Looks like no one added any tags here yet for you.
same allele freq means what for genotype distribution (50% A 50% B)
can have diff genotype distribution (50% AA, 50% BB, or 25% AA, 50% AB, 25% AB)
population definition (biologist, pop. genetics, & genetic)
biologists: aggregates of similar individuals
population geneticists: limited to describe groups of sexual forms that associate for reproduction as well as for ecological reasons
*genetically: defined as a genetic population that spatially and temporally restricts a group of conspecific interbreeding individuals → this particular use of the term permits a population to persist through time while the indiv. in it are dying and being replaced on a continual basis
genetic population is maintains continuity over time because of…
it might change because of…
reproductive interconnections between generations, and it is endowed with spatial unity due to interbreeding among its members
a population may grow in size or become smaller due to migration of indiv. in/out or by alterations in birth and death rates. it may fuse with other populations and may become extinct either by total elimination or by the complete dispersion of its members
gene exchange between members & its importance in population genetics
(gene pool definition and role as a result of meiosis—describe process)
in pop. genetics, populations are considered populations of genes rather than individuals.
in this light, all genetic information distributed among the individuals of an interbreeding group collectively forms a gene pool.
such a gene pool is temporarily dispersed and held as a set of particular genotypes in each generation (keeping in mind that zygotes—genotypes—that develop into individuals of one generation result from the union of gametes produced by the process of meiosis. alleles are sorted, shuffled, and distributed to gametes by the process of meiosis. gametes combine during fertilization to produce a new set of genotypes).
consequently, a reconstituted gene pool is produced with each generation (assuming non overlapping generations)
how is the gene pool concept best represented
by small isolated or semi isolated colonies whose members can interbreed in a manner approximating panmixia (random mating/mating pairs are a random association of genotypes.)
“panmictic unit” and “local mendelian population” (definition and other, more modern names)
have been applied to such populations (small isolated/semi-isolated colonies used to describe a gene pool).
the common designation for such populations can also be called “genetic population” or “deme”
metapopulation → subpopulation relationships
meta= population within a population (large area, thus pops cluster in certain smaller regions)
some gene flow, but less than within sub population
enough gene flow is occurring to maintain a connection between subpopulations
change in gene frequencies & how measured
“evolution”
can be measured at a single loci or at multiple within the genome of the organism being studied
how much do species differ genetically?
(& genetic similarity def.)
depends on organism and the genes surveyed;
genetic similarity is a phenetic measurement that provides an estimate of the genetic identity among two species that are being compared.
genetic identity of 1, <1, vs 0
1: if the two populations have the same alleles in the same freq. at all loci sampled
<1: if the gene freq. of any alleles at any loci vary. less similar the sets of allelic freq. at each locus, the lower the value
0: if the two species share no alleles at any sampled locus in common
phenetic vs phylogenetic and homologous vs nonhomologous
phen: overall similarity (wings)
phylo: evolutionary relationships (mammal bone structure); can be homologous structures or non homologous →
homo: identical trait acquired by shared evolutionary event/ same distant ancestor
nonhomo: paralogs (developed separately for same function)
hardy weinberg equilibrium general overview/ main notionsHW
in the absence of evolutionary processes (destabilizing forces), the allelic freq. within a pop. will remain constant over time. this behavior of population level genetics can best be studied through applications of HWE and an understanding of assumptions for this model;
no evolution means allele freq. will not alter after first generation bs no mechanisms for change or destabilize genotypes (no change in gene freq over time)
HWE
note that several pops can have the same gene freq. but diff genotypic freq.
however, after one gen of random mating, if all assumptions are met, the the pop will achieve HW expected genotypic freq.
HW assumptions (stabilizing forces vs destabilizing and why it acts as destab.)
any violation will cause gene freq. to shift over time and thus allow evo processes to occur
large populations meaning in HW
infinitely large (not realistic). assumed bc sampling error from one gen to the next changes gene freq. and this probability of sampling error inc as pop. size dec.
as pop depart from being infinitely large, the relative contributions of genetic drift and the founder effect to the evolutionary process becomes much more significant
HW assumption of random mating/panmixia
so that a given indv. has numerically/statistically equal probability of mating with any other indiv. of opposite sex in the population.
thus, any tendencies toward inbreeding or self fertilization or any tendency toward some form of assortative mating will change gene freq. in the pop from one gen to the next
mutation & HW assumption
mutation changes gene freq. unless the rate of mutation to a new allele occurs at the same rate that mutation back to the original allele occurs (small probability of actually happening).
commonly some sort of mutation pressure is observed to exist so that mutation is preferentially one way—that is while some back mutations may occur, it most typically occurs in one direction
migration and HW
gene flow changes gene freq in both the source and the recipient populations (even one indiv. causes small change). immigration and emigration are both important factors
those indiv. that do migrate and introduce their genes into a new pop are unlikely to be fully representative of the donor gene pool or the recipient gene pool.
how powerful a force gene flow is in changing gene freq. in the pop dep directly on how many indiv. are dispersing and how large the population is that they are dispersing into. the smaller the pop that migrates into and the larger the number of migrants there are, the greater the overall power of this process to change gene freq. and thus cause evo change
natural selection and HW
NS results in the differential perpetuation of genotypes or in other words the “unequal transmission of genes to subsequent generations by diff genotypes”
if NS does confer some sort of incredible inc in fitness to carriers of a certain allele at any given locus, clearly this could cause rapid changes in gene freq.
the greater the degree of selection for this novel allele, the greater the rate of change
while NS is a force operating in the overall evo process and is thought to provide the overall directional force in the evo process, the other HW assumptions can also be influential in the evolution of new species
stabilizing selection
when selection favors indiv. in the center of the normal distribution of phenotypes.
in this instance, the phenotypes at the center become more strongly represented while the tails (bell shaped) becomes truncated
selection
some alleles confer some sort of benefit with it thus alleles appear more in subsequent generations over others without the benefit.
selection for and selection against are one in the same (against is a trait removed from pool bc transmitted at lower rate, thus there is another allele that is selected for and has an inc in freq.)
directional selection
when selection favors one end of the range of a normal distribution of phenotypes
the overall effect is to shift to either the right or the left in favor of those phenotypes that are being selected for
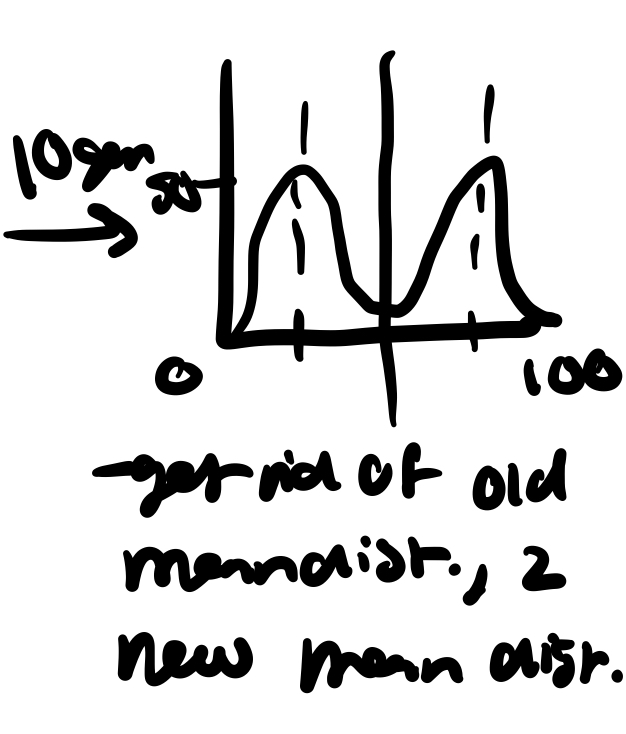
disruptive selection
what was the most frequent phenotype may no longer be adapted to new conditions and the freq. of phenotypes around the former distributional mean dec and the two new peaks of high freq. develop with the overall result of broadening the dist. base
this may occur if a species is splitting and moving into two or more new habitats
genetic drift (& three conditions)
the most unrealistic of HW assumptions are that pop are infinitely large and mating is random; while panmixia within a species assures that no genes will be lost from gen to gen, any deviation from this will necessarily introduce some form of sampling bias that will change the gene freq. over time
genetic drift is the alteration of gene freq. through sampling error. 3 general conditions:
1) continuous drift—when a pop remains small gen after gen, sampling error is a continuous process and can greatly alter gene freq.
2) intermittent drift—a large pop may occasionally shrink to a small size. at this point in time tremendous sampling error may be introduced
3) founder effect—when a few indiv. or only one reproductively competent indiv. founds a new pop that indiv(s). is seldom representative of the pop it came from and thus the new pop may have a set of gene freq. that are tremendously different from the parental pop
“place and powers” date, name, and authors
1979, by allen r place and dennis a powers
genetic variation and relative catalytic efficiencies: lactate dehydrogenase B allozymes of fundulus heteroclitus
place and powers main ideas and concepts
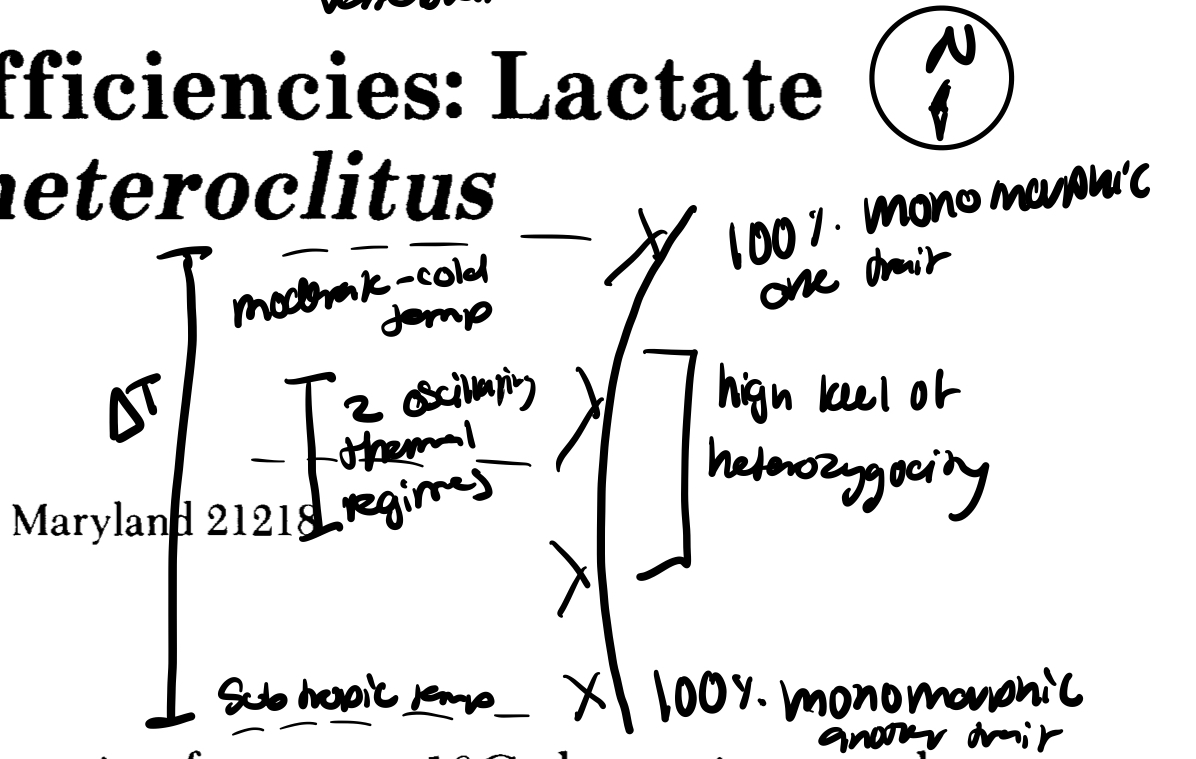
evidence suggest that there is a preference toward one homozygote>other, no advantage to being heterozygous (at either temp), and consistency with the hyp. of NS
naturalistic perspective (majority of variety is selectively neutral) vs selectivist perspective (selection operates to maintain protein polymorphism like in sickle cell hemoglobia where heterozygotes get malaria resistance)
used fish bc endothermic so their body temp reflects env.
got lucky with data set from region perfect conditions
what prevents speciation in two sister species at diff temps? → gene flow bc no profound barriers to prevent mating thus no geographic isolation that is needed for vertebrate speciation
polymorphism
the condition of having more than a single allele at a locus
thought of at population level or above, hence, does the gene locus being considered code for only a single allelic product within a population/species, or does it code for more than a single allelic product?
poly vs monomorphic work better in certain env more than others if NS at work (not necessarily selection for one, but exclusion of rest of traits if one trait is advantageous)
heterozygosity
discovered after the invention of DNA studying techniques
in a diploid organism, this is the condition of having two alternative alleles present at a gene locus
produce two diff types of gametes with respect to this single locus
two ways heterozygosity is measured
population geneticists often estimate the average number of loci within a population that are heterozygous
the average number of individuals that are heterozygous for a particular gene locus is another way to look at data
these enable one to study diff aspects of the dynamics of the population under the study
plants and polymorphism
polymorph at 50% of their loci with a mean of 1.96 alleles per locus
gymnosperms—wind pollinators like pine trees—have highest level, more than monocot angiosperms—flowering plants—which is higher than dicot angiosperm—fruit plants (who are most selective with mating and less offspring that dec chance of fertilization and thus less reproduction events to allow NS and evo to occur)
why? bc gymnosperms are long lived, have wide geographic ranges, are widely outcrossed due to their wind pollination
animal polymorphism
marine invertebrates > invertebrates > vertebrates (reasons similar to plants; far reaching, non specific mating strategies are favored for polymorph)
vert. have lowest levels of polymorph and heterozygosity with mean values of 17.3% and 4.9% respectively
*water mimics wind so why marine invert. so similar to gymnosperm rep.
does variability in the substrate that a particular enzyme acts upon account for higher levels of polymorphism? is there a heterozygote advantage?
homozygotes at a locus encoding a protein only make one form of that protein, heterozygotes at a locus encoding a protein make two diff forms of that protein; advantages for hetero could be 2 diff forms achieve levels of catalysis at diff temps, thus creature can expand enviro. if needed/encouraged
if that protein is an enzyme, having two forms could allow catalysis in a wider array of substrates and thus be better off, or allows one to act on two or more forms of a substrate that is routinely encountered in the env. it lives in, then there may be a heterozygote advantage as a selective advantage
however, much of genetic variation observed seems to have no apparent advantage, that is, in many cases, one form on an enzyme functions the same as another allelic form at the same locus. thus, much of the variation we do observe seems to be selectively neutral
DNA
all the genetic information in a cell is contained in the sequence of four subunits along the molecule
deoxyribonucleic acid or DNA for short.
forms a template from which the cell manufactures proteins
molecular subunits of DNA
guanine, adenine, thymine, and cytosine
GATC
proteins consist of chains of up to how many diff types of AA; and how many nucleotides it takes to encode an AA
20, each encoded by a sequence of three nucleotides
codon
coding triplet
redundancy of codons and wobble position
(how relates to substitution mutation)
because 4 codons taken three at a time yield 64 different sequences and there are only 20 amino acids to encode, there is considerable redundancy in the genetic code
mostly seen in the wobble position —> 3rd codon, bc first two determine AA that gets coded for
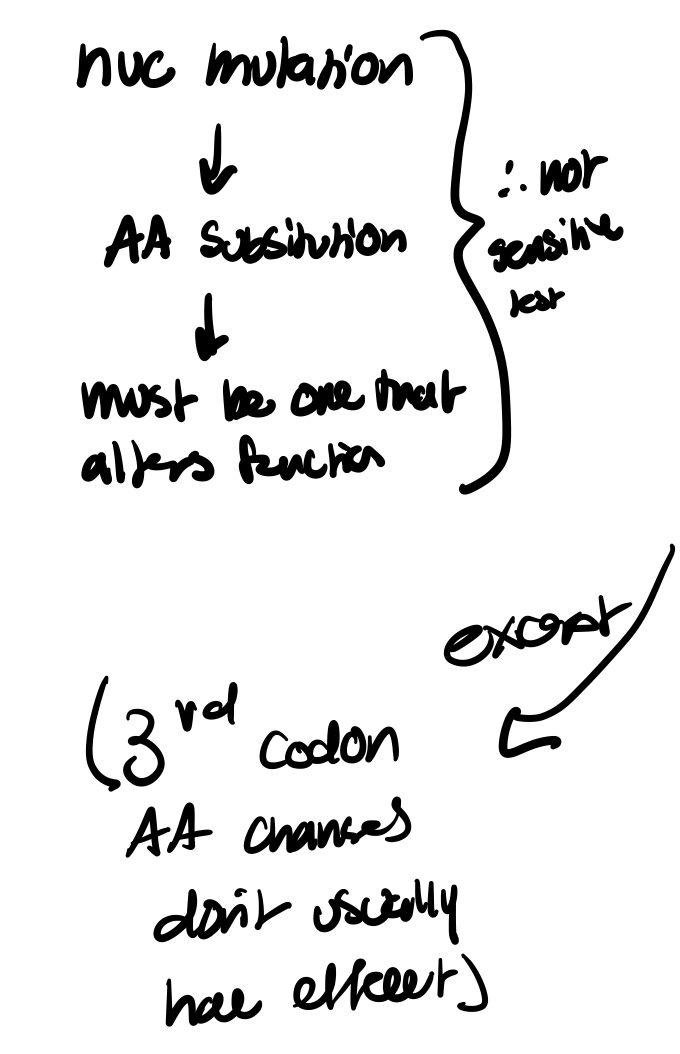
thus, a change in one of the nucleotides in a DNA codon may alter the AA that is encoded for or not dep on what position is changed
(substitution mutation: dep is resultant change causes 3 nucleotide codon to actually code for a diff AA)
substitution mutations; do all mutations have an effect on fitness?
form one class of changes to the DNA molecule that are referred to collectively as mutations
mutations can result from random copying error when the genetic material replicates during cell division, by the action of certain chemical agents, or due to the effects of ionizing radiation.
keep in mind that many mutations that occur have neither deleterous nor advantageous effects on the organism in which they occur bc they occur in non-coding portions of the DNA seq.
third codon
wobble position
in protein coding seq, changes at the third codon position are observed w greater freq than at either of the first two positions. presumably bc these changes are less likely to alter the AA being encoded
deleterious effects of mutations
most mutations are felt to exert a deleterious effect through the following reason:
Over the course of evolutionary history, natural selection has operated to weed out most of the deleterious alleles, leaving behind only those alleles that suit the organism to its environment.
Any new variant allele resulting from a mutation is likely to disrupt the well-tuned interaction between the organism and its environment.
New proteins produced by mutant genes might have different properties from the protein that the mutant is based on. Some of these new mutants might be beneficial, but most are thought to be
deleterious.
That is, they are felt to most likely decrease the fitness of the individual in some way.
HW test
only validates HWEQ for specific gene at specific locus at specific time, still could have mutation, selection, etc. on other loci
more likely to occur in larger pop
why HW? (3 reasons)
foundational principle that pop. genetics is based — all measurable factors derived from HW
can do multiple loci and multiple alleles and still use HW
1) mathematically explains why we don’t lose variation from one gen to the next (mendelian inheritance is verified)
2) HW proportions are null hypothesis; 5 assumptions (no evo). if proved invalid, can then perform tests on which of 5 hyp. broken—test acts like springboard for why pop out of HW (selection, mutation, etc)
heterozygote freq. usually too low in nature
3) 5 stages in general model of genetic, HW simplifies
why mutation theory that most mutations have a deleterious effect is not reasonable
neutral or neg alleles can be drug along w selection of pos (selected for) alleles due to linkage (close prox)
presupposes that everything ab organism was shaped by NS; not everything is selected for bc some alleles drug along (wisdom teeth) if no overt deleterious effect
genotype
the sum total of all the genetic characteristics that determine the structure and functioning of the organism
phenotype
the physical expression of the genotype in the organism
it is a combination of morphological and physiological characters
each gene codes for what & two general functions of this term
a particular protein
these proteins may be structural in nature (may act in physical composition of the organism) or they may form hormones or enzymes critical to functioning of the organism
alleles when mutated
different sequences of genetic subunits in a particular gene that are caused by mutations
what can have an impact on an organism’s phenotype
many alleles or different forms of a given gene can impact an organism phenotype (morpho or physio)
that is they may code for some protein product that lends the organism a diff appearance
diff alleles or forms of a gene can impact an organism how, beyond phenotype changes?
alternatively, they may code for an enzyme that contains a slightly diff composition of AA—while this causes no visible change, the implications of this sort of variation can be profound.
perhaps one allele of the gene codes for an enzyme that has a much greater affinity for a certain substrate in an enzymatic reaction. such allelic products, or alternative forms of e gene can confer some sort of selective advantage on an organism
diploid organisms and their somatic cell genes
diploid organisms have two copies of all genes that are not sex linked
homozygote vs heterozygote & dominant vs recessive genes
if two copies of a gene in an organism are identical (homozygote), if they are diff, meaning that an individual has two diff alleles of a gene (heterozygote)
in heterozygous indiv, the transcription on a DNA seq to RNA and translation of that RNA into protein will produce two diff gene products (more complicated if more than a monomeric protein)
if one of these gene products completely masks the expression of the other gene product, we refer to the allele coded for that the protein is dominant. the alternative allele is recessive. alternatively, these two alleles may result in an intermediate phenotype
what deviations of HW mean
it is important to know equilibrium proportions of homozygotes and heterozygotes in HWEQ, bc deviations from the expected proportions given measured levels of allele freq. are indicative of the effects of selection, nonrandom mating, or other factors that influence the genetic makeup of the population.
diff phenotypes interactions in the envir.
diff phenotypes interact with the envi. in slightly diff ways. this can lead to variation in survival and fecundity among indiv. in a pop.
fecundity & the measure of fitness (and in sense of NS and evo)
the rate at which an indiv. produces offspring
the rate of reproduction of a given phenotype is a measure of fitness
when differences in fitness among indiv have a genetic basis, those alleles that confer higher fitness will tend to reproduce faster and take over the population.
those phenotypes having the highest fitness are said to be “selected”, and the change in genotype freq. resulting from NS is referred to as evolution
research on selection in English Peppered Moths by HD Kettlewell
Briefly, there are two color morphs of the English peppered moth, one dark and one light. The light morph had historically been the dominant form with relatively fewer of the dark morphs. It was discovered that in forested areas near centers of industry, there seemed to be a greater number of the dark morphs relative to the light morphs, and in forested areas that were remote from industrial centers, the light morph was still predominant.
Through a fairly elegant series of experiments the English physician H.D. Kettlewell, turned naturalist,
determined that the darker morphology conferred a selective advantage on the moth in industrial areas where the trees had darker trunks. Industrial pollution turned normally light-colored tree bark darker which would make a darker morph of the moth harder to see by predators.
In the non-industrial areas, the lighter moths had a selective advantage as tree trunks were lighter in color due to an absence of excessive industrial pollutants.
The agents of the selection in this instance were insectivorous birds that routinely searched the trunks of trees to prey on these moths. Those morphs easiest to observe were taken as prey items in greater numbers. Hence the morph that was more cryptic in a given area was selected for and subsequently had higher fitness.
Thus, the increase of the dark morphs in polluted environments.
correlated responses to selection (interdep of traits)
mice ex.
evolutionary responses often involve traits other than those being selected. both development and function tend to integrate the parts of organisms bringing about an interdep. of certain phenotypic traits.
for ex) there are several traits that are highly integrated into discrete clusters that involve osteology in mice. therefore in mice, if you were to develop a breeding program to select for an increase in body weight, you would along with the desired inc in body weight see changes in skull width and tail length
can’t separate corresponding traits bc either close on chrm are of DNA and thus don’t recombine and/or is linked, or bc change in one would be harmful and unsupported by other body parts—the body is an integrated system that works in perfect tandem to meet needs (selective constraints).
NS in relation to interdependent traits
NS operates through a system of phenotypic compromise that is tempered by the genetic makeup of the organism and the physical linkage of the traits that are being selected
inbreeding and why bad
mating among close relatives (siblings, or in plants the process of “selfing” aka autozygous)
most indiv. are heterozygous for at least some rare, deleterious recessive gene across the entire genome
this means that the freq of the allele is low in the pop at large (rare) that the results of the gene product of this allele are negative in some way (deleterious) and that in a heterozygous condition the gene is not expressed (recessive)
if full sib cross (same gene combo), some offspring will have trait and there is an inc in proportion of homozygous recessive for rare deleterious trait in overall pop
in depth why no inbreeding in context or recessive, rare, deleterious trait
Hence the negative consequences of having this rare allele is only expressed in the homozygous condition.
Most individuals are felt to have at least some of these genes.
If an individual that carried the recessive trait in
the heterozygous condition were to mate with an individual from the population at large, the probability would be fairly low that that individual carried the same recessive trait, this of course depends on the frequency of the allele.
The results of such a union would be that half of their progeny would be heterozygous for the trait, and
the other half would not carry the allele at all.
None of the progeny would however be at a disadvantage by simply carrying the trait. If on the other hand a carrier of the trait reproduced by “selfing”, as you see in some plants, one fourth of its offspring would be homozygous for the deleterious allele and would suffer a loss in fitness as a consequence.
Mating’s between siblings would produce similar results only at a reduced level. This is why inbreeding is bad.
how most species reduce freq. of inbreeding (hermaphroditic plants and animals)
Most species have a variety of mechanisms to reduce the frequency of inbreeding.
Among these are dispersal of progeny, recognition of close relatives, and negative assortative mating (preferential mating among phenotypically unlike partners).
Hermaphroditic species of plants (those in which individuals bear both male and female sexual organs) have additional mechanisms to prevent “selfing”, such as self-incompatibility, temporal separation of male and female function, and elaborate flower structures designed to make “selfing” difficult.
genetic changes in small populations (genetic drift definition)
in small pop, the freq of alleles and genotypes may change bc of random variations in birth and death rates (statistical anomalies) due to change events—even in the absence of selection—this is called genetic drift
founder events
can also have profound impact upon the genetic structure of a pop
these events occur when a few indiv. from a larger pop colonize a new habitat. the significance is that these new pop have a reduced sample of total genetic variation found in the parents
hence, drift and inbreeding (dec genetic variation) can result in fixation of alleles that were at a relatively reduced freq in the population as a whole
this can and does lead to considerable population subdivision and heterogeneity
inbreeding (& consanguineous mating def)
when individuals are more likely to mate with relatives than with nonrelatives (nonrandom—no panmixia aka the HW assumption).
this is consanguineous mating (“of the same blood”).
autozygous
more precisely, when the alleles carried by gametes that unite to form a zygote are identical by descent. such alleles that are the result of replication of a single allele in some ancestral population are said to be autozygous
overall effect of inbreeding
is to inc (in comparison to HWEQ) homozygosity in a population
it is important to note that homozygotes is increased for ALL gene loci (entire genome is undergoing inbreeding, not a specific loci selected)
as the freq of homozygotes inc, the freq of heterozygotes must dec
inbreeding from DL Hartl 1987; a primer of population genetics; Sinauer (with comparison to assortative mating)
mating between relatives is inbreeding
like positive assortative mating (when individuals that are similar with respect to some set of traits mate with one another), inbreeding inc the homozygosity in a pop
unlike assortative mating, which affects only those genes on which mate selection is based (and genes in linkage disequilibrium with them), inbreeding affects all genes
in humans, the closest degree of inbreeding that commonly occurs in most societies is first cousin mating, but many plants regularly undergo self-fertilization
genotype freq with inbreeding
the principle effect of inbreeding in a pop is to inc the freq of homozygous genotypes at the expense of the freq of heterozygous genotypes
this effect can most easily be seen in the case of repeated self-fert.
display of how homozygosity inc and heterozygosity dec in a completely selfing plant pop
true or false, inbreeding by itself does not change allele freq.
true (freq of p and q stay same, genotype freq like AA or Aa will change)
the self fert. example illustrates the general principle that inbreeding by itself does not change allele freq
the constancy of allele freq with inbreeding occurs only when all genotypes have an equal likelihood of survival and reproduction, which is to say that no natural selection occurs,
if selection does occur, then allele freq can change with inbreeding or even with random mating
quantitative effects of inbreeding
in terms of the reduction in heterozygosity that occurs
we can measure the amount of inbreeding by comparing the actual proportion of heterozygous genotypes in a pop with the proportion of heterozygous genotypes that would occur with random mating (HWEQ)
actual freq of heterozygous genotypes and random-mating het. genotypes eq to find the inbreeding coefficient:
when F= 0 (no inbreeding/random mating), the genotype freq. are…
when F=1 (complete inbreeding), the population consists…
in the familiar HW proportions of p2, 2pq, and q2
entirely of AA and aa homozygotes in the freq p and q respectively
*assumption being tested via HW is panmixia, does H=2pq equal the null, Ho
hierarchical F statistics
Fst, Fis, and Fit;
population subdivisions
analog for inbreeding bc can compare inbreeding coefficients of fractional reduction pq to the “grand” pq of the rest of a species
measures how separated pop are and how close to speciation
what other than inbreeding can lead to homogenization of pops
gene flow if unopposed by other factors
the extent to which there is a reduction in gene flow between pop of a species (pop subdivision) can be quantified
Fst equation
Fst = (Ht - Hs) / Ht
Ht
what heterozygosity would be if all subpopulations were pooled together and mated randomly (2pq for total population)
heterozygosity of total pop
2pq is expected heterozygote freq. in a population
Hs
2pq for the subpopulation
Fst
measuring the reduction of heterozygosity
between 0 and 1
fixation index that measures population division among subpopulations (measurement of population subdivision)
either much gene flow or diversion
natural populations are distributed in pockets (subpopulations) with much gene flow
=> what these statements mean in terms of Fst equation
all same species distributed across a landscape they ca be found, in clusters, thus don’t get completely even dist. of indiv. bc not every area is optimal for living based on resource availability
clusters closer to where good habitat
higher likelihood of reproducing with closer individuals than other sub pops thus not panmictic
need some gene flow to keep species together cohesively bc mutations are a spontaneous and continuous process in every indiv. otherwise we get speciation (Fst closer to 1)
panmixia
total random mating between subpopulations
how evolution occurs in 3 terms
1-isolation
2-lack of gene flow
3-constant mutations occuring
thus result in two different populations
if Fst = 0 (Hs=Ht)
no population subdivision
HW heterozygosity is similar to heterozygosity frequency in subpop
same species
if Fst = 1
then populations are fixed—freq 100%—for alternative alleles at the gene locus analyzed
no heterozygotes (Hs = 0)
essentially two diff populations or even diff species—no gene flow
homogenization of subpop can have two results:
if gene flow, then remain same species
or sub pop divides as first step toward speciation
what keeps two pop from diverging?
All you need is 1 individual to have successful gene flow between populations to keep them from diverging.
With no gene flow, populations will continue to accumulate mutations independently of one another. Mutations will introduce new alleles into the populations and without gene flow, the populations will diverge and may become different species.
This is a process of speciation without the necessity for natural selection. Purely random and mechanical processes to drive speciation.
genetic drift
the alteration of allele frequencies (diff expressions of a gene) through sampling error
it can be a very powerful evolutionary force under three general sorts of conditions
occurring all the time in nature
Two assumptions of the Hardy-Weinberg model are that populations are infinitely large and mating is random.
In nature these two assumptions are rarely met. why?
While panmixia within a species assures that no alleles will be lost from generation to generation, any deviation from this will necessarily introduce some form of sampling bias that will change the allele frequencies over time.
where is sampling error from
error from not randomly mating
genetic sampling less than 100%
say pop 1 has 100000 and the next gen only 500 reproduce. some alleles will be lost because only a subset of the population participated in reproduction
continous drift
when a pop remains small generation after generation
sampling error is a continuous process and can greatly alter allele freq
intermittent drift
“bottleneck”
a large population may occasionally shrink to a small size
at this point in time tremendous sampling error may be introduced
at bottlenecking event, must of diversity is lost and cannot be regained
founder effect
when a few indiv or only one reproductively competent individual founds a new population, that individual(s) is seldom representative of the pop it came from and thus a new pop may have a set of allele freq that are tremendously diff from the parental pop
ex) since hawaiian islands are formed by volcanoes, no species were formed in situ thus all living forms came from elsewhere/mainland as migrants to the area
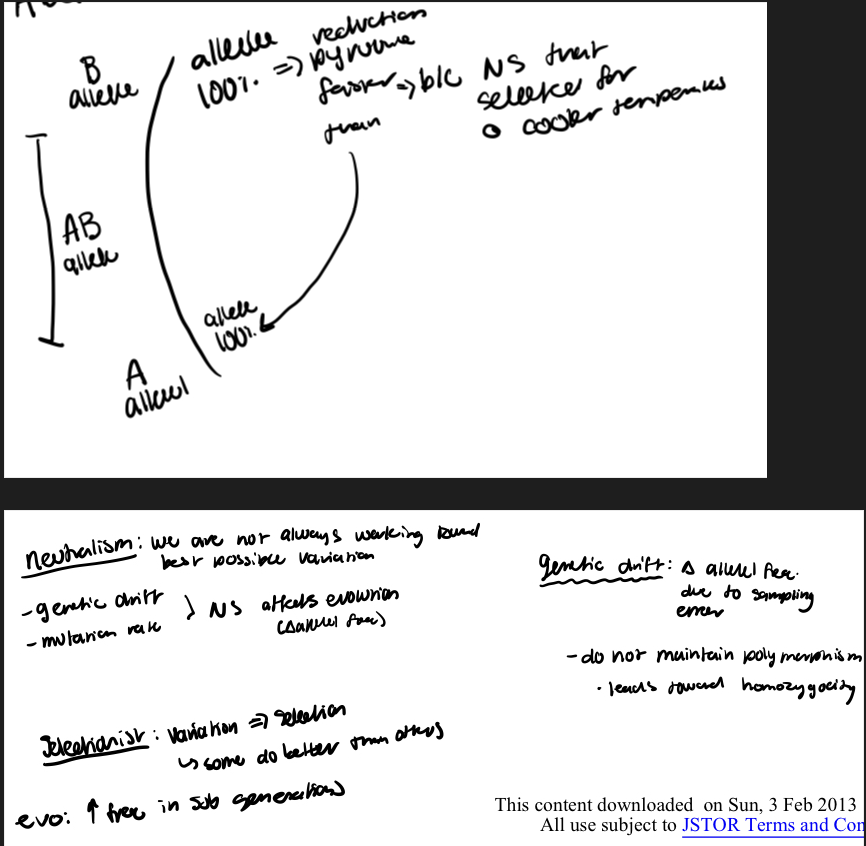
neutral selectionist controversy summary (two sides, then shift with new data and two possible outcomes)
1. Until the 1960’s one of the chief issues in population genetics was whether populations contained much or little genetic variation. —didn’t know bc we didn’t have the technique to directly quantify genetic variation (allele freq variation). this changed with protein gel electrophoresis
2. Now this has been answered and the question has shifted to whether most of the variation is selectively neutral and thus irrelevant to selection (does not allow the population to respond to different selective regimes) or whether the genetic variants do differ in fitness and thus constitute the raw material for adaptation to new selective regimes. —we can’t have evolution or change in characteristics if not selected for by some force
3. The evidence that evolutionary rates are highest in non-transcribed DNA and for relatively functionless polypeptides argues strongly for considerable genetic drift at the molecular level; but the question then becomes, how much molecular variation and evolution does is explain. —found more genetic variation than expected
4. The observation that enzymes are as variable in the haploid bacterium E. coli as in diploid organisms indicates that heterozygote advantage (powers and place demonstrated: heterozygotes with two protein types function better than homozygous colleagues) is not a general explanation for allozyme polymorphism (does not rule out other types of selection though). —most early research was only on functional enzymes that “did” something; thus only ones with outcomes were tested; most AA don’t make a functional difference in proteins thus, no functional advantage
5. In contrast the nature of polymorphism at some loci (such as ADH in Drosophilia) suggests that selection is operating.
neutral theory: non-darwinian evolution summary; selection as a force of evolution limitations
A cornerstone of the theory of evolution by gradual change is that the rate of evolution is absolutely limited by the amount of genetic variation in the evolving population (Hubby and Lewontin, 1966).
Thus even if selection is the force that drives the establishment of overall morphology and function for an organism (this is an assumption that we will allow in this instance), selection can only operate on genetic differences that already exist in the species. That is selection only acts to weed out the non-functional or less fit variants that exist, it does not drive the formation of these variant forms.
*selection does not make new variants, it is a thoughtless process with blind mistakes
protein electrophoresis overview
Estimates of the amounts and kinds of genetic variation that exist in populations is one of the primary foci of evolutionary studies.
Understanding the details of this genetic variation was greatly facilitated in the 1960’s with the advent of protein electrophoresis.
This is a technique that allows the visualization of many proteins within an organism in a manner that allows a gross estimate of the amino acid changes that exist within various forms of the protein commonly referred to as allelomorphs (diff morphotypes, of a protein=diff alleles).
Protein electrophoresis takes advantage of the fact that most mutations in structural genes should result in a substitution, deletion, or addition of at least one amino acid in the polypeptide produced by the gene.
In a certain proportion of the cases this mutation in the gene will cause changes in the net electrical charge on the polypeptide and the charge on the enzyme or protein of which it is a part.
Keep in mind however that many mutations in the DNA sequence do not cause changes in the Amino Acid being coded for and thus the estimates generated with this technique are generally considered to be underestimates of the true level of genetic variability within a population at the DNA level.
—inevitably going to underestimate AA changes bc won’t pick up alterations with no functional difference, like in the wobble position
usefulness of protein electrophoresis
This technique proved very useful in determining the amount of genetic variation within populations, and the level of heterozygosity within individuals.
It is still very widely used by a number of investigators for evolutionary investigations in areas including the estimation of phylogenetic history and the determination of population structure and intraspecific heterogeneity (determining the level of within species genetic variability).
Lewontin and Hubby electrophoretic study 1966
In one early electrophoretic study (Lewontin and Hubby, 1966) it was reported that within five populations of Drosophila studied at 18 different gene loci, that the average population was polymorphic at 30% of its loci and that on average, 18% of the loci within any one individual were heterozygous.
Given what was stated above regarding these estimates as necessarily underestimating the true levels of genetic diversity present, this immediately posed a problem for evolutionary studies.
In essence, this problem was a question of how this level of variation could persist in natural populations, and what sort of mechanisms influenced this genetic diversity.
Three possible explanations were offered to account for these observed levels of genetic variation.
Lewontin and Hubby electrophoretic study 1966: “alleles are irrelevant to NS” possible explanation to account for observed levels of genetic variation that were higher than expected
what would this look like/mean if it were true
what is counterargument
main takeaway
Alleles are irrelevant in natural selection--essentially this argument is that all the alleles that exist are adaptively equivalent to one another. While genetic drift would tend to drive populations toward homozygosity, recurrent mutation and migration would tend to counteract such a trend.
If this were the case local populations would tend to have one allele or the other due to their equivalence.
This is not what was observed. Instead researchers saw a high frequency of heterozygotes in the natural populations studied which tended to counter this notion. This data does lend support to the selective neutrality hypothesis.
However, this is not true of all natural populations. Many of which are completely fixed (all individuals homozygous) for a given allele. Neighboring populations in many instances are fixed for alternate alleles.
Therefore it is not quite as cut and dry as the author of many textbooks make it out to be.
Lewontin and Hubby electrophoretic study 1966: “mutation rates are high” possible explanation to account for observed levels of genetic variation that were higher than expected
what would this look like/mean if it were true
Mutation rates are high--selection eliminates alternative alleles, but mutation rates are so high that the alleles are restored.
This suggests that (again, alleles are close to selective neutrality).
Problem: Mutation rates would have to be unusually high for this to occur. Much higher than we have observed.
Lewontin and Hubby electrophoretic study 1966: “selection favors heterozygotes” possible explanation to account for observed levels of genetic variation that were higher than expected
2 assumptions needed to be true
Selection favors heterozygotes--requires two assumptions:
1. Weak selection against homozygotes.
2. Heterozygotes have some sort of adaptive superiority.
—possible like in powers and place
neutral theory of molecular evolution (logic mechanistic pathway, and null hyp)
the electrophoretic data gathered in many early studies set the stage for this theory
This theory does not deny the existence of natural selection but, asserts that at the molecular level most polymorphisms and mutations that become common are adaptively neutral. —evolution w/out NS=> most mutations selectively neutral (non-lethal).
Point mutations occur within protein encoding genes on a constant basis (not at a constant rate).
As such the genome of any organism becomes saturated with any changes that are not in essence lethal to the organism (as these would be weeded out by natural selection).
Thus in portions of a protein coding locus you might see a great number of amino acid substitutions (perhaps in areas that are not encoding the portion of the polypeptide forming an active site for instance) whereas in other portions of the same locus, there may be areas that are highly conservative across a wide array of taxa (perhaps in areas that do code for the active domain, or encode an amino acid sequence that is critical to folding, net charge, etc.).
The null hypothesis then (for the neutral theory) is that evolutionary change at the molecular level occurs by random mutation and random drift. —not saying NS doesn’t occur, but saying at a molecular level, most changes are due to random mutation and drift