Michaelmas building a phenotype
1/112
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai | Chat |
|---|
No analytics yet
Send a link to your students to track their progress
113 Terms
what are the psi and phi angles?
the psi angle is formed between the alpha carbon and the carbonyl carbon in a residue
the phi angle is formed between the alpha carbon and the nitrogen in a residue
these angles are found in combinations that minimise the steric hindrance between the atoms, and dictate the rotation on either side of the peptide bond within the amino acid backbone
this determines which secondary structures can form according to the ramachandran plot
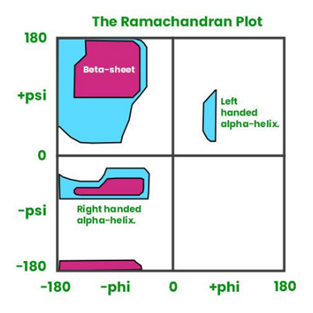
describe alpha helices
hydrogen bonds form between carboxyl and amino grups within the backbone, each residue connected to the fourth residue following it in the chain
3.6 residues per turn
12 residues long on average
side chains project outwards from the main chain in a helical array
large side chains and charged side chains aren’t tolerated due to steric hindrance
describe beta sheets
2-22 neighbouring polypeptide chains
chains are antiparallel or parallel- antiparallel are more energetically stable because the H bonds are at 180 degrees and are regular lengths
side chains project perpendicularly outwards, alternating sides of the plane
what is needed to carry out PCR?
needed to amplify starting matter + add restriction sites in gene modification
the target DNA is added to a mixture containing:
an excess of deoxyribonucleoside triphosphates (dNTPs) for all four bases
a heat-stable DNA polymerase- this can be reused for many cycles without needing to be replaced (taq is the textbook one, but not often used because it can’t proofread)
DNA primers for the sequences flanking the target sequence, typically 20-30 nucleotides long- for genetic modification these are normally edited to include restriction sequences for the endonucleases used
a buffer
magnesium (the cofactor for DNA polymerase)
what is the process of PCR?
heated to 95 degrees to break the H bonds (denature) between the strands
cooled to 50-60 degrees (dependent on length + melting point of the primer- higher proportion of G+C = more H bonds, so higher) to anneal the primers to each strand
heated to 72 degrees (optimum for taq polymerase) to elongate the chain using complementary dNTPs
why does DNA take a helix form?
this allows pi-pi interactions to form between the bases within a strand, which are highly stabilising
also, rotating the bases in the helix offsets the relative electron positions in the rings
the nitrogens in the rings repel delocalised electrons more because they have a lone pair- so the delocalised electrons are more likely to be found in the same positions in adjacent bases, which would repel each other
also due to stabilisation by H bonds, van der Waals and the hydrophobic effect- helix allows water to be most excluded
describe the different DNA helices
B-DNA (hydrated form):
right-handed coil
~10.4 nucleotides per turn
each turn has a length of 34 Å
most compact
A-DNA (dehydrated form):
right-handed coil
bases are more tilted
11 nucleotides per turn
Z-DNA (temporary, more common with C and G repeats, often formed under torsional stress):
left-handed helix
12 bases per turn
the phosphate backbone zigzags
what is the acid in DNA/RNA? why are they called bases?
acid- the phosphate groups have two hydroxyl groups that can easily donate hydrogens
bases- the nitrogens in the bases can accept hydrogens
where do the RNA components come from?
phosphate is a common mineral
ribose comes from formaldehyde (H2C=O)
bases come from cyanide (HC=N)
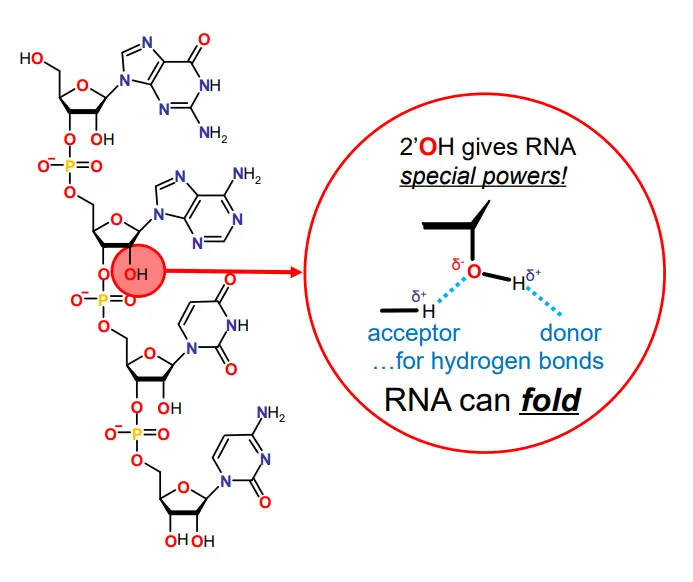
why can RNA fold?
the 2’ OH group (which is missing in DNA) allows hydrogen bonds to form, both as an acceptor and a donor
this allows a tertiary structure to form
(the primary structure is the base sequence and the secondary structure is the base pairing )
why is RNA a valid theoretical starting molecule for life?
it can:
store information- base sequence
evolve- mutations + selection
sense the environment- some RNA molecules called riboswitches (eg. glutamine riboswitch) can bind to small molecules to detect changes in pH, temp, ions, salts, etc
catalyse reactions- ribozymes are RNA enzymes
probably replicate itself (but no real RNA replicase has been created yet)
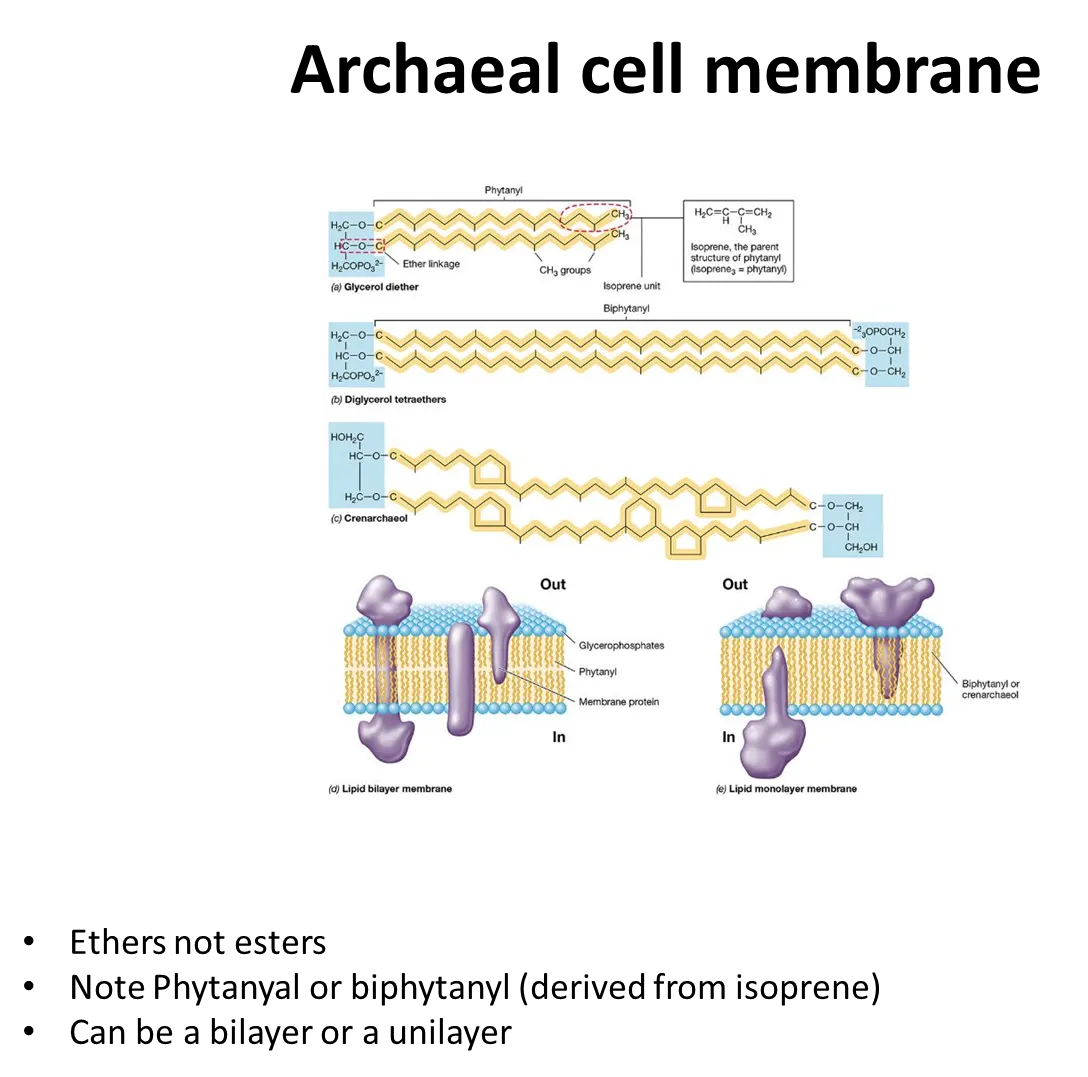
how are cell membranes in archaea unique?
phospholipids in archaea are joined by ether bonds, not ester bonds
diglycerols can be formed, so they can form monolayers as well as bilayers
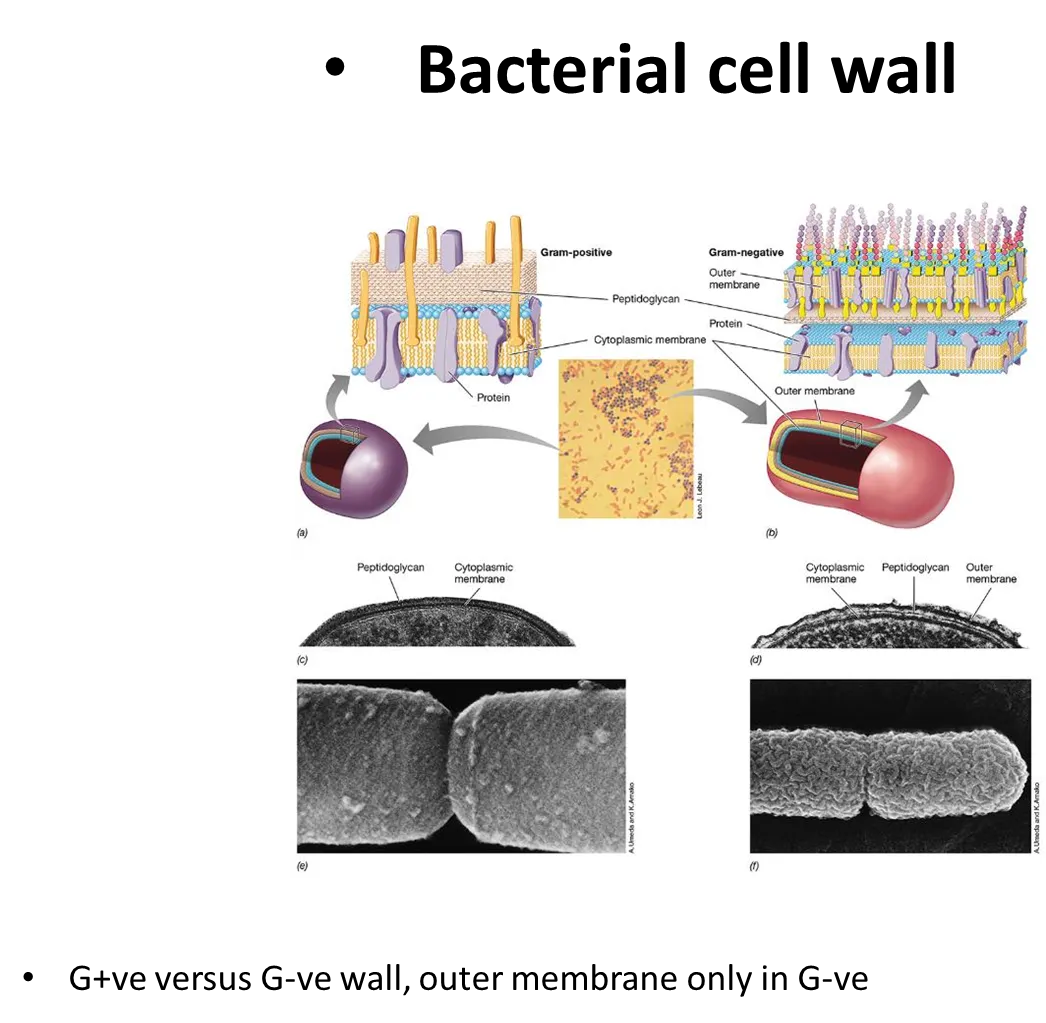
how are the structures of G+ve and G-ve bacterial cell walls different?
G+ve bacteria have a cytoplasmic membrane and a large layer of peptidoglycan over it
G-ve bacteria have a thin layer of peptidoglycan around the cytoplasmic membrane, followed by a second outer unit membrane (so they don’t take up gram stain), with large carbohydrate chains sticking off- inbetween the two membranes is the periplasm
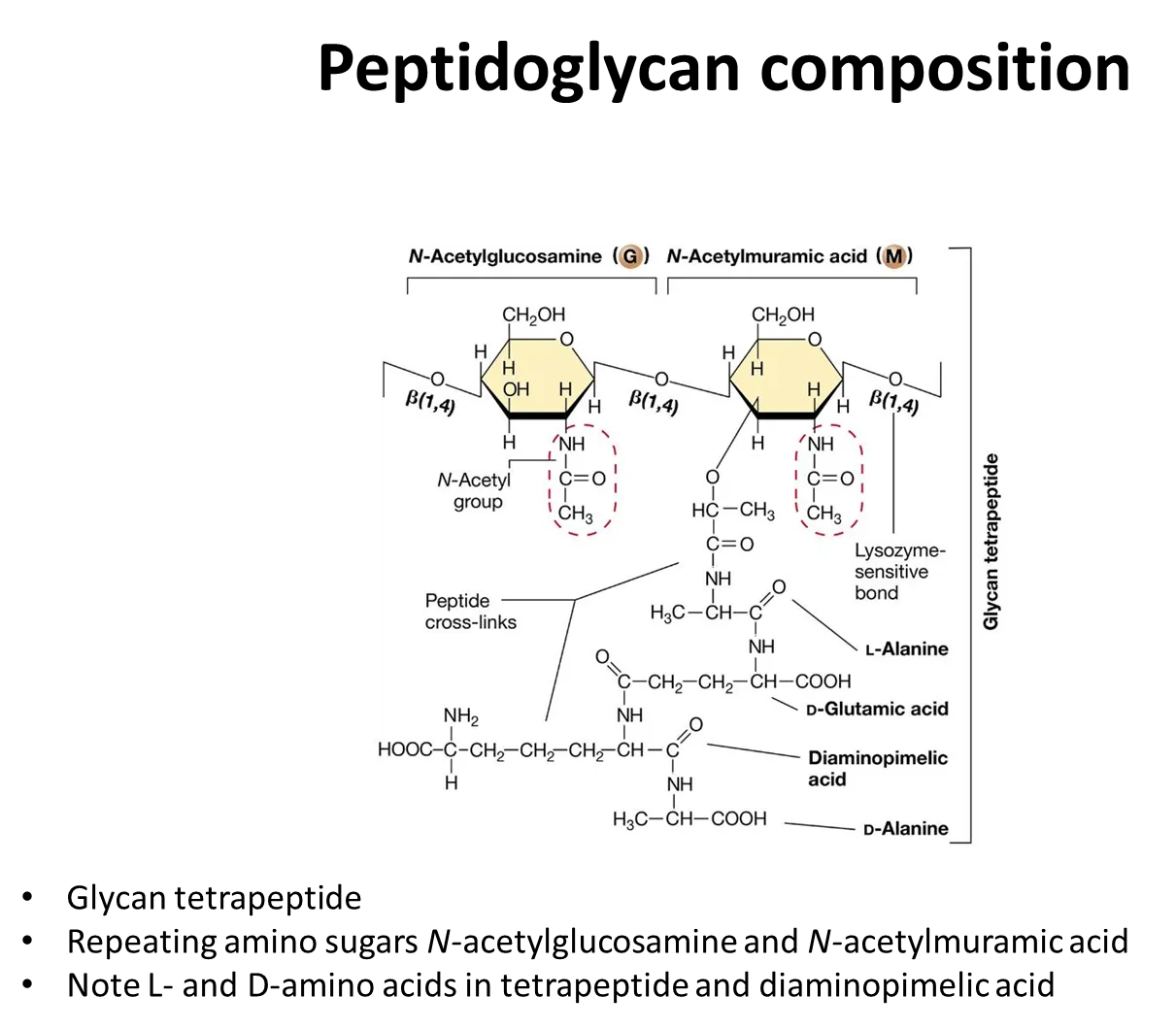
what is the structure of peptidoglycan?
the sugar backbone is composed of alternating monomers of N-acetylglucosamine, NAG, and N-acetylmuramic acid, NAM, (modified glucose residues) joined by a beta-1,4 linkage
a short peptide side chain is attached to N-acetylmuramic acid- the amino acids vary between species
strands of peptidoglycan cross-link by peptide bonds between the amino acids in these peptide side chains
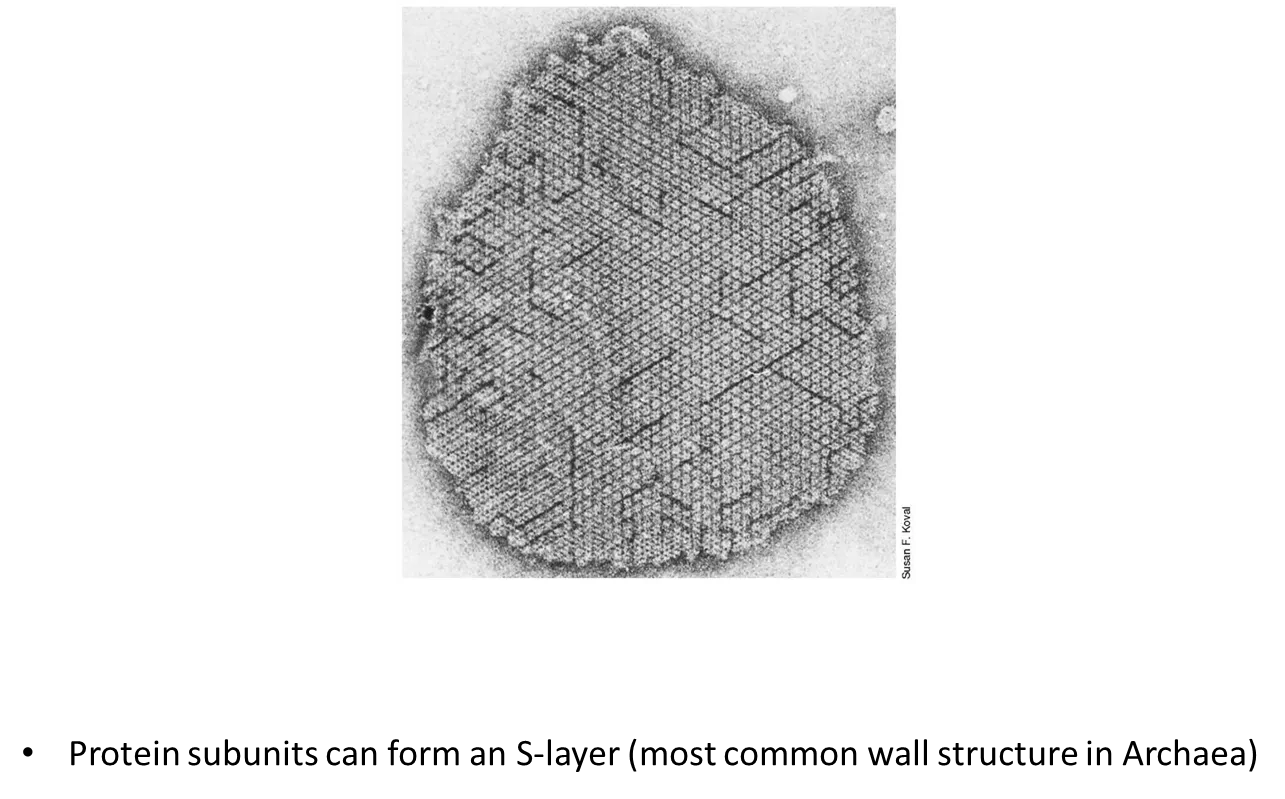
how are archaeal cell walls different to bacterial cell walls?
archaea don’t have peptidoglycan in their cell walls
they don’t have an outer membrane like G-ve bacteria
protein subunits can form an S-layer
the structure of the cell walls varies a lot between archaeal species
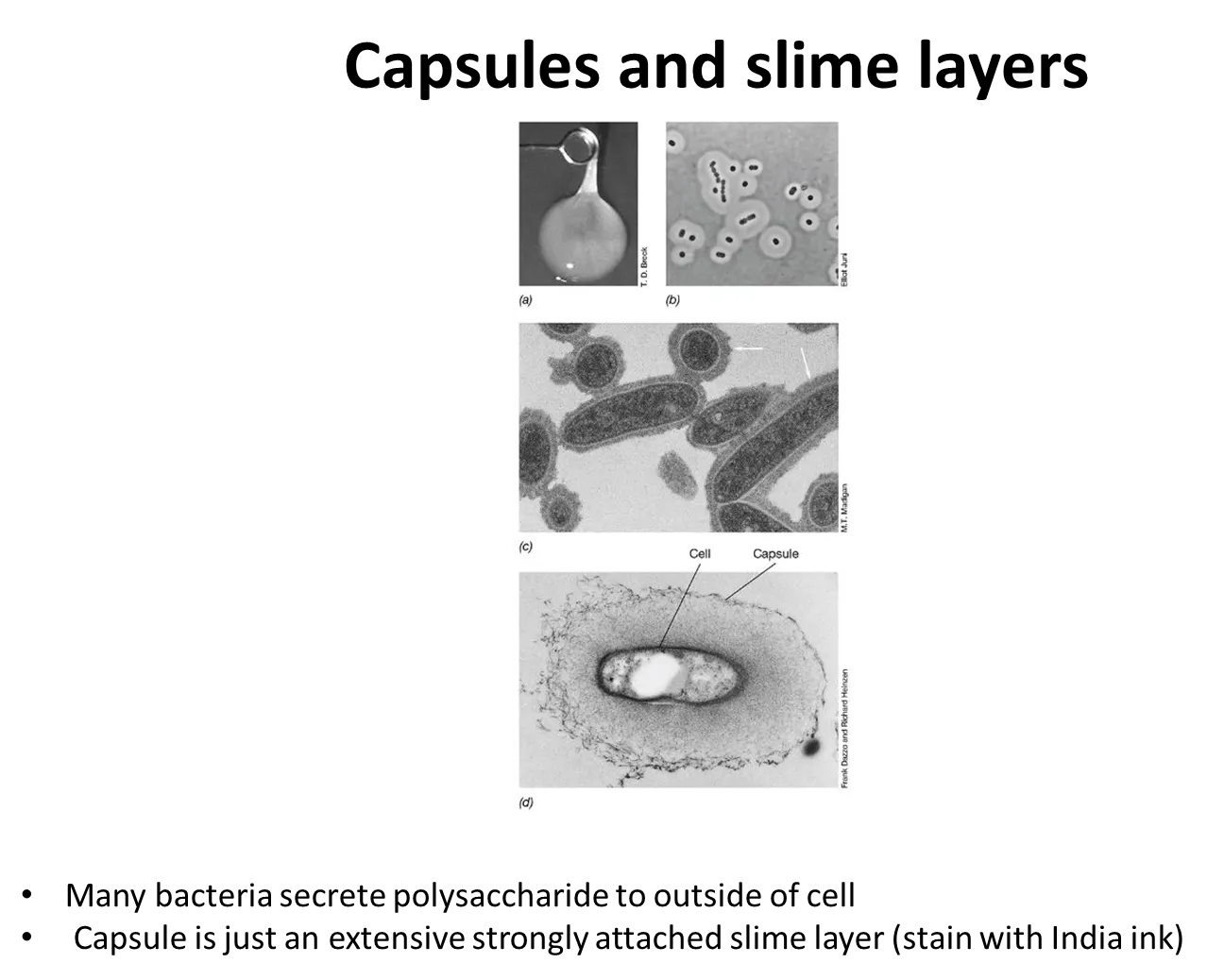
what are capsules and slime layers?
many prokaryotes secrete sticky/slimy polysaccharides on their cell surface, outside the cell envelope:
a capsule is a tight matrix that excludes small particles
a slime layer is easily deformed and loosely attached, and doesn’t exclude particles
these mediate attachment, protect the cell from attack and environmental stresses eg. dehydration
they are often used by nitrogen-fixing bacteria to exclude oxygen, because nitrogenase is destroyed by oxygen
what are pili used for?
sticking to surfaces
sticking to other cells (helpful for pathogens/conjugation)
forming thin sheets of cells: biofilms (on solid surfaces) and pellicles (on liquids)
movement via twitching motility (by attaching to a surface then retracting)
all G-ve have pili, some G+ve do too
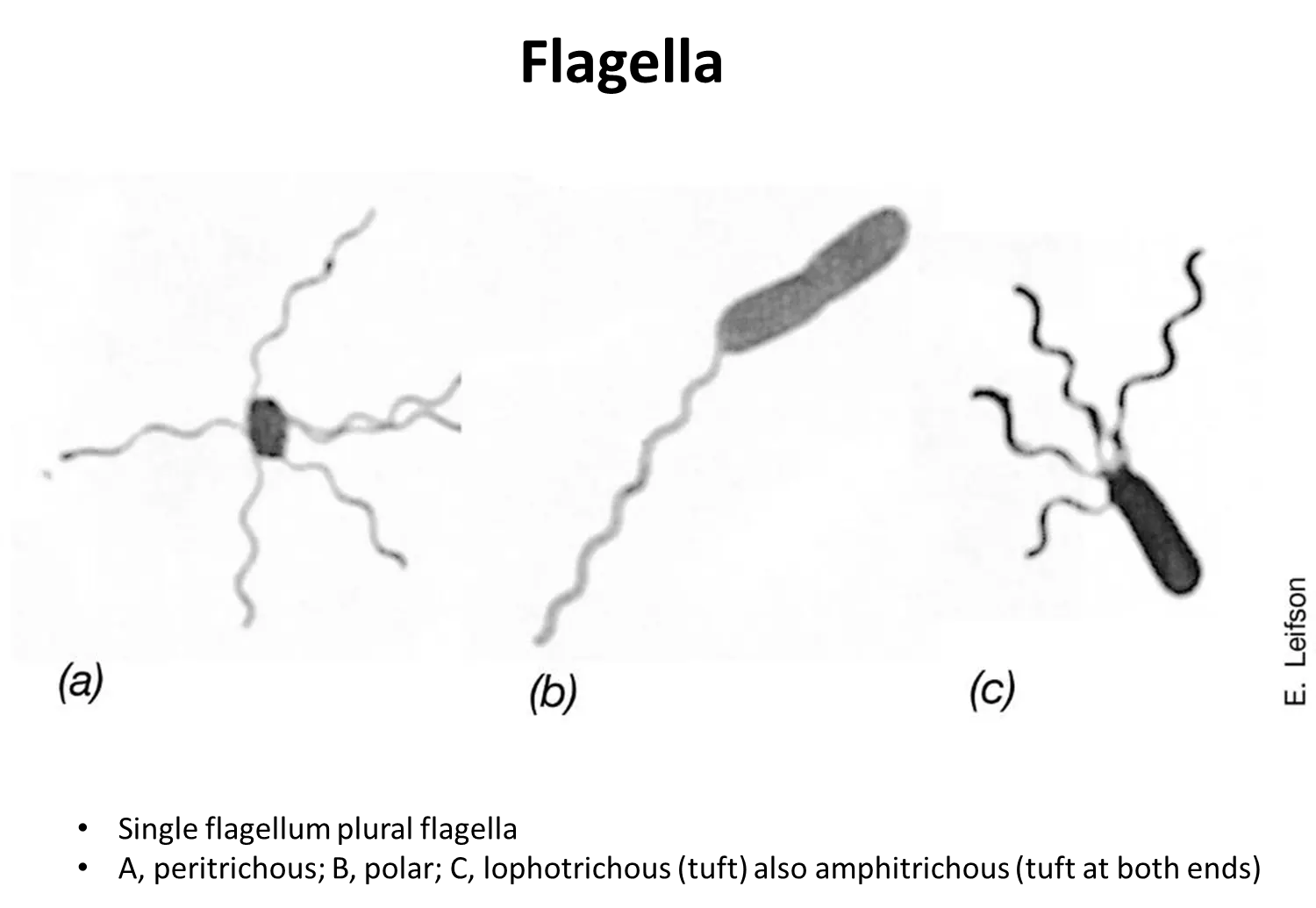
what are the four distributions of flagella?
peritrichous - scattered around the cell surface
polar- one flagella at one end
lophotrichous- bundle at one end
amphitricous- bundles at both ends
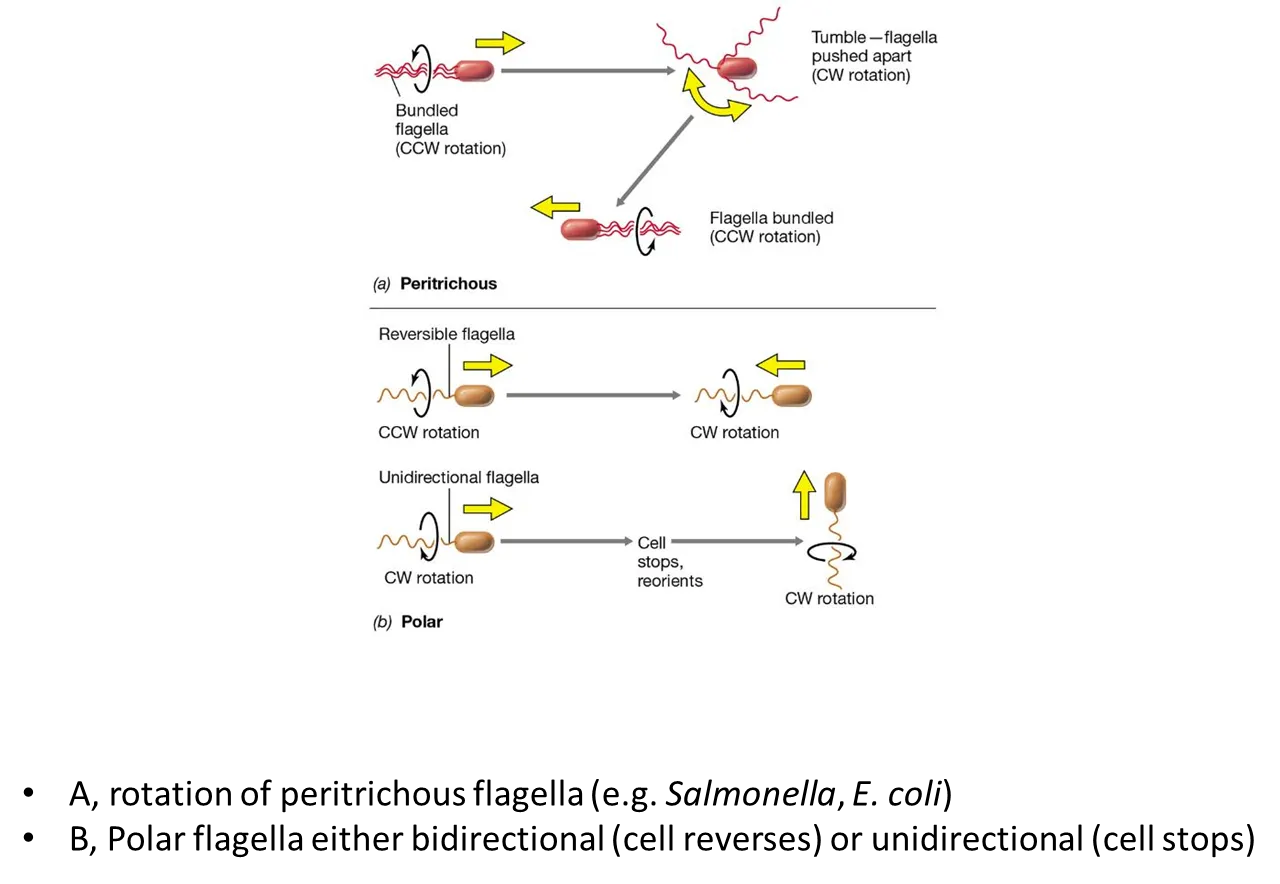
how do bacteria move using flagella?
in peritrichous bacteria:
all the flagella bundle together and rotate counterclockwise
to turn, one flagellum turns clockwise, causing a ‘tumble’ where it will change direction
then the flagella bundle back up and rotate counterclockwise in the new direction
in polar bacteria:
some cells can reverse by turning their flagella counterclockwise or clockwise (reversible flagella)
some cell have to stop and reorient to turn (unidirectional flagella)
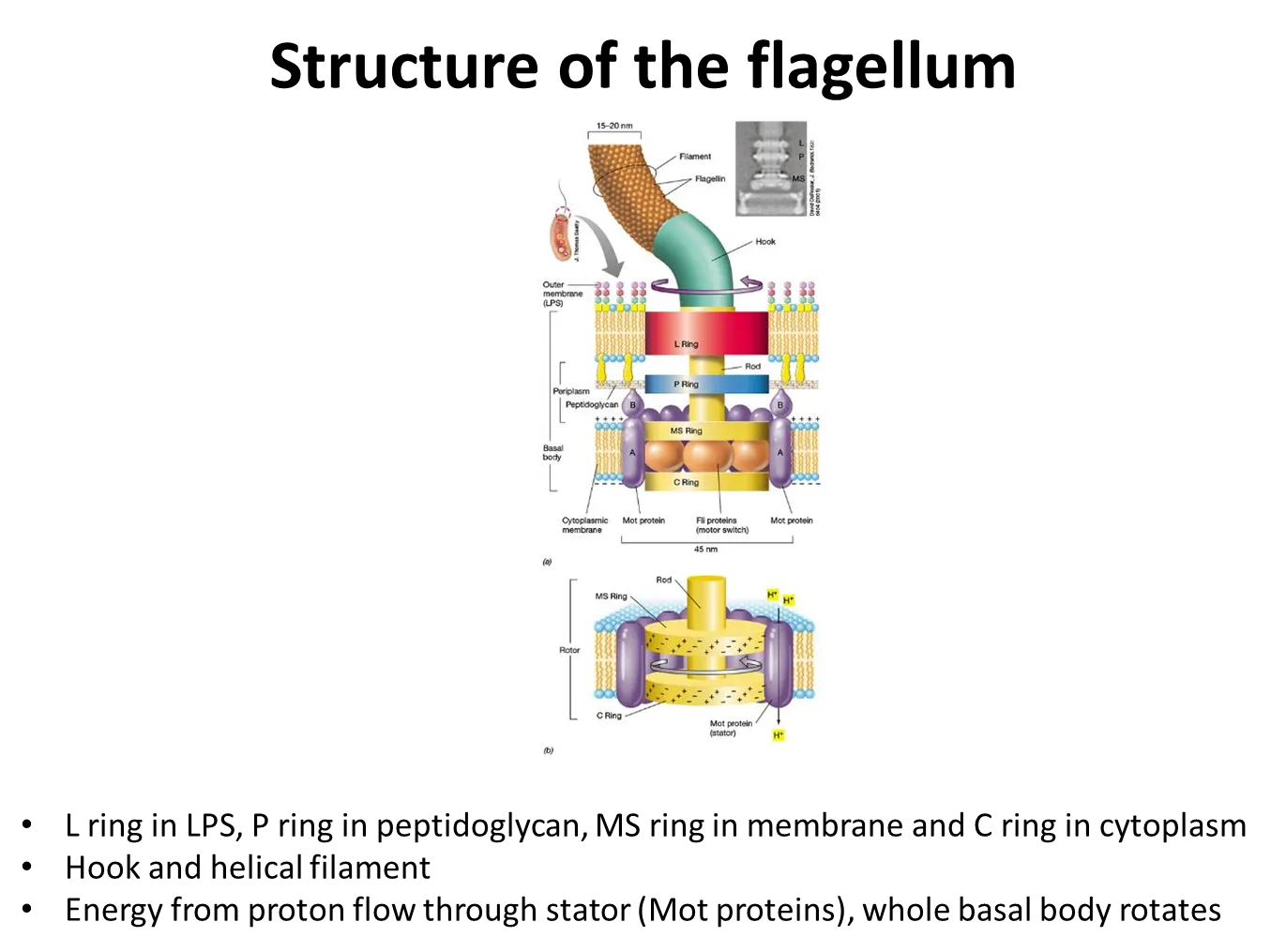
how do flagella rotate?
bacterial flagella are rigid and helical
the basal body is anchored in the cytoplasmic membrane
this contains a proton motor- proton movement through channels and electrostatic forces cause the rotation of the proteins
they change their speed in relation to the strength of the proton motive force
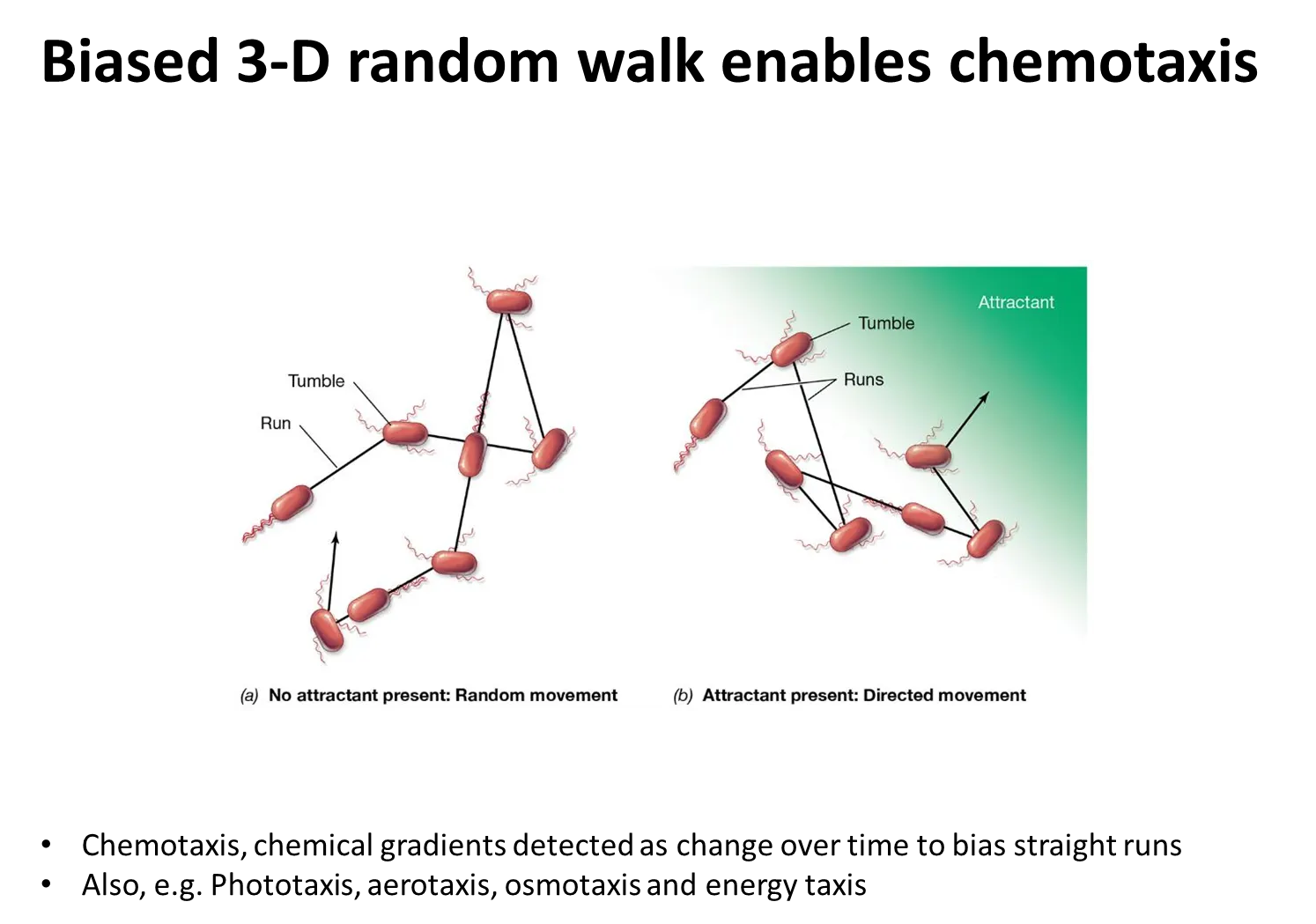
how do flagella enable chemotaxis?
positive chemotaxis towards an attractant (eg. sugar):
if the bacteria are swimming away from the attractant, the frequency of tumbling increases so that the bacteria turn towards it, and the frequency of tumbling then decreases
the opposite occurs for negative chemotaxis away from a repellent (eg. toxin)
this is called the biased 3D random walk
what is the most common structure of gene regulators?
regulators are usually dimers (two very similar protein monomers)
the helix-turn-helix motif is a very common motif
these form two domains: a stabilising helix and a recognition helix
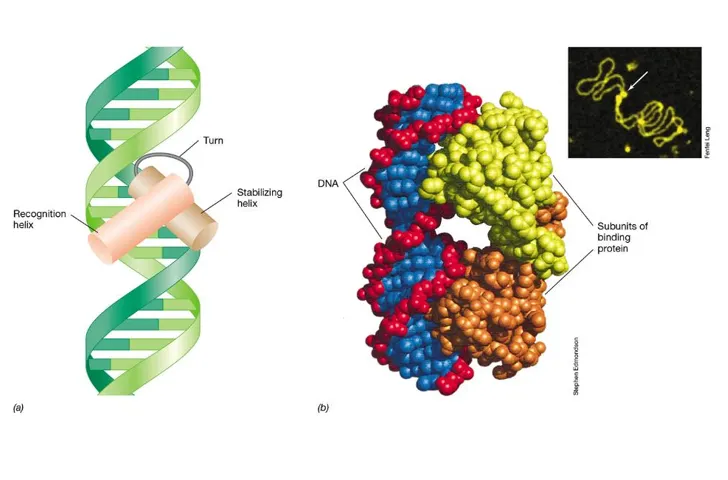
what are the three types of gene regulation in prokaryotes?
negative regulation always has a repressor which binds to the operatore (downstream of the promoter) and stops RNA polymerase:
negative regulation resulting in repression eg. lac operon
a corepressor binds to the repressor protein and causes it to attach to the operator so that RNA polymerase can’t transcribe DNA
negative regulation resulting in induction eg. arginine biosynthesis
an inducer binds to the repressor and causes it to detach from the operator, so that RNA polymerase can transcribe DNA
positive regulation always has an activator which binds to the activator binding site (upstream of the promoter) and permits RNA polymerase:
positive regulation resulting in activation eg. maltose uptake operon
an inducer binds to the activator protein which attaches to the activator binding site and allows RNA polymerase to attach to the promoter so that it can transcribe DNA
the binding site can be quite far upstream of the promoter, so the dna needs to bend

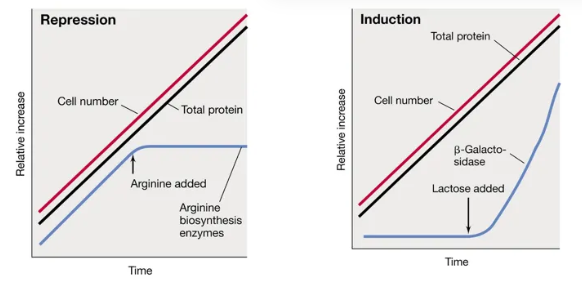
what is the difference between the two types of negative gene expression in prokaryotes and when are they used most? include an example of each and the graphs
negative gene regulation always involves a repressor protein
anabolic genes (biosynthetic) are typically subject to repression by the end-product
the repressor protein only attaches when the end-product (the corepressor) is present
when arginine is present, the repressor protein attaches and arginine biosynthesis enzymes stop working
catabolic genes (degradative) are typically induced by a substrate
the repressor protein only detaches when the substrate (the inducer) is present
when lactose is present, the repressor protein detaches and the lactase enzymes start working
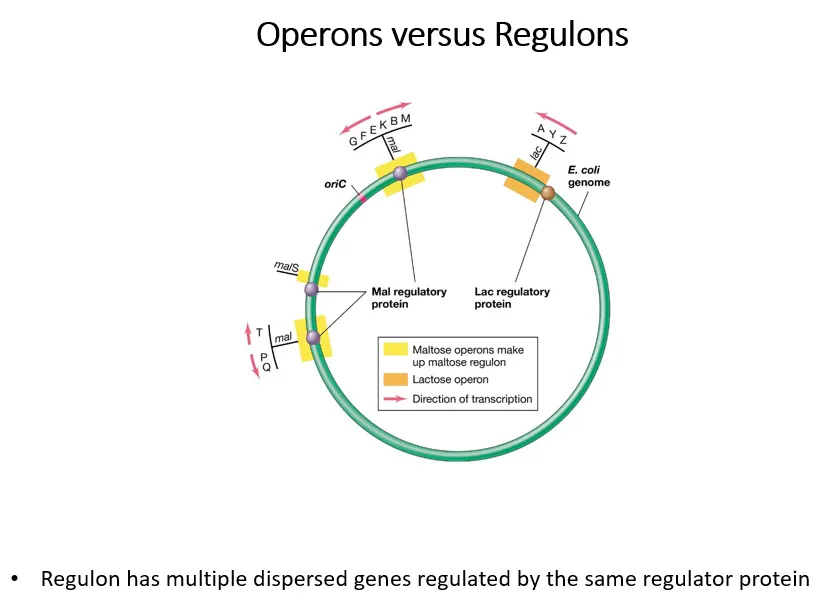
what are regulons and operons?
operons are a group of genes controlled by a regulator protein
regulons are multiple groups of operons controlled by the same regulator protein
the yellow operons are part of the maltose regulon (all controlled by the mal regulatory protein), the orange operon isn’t part of a wider lac regulon
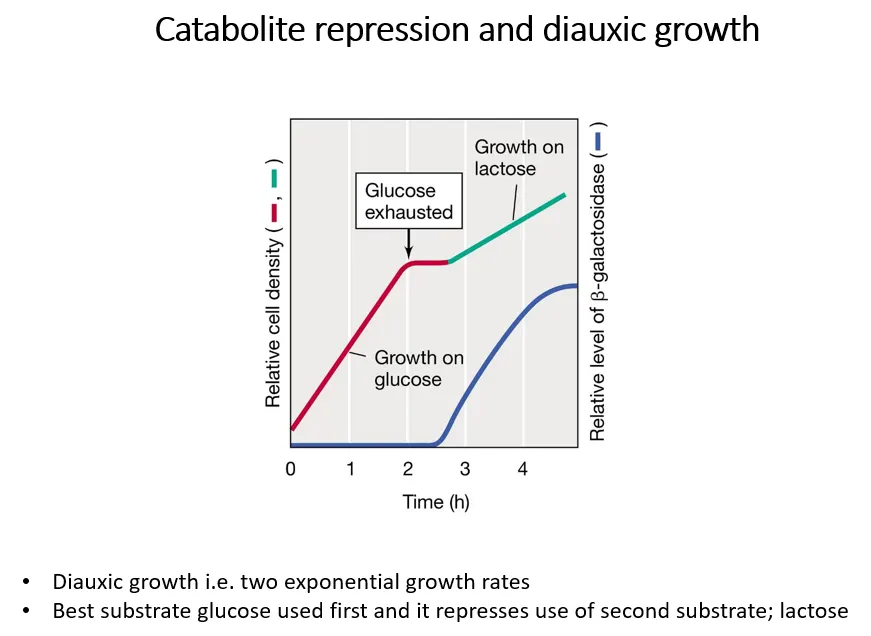
what is diauxic growth?
diauxic growth is shown when two metabolic substrates are present, eg. both glucose and lactose
glucose is a better substrate, so bacteria grow on it preferentially at first, then switch to lactose when it gets used up
if glucose is present, the lactase enzymes aren’t transcribed
when glucose runs out, bacterial growth stops while the lac operon is being induced to produce the lactase enzymes
this means the lac operon is under two levels of regulation:
positive regulation dependent on glucose concentrations
negative regulation dependent on lactose concentrations
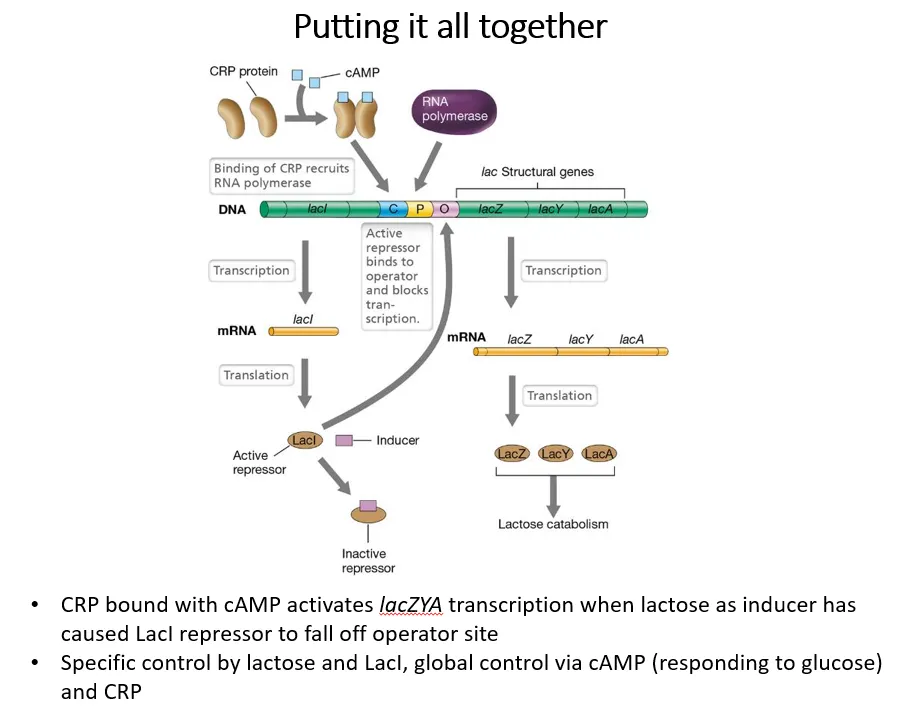
how do glucose and lactose concentrations regulate the lac operon?
positive regulation dependent on glucose:
adenylate cyclase converts ATP into cyclic AMP
cAMP acts as an inducer, binding to CRP (cAMP receptor protein), which is an activator for the lac operon
glucose inhibits adenylate cyclase, so when glucose is present, little cAMP is produced
this means less CRP can bind to the activator binding site, so RNA polymerase can’t bind
negative regulation dependent on lactose:
lactose acts as an inducer for the LacI repressor protein
when lactose is present, the repressor detaches from the operator region, so RNA polymerase can bind
this means transcription is only possible if CRP is present, due to the absence of glucose, and if the LacI repressor is inactivated, due to the presence of lactose
then, LacZ (b-galactosidase), LacY (lactose permease) and Lac A (galactoside acetyltransferase) can be produces
how do bacteria regulate gene expression based on the environmental signals?
bacteria have many sensors, autokinases that tend to be membrane proteins
these can recognise different environmental signals and phosphorylate themselves using ATP
they can then transfer this phosphate onto a response regulator, which is typically a transcription factor
the phosphate acts as an activator, corepressor or inducer on different genes
this is called a two-component system

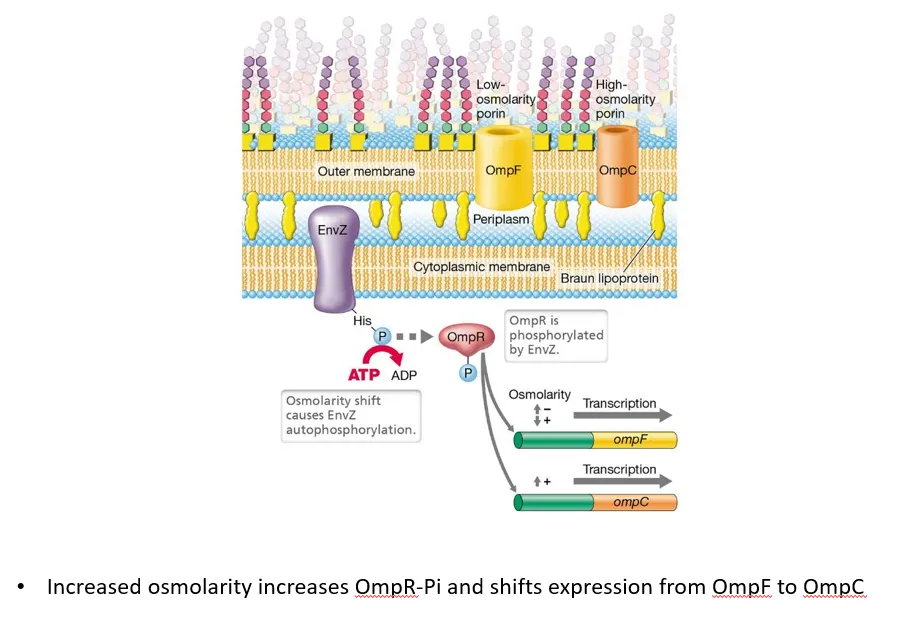
what is an example of a two-component system in G-ve bacteria?
the sensor envZ, a transmembrane protein in the cytoplasmic membrane of G-ve bacteria, detects changes in the osmotic potential of the periplasm
when the osmolarity shifts, it autophosphorylates, then transfers the phosphate to the ompR regulator protein
low osmolarity (high solute concentration outside the cell) causes ompR to induce ompF (large protein channel) production and repress ompC (small protein channel), so more solutes diffuse in
high osmolarity (low solute concentration outside the cell) causes ompR to repress ompF production and induce ompC, so more water diffuses in
this helps achieve the correct osmotic pressure inside the cell, to maintain turgor
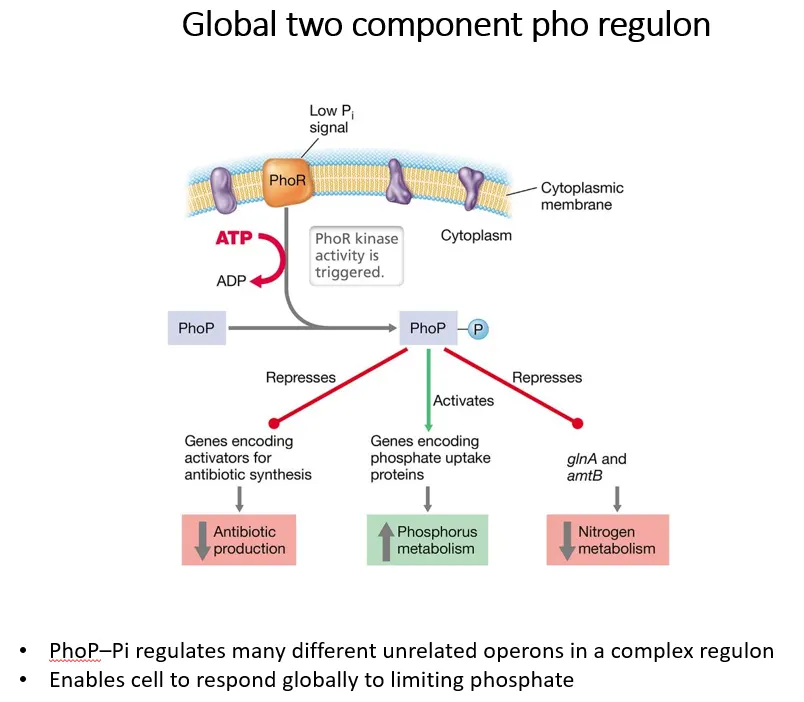
what is an example of a two-component regulon in G+ve bacteria?
the sensor PhoR in the cytoplasmic membrane in G+ve bacteria detects low phosphate in the environment and autophosphorylates using ATP
it then transfers this phosphate to PhoP, which regulates many different operons in a complex regulon to respond to low phosphate
what is an example of transcriptional control in archaea?
amino acids are normally produced by the transamination of oxaloacetate by glutamate which is converted into alpha-ketoglutarate
if nitrogen is limiting, glutamate isn’t reformed from a-ketoglutarate
so if a-ketoglutarate is high, nitrogen-acquiring genes must be turned on
a-ketoglutarate is an inducer, which binds to the nrpR (nitrogen regulatory protein repressor) protein and causes it to drop off the TATA and BRE boxes so TBP (TATA binding protein) and TFB (transcription factor B) activators can bind, allowing RNA polymerase to attach and transcribe the DNA

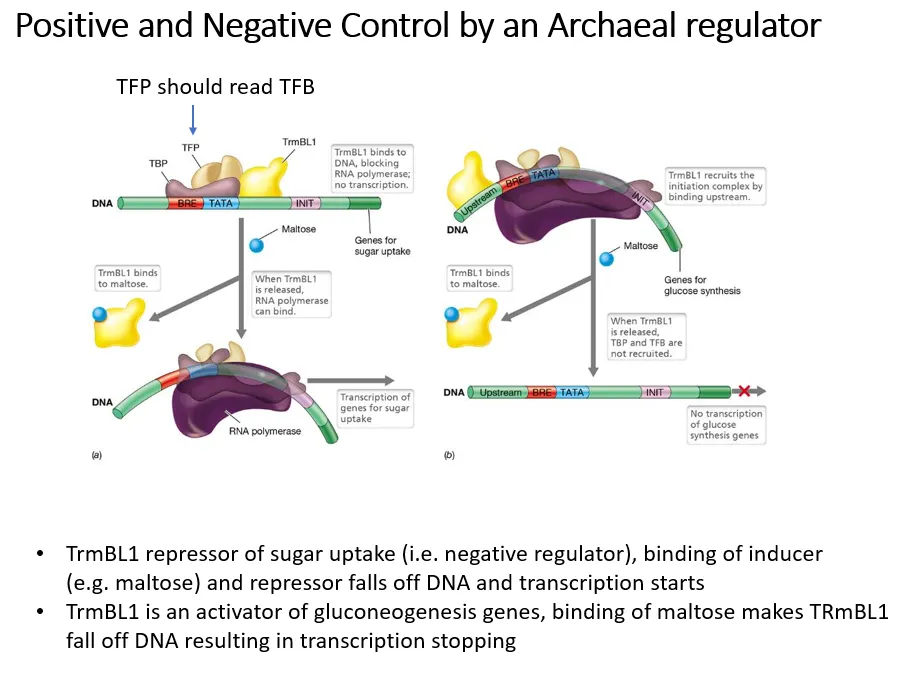
what is an example of a regulator that can both positively and negatively control genes in archaea?
negative control by induction:
trmBL1 acts as a repressor of sugar uptake genes
when sugars are present, maltose acts as an inducer and binds to trmBL1 so that it drops off
this means transport systems to take up the sugars can be transcribed
positive control:
trmBL1 also acts as an activator of gluconeogenesis genes
when sugars are present, maltose binds to trmBL1 and causes it to drop off
this means the gluconeogenesis genes can’t be transcribed when there are sugars already present
explain what sigma factors are
sigma factors are cofactors for RNA polymerase in prokaryotes- they recognise and bind to the promoter sequence, then recruit RNA polymerase to begin transcription (and form a holoenzyme together), then drop off
the main sigma factor is sigma 70, which is produced from the rpoD gene (the housekeeping gene)
this binds to pribnow boxes- the more similar the sequence is to the TATA box, the more often sigma 70 will bind and the more the gene will be expressed
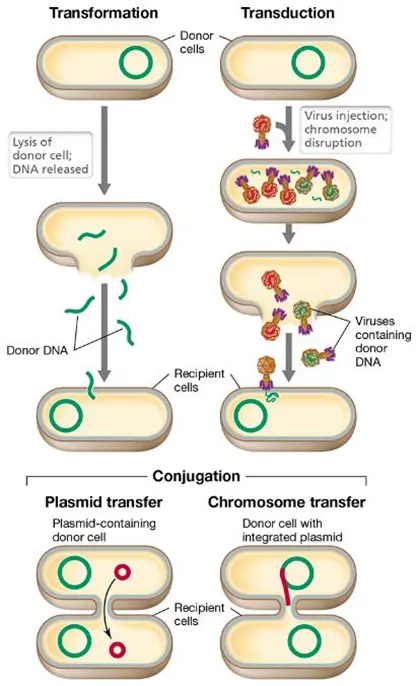
what are the 3 main ways bacteria acquire dna?
transformation- a single strand of DNA is taken up after a donor cell lyses
transduction- a virus called a bacteriophage transfers DNA
conjugation- bacteria form a mating bridge to transfer plasmids (and sometimes chromosomes)
why does foreign DNA need to be recombined into the host DNA in HGT?
so that it isn’t recognised as foreign and broken down:
restriction enzymes in bacteria degrade specific restriction sequences in DNA
when these sequences occur in host DNA they get methylated by the bacteria so that they aren’t broken down
foreign DNA isn’t correctly methylated, so it has to be recombined to not get attacked
this is with the exception of if the DNA introduced is a self-replicating plasmid
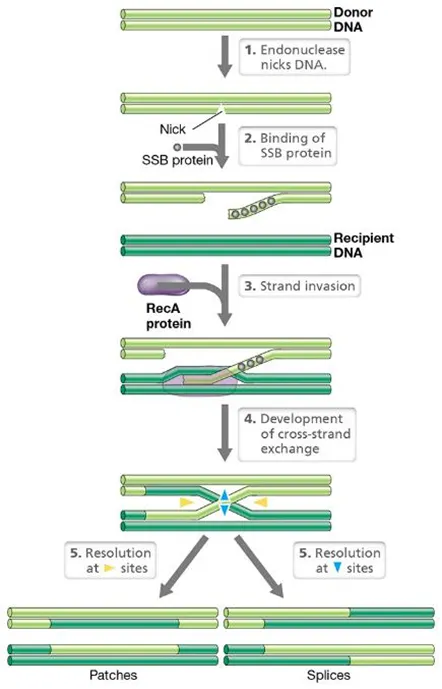
how can foreign DNA be recombined?
DNA gets nicked at specific sites by endonucleases
the single stranded binding protein (SSB) binds to the start of the nicked sequence
recA, the main recombinating protein, triggers a cross-strand exchange
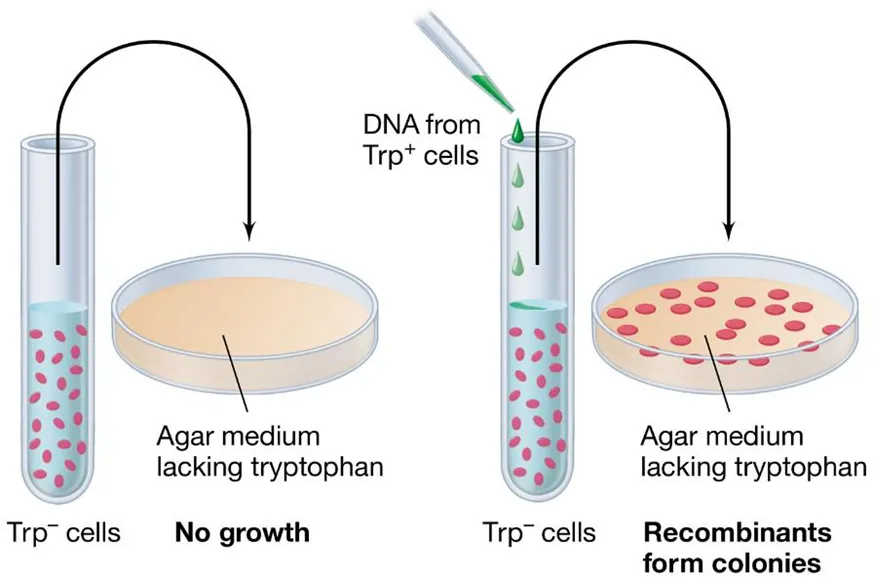
how can we measure DNA uptake after recombination?
we need a selective marker to see which recombinations were successful eg. antibiotic resistance or essential compounds like the amino acid tryptophan
auxotrophs are bacterial strains which cannot synthesise a particular compound
the wild type, which can produce the compound, is called a prototroph
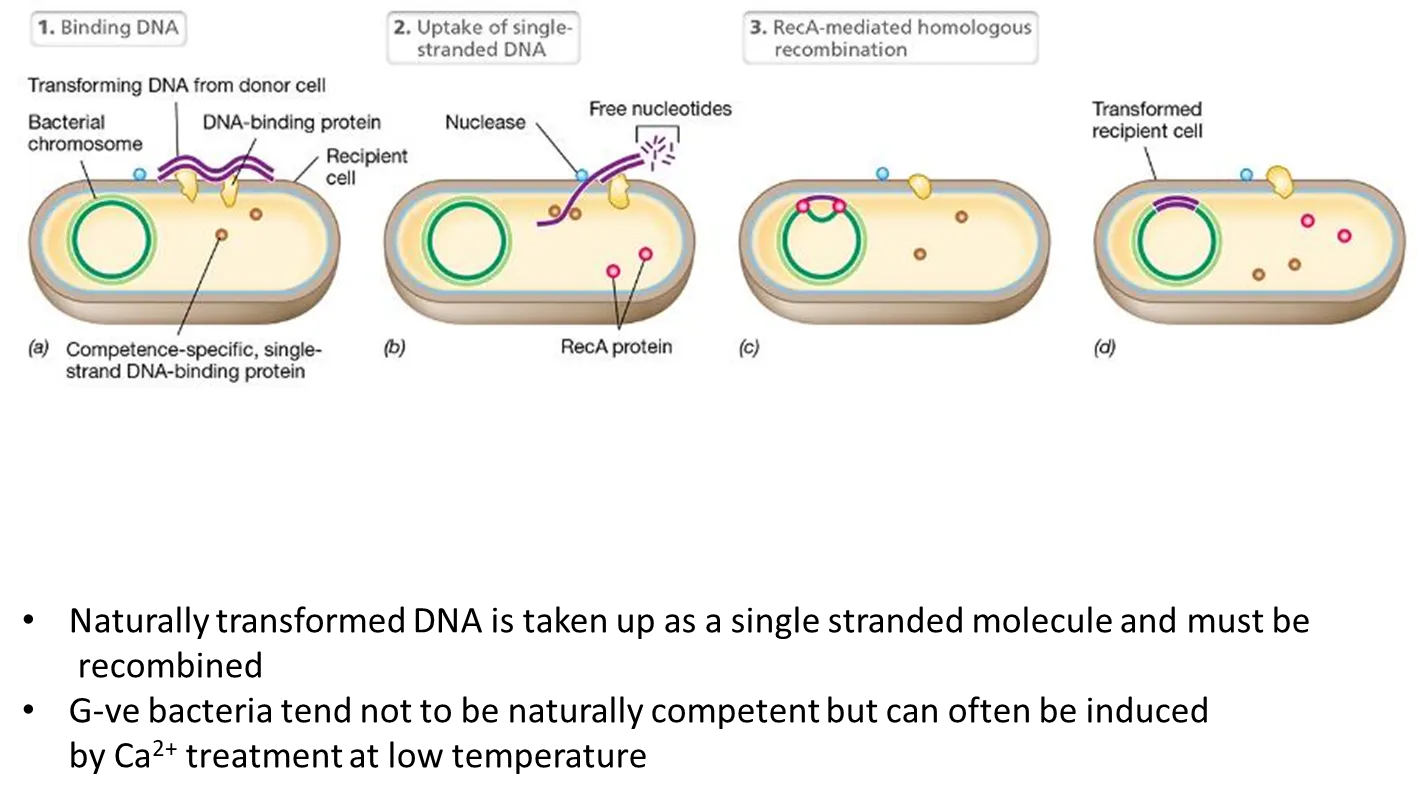
what is the process of natural transformation?
DNA from the donor cell is taken up as a single-stranded molecule (cut by single-stranded DNA binding proteins) and must be recombined by recA
this happens most in G+ve bacteria
G-ve bacteria can be artificially induced using calcium ion treatment (normally using self-replicating plasmids so they don’t have to be recombined, as this is an unnatural process)
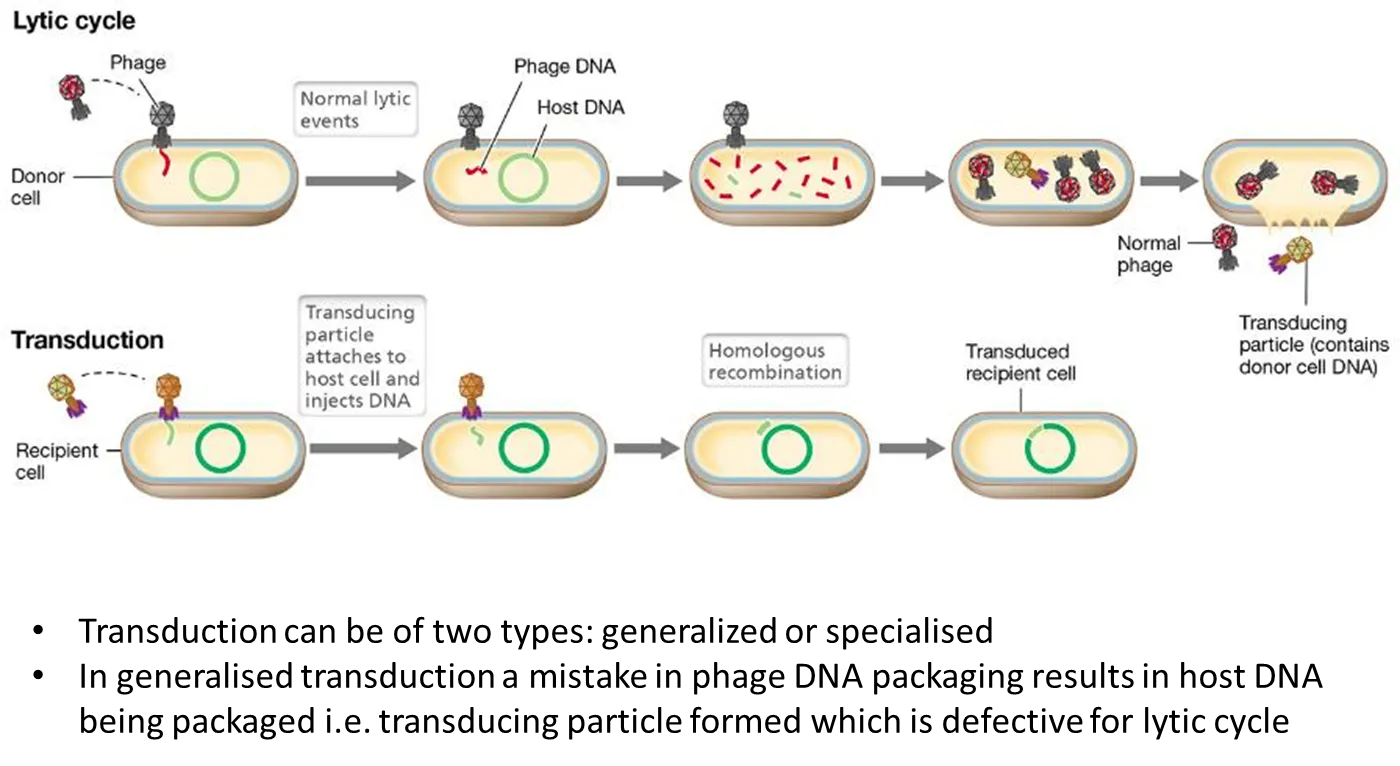
what is generalised transduction?
when a bacteriophage virus infects a cell, it injects its DNA into the cell, replicates its genome and goes into a lytic cycle
generalised transduction can sometimes occur when the nucleases injected also break up the bacterial DNA
this means that when the viruses are reassembled, some bacteriophages will include bits of bacterial DNA, and miss some of their normal DNA
this will make them defective- they can inject their DNA when infecting the host cell, but cannot cause it to lyse
this is a transducing particle, as it also injects the DNA from the original donor cell, which can be recombined
what is specialised transduction?
when some viruses inject their DNA into the host cell, it gets integrated into the host DNA as a prophage, using site-specific endonucleases
this can stay there, being normally transcribed by the bacteria, until some kind of stress, when it excises and replicates (by rolling-circle replication, which is very efficient) when the lytic cycle is induced
rarely, when the prophage enters the lytic cycle, it doesn’t excise correctly and takes some bacterial DNA with it, missing a bit of viral DNA, and so producing a defective phage that will transduce the donor DNA into a host cell
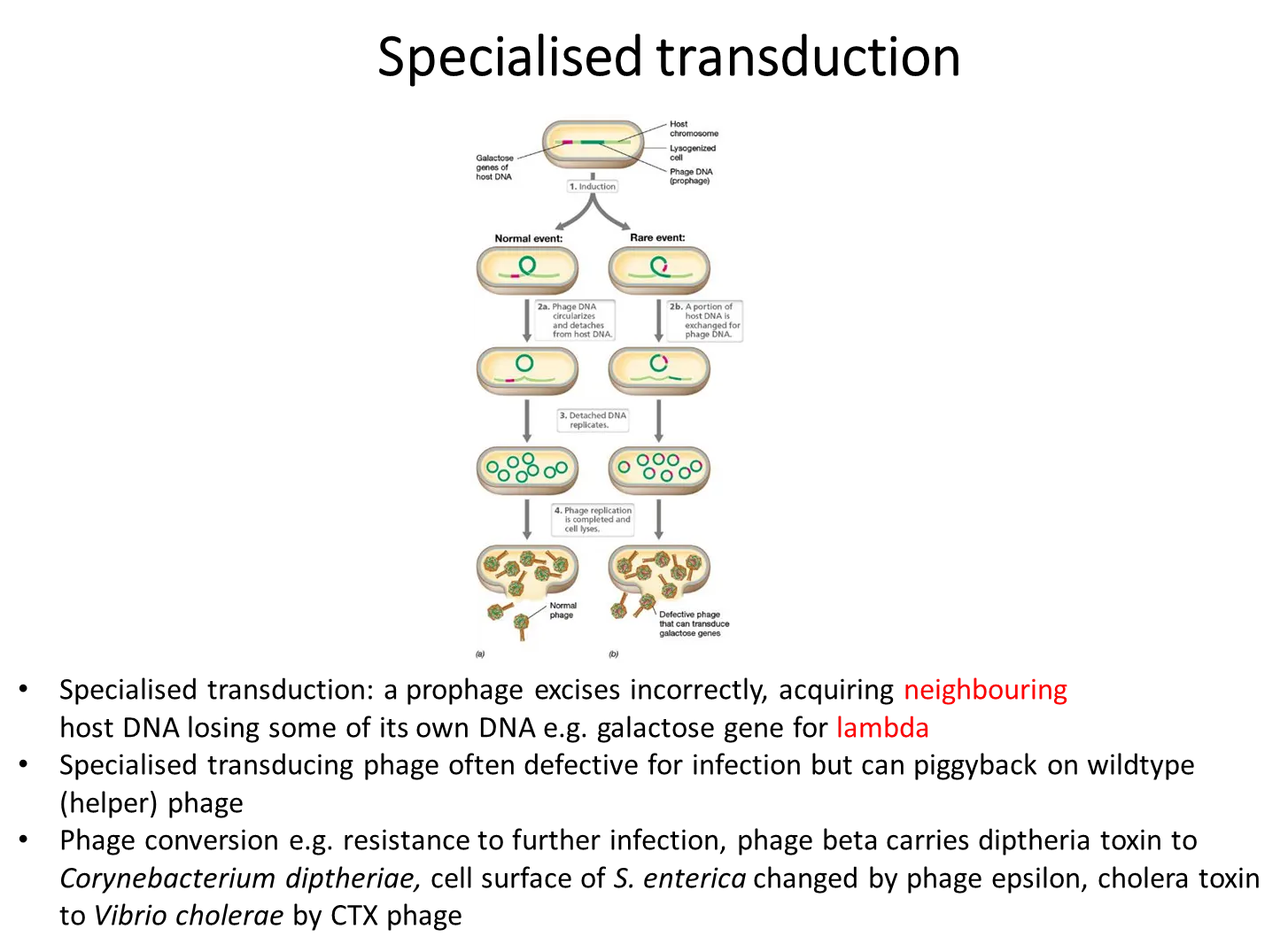
what is an example of specialised transduction?
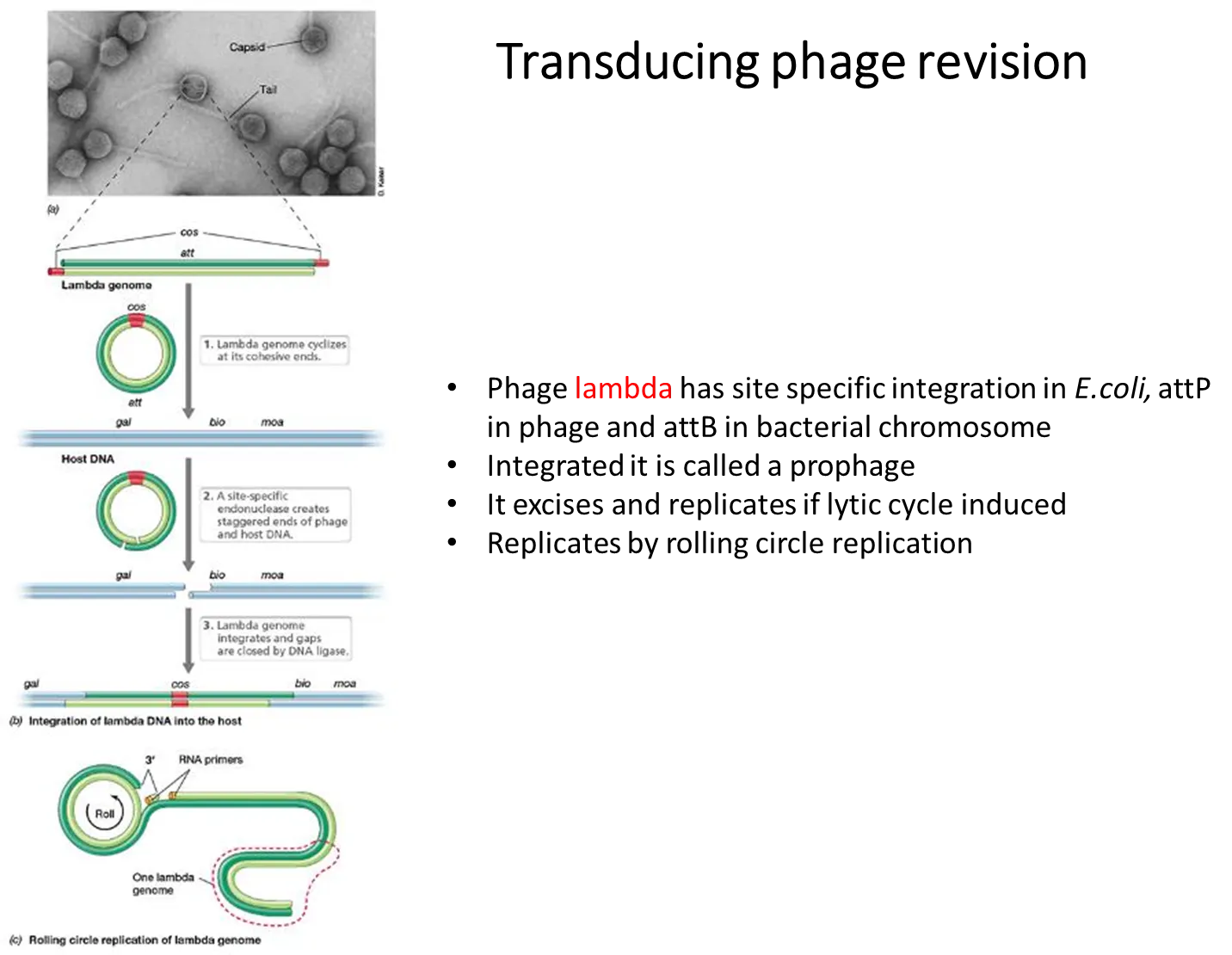
the lambda genome has cos sites at either end that are complementary and can cause the lambda genome to become cyclical
this can then integrate itself into bacterial host DNA at a specific site, through the use of site-specific endonucleases
if the prophage then doesn’t excise correctly, it could take a section of the gene on either side in the host DNA eg. the galactose operon, which will be transduced to another bacterium
this is specialised because only a select few genes on either side can be transduced, not just any gene from the digested host DNA in generalised transduction
what are gene transfer agents?
defective phages that have been hijacked by bacteria to transfer its host DNA to other bacteria and not be lytic
what is the process of conjugation? use an example
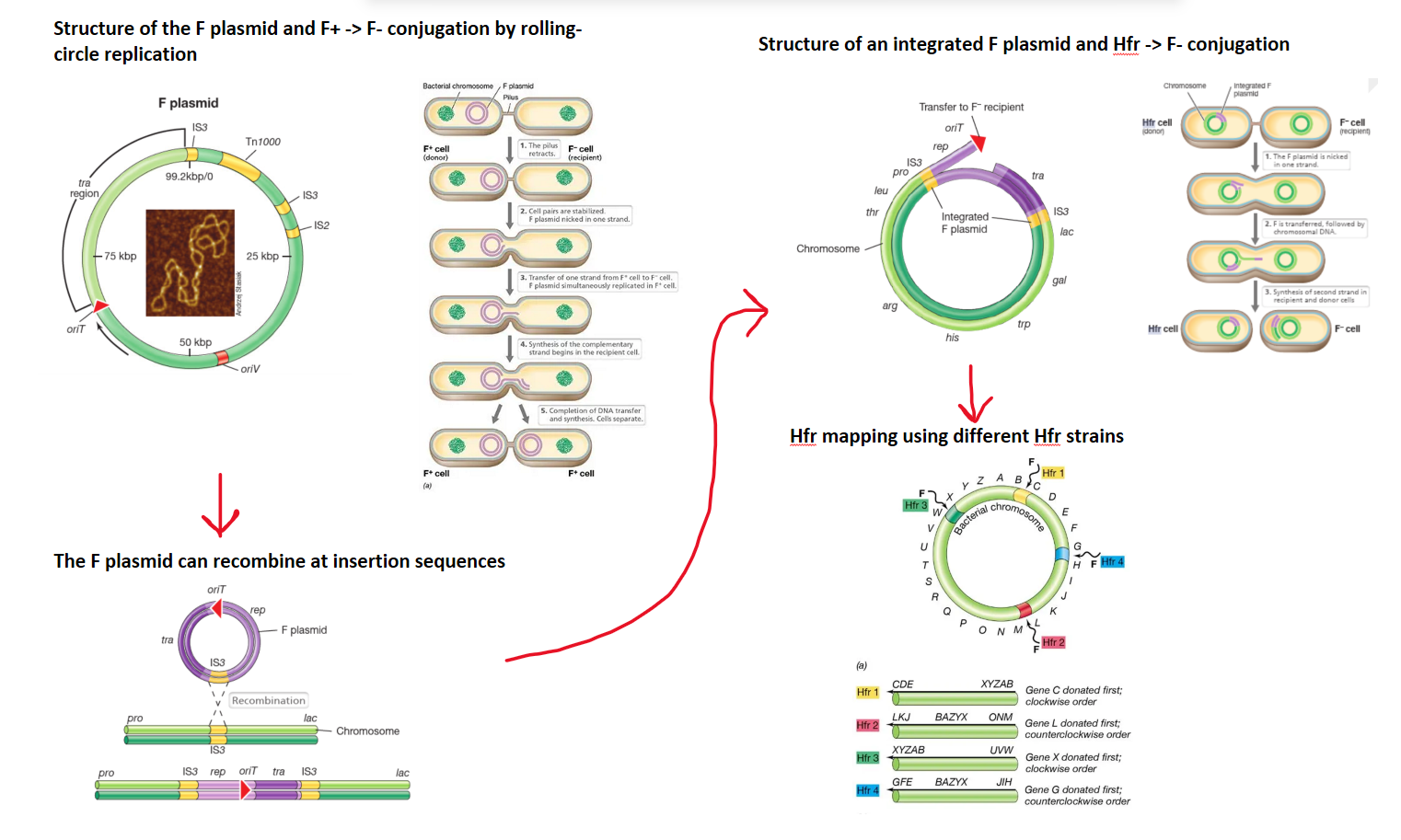
the fertility plasmid has all the genes needed to carry out conjugation, and can be transferred from a donor (F+) to a recipient that doesn’t have it (F-)
it has tra genes which allow it to form a sex pillus- a bridge that draws the two cells together to form a mating junction
the plasmid is nicked in one strand by the traI enzyme, and this strand moves into the F- cell, while both strands are simultaneously replicated, to form two double stranded plasmids, so that both cells are F+ (rolling circle replication)
in this way, the F plasmid can transmit iself between different cells
the F plasmid is an episome (a plasmid that can recombine and integrate into the chromosome), as the plasmid has multiple insertion sequences complementary to sections of the chromosome
if the F plasmid integrates into the chromosome it produces an Hfr cell (high frequency of recombination)- different Hfr strains are produced when different insertion sequences are targeted
consequent conjugation between an Hfr donor and an F- cell will then transfer a section (normally the pilus is broken before the whole chromosome can be transferred) of the donor’s chromosome
this must recombine into the F- cells chromosome because it can’t replicate itself- this is because it doesn’t contain the tra operon, which is always at the end of the transfer (origin of transfer, oriT, is next to the tra gene in the plasmid, but goes in the opposite direction),
different Hfr strains will have the origin of transfer at different points in the chromosome, and will transfer in different directions, so they can be used to map the genome of an organism by conjugation interruption experiments
what are transposable elements and what are the two types found in bacteria?
transposable elements are stretches of DNA that can move from one site to another, and are found across all three domains
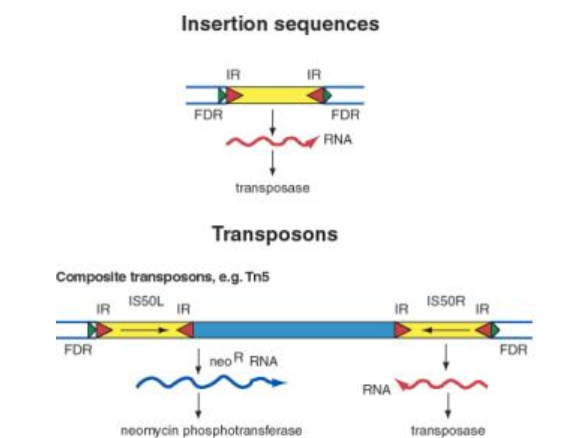
insertion sequences- short DNA segments that code for transposase, flanked by inverted repeats
transposons- longer DNA segments, essentially two insertion sequences with an extra gene (or multiple) inbetween, which also gets moved as part of the transposon
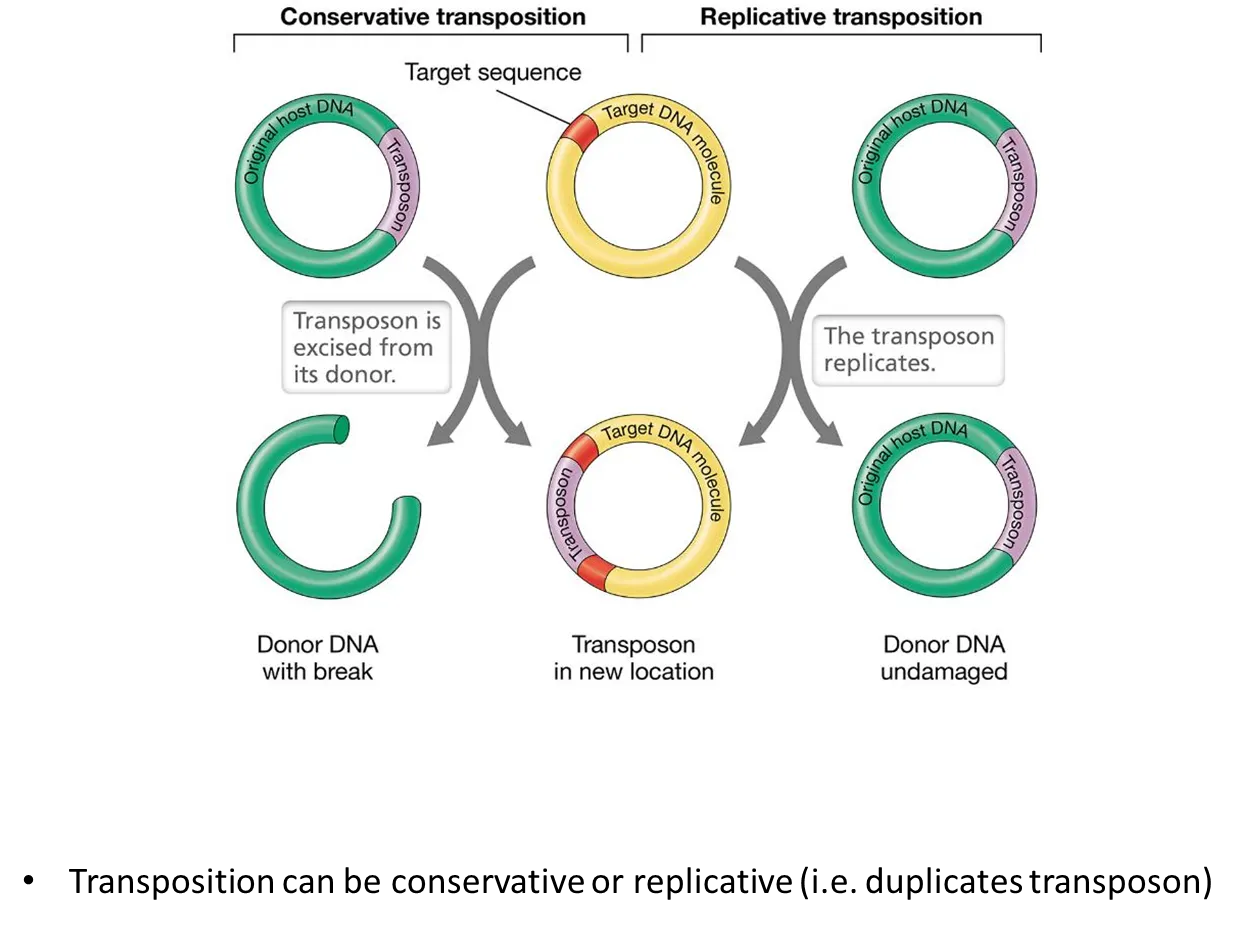
what are the two mechanisms of transposition?
in conservative transposition, the transposons just get moved to the target DNA
in replicative transposition, the transposons get copied into the target DNa, and the donor DNA is unchanged
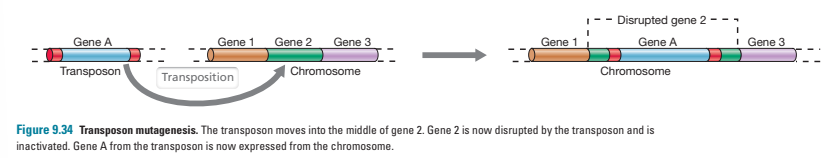
what is transposon mutagenesis?
when a transposon inserts itself into a gene, the gene is split into two parts and the sequence is altered- this essentially creates a mutation
this occurs naturally but can be used to create mutants in labs- transposons carrying antibiotic resistance genes are normally used so that cells that didn’t successfully recombine will be killed when cultured in antibiotics
why do we need DNA?
RNA mutates spontaneously- the deamination of cytosine into uracil is very common, and hard to detect because it is single-stranded and because U is one of the bases anyway (unlike in DNA)
it also has the 2’OH group, which allows it to form more H bonds and fold, and nucleophilically attack the positive phosphate groups to cleave the backbone
RNA itself is generally stable, but is unstable in the current protein world because of the presence of RNAases
we need long stable sequences of information to produce enough proteins
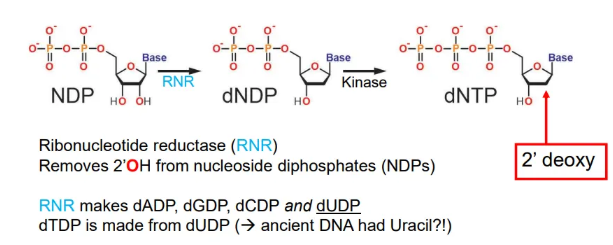
how are the DNA building blocks produced?
nucleoside diphosphates (NDPs) are converted into deoxynucleoside triphosphates (dNTPs) by ribonucleotide reductase (RNR), which removes the 2’OH, and kinase enzymes which add a third phosphate group
how does DNA replication occur?
DNA helicase separates the two strands by breaking the H bonds
complementary RNA primers are attached by primase at the start of the leading strand (replicated 5’ → 3’) and at regular intervals in the lagging strand (3’ → 5’)
DNA polymerase adds on the dNTPs following base pairing rules (condensation reaction releasing pyrophosphate), producing Okazaki fragments in the lagging strand
RNAse H degrades the RNA primers, and the fragments are extended until ligase joins the phosphate backbones together
topoisomerases remove DNA positive supercoils from helicase unwinding
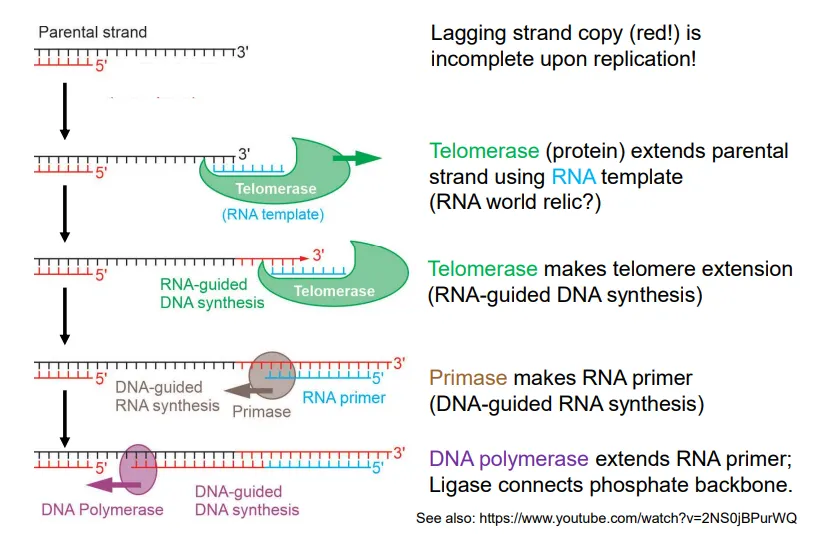
what happens at the end of DNA replication in the lagging strand in eukaryotes?
to form the final Okazaki fragment, telomerase extends the parental strand using an RNA template
primase attaches an RNA primer to this extended DNA strand
DNA polymerase extends the primer until the strands can be connected by DNA ligase
how is transcription initiated in prokaryotes?
the sigma factor (a cofactor of RNA polymerase) recognises and binds to the Pribnow and TATA box motifs in the promoter region upstream of the initiation site
this recruits RNA polymerase to bind to the DNA, produce a transcription bubble and begin RNA synthesis in the 5’ to 3’ direction using NTPs
the sigma factor dissociates

how is transcription terminated in prokaryotes?
after the stop codon is transcribed, termination signals found in the 3’UTR of the mRNA strand are also transcribed
these can either be:
inverted repeats, which cause hairpin loops (through base pairing) that will terminate transcription by RNA polymerase
a rut termination sequence, which is recognised by the rho protein that binds to RNA polymerase, terminating transcription

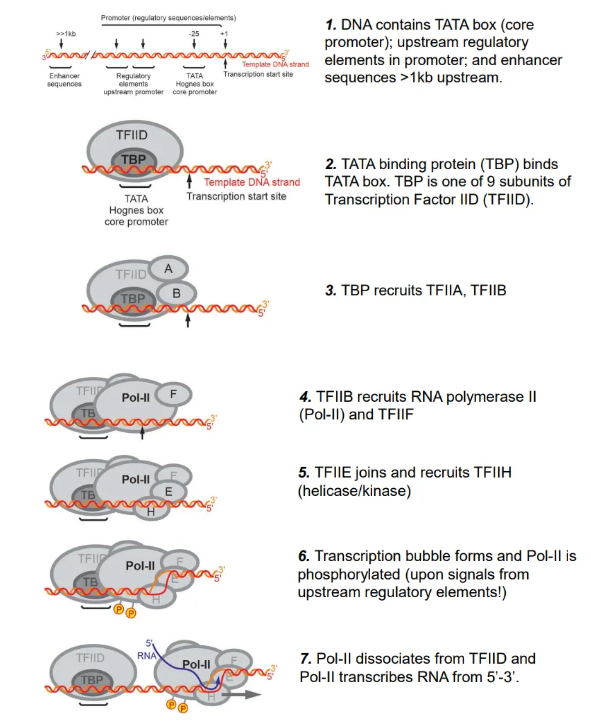
how is transcription initiated in eukaryotes?
the TATA binding protein (TBP), a subunit of transcription factor IID (TFIID), binds to the TATA box in the promoter region upstream of the initiation site
this recruits multiple proteins, including RNA polymerase II (Pol-II), which forms a transcription bubble, detaches from TFIID and begins to transcribe the RNA upon phosphorylation using NTPs
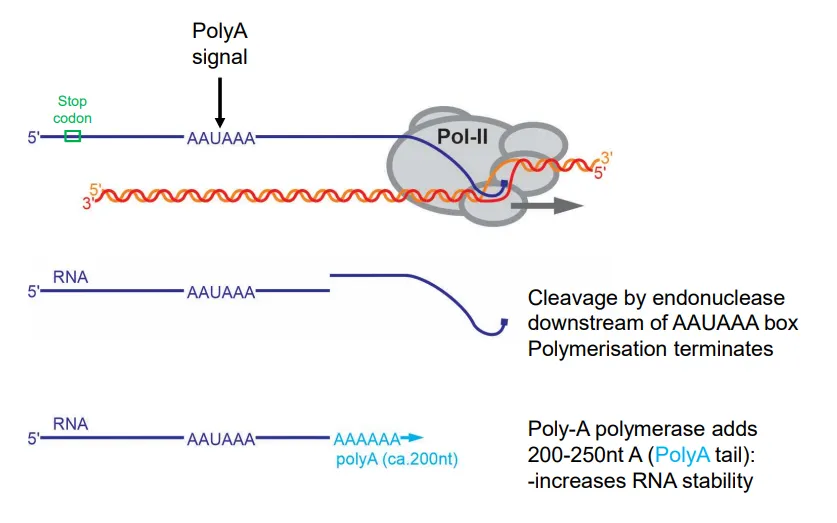
how is transcription terminated in eukaryotes and how is the mRNA modified?
after the stop codon is transcribed, a polyA signal (AAAUAAA) is found in the 3’UTR
this causes cleavage downstream by endonuclease enzymes, terminating transcription
a polyA tail (200-250 A nucleotides) is added to the 3’ end to increase the mRNA stability
the introns are then removed from this preRNA by splicing, catalysed by spliceosome (a ribozyme)
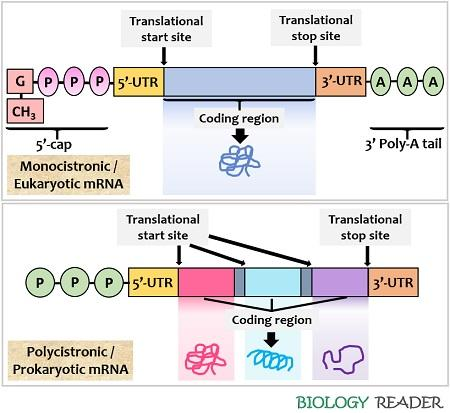
how is mRNA modified in prokaryotes and eukaryotes and why?
prokaryotes don’t have membrane-bound nuclei, so transcription and translation can occur simultaneously
at the 5’ end, they just have a triphosphate purine nucleotide
eukaryotic mRNA has to be transported out of the nucleus, so it is modified in a more complicated way:
a 5’ cap is added (made from guanosine triphosphate, and involving the methylation of the first two bases)
at the 3’ end a poly(A) tail is added by poly(A) polymerase (200-250 A nucleotides)
introns are also removed by splicing
these modifications aid in recognition as mRNA + export, translation initiation + efficiency (by cirularisation) and stability
how is the amino acid code redundant?
multiple codons can code for one amino acid
in these cases, it is the third base in the codon that is unimportant and can be different
these are called ‘wobble bases’
normally the interchangeable bases will be the two purines or the two pyrimidines
in a few codons the third base can be anything
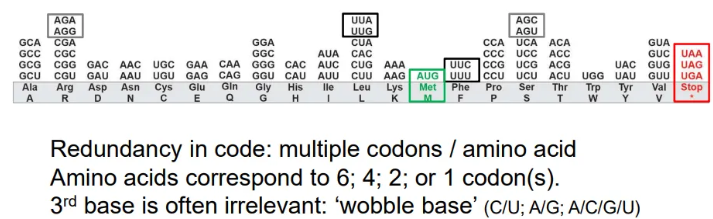
what are the start and stop codons in mRNA?
start
AUG
stop
UAA
UAG
UGA

how is the correct open reading frame selected?
the first start codon (AUG) found in the mRNA strand will determine where translation starts, and hence the reading frame
since prokaryotic mRNA is polycistronic, this is the first AUG after the Shine-Dalgarno sequence
whereas in eukaryotes, this is just the first AUG from the 5’ end
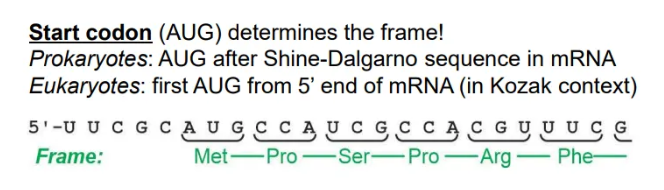
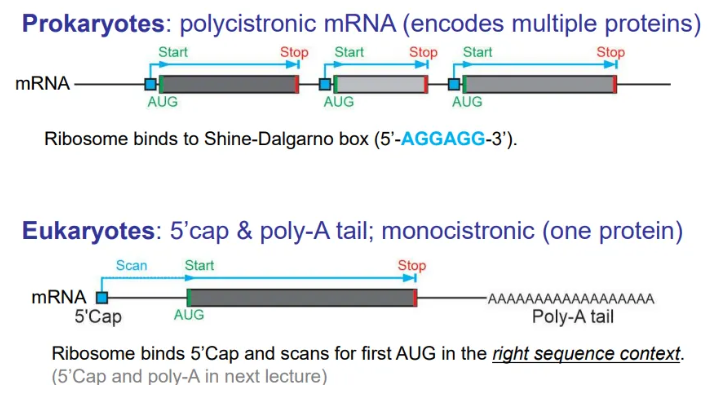
what are the general layouts of mRNA in prokaryotes and eukaryotes?
translation of prokaryotic mRNA begins before transcription is complete, because they don’t have membrane-bound nuclei, so the mRNA is polycistronic (encodes multiple proteins)
in eukaryotes the mRNA strands are cleaved and packaged to be sent to the ribosomes for translation, so the mRNA is in discrete chunks for each protein (monocistronic)
how do tRNA molecules deal with the redundancy of the genetic code?
there are only 31-41 distinct tRNAs per cell, compared to 60 codons, so many must have anticodons that can recognise the wobble bases
they do this by having their own modified RNA bases
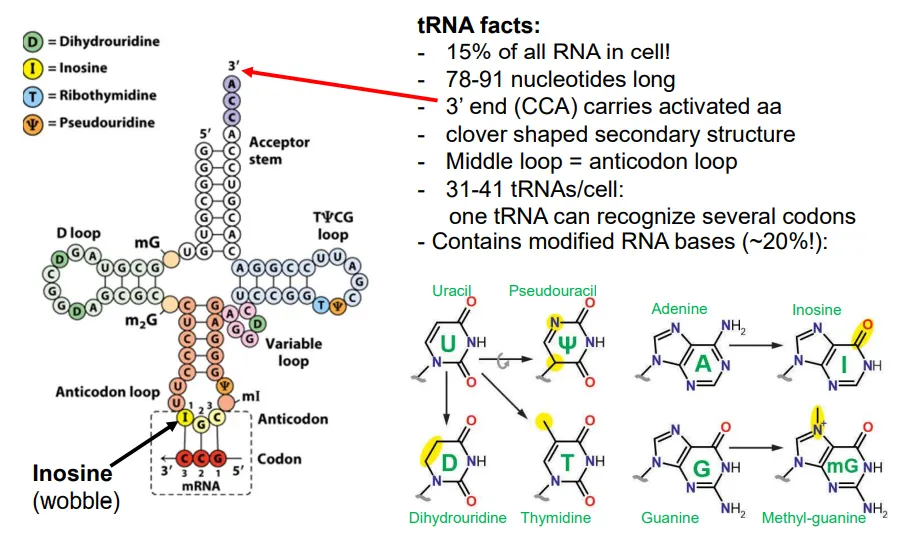
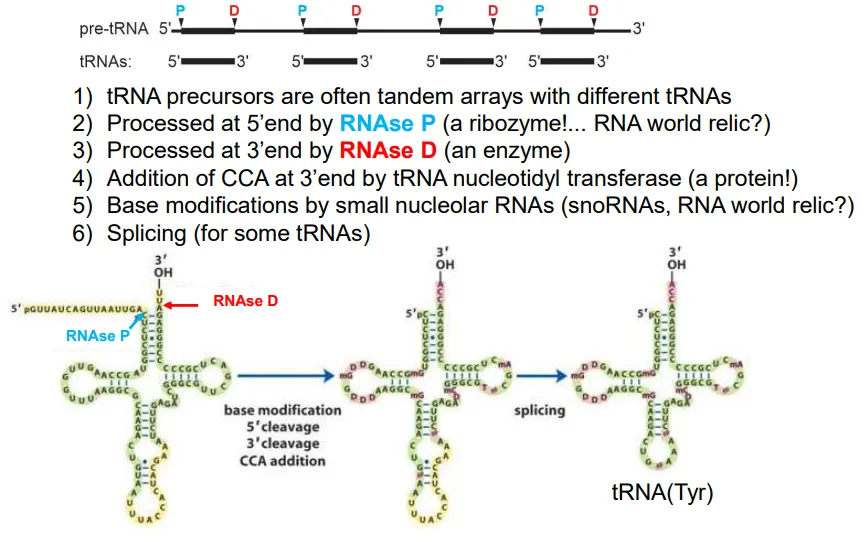
how are tRNA molecules produced?
the pre-tRNA molecules normally contain the sequences for multiple tRNAs (tandem arrays)
the sequence is cleaved at multiple points
at the 5’ end of each tRNA this is done by RNAse P, and at the 3’ end it is RNAse D
then the enzyme tRNA nucleotidyl transferase adds CCA to the 3’ end
base modifications occur in some tRNA molecules to deal with the wobble bases, and some get spliced
these molecules are then ‘charged’ by an aminoacyl-tRNA synthetase (aaRS) to add on the relevant amino acid to the new 3’ adenosine (at the 3’OH) using ATP
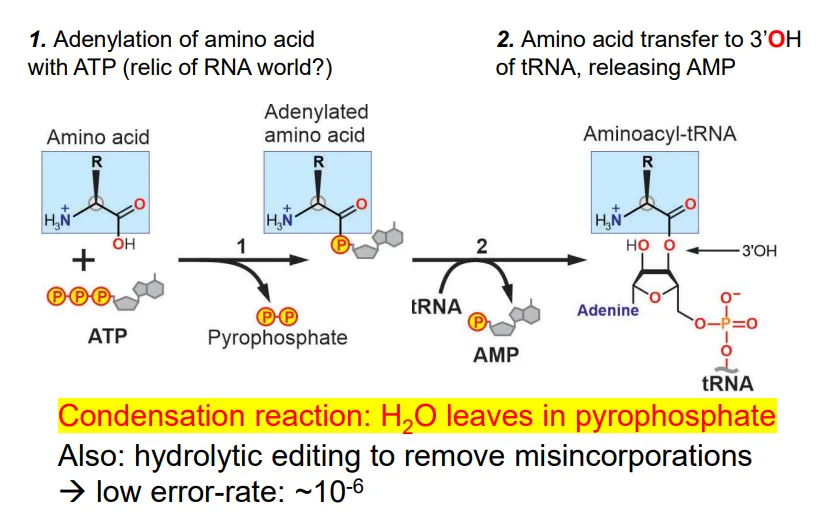
how are tRNA molecules charged?
the amino acid reacts with ATP to produce an adenylated amino acid (bound to AMP) and a pyrophosphate (contains water- this is a condensation reaction)
the AMP is replaced by a reaction with the 3’OH of the 3’ adenosine in the tRNA molecule
this reaction is catalysed by aminoacyl-tRNA synthetase enzymes (aaRS)- only 1 per amino acid)
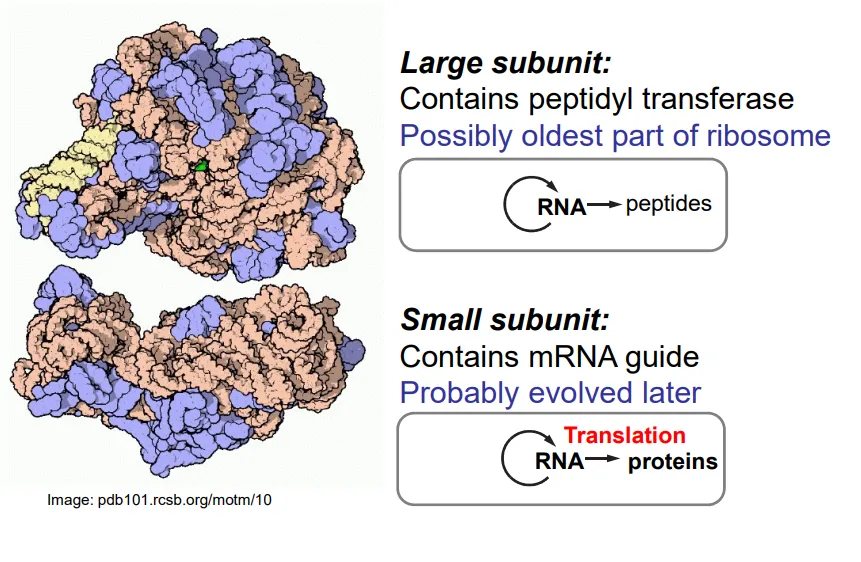
what is the structure of ribosomes?
ribosomes are mostly composed of RNA, with some peripheral auxiliary proteins
the large subunit contains peptidyl transferase (a ribozyme), which connects the amino acids by catalysing the formation of peptide bonds
the tRNA molecules bind to the large subunit
the small subunit binds to the mRNA so that codon-anticodon pairing can occur with the tRNA
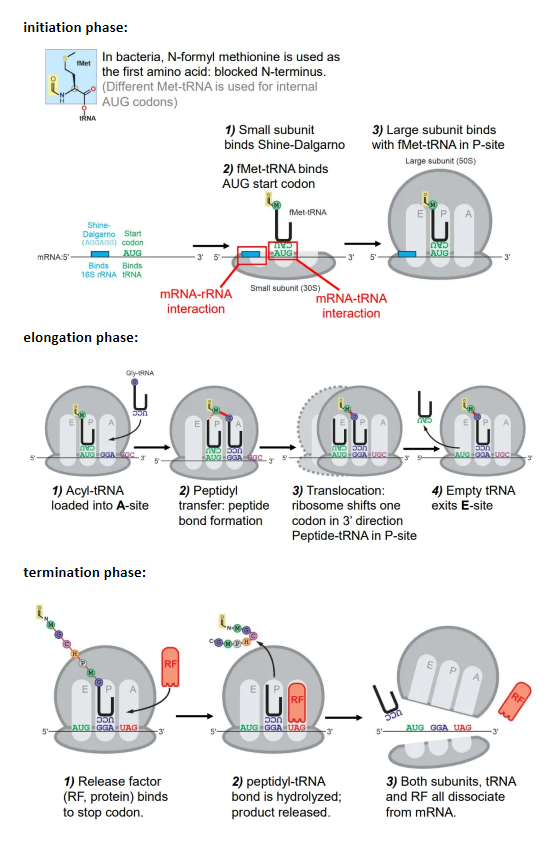
what happens in the phases of translation in prokaryotes?
initiation:
the small subunit of the ribosome binds to the Shine-Dalgarno box
the Met-tRNA (methionine = start aa) binds to the first AUG codon after the Shine-Dalgarno box
the large subunit then binds to the Met-tRNA at the P site
elongation:
the charged tRNA molecule for the next codon is loaded into the A site, and a peptide bond forms between the two amino acids
the amino group of the incoming amino acid attacks the carbonyl carbon nucleophilically
this is catalysed by the peptidyl transferase ribozyme, which contains an adenine base that accepts and donates a proton to hydrolyse the peptide-tRNA bond
the ribosome shifts one codon, so the first tRNA is now in the E site, and exits
the second tRNA is now in the P site, so the process repeats, with new tRNA molecules being loaded into the A site and peptide bonds forming
termination:
a release factor (RF) protein binds to the stop codon at the A site (molecular mimicry)
this causes the peptide-tRNA bond to be hydrolysed, so the peptide is released
both subunits, the tRNA and the RF protein dissociate from the mRNA
why are proteins better catalysts than RNA?
proteins are much more compact + detailed than ribozymes
they have a hydrophobic core, so water is excluded from the active site
20 amino acids vs 4 bases, so more variability
amino acids are smaller units than nucleotides
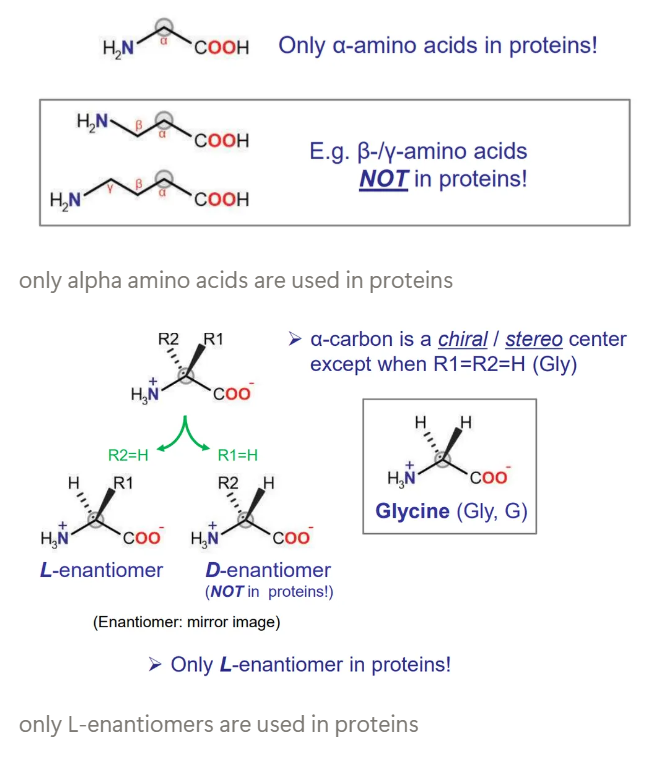
what kinds of amino acids are used in proteins?
alpha-amino acids, not beta or gamma
L-enantiomers, not D
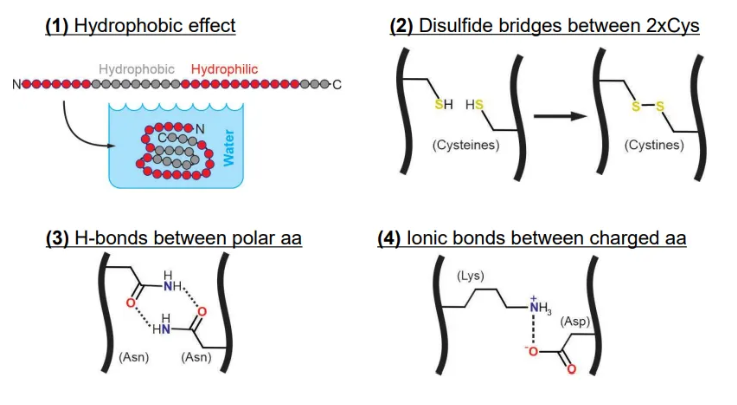
what forces dictate the tertiary structure of proteins?
the hydrophobic effect (higher entropy because water molecules can’t form cages around hydrophobic residues, so they become less ordered)
H-bonds between polar amino acids
ionic bonds between oppositely charged amino acids
disulphide bridges between cysteines can be involved but aren’t very useful in defining structure because they need oxidising conditions to form (extracellular)
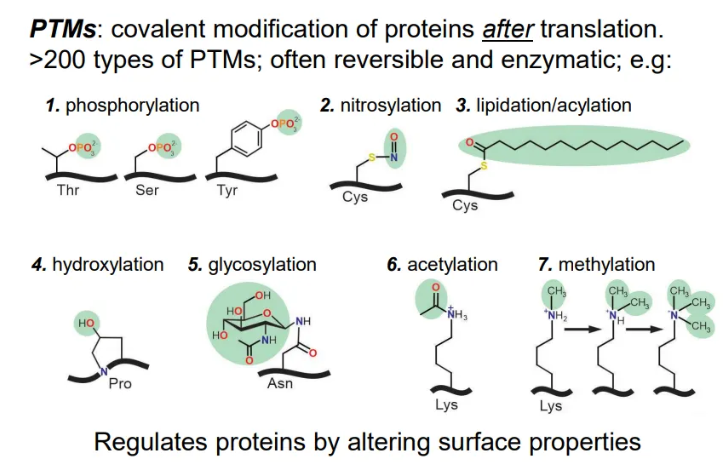
what are the 7 types of PTM?
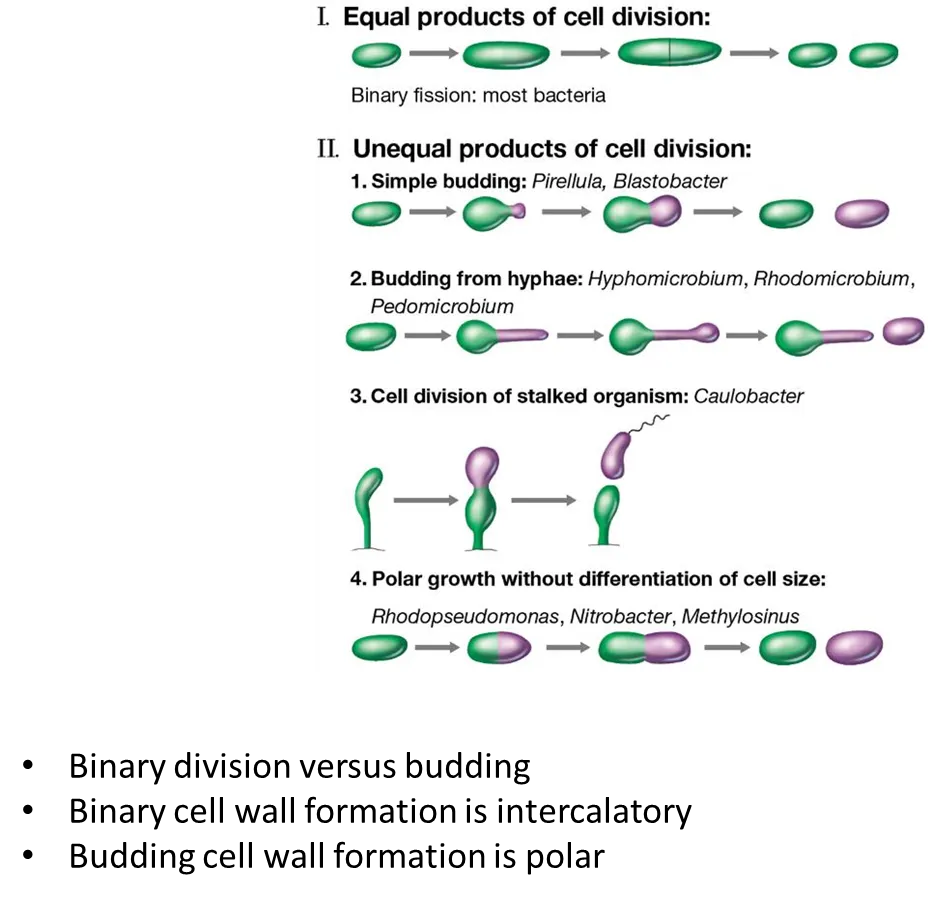
how can bacteria divide?
by binary fission (symmetrically):
cell elongates then forms a septum in the centre, which leads the cell to separate
this is intercalatory growth- new bits of cell wall are inserted all the way around the cell
by polar growth (asymmetrically):
budding
budding by hyphae
from stalked (specialised) cells producing a motile cell
in polar growth, the new cell wall is all inserted on one side (the purple cell will be all new cell wall, the green cell will be all old cell wall)
how can the growth of bacteria be measured?
viable cell counts- culturing the sample on a solid medium and counting the colonies produced
turbidity- the optical density (due to light scattering, not absorption) can be measured by spectrophotometers, which is much faster than culturing
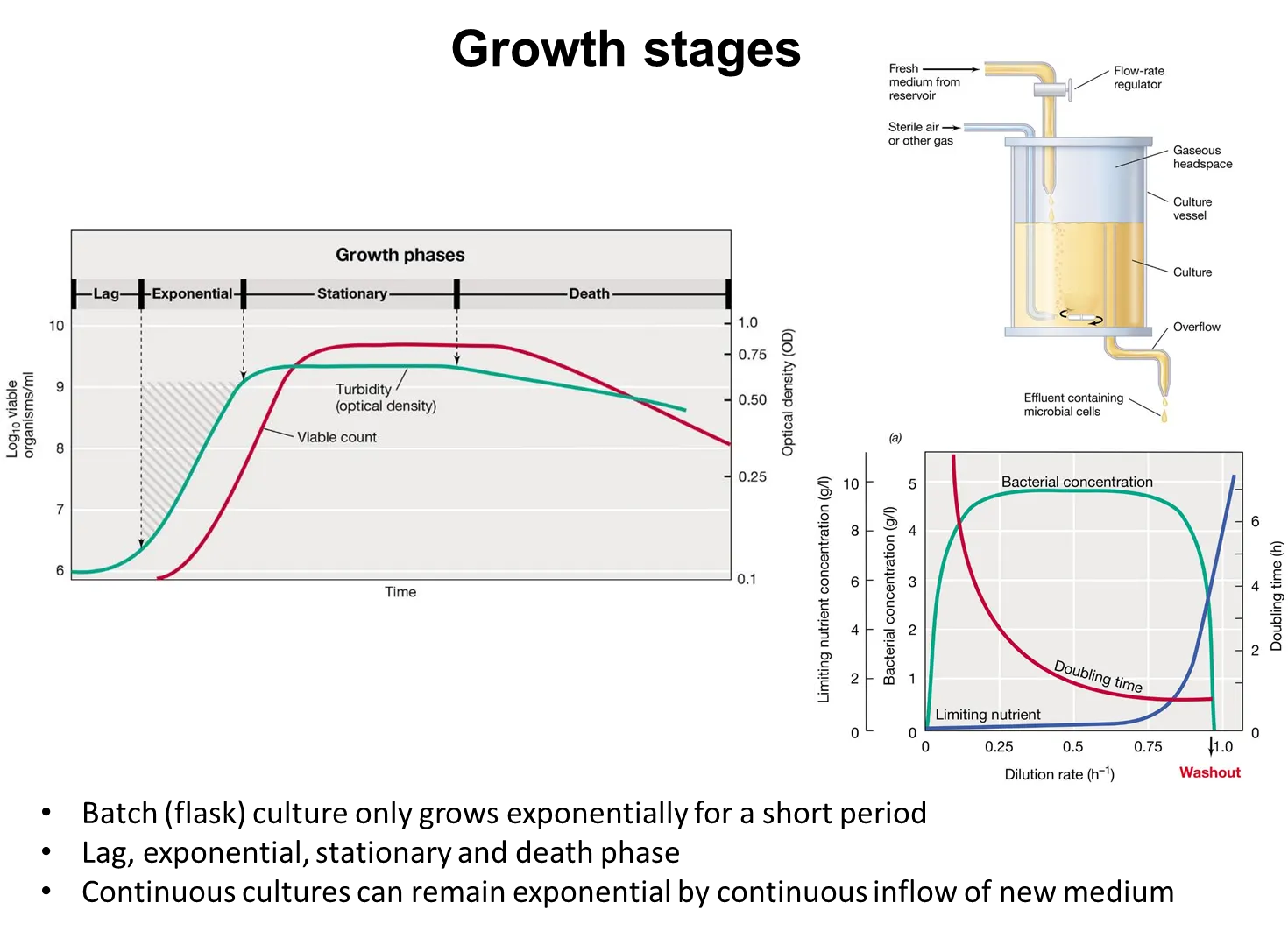
how does bacterial growth vary with the dilution rate in continuous cultures?
in a chemostat (continuous culture) at steady state, the cells grow at the same rate as they are removed from the system, because the cells are competing for a limiting nutrient, so the nutrient added is consumed rapidly
as the dilution rate increases, the cells can grow faster and the doubling rate decreases
they keep pace with the dilution rate until it is so fast that it exceeds the maximal growth rate, where the cells can’t keep up and get washed out
what are compatible solutes and how are they used?
compatible solutes such as sugars, alcohols and amino acid derivatives, increase in concentration in the cell when in high osmolarity environments (low water potential), either by being pumped into the cell or synthesised
these are compatible with the cell’s biochemistry because they don’t mess with cell processes when in high concentrations, like other solutes eg. sodium
this prevents the cell from becoming dehydrated, because water will flow into the cell (positive water balance)

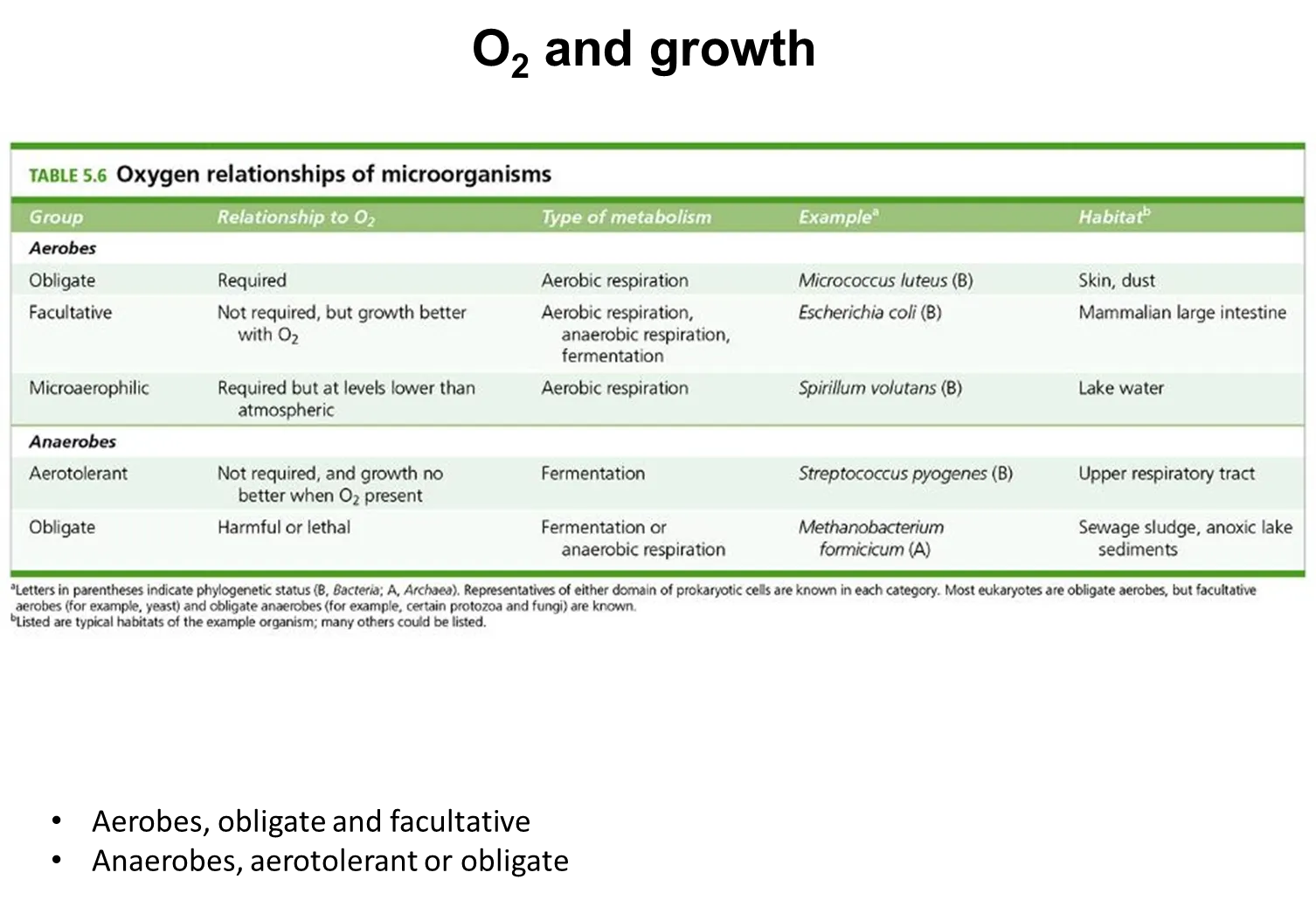
how do different microbes respond to oxygen availability
obligate aerobes need oxygen to survive- they can’t survive in anaerobic conditions
facultative aerobes grow better in oxygen, but can survive in anaerobic conditions
microaerophilic aerobes need oxygen to survive, but at less than atmospheric concentrations
obligate anaerobes need the absence of oxygen- they can’t survive in aerobic conditions
aerotolerant anaerobes don’t need oxygen, and don’t grow any better or worse in aerobic or anaerobic conditions
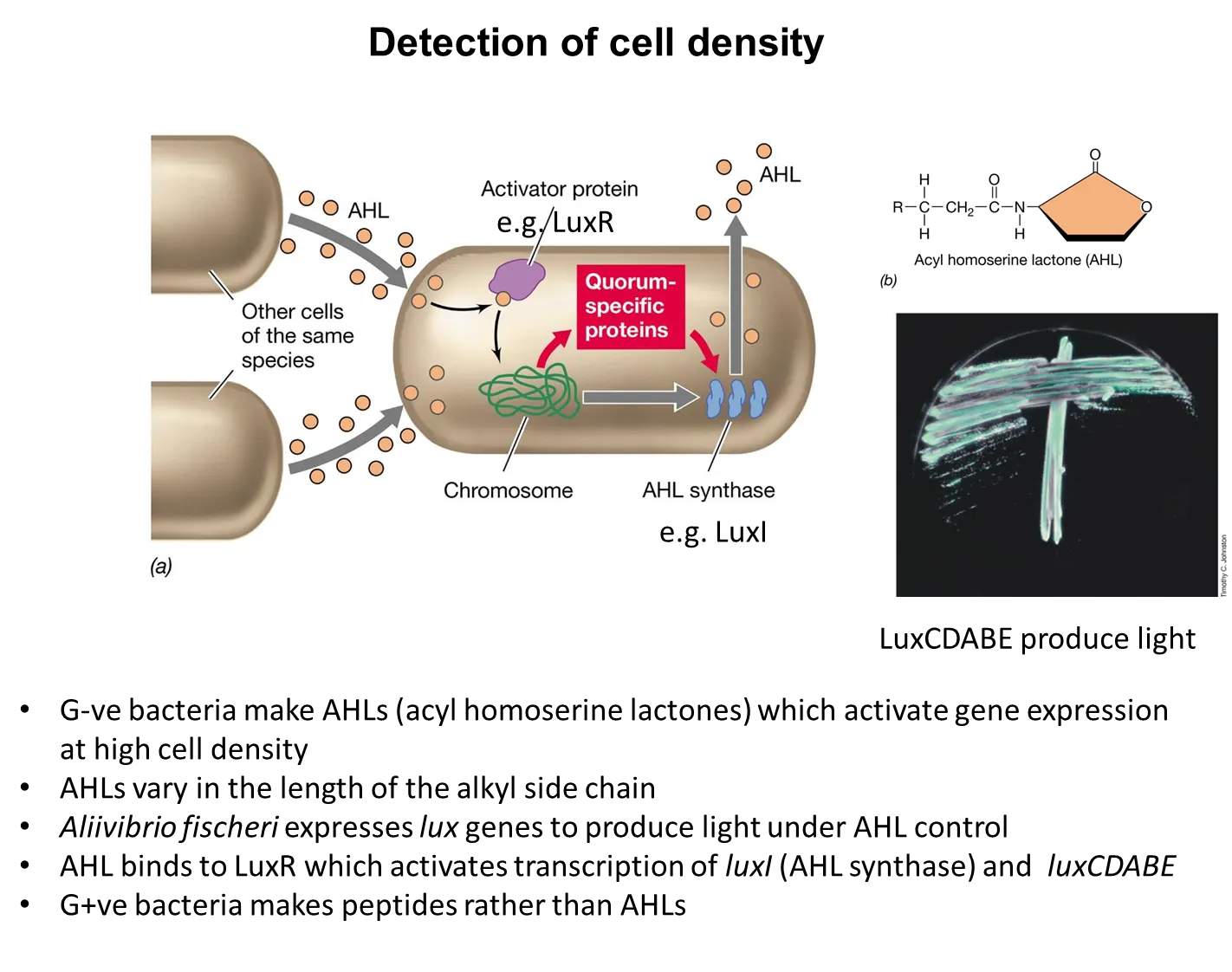
how do G+ve and G-ve bacteria detect cell density and change their behaviour?
bacteria produce signalling molecules, which build up when the cell density is high
they change their behaviour based on these concentrations, by changing gene expression
eg. pathogens don’t produce toxins with low cell density because they won’t make enough difference, but then switch on toxin production in high cell density
G-ve bacteria use acyl homoserine lactones (AHLs) whereas G+ve bacteria use peptides
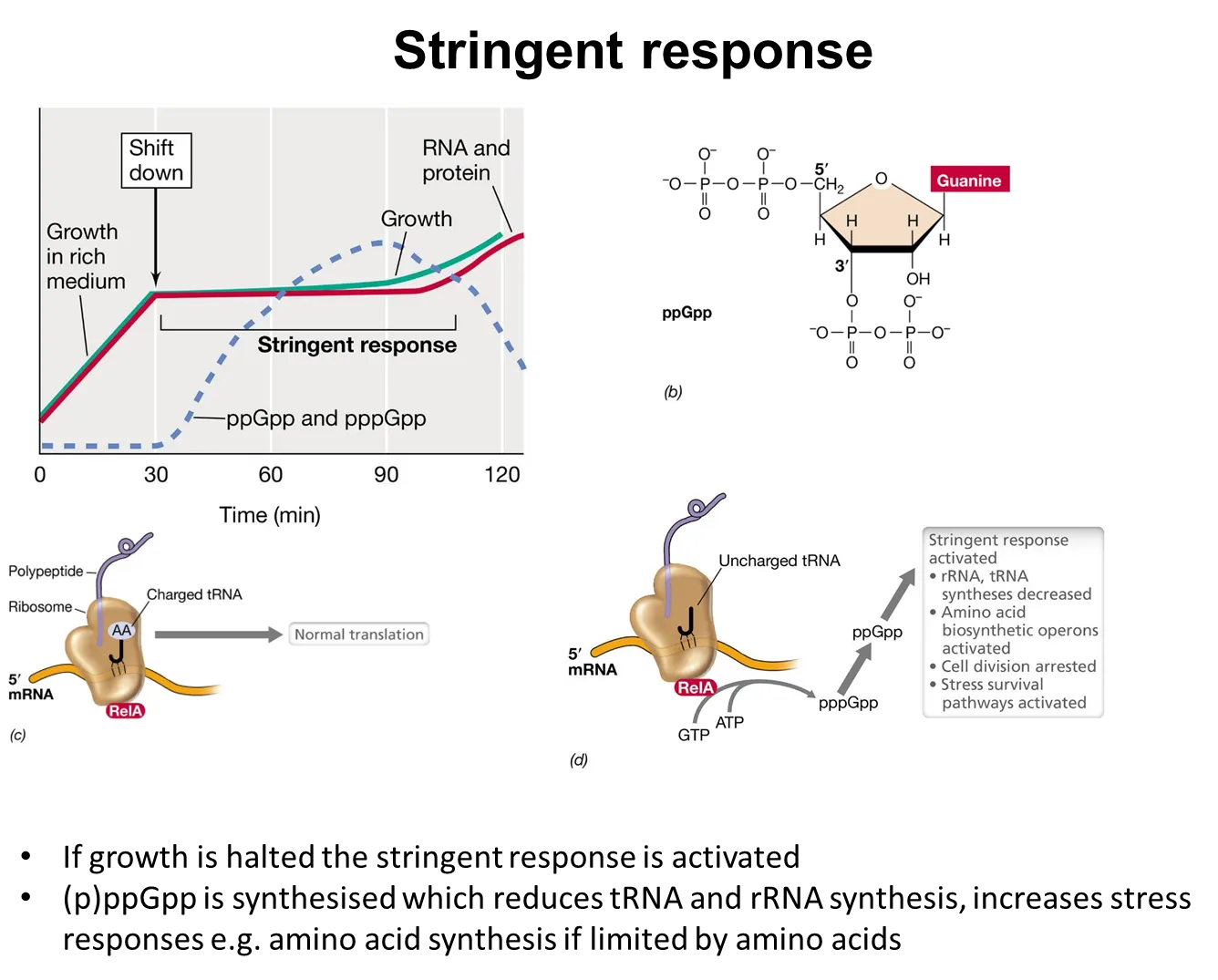
what is the stringent response?
bacteria have to respond to nutrient limitations and stressors
if growth slows, the stringent response is activated
this is triggered when charged tRNAs aren’t used in translation (because there aren’t enough amino acids to charge them)
this causes the relA protein to produce tetraphosphate guanosine, which binds to RNA polymerase and changes its promoter preference
this shifts gene expression to activate stress and survival pathways to adapt to the environment eg. decreasing translation, increasing amino acid biosynthesis, stopping cell division
(the SOS response is triggered by significant DNA damage)
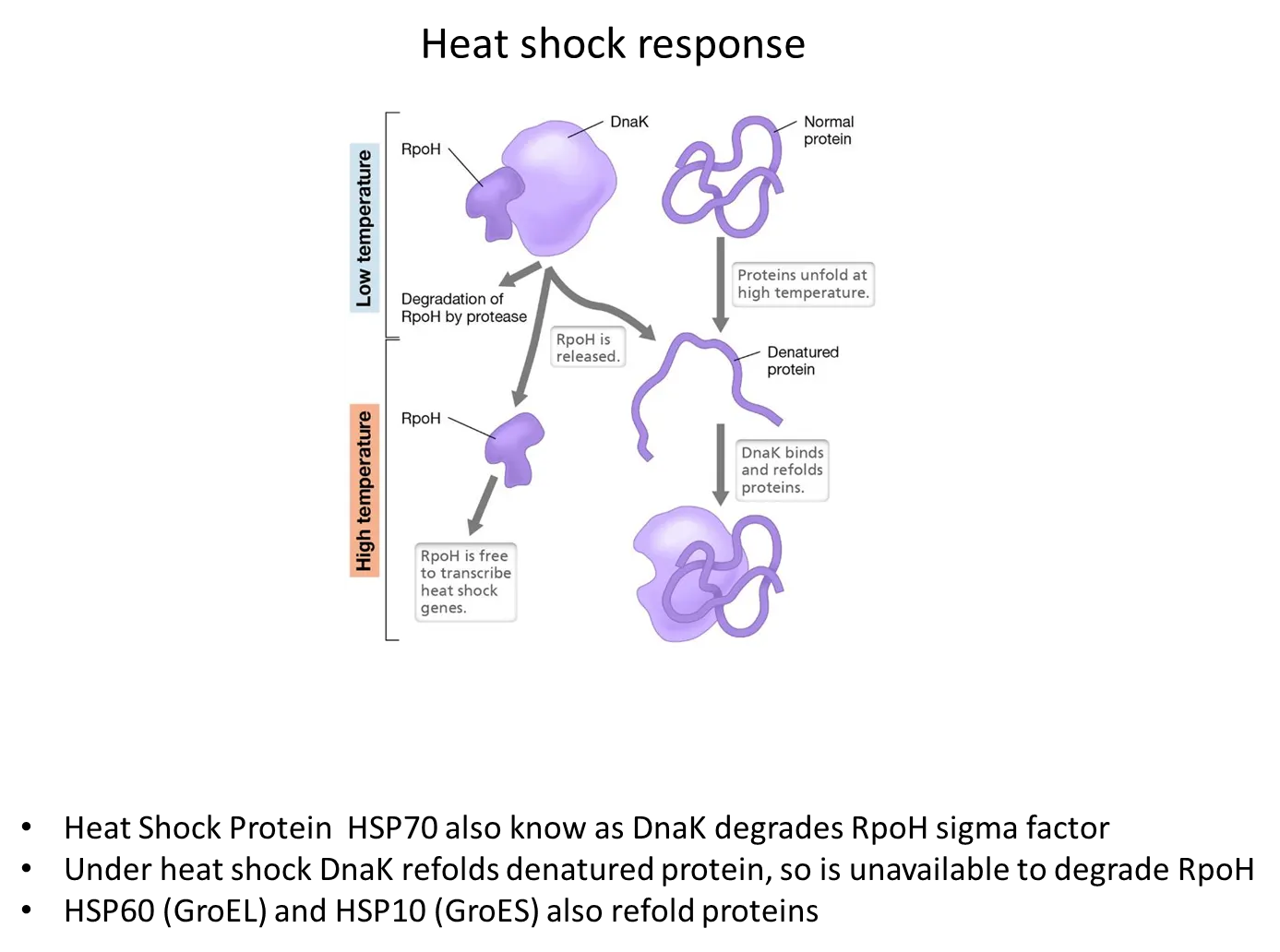
what is the heat shock response in bacteria?
at low temperatures, the DnaK protein has a high affinity for the RpoH sigma factor and degrades it so heat shock genes aren’t expressed
At high temperatures, DnaK releases RpoH (which can then transcribe heat shock genes) and starts to bind to and refold denatured proteins instead
what are bacteriocins?
bacteriocins are antibiotics that bacteria can produce to kill closely related bacteria (other strains of the same organisms but not the same strain) to compete to pass of genes
these are normally contained in plasmids, which can be transferred horizontally
what are incompatibility groups?
two different plasmids in an incompatibility group will use the same replication mechanism
this makes them incompatible because they ‘mess up’ each other’s replication
only one of these can exist at once in a cell
how is the bacterial cell cycle initiated and prevented from initiating?
DnaA protein binds to oriC (the origin of the chromosome) using ATP, initiating bidirectional replication of the chromosome from the origin by recruiting other proteins
it is only able to do this when the chromosome is fully methylated (on both strands)
when the cycle of replication is unfinished, only the parental strand will be methylated, while the new daughter strand will be unmethylated
the SeqA protein binds to this hemimethylated DNA, preventing DnaA binding
this means that DNA replication is only initiated when the previous replication cycle is complete
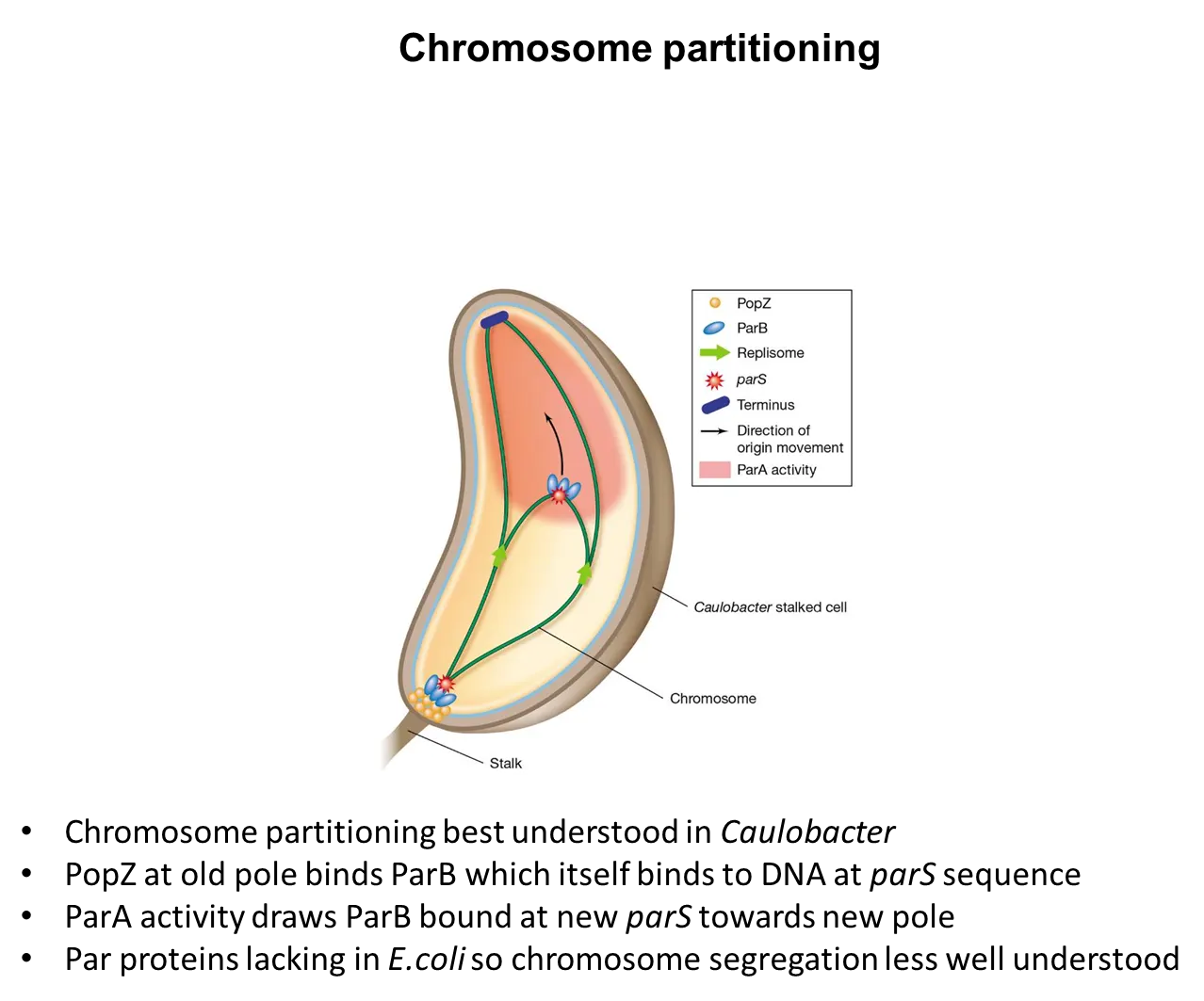
how are bacterial chromosomes partitioned without having spindle fibres?
the PopZ protein found at one pole binds to the ParB protein, which binds to the chromosome at its parS sequence
ParA protein activity at the opposite pole draws the ParB towards the new pole
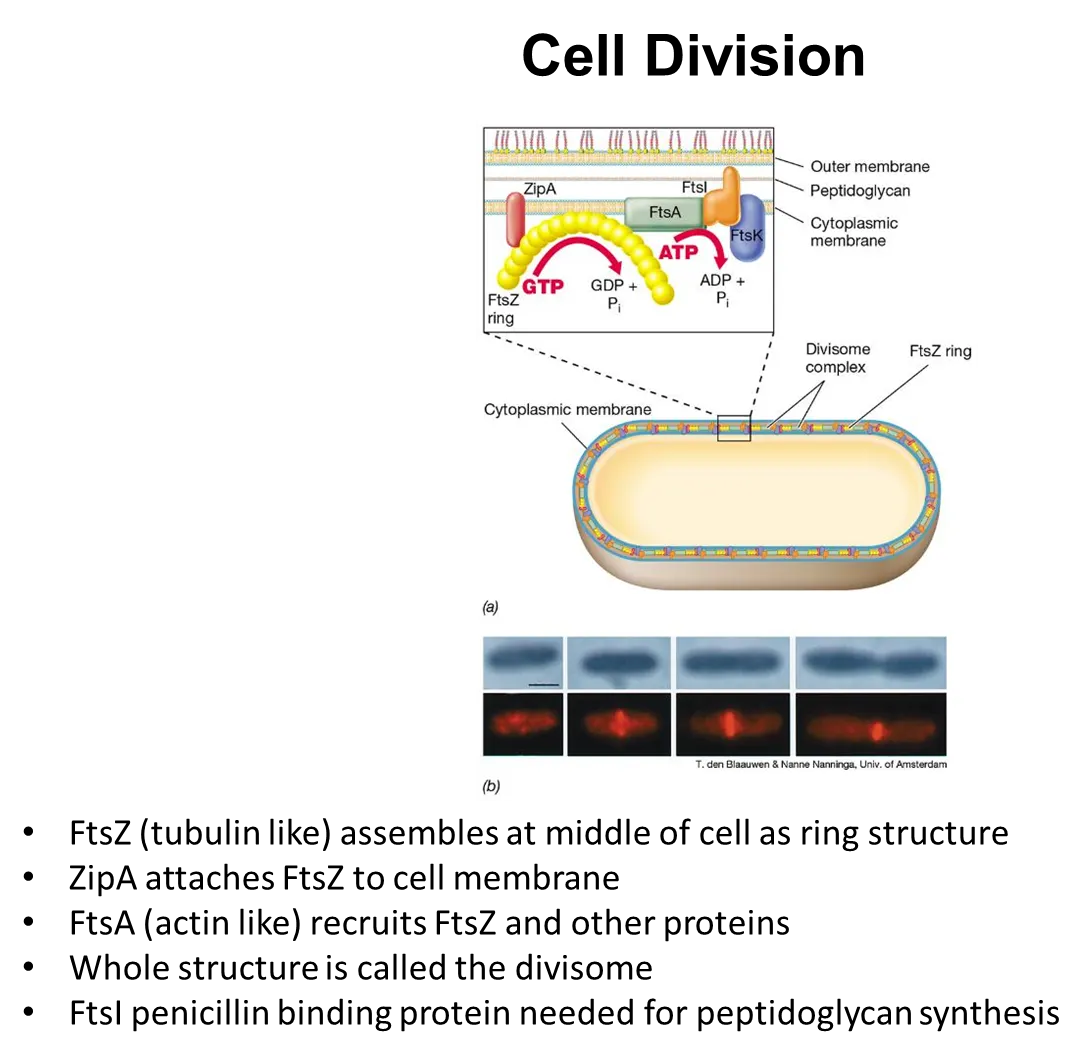
how do bacterial cells contract to divide?
the contractile FtsZ protein (analagous to tubulin) assembles to form a ring in the middle of the cell
the ZipA protein attaches this ring to the membrane
the FtsA protein (analogous to actin) recruits FtsZ and other proteins into a structure called the divisome
the ring gets pulled together to contract the cell
this divisome includes FtsI penicillin-binding protein, a transpeptidase, which catalyses the reaction that cross-links peptidoglycan when forming the septum
meanwhile (in most rod-shaped bacteria), the MreB protein forms in bands around the cell wall, which directs peptidoglycan synthesis, allowing the cell length to increase
how do bacteria find the middle of the cell to divide?
the MinCD protein oscillates from one pole to the other in a spiral
the MinE protein is found near the centre and disperses MinCD towards either pole (so that it has a lower conc in the middle)
MinCD prevents FtsZ polymerisation (the protein that forms a contractile ring in the middle of the cell) when it is in high concentrations, so FtsZ can only work in the middle of the cell

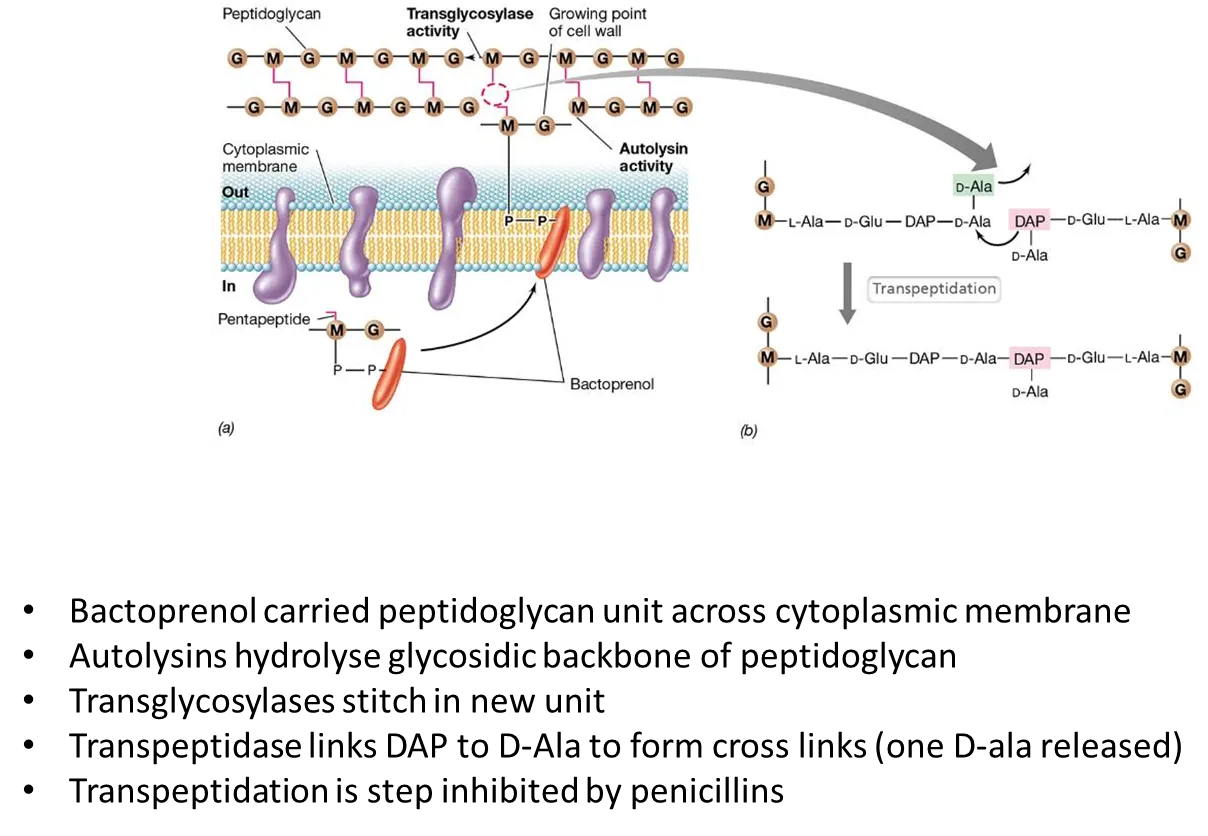
how are peptidoglycan cell walls synthesised?
the NAG-NAM pentapeptide (peptidoglycan precursor) is attached to a large non-polar carrier molecule called bactoprenol that allows it to be flipped across the cell membrane, since it is too polar to cross by itself
autolysins hydrolyse the glycosidic backbone in the existing peptidoglycan
transglycosylases connect the new unit using beta-1,4 glycosidic bonds, and transpeptidases form cross-links to connect the strands (this is the step inhibited by penicillin)
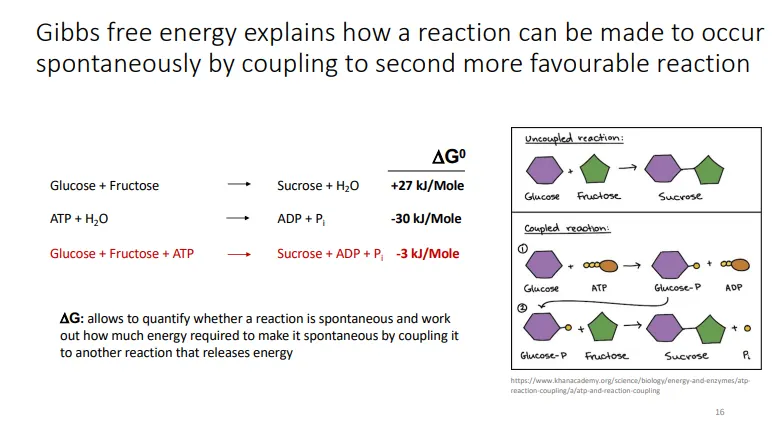
how does the organisation of life not defy the second law of thermodynamics?
the second law: the total entropy of a system and its surrounding increases for a spontaneous process
life seems to disobey this because small molecules spontaneously assemble into a highly organised state (decrease in entropy)
however, local entropy can spontaneously decrease as long as the total entropy of the system increases
life is possible because its biochemical processes:
increase the entropy of the environment by taking up highly ordered forms of energy and releasing less ordered forms (and using the energy released in catabolism to produce ATP)
couple non-spontaneous reactions with more energetically favourable ones (primarily the hydrolysis of ATP)
the energy released can be used to produce macromolecules and decrease the entropy within the cell
what is entropy?
entropy is a measure of disorder in a system
S = k x lnW, where:
S = entropy, k = constant and W = the number of possible microstates for the macrostate of the system

why is ATP the universal energy currency?
can be produced from adenine nucleotides
adenine is the only base that doesn’t include oxygen (so can be produced easily in anaerobic conditions)
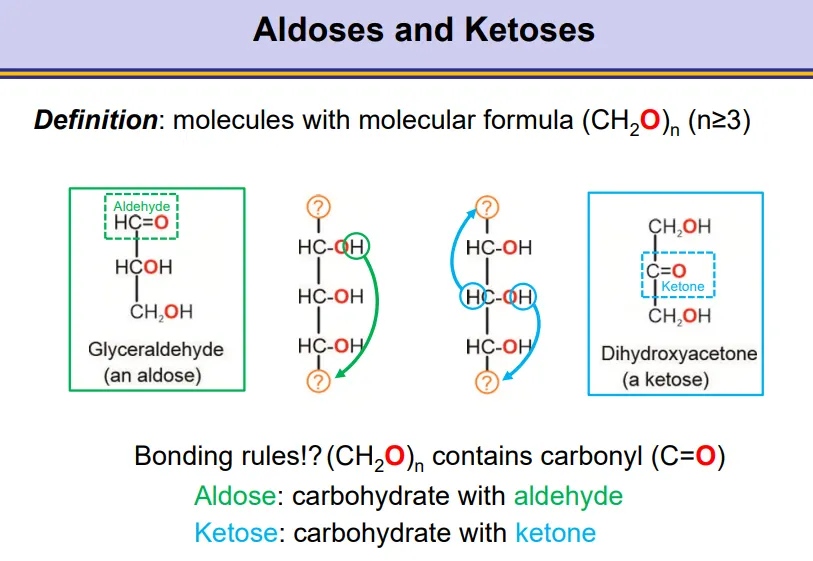
what are aldoses and ketoses?
aldoses are carbohydrates containing an aldehyde group
ketoses are carbohydrates containing a ketone group
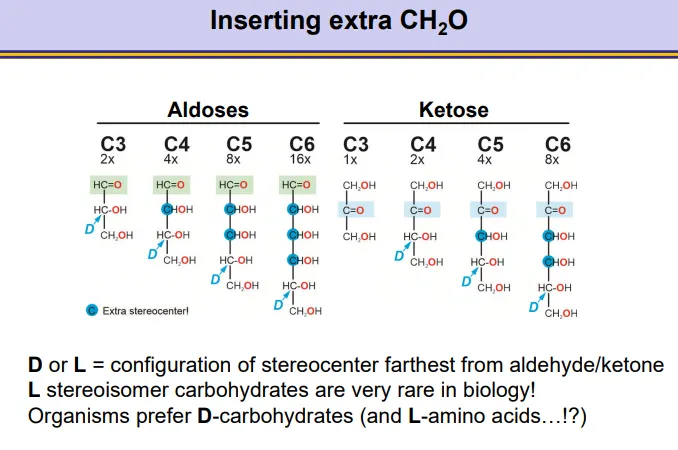
how can carbohydrates have stereocentres and how do these change with added carbons?
C3 (triose) aldoses have one stereocentre, and gain one more with each added CHOH
the same goes for ketoses from C4 (tetrose) onwards
the D or L enantiomer is defined by the stereocentre furthest away from the carbonyl group because this will have the secondary hydroxyl group that is used in cyclization
only D enantiomers are used in nature
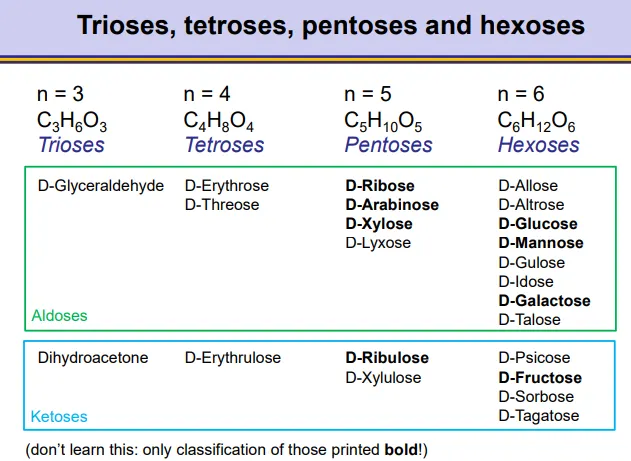
what kind of carbohydrates are galactose, mannose, xylose, fructose, ribulose, arabinose, glucose and ribose? (all D forms)
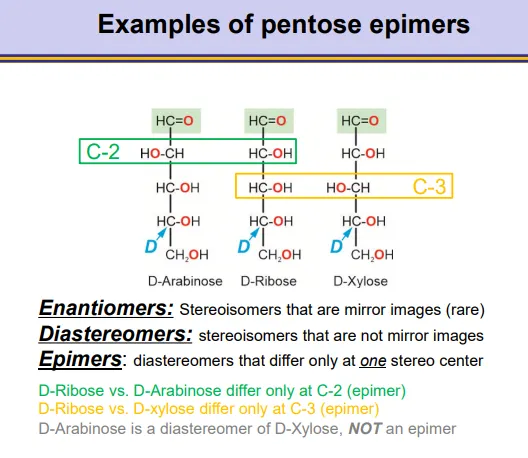
what are the different kinds of stereoisomers in carbohydrates?
enantiomers- complete mirror images, for all stereocentres (rare)
diaestereomers- stereoisomers that aren’t mirror images, these include:
epimers- diastereomers that differ at only one stereocentre and are fixed
anomers- diastereomers that differ only at the anomeric carbon (alpha vs beta, dynamic)
conformational isomers- chair vs boat, envelope
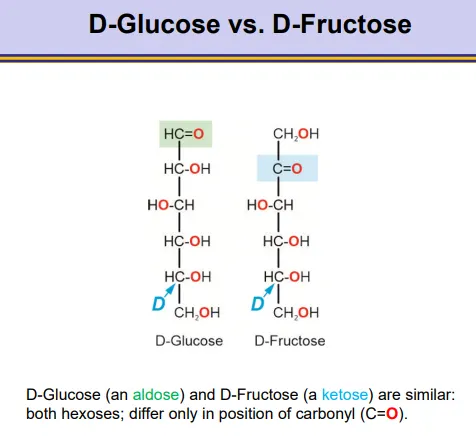
what is the difference between D-glucose and D-fructose?
they are very similar, just with the carbonyl in a different position
glucose is an aldose, and fructose is a ketose
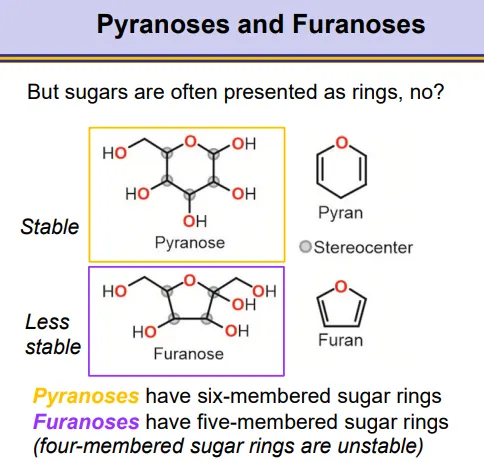
what are pyranoses and furanoses?
pyranoses and furanoses are sugar ring structures
pyranoses have six-membered rings
furanoses have five-membered rings, which are less stable
four-membered rings are unstable, as well as any bigger than six
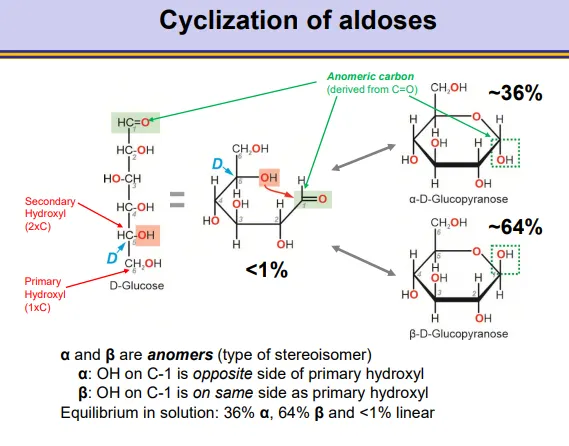
how does intramolecular cyclization occur in aldoses and how does this result in stereoisomers?
a hydroxyl from the bottom of the molecule can attack the carbonyl at the top of the molecule nucleophilically to produce pyranoses and furanoses
in aldoses this produces a hemiacetal around the anomeric (previously carbonyl) carbon
this occurs preferentially by a secondary hydroxyl, because they are more reactive than primary hydroxyls
when the D-enantiomer is used, the CH2OH will point up
this will produce anomers, depending on whether the hydroxyl on the anomeric carbon points down (alpha-opposite the CH2OH) or up (beta- same side as the CH2OH)
the ring closing is reversible and the ring can close into either form, so an equilibrium exists between the two forms
in water the equilibrium leans towards the beta anomer (hydroxyl groups in equatorial position in the pyranose chair conformation)
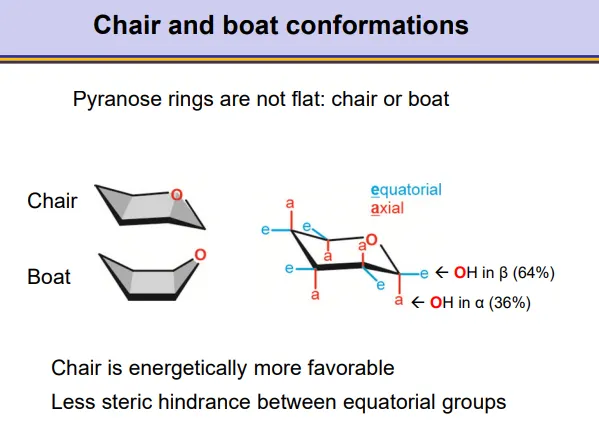
how do pyranose rings have different conformations?
pyranose rings take up a chair or boat conformation
the chair is more energetically favourable
this is why beta anomers are favoured, because there is less steric hindrance when the C1-hydroxyl group occupies the equatorial position (on roughly the same plane as the carbons)
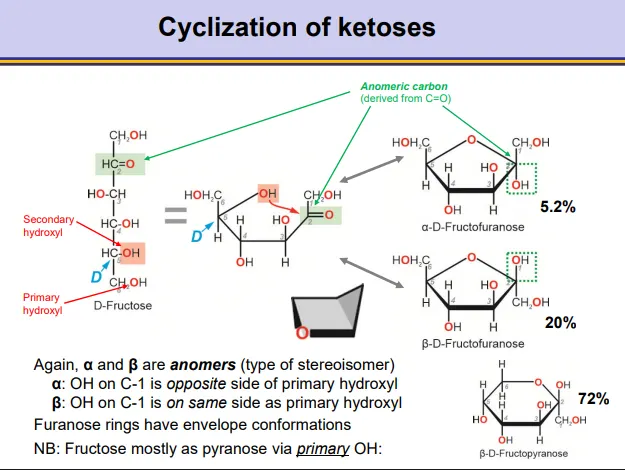
how does intramolecular cyclization occur in ketoses and how does this result in stereoisomers?
a hydroxyl from the bottom of the molecule can attack the carbonyl near the top of the molecule nucleophilically to produce pyranoses and furanoses
in ketoses this produces a hemiketal around the anomeric (previously carbonyl) carbon
this normally occurs preferentially by a secondary hydroxyl, because they are more reactive than primary hydroxyls
when the D-enantiomer is used, the CH2OH will point up
this will produce anomers, depending on whether the hydroxyl on the anomeric carbon points down (alpha-opposite the CH2OH) or up (beta- same side as the CH2OH)
the ring closing is reversible and the ring can close into either form, so an equilibrium exists between the two forms
in water the equilibrium leans towards the beta anomer (favourable in envelope furanose conformation)
note: fructose prefers to form a pyranose ring via its primary OH
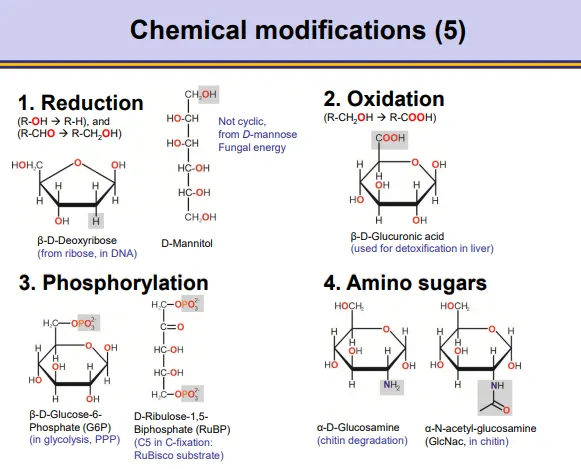
how can carbohydrates be chemically modified?
reduction- remove oxygen/add hydrogen eg. ribose → deoxyribose
oxidation- add oxygen
phosphorylation- add phosphate to replace hydroxyl groups
amination- add amino/acetylate group etc
glycosidic bond formation- in acidic conditions, the anomeric carbon can react with alcohols
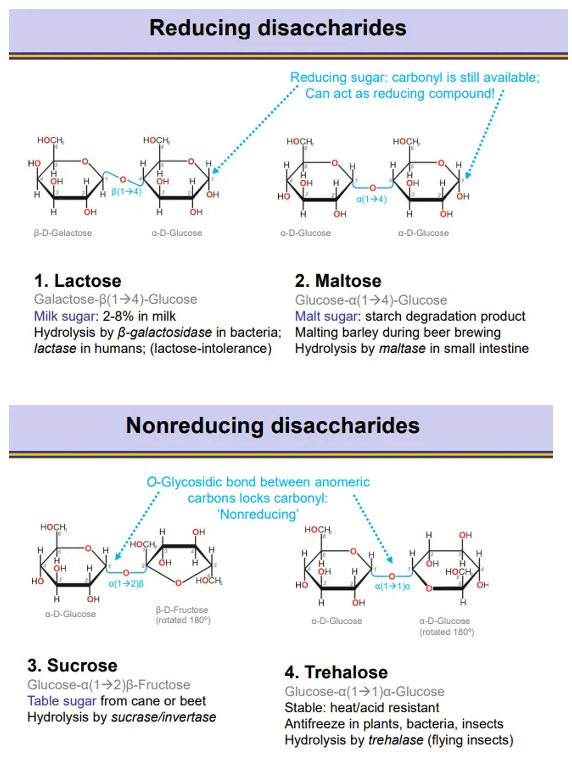
what are reducing and non-reducing disaccharides and what are the two examples of each?
in reducing disaccharides, there is always one anomeric carbon still available, so the ring can still open and the carbonyl can be used as a reducing agent (becomes COOH)
in non-reducing disaccharides the gycosidic bonds are flipped 180 degrees so both anomeric carbons are involved, so the carbonyl isn’t available as a reducing agent
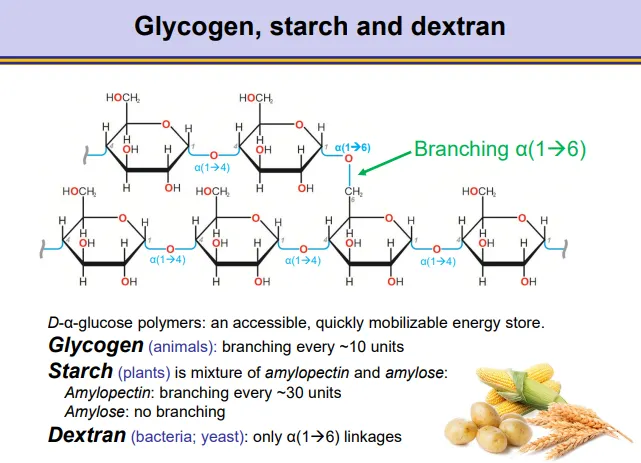
what are the structures of dextran, glycogen and starch?
these are all accessible energy stores based on alpha glucose
dextran (bacteria + yeast)- only 1,6 glycosidic bonds
glycogen (animals)- mostly 1,4 bonds, with 1,6 branching every ~10 units
starch (plants), a mixture of:
amylopectin- mostly 1,4 bonds with branching every ~30 units
amylose- only 1,4 bonds
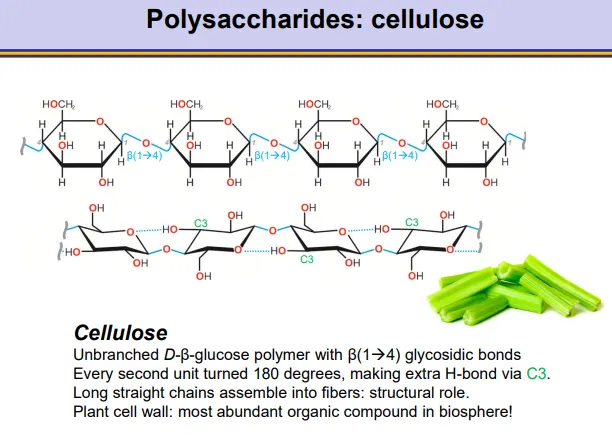
what is the structure of cellulose?
cellulose makes up plant cell walls:
unbranched beta glucose polymer with beta 1,4 glycosidic bonds
alternating units are flipped 180 degrees so that a hydrogen bond can form between the C3 and the oxygen in the ring
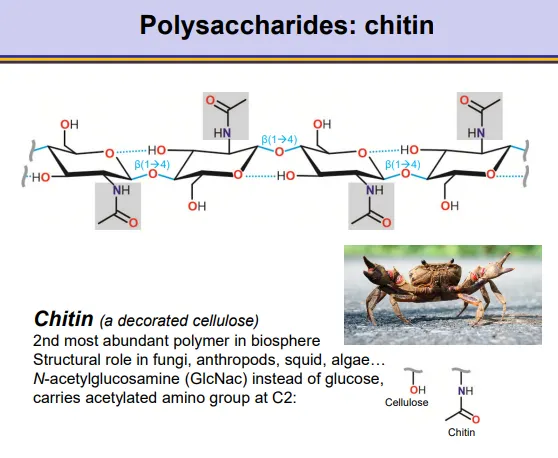
what is the structure of chitin?
chitin is a decorated cellulose, found in fungal cell walls, insects and other organisms:
the beta glucose monomers are chemically modified by amination to add an acetylate group to the C2
this produces C-acetylglucosamine monomers
unbranched, beta 1,4 glycosidic bonds
alternating units are flipped 180 degrees so that a hydrogen bond can form between the C3 and the oxygen in the ring