Burkitt’s lymphoma?
Myc gene translocates from Ch8 to Ch14, falling within the regulation of strong promoter of immunoglubulin genes
What is the role of steroid hormone receptors?
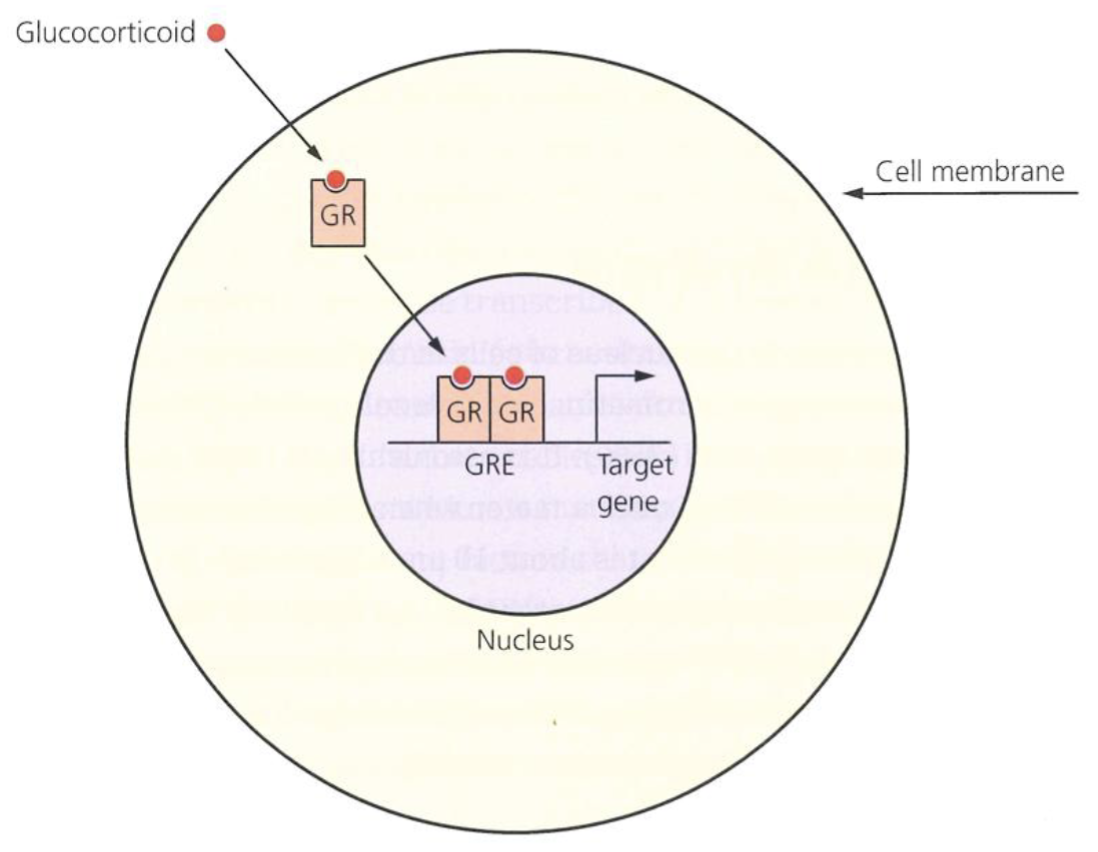
Steroid hormones pass through the cell memebrane and bind to their particular intracellular steroid receptors (aka nuclear receptors) in the cytoplasm
Acts as ligand dependent TF
Contain zinc finger type DNA binding domain, ligand binding domain for a specific steroid molecule, and dimerisation domain as they activate transcription as a dimer
What is the role of retinoic acid signalling?
Retinoic acid receptor (RAR) is a part of the steroid hormone receptor family and acts as RA-dependent transcriptional regulator - important during differentiation
RAR constitutively located in nucleus and acts as transcriptional repressor in the absence of RA
Binds to RA response element (RARE) in target genes as heterodimer with another member of the family called RXR
Aberrant forms of RARs, often due to chromosomal translocations, are characteristic of several leukaemias
lncRNA
Endogenous polyadenylated ncRNAs which are >200nt and lack an open reading frame
What are the functions of lncRNAs?
Transcribed from gene enhancers and modulates gene activation and silencing, X chromosome inactivation, alternative splicing, and post translational regulation
Involved in epigenetic modifications and are themselves epigenetically regulated
Thousands ofn lncRNAs are differentially expressed in tumours vs normal tissue
The steroid receptor activator is a lncRNA
They can act as molecular scaffolds to guide chromatin modifying enzymes (eg. HOTAIR or DLX6AS24)
Can act as competing endogenous RNAs (ceRNAs) that sponge miRNAs or proteins
They may inhibit long range chromatin interactions
Can function through the act of transcription itself
Additional mechanisms emerging - eg. orchestration of nuclear archiecture, forming circular lncRNAs, destabilising interacting mRNAs
What is the fucntion of lncRNA-p21?
lncRNA-p21 involved in downstream effects of p53
In DDR, p53 binds to promoter of lncRNA-p21 and mediates repression of genes involved in cell cycle arrest and apoptosis
What is the fucntion of HOTAIR lncRNA?
HOTAIR lncRNA is transcribed from HoxC gene cluster at Ch12, interacts with polycomb repressive complex 2 (PRC2) at 5’ end and lysine specific demethylase 1 (LSD1) complex which induce epigeneticc aleterations and cause gene silenceing/oncogenesis
HOTAIR dysregulation associated with cancer progression in 26 tumour types and predictive of metastasis in breast cancer
Can act as a sponge by binding/inhibiting specific repressive miRNAs
miRNA
Small ncRNA, 18-25nt, regulates mRNA expression
What are the steps of miRNA biogenesis and function?
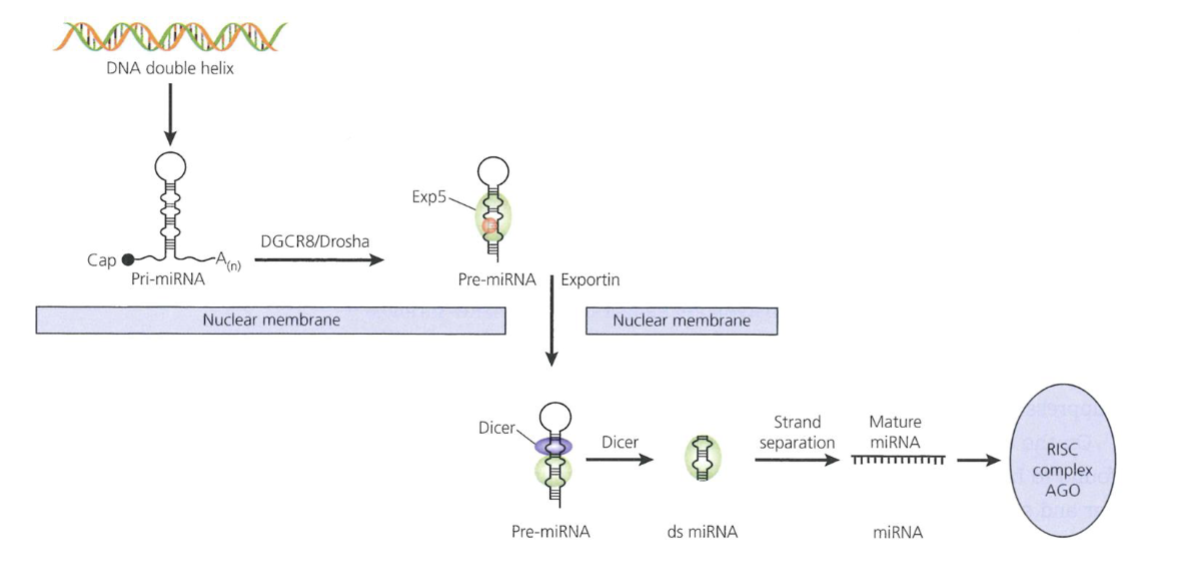
After being transcribed by RNA pol II from intergenic or intronic regions, primary transcript is processed by ribonucleases DGCR8 and Drosha in the nucleus, producing pre-miRNAs (hairpin shaped, 70-100nt intermediate)
Exportin-5 transports pre-miRNA into cytoplasm to by processed by the ribonuclease Dicer into double stranded miRNA
Strands separate and the mature single stranded molecule joins RNA-induced silencing complex (RISC) which contains an endonuclease component Ago which can degrade target mRNA
miRNA can hybridise perfectly to the 3’ UTR of the mRNA within RISC, causing mRNA cleavage/degradation, or it may bind to imperfect complementarity sites in the 3’ UTR in the RISC to block translation
How can miRNAs be implicated in cancer?
Reduction/deletion of tumour suppressing miRNAs promotes tumourigenesis, which could happen due to defects at any stage of miRNA biogenesis (amplification of miRNA gene, constitutively active promoter, increased processing or stability of miRNA)
Some miRNAs may act as oncogenic oncomirs (aka oncomiR) which function to suppress tumour suppressor mRNAs may be amplified in cancers
Evidence that miRNA expression profiles may help discriminate different types of cancers, may be useful for diagnosis/prognosis where mRNA profiles fall short
What is the function of the Let-7 family of miRNAs?
Let-7 family of miRNAs repress Ras oncogenes, refuced in lung, breast, urothelial, and cervical cancers
What is the function of miR-21
miR-21 inhibits apoptosis and is upregulated in glioblastoma and breast cancers
What is the function of miR-155?
miR-155 cooperates with Myc to promote B cell malignancies, overexpressed in Burkitt lymphoma
What are the broad steps involved in translation?
Pre-initiation complex (43S)
mRNA recruitment
43S recruitment to mRNA
Scanning and AUG selection
Formation of 80S initiation complex
Elongation and termination
Recycling
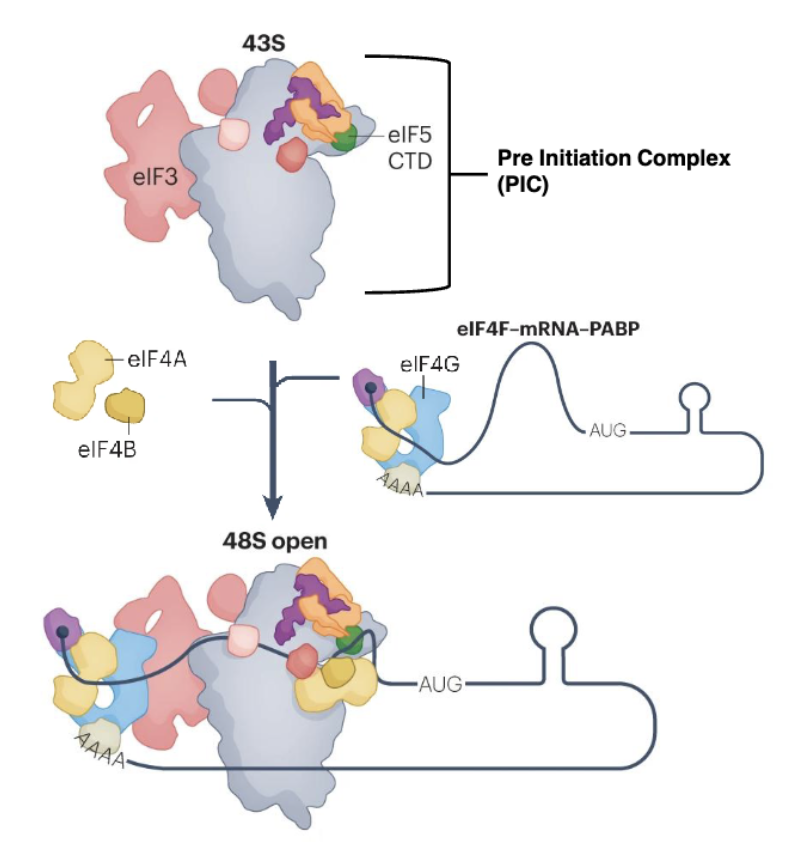
How is the 43S PIC formed?
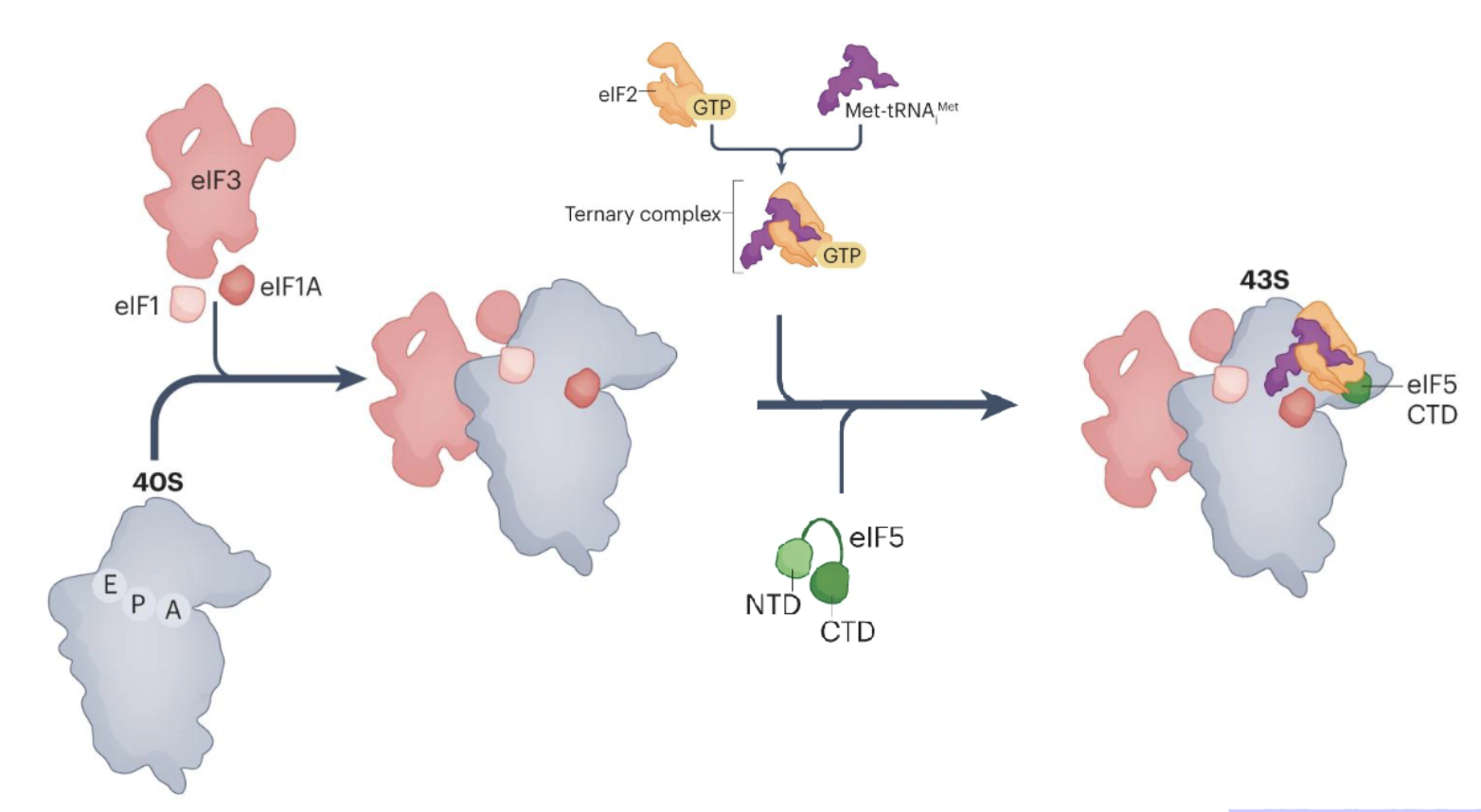
Binding of eukaryotic translation initiation factor 1 (eIF1), eIF1A, and eIF3 to the 40S subunit
eIF5 and ternary complex of eIF2, GTP and met-tRNA then bind to form a 43S translation PIC
What are the steps in mRNA recruitment for translation?
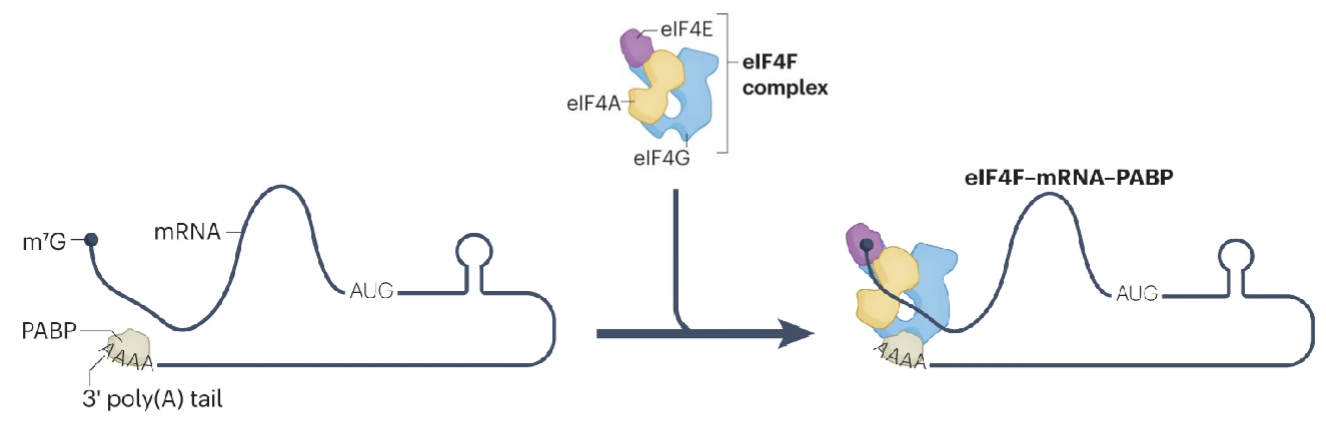
Activation of mRNA by cap binding complex eiF4F is crucial for recruitment
eIF4F and polyadenylate-binding protein (PABP) bind to the poly(A) tail, selecting mRNA for recruitment
What are the steps in 43S recruitment to mRNA?
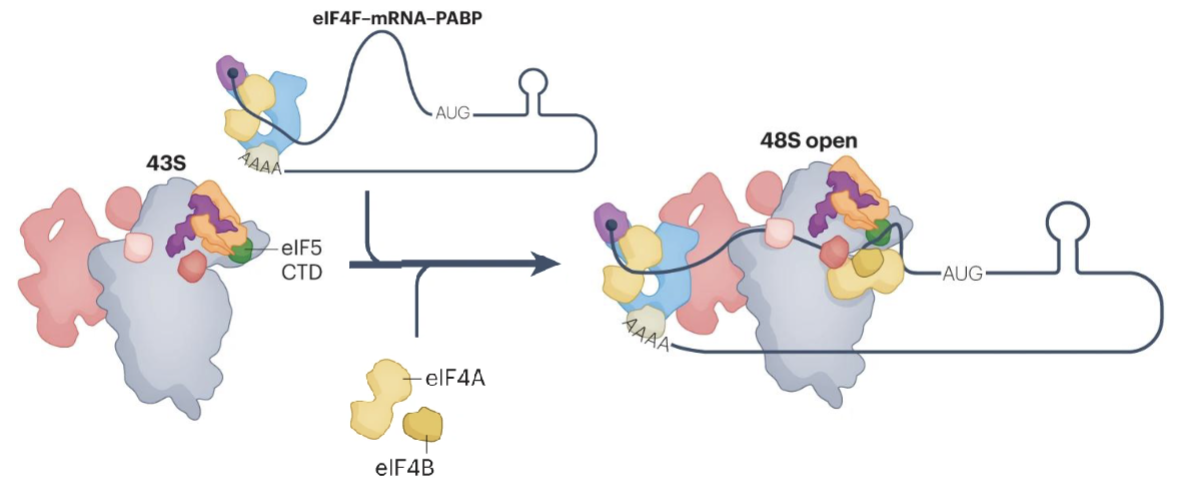
Assembled 43S is recruited to mRNA to form the scanning-competent 48S complex (48S open)
During scanning of 5’ UTR of mRNA, eIF5 interacts with eIF2 and accelerates eIF2-bound GTP hydrolysis
What are the steps for scanning and AUG selection?
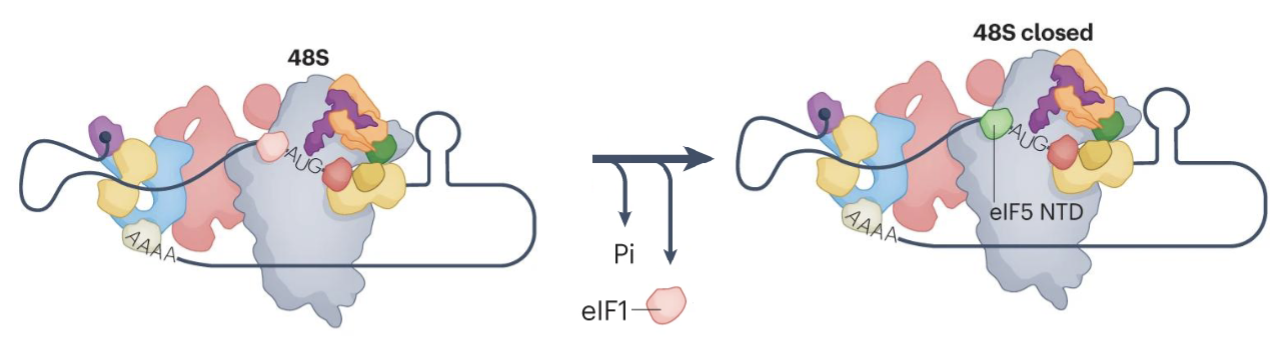
Start codon selection triggers release of eIF1 and Pi from the complex
What steps are in the formation of the 80S initiation complex?
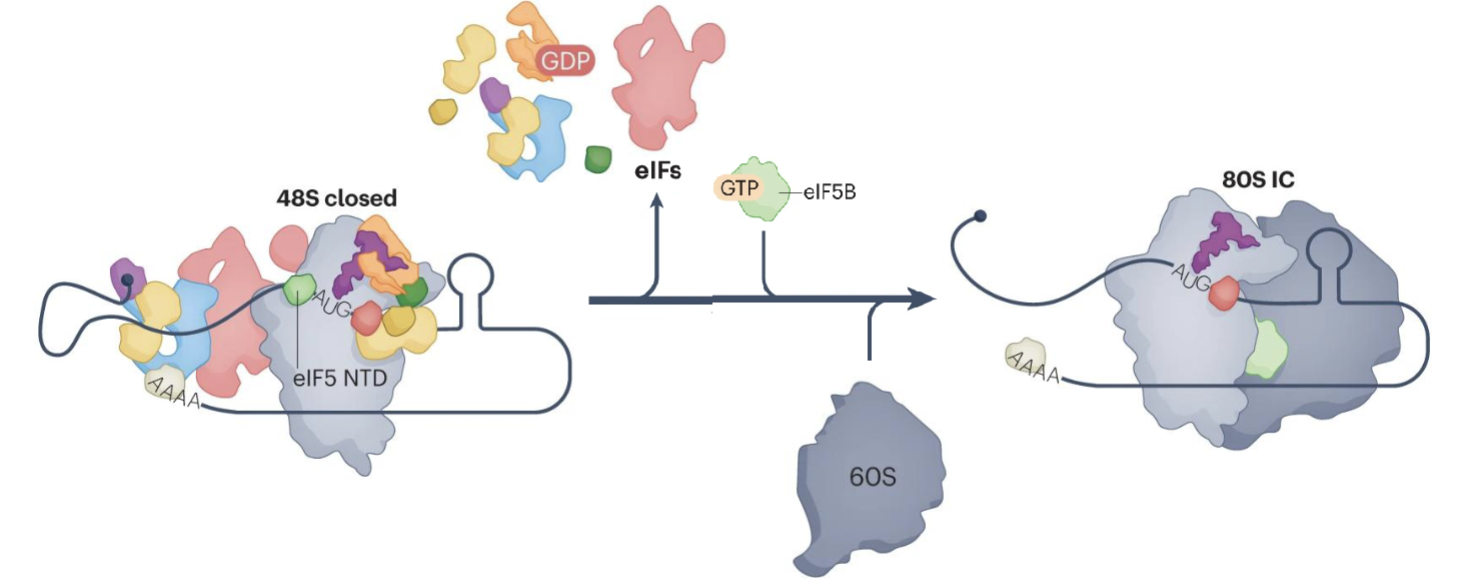
N terminal domain of eIF5 occipies the position vacated by eIF1 near the P site of the ribosome
eIF2-GDP has lower affinity for Met-tRNA so the release of Pi triggers the release of eIF2-GDP, eIF5, eIF3 and eIF4
eIF2-GDP release allows eIF5B to bind, promoting the joining of the 60S large subunit, forming the 80S initiation complex (80S IC)
What steps are of elongation in transaltion?
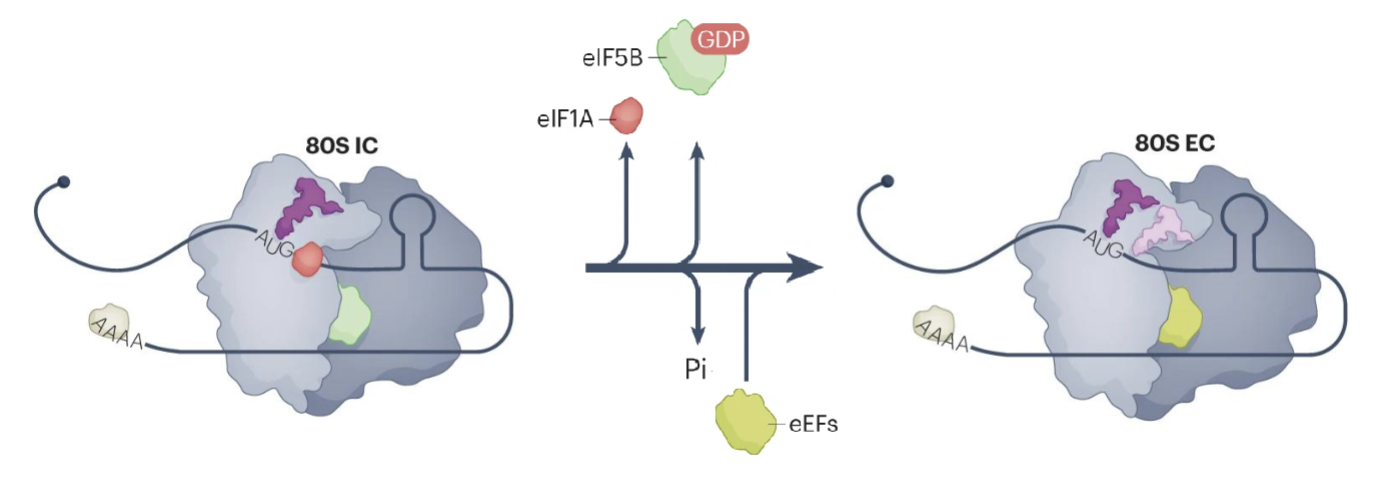
80S IC formation triggers hydrolysis of eIF5B-bound GTP and release of eIF1A
eIF5B undergoes conformational change that places the amino acylated end of Met-tRNA in the peptidyl transfer centre of the ribosome
eIF5B release marks the end of initiation and beginning of elongation (80S EC)
Eukaryotic elongation factor 1A (eEF1A)-GTP delivers amino-acylated tRNA to the A site of the ribosome
eEF2-GTP promotes tRNA translocation from A site to the P and E sites
Release of eEF2-GTP and deacylated tRNA from the E site allows a new elongation cycle to occur
What steps are of translation termination?
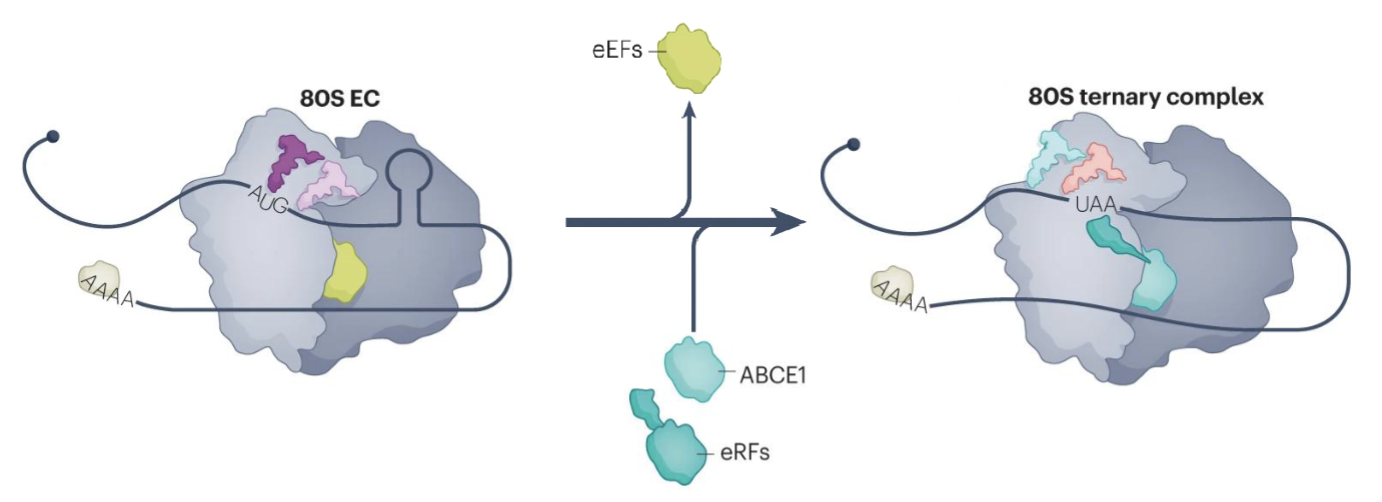
Translation termination by eukaryotic release factors (eRFs) occurs upon encountering a stop codon
ABCE1 binds to 80S termination coplex and stimulates peptidyl-tRNA hydrolysis by eRF1
Describe recycling after translation termination
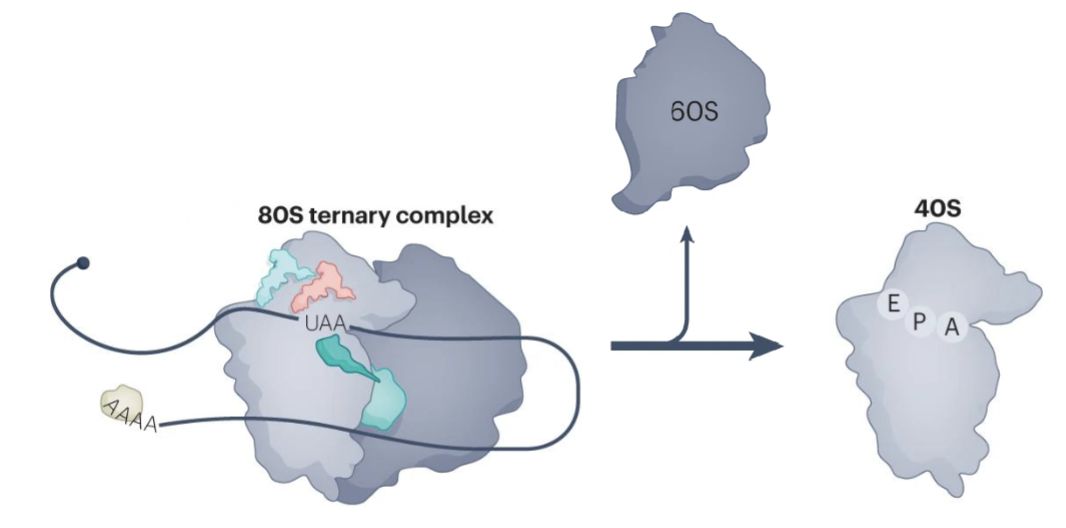
ABCE1 splites 80S into the 40S and 60S subunits
mRNA and tRNA are removed from the 40S by recycling factors, freeing the 40S subunit for a new round of translation
What is the relevance of eIF4F complex formation in cancer?

Ribosomes are recruited to the 5’ end of mRNA via the eIF4F complex, consisting of eiF4E, eIF4G, and eIF4A - these are all targets of Myc and can be dysregulated in cancer, amplified in human tumours
eIF4E and eIF4G are classical oncogenes
These components can also regulate translation initiation, eg. eIF4E phosphorylation by MNK1 and MNK2 can promote tumour development and dissemination
MNK-mediated phosphorylation of eIF4E is also involved in translational reprogramming driving tamoxifen resistance in ER+ breast cancer
eIF4A can be sequestered by tumour suppressor protein PDCD4, preventing formation of eIF4F complex
Loss of PDCD4 associated with cancer cell invasion and poor patient survival
eIF4E-binding proteins (4E-BPs) compete with eIF4G to bind to eIF4E, acting as tumour suppressors by inhibiting cap dependent translation
Loss of 4E-BP expression can also be lost or have impaired function due to inhibitory phosphorylation
4E-BP expression is increased in stage III non-metastatic oesophageal, breast, and prostate cancers, where is is proposed to oppose metastasis but lead to large, locally advanced tumours
What is the relevance of ternary complex formation in cancer?
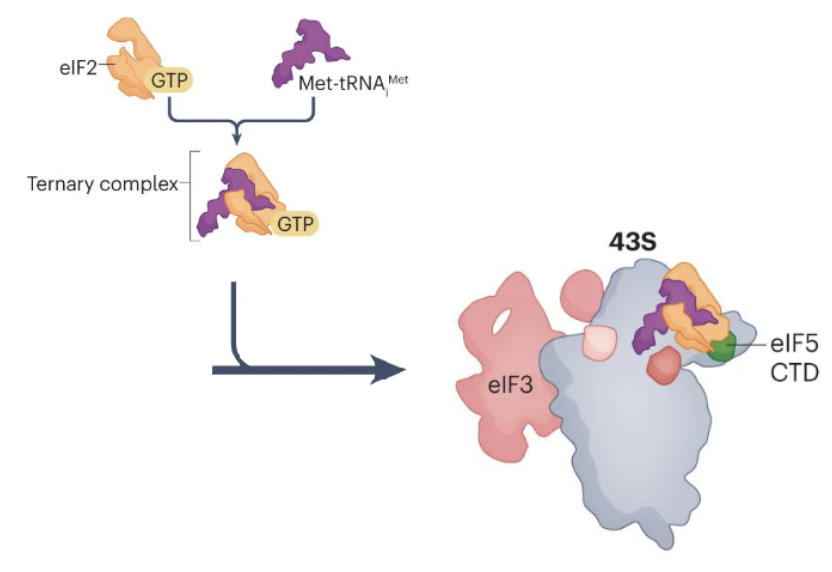
TC is composed of eIF2, GTP, and Met-tRNA
Deregulated TC formation in cancer has caused different findings relating to eIF2α phosphorylation
Generally thought that more eIF2α phosphorylation increases ability to respond to stress by promoting translation of upstream open reading frame (uORF) containing stress response mRNAs
Accordingly overexpression of eIF2α or its kinase PKR has been shown to promote transformation in some contexts, but the mechanism is unclear
In contrast long term eIF2α promotes apoptosis and prompted research on upregulating eIF2α activity or inhibiting eIF2α phosphatases for cancer therapy
Outcome of eIF2α phosphorylation is highly context specific and may be related to site of disease or underlying mutations
Additional ways to modulate TC activity could also include overexpressing eIF5 or its mimic proteins (MPs) 5MP1 and 5MP2 which can bind to and sequester eIF2 from the 40S ribosome when present ine xcess
eIF2 binding by eIF5 or 5MPs reduces global protein synthesis but enhances translation of uROF containing mRNAs which could be important for some cancer properties
What is the relevance of eIF3 connecting eIF4F and PICs in cancer?
eIF3 is a multi-subunit complex that binds to eIF4G, bridging it to the PIC and thereby connecting mRNAs with the 40S subunit for scanning to occur
Increased eIF3 levels should therefore increase rate of translation initiation
What is the relevance of translation elongation/termination in cancer?
Literature has focused on translation initiation but oncogenic changes in elongation and termination is also emerging, eg. dominant role for loss of inhibitory regulation of elongation by eEF2K for intestinal tumour formation
Increased availability of specific tRNA isoaccepting species in cancer cells has a role in tumourigenesis
Speed of amino acid incorporation during elongation is dependent on availability of corresponding charged tRNA
Several studies have reported translation programs in which proliferating undifferentiated cells and cancer cells express tRNAs optimised to correspond to codon usage of pro-proliferative mRNAs
Elongation can be deregulated in cancer via programmed -1 ribosomal frameshifting (-1 RPF), a process by which sequence elements force elongating ribosomes back by one base, leading to frameshift, premature stop codons, and nonsense-mediated mRNA decay (NMD)
May explain the oncogenic role of silent mutations inducing frameshifting in tumour suppressors
Termination at premature stop codons can be a cancer driver if it occurs as a result of somatic mutations in tumour suppressor genes, resulting in NMD of the corresponding transcripts
eIF6 initiation factor has confusing multiple roles in mRNA translation and associated with altered translational regulation in cancer
Ribosomal subunit anti-association factor that prevents aberrant interactions between the 40S and 60S ribosomal subinits
eIF6 must be displaced from the ribosome for the final step of 60S ribosome biosynthesis in the nucleolus and can promote 80S ribosome disassemby in cytosol by preventing the reassociation of post-termination 60S ribosomes
Aberrant expression observed in some cancers in which it accumulates in the nucleolus