Ecology - Midterm 2
1/97
Earn XP
Description and Tags
Lectures 8-13 and Chapters 8, 10, 11
Name | Mastery | Learn | Test | Matching | Spaced |
|---|
No study sessions yet.
98 Terms
there are three common forms of exploitation
predation: predatory/prey
parasitism: parasites or pathogens/hosts
herbivory: herbivores/plants
we can adapt the lotka-volterra model: the prey equation
change in prey population density = exponential population growth of prey - number of prey lost to exploiters

we can adopt the lotka volterra model: the exploiters equation
change in exploiters population density = births possible form prey consumption - natural deaths of exploiters
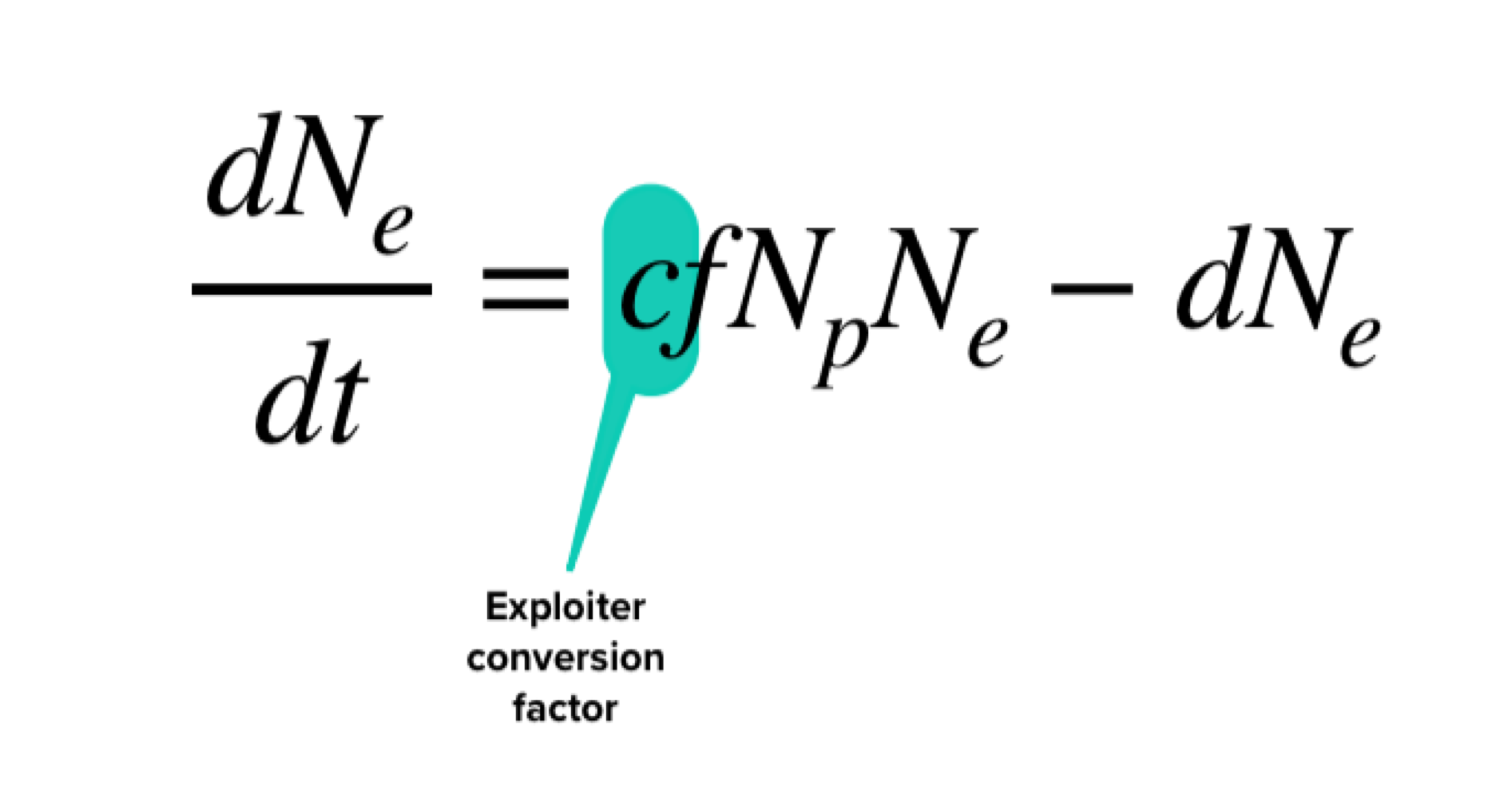
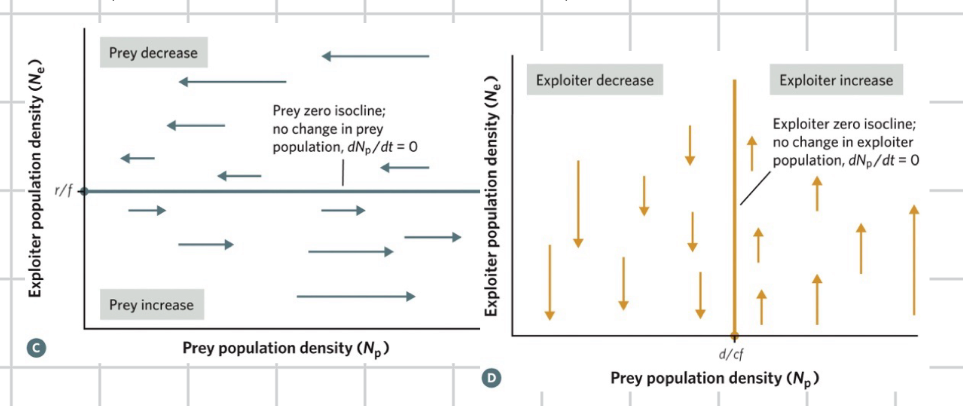
each species phase plant graph is divided into two areas
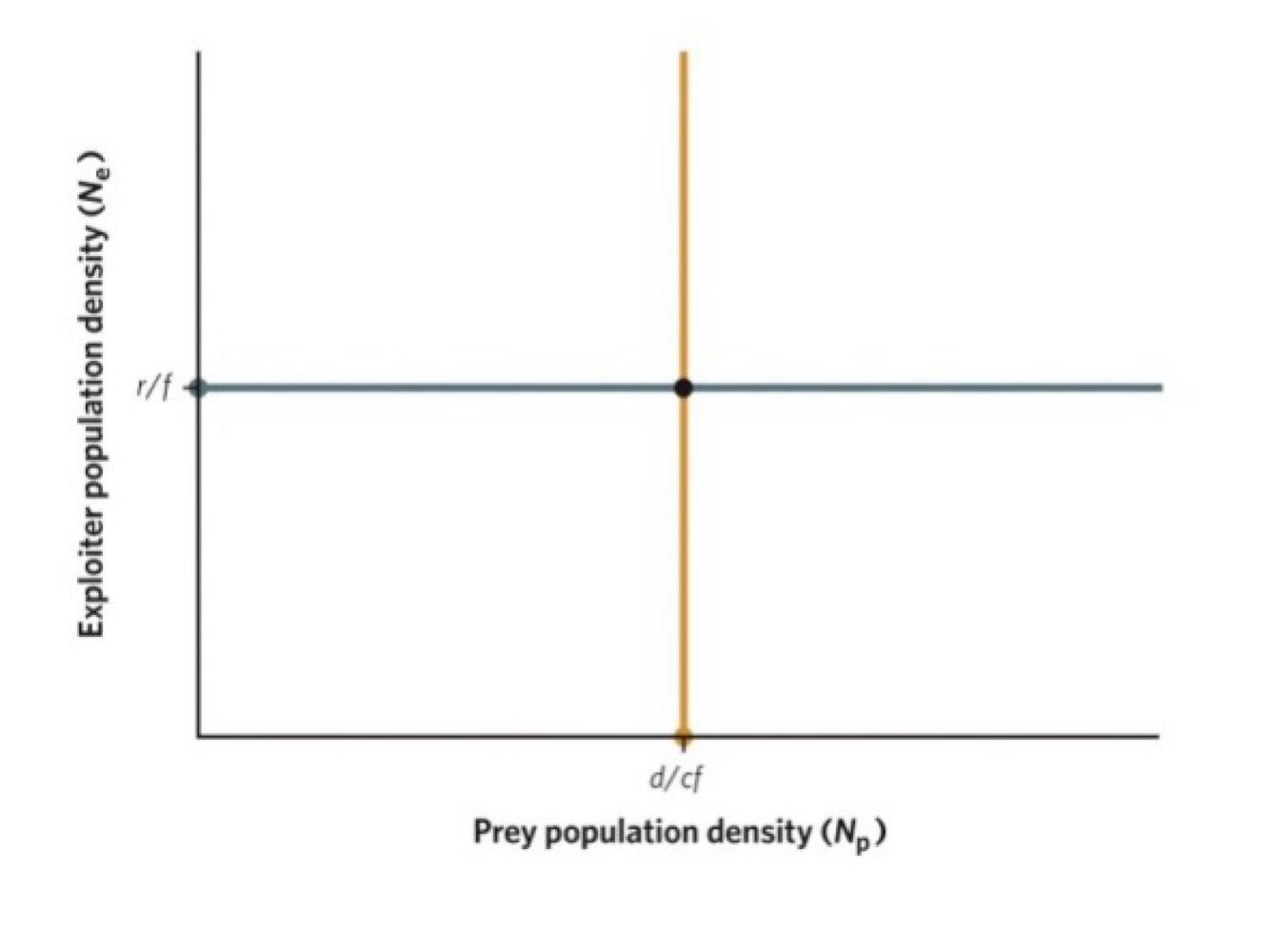
population cycle around a neutrally stable equilibrium point
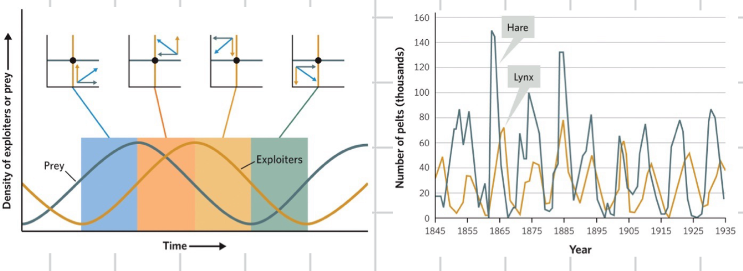
the cycling explains the offset cycling of population
making the model more realistic: prey density dependence
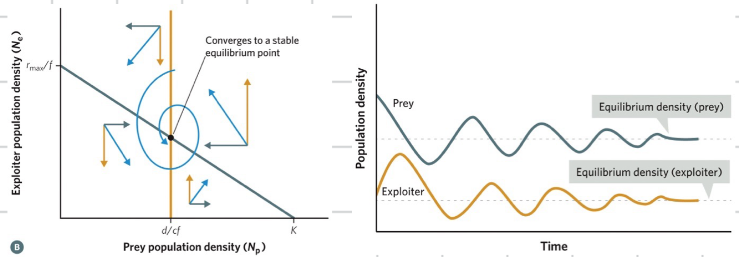
making the model more realistic: exploiters functional response
exploration model outcomes depends on location of intersection of isoclines
all living organisms have to eat
for species that photo synthesize energy intake involves the capture of sunlight
for the rest, the need for energy requires the capture, consumption and assimilation of energy contained in another organisms tissues
exploitation
an interaction in which the individuals of one species increase in fitness by consuming individuals (or part of individuals) of another species
comes in many forms, but the common factor among them is that individuals of one species benefit by consuming individuals of one species benefit by consuming individuals of another species
predation
the most familiar form in which a predator kills and consumes prey
generally they will consume many different prey individuals in its lifetime
this form of exploitation is so common and familiar that predation is often used as a synonym for exploitation
parasitism
a parasite obtains energy from their hosts by attacking the hosts body or organs and consuming tissues or fluids, often without killing the host
this category is quite broad and includes disease organisms (or pathogens) and parasites that can live either inside (endoparasites) or outside their hosts (ectoparasites)
parasites are gnerally smaller then their hosts and each parasite tend to use only one host for its whole life, or at least for one whole stage of its life
herbivory
exploiters consume plants, with either plants or select parts of them
herbivores do not typically kill the plant they eat
like predators, through, they can eat many individuals plants in their lifetime
terminological issues
some resources are not biotic and therefore are not ingested
event if we focused on ingested, biotic resources, there are many types of exploitation
the common thread for a lot of them is that the consumer individual receives a fitness benefit, and the consumed individuals experience a decrease in fitness
for this to hold true the prey must be alive when the consumer encounters them
detritivore
any organism that feeds on dead plant and animal parts (detritus) as its primary source of energetic nutrition
what ecologists focus on
they focus on whether the consumed resource is alive (or apart of a living organism) and not on its reproductive status
this assumption works pretty well in practice when defining what counts as prey because many organisms in the wild do not live much past reproductive age
possible outcomes of exploitation
the (+, -) schematic works as a simple conceptual model and suggests patterns that we might expect to see as a result of exploitation
with interspecific competition, this was easier to conceptualize because the negative effects worked in both direction and lead to declines in population abundance for both species
if we apply the tilman model, we can imagine that the exploiter species should drive the prey abundance down to a consistently low level, but only if there is a consistent resupply of prey
exploiter prey cycle
a complex cycle between predators and prey in which the density of abundance of each species alters the density or abundances of the other species
fluctuation in weather or other external variables may lead to large changes in the abundance of either population but without cycle or imminent risk of extinction
four possible outcomes of exploitation
prey extinction that leads to exploiter extinction
exploiter extinction with surviving prey
exploiter prey cycles and stable coexistence
stable coexistence
the prey equation
the model for the prey needs an intrsinsic rate of population growth that reflects natural birth and death rates without the influence of an exploiter series
(change in prey population density) = (exponential growth of prey) - (number of prey lost to exploiters)
population growth equations always explore population growth over some chunk of time
the left hand side of the equation represents the change in the popualtion in the smallest possible slice of time
the speed of a chemical reaction
it is dependent on the concentration of chemicals, because a higher concentration means the chemical elements are more likely to randomly bump into each other
capture rate (f)
functions as a conversion factor and helps to make the units consistent because we are interested in the proportion of encounters that are successful
the actual number of prey captured will equal the capture rate multiplied by the number of encounters between exploiters and prey
putting the prey equation all together
the term dNp/dt is the instantaneous change in prey population density over continuous time
r is the intrinsic rate of increase for the prey population
Np is the prey population density
Ne is the exploiter population density
f represents the capture rate
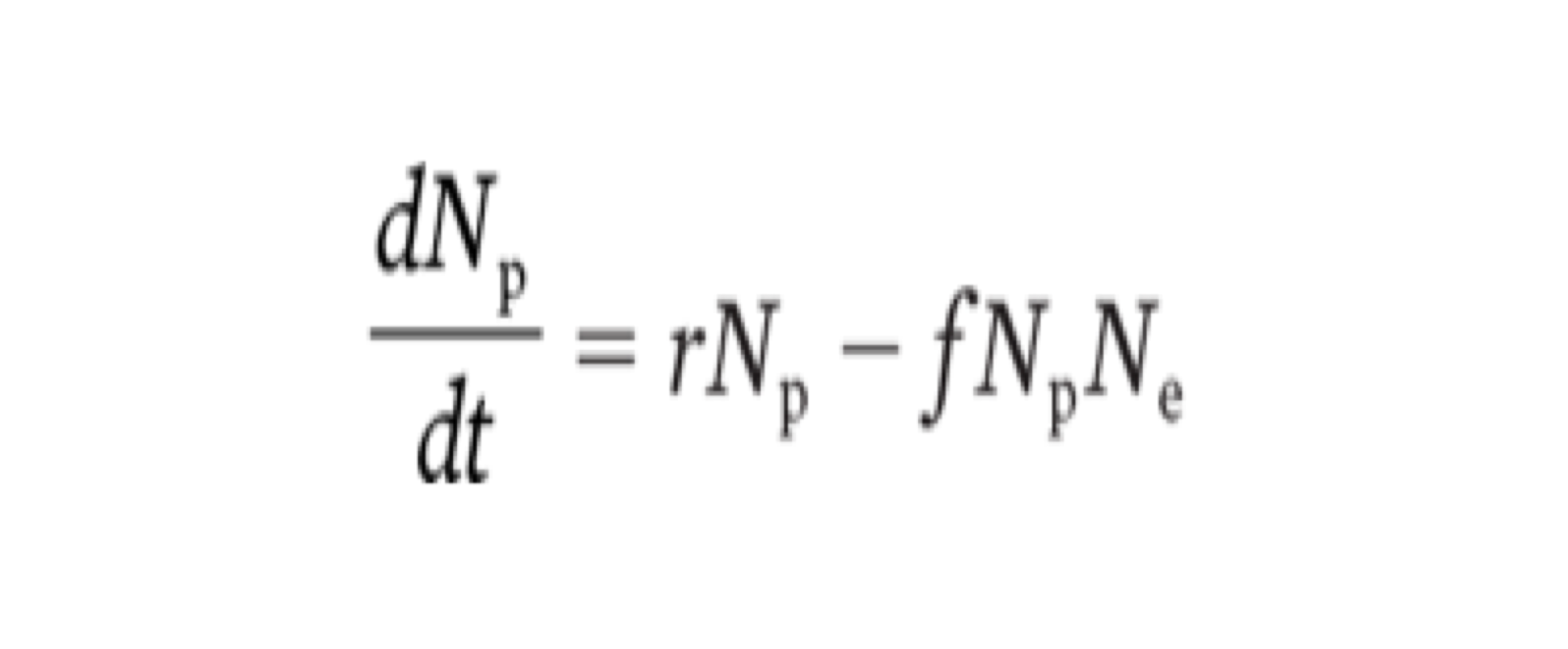
the exploiter equation
change in exploiter population density = births possible from prey consumption - natural death of exploiters
exploiter conversion factor (c)
a term that translates consumed prey into an expected number of offspring for a focal species
we can determine the births in the exploiter equation by multiplying the number of captured prey individuals from the prey equation by c to convert consumed prey into expected broths of exploiters
putting the exploiter equation together
the cfNp term is like the instantaneous per capita brith rate, b, but it is not constant the way b was
the birth rate is dependent on the density of prey
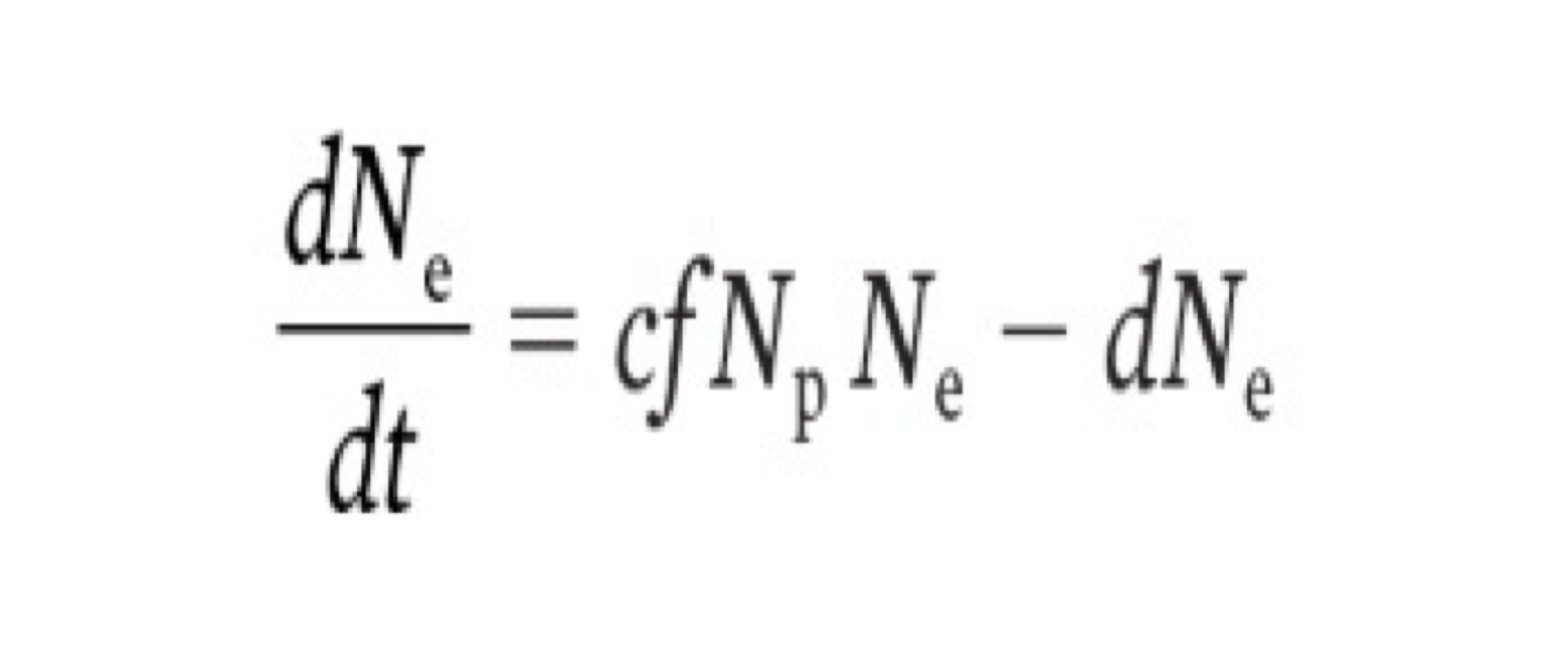
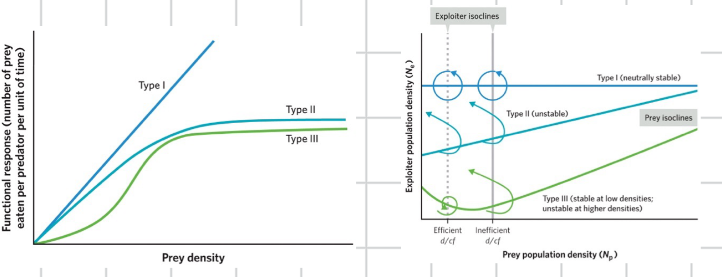
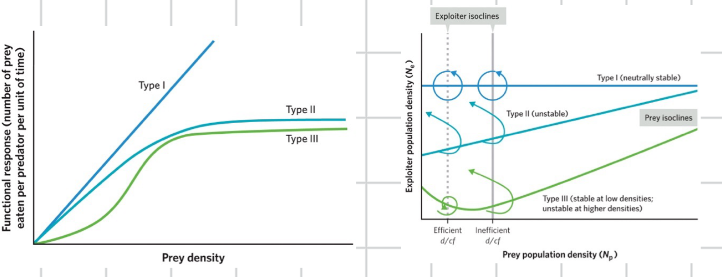
exploiter functional response
another unrealistic assumption built into the lotka volterra model is that exploiters will consume a constant fraction of the prey they will encounter, regardless of increases in the prey density
we call the factor that describes this consumption rate the functional response
as prey density goes up the number of prey eaten per predator goes up linearly
type I functional response
this type is mathematically easy, but organisms in the real world cannot increase their consumption endlessly
leads to stability and cycles
type II functional response
this natural constraint means that exploiter consumption levels off at high densities of prey, in the real world
much more common than type I
divides the graph into four uneven sections
type III
acknowledges that consumption may be slow when prey population densities are very low
if the exploiter isoclines intersects the prey isocline at a place where the prey isoclines has a negative slope, we see damped oscillations and stability
allee effects
sometimes individuals will group together, or aggregates (meaning that their average density across a large space may be low, but their local density is high)
not only mammals aggregate but many species of fish, birds and insects
allee effect
a pattern, observed in some species, in which there is a fitness decline as density declines
allee effects may also result from increased reproduction as density increase as from very low levels
because allee effects lead to lower predation rates, adding them to the lotka volterra model should influence the outcomes
a phase plan analysis of allee effects suggest three outcomes
stability, extinction, or cycles
allee effects can be strong or weak, so the specific shape of the prey isocline can change, but it should always have a positive slope to show inverse density dependence at low prey density, and then.a negative slope as the allee effect fades and density dependence takes over
if the predator isocline crosses the prey isocline to the right of the hump in the prey isocline, population cycle decreases in amplitude and spiral toward stability
predation
carnivores are organisms that eat animal tissue
herbivory
the feeding of individuals of one species on individual, or parts of individuals, of a primary producer species
the primary producers are usually terrestrial plants, but ecologists often also refer to organisms that eat seaweeds and algae as herbivores
what herbivores typically consume
they typically only consume portions of individuals
we still see the same general range of population dynamics for herbivores and their plant “prey” partially because chomping on a plant may mean that the plants offspring or reproductive structures have been removed, directly reducing the plants reproductive fitness
generalists herbivores
although predators of animals may have preferences as to which prey species they eat, they rarely refuse available prey organisms and their diet tends to include a diverse array of prey
specialists
many consume a very narrow range of species, often sticking to just one family or genus
some may feed exclusively on the leaves, fruit, pollen, or flowers of a plant, whereas others may eat only the seeds, bark, roots, sap or woody parts
parasitism
falls within the definition of exploitation as an interaction in which one species gains in fitness and another loses fitness
an organism that obtains it nutrition form one, or a very few host individuals
it causes harm to the host but often does not kill the host
it is a bit like herbivore in that the fitness cost to host are often lower than they are in predator prey interactions
macroparasites
are large enough to be seen with the naked eye
they may live on or in a host, but they often reproduce with infective stages outside the host
animal macroparasites live on the blood or in the gut cavities of their hosts
in plants, macroparasites often penetrate the hosts tissue in order to tap into the xylem and phloem
microparasites
tiny are often occur intracellary
they reproduce directly within their hosts, where can achieve very high densities
if it produces symptoms that are clearly harmful, the host is said to have a disease
parasitoids
insect exploiters that fall somewhere between specialist predators and parasites
some parasites have fairly simple life cycles, requiring only food to consume and a place to grow and reproduce
other parasites have extremely complex life
definitive host
a host that carries the reproductive stages of the
intermediate host
a host that supports the non adult form of a parasite
evolutionary outcomes of exploitation
species evolve defensive traits that lessen the fitness impacts of their exploiters, and exploiters evolve traits that allow them to overcome these defences (co evolution)
this is evolution by natural selection through reciprocal selective effects between two or more closely interacting species
exploiter prey convolution
an individual exploiter that successfully eats prey is more likely to survive and reproduces
a prey individual that successfully avoids being eaten by an exploiter is also more likely to survive and reproduces
life lunch concept
the evolutionary consequences for prey are more dire than for exploiters
the strong evolutionary pressure on the prey to avoid being consumed has produced an array of adaptations into three broad categories
categories of adaptations
primary
secondary
tertiary
primary defensive strategies
serve to prevent the exploiter form ever finding the prey
an effective way to avoid being eaten is not to be seen in the first place
many organisms use some form of camouflage to avoid being seen by their predators
this is often referred to as crypsis, or cryptic colouration
secondary prey defence strategies
often draw attention to the prey but also send a message that deters the exploiter from attacking
once an exploiter has spotted prey individual, different secondary defence strategies kick in to reduce the likelihood of capture or increase the change of escape once capture
one common strategy is for prey to announce themselves not as themselves but as something other than what they are
some secondary defence strategies involve sending a message to the exploiter to fool it into thinking the prey are dangerous, sometimes they are all dangerous
aposematic colouration
a colouration pattern found in prey organisms that signal dangers to a potential predator
also known as warning colouration
types of mimicry
mullerian and bateasian
mimicry
an adaptation that benefits individuals that look very similarly to individuals from another species
common form is a type of secondary defence strategies involve used by prey species with similar appearances to other species that advertise their toxicity through colouration and patterning
mullerian mimicry
a form of mimicry in which there is a selective advantage for both the mimicry and the model to closely resemble each other
reinforces a message of toxicity
bateasin mimicry
a form of inter species mimicry in which the mimic sends a false signal to interacting species
common in prey species that do not actually have a mechanisms to kill or harm the predator but show the same signal or pattern as a species that does
bastesian mimic dilute the message
tertiary defence strategies
in general it involves a way to escape or fight off an attack once it has already started
sometimes the line between a secondary and tertiary deference is blurry, as it is hard to tell whether the strategy prevents attack or interrupts the attack
the most common tertiary defence strategies involve physical defences that allow a species to escape or fight back
thanatosis
some species respond to attacks not by fleeing but by playing dead
aggregating behaviour
this can also be a tertiary defence strategy if the whole group takes defensive action to confuse the exploiter
false target
some prey will dangle a false target, such a a brightly coloured tail that is distracting and enticing, to draw the exploiters attention away form the preys head or vital organs
some prey will even sacrifice a body part to a predator to remove the threat of death
fighting back
fighting back can be an excellent tertiary defence strategy, particularly for big prey
evolutionary arms race
a constant cycle of trait evolution which two coevolved species respond to each others adaptation over time
in an evolutionary arms race some fraction of prey individuals will have a genetic variation that allows them to escape or avoid predations better than their peers in the same population
in most cases an arms race is not likely because most predators are generalists and switch to a new prey species if their current one becomes rare
red queen hypothesis
the evolutionary hypothesis that all species must constantly evolve in order to maintain their fitness in the face of a changing environment full of other evolving species
plant herbivore convolution
because individual plants are often simultaneously attacked by a suit of herbivores rather than just one, the adaptations developed by plants to avoid consumptions are of a somewhat different character than those of animals
constitutive defences
some defences are always present
inducible defences
defences that are only present when needed
toxins
are a common plant defence and are often described as secondary metabolites
origin chemicals that are typically not involved in the plants basic growth and development but can be essential to its survival
secondary plant chemicals
come in varieties of including alkaloids: such as caffeine, nicotine, morphine, and quinine, terpenoids, such as camphor, citronella, methanol, and THC
many of our drugs and pharamceuticals take advantage of these plant defences, as do quite a few of our much loved dietary produces
mechanical defences
these are to deter herbivores
plant defence strategies can add a level of complexity to herbivorous interactions by attracting enemies of their own enemies
hunting success
pack hunters: wolves 20% african wild dogs 60-90%
pursuit hunters: cheetah 40-50% dragonflies 95%
white clover (trifolium reopens)
produces cyanide to deter herbivore
the further from the city the more cyanide is produce
indirect effects
the overall effects of one species on a second species when both species interact with a third species or with a shared resource; the third species or shared resource mediates the interaction in such a way that the effect may even change in sign or direction
the idea this the one species affects a second species through a “change” in an intermediate, or mediating species
tropic interaction
an interaction between species in which the individuals of one species eat the individuals of the others
we can link tropic interactions in a food chain
the most common food chains differ slightly from this example by including a primary producer at the bottom level
primary producers
are autotrophs because they produce their energy form inorganic sources rather than from eating other biological organisms
autotrophs are generally plants and algae or bacteria and archaea
primary consumer
any orangish that mainly eats plants or other autotrophs is a primary consumer
secondary consumer
a primary consumer is then consumed by the secondary consumer
consumer trophic levels
each step in this chain represents a trophic level, meaning feeding or eating level
we can see the chain aspect of this basic level model if we link each level with the arrow to indicated the upward direction of the energy flow
aquatic ecosystems - food chains
four- even five- level food chinas are common, partially because of difference in the primary producers found in aquatic versus terrestrial ecosystem
terrestrial plants
growth with very stiff cell walls and are generally much larger and more fixed in place than the tiny mobile algae called phytoplankton that dominate in aquatic systems
a food chain for a lake or ocean will always include primary consumers that are zooplankton
trophic energy transfer efficiency
primary producers have access to to huge store of energy from the sun, but we know that this source is not unlimited
the availability and intensity of sunlight in different habitats, the availability of carbon dioxide or water, and the speed of chemical reactions all place physical limits on the rate of photosynthesis and therefore on primary producers energy acquisition
for heterotrophs energy acquisition is limited by the abundance of their food
this trophic connection suggest that the biomass of primary producers should affect the biomass of herbivores, which in turn should affect the biomass of predators
access to energy is just the first piece of this puzzle
low energy transfer efficiency across trophic levels places a physical limitation on biomass accumulation that holds for the vast majority of non photosynthetic organisms in the world
the inefficiency of energy transferred up a food chain means that a top predators body mass is derived form an incredibly large amount of photosynthetic production at the base of the food chain
trophic cascade
a pattern of altering effects on abundance and biomass that moves down a food web
evidence for bottom up control
one of the fundamental ways ecologists categorize lakes by trophic status, which they can determine by the clarity of the water
oligotrophic
lakes that have very clear, blue water generally support only a few primary producers and thus have a lower overall biomass per unit area
eutrophic
lakes that have more opaque, green water generally support an abundance of basal autotrophic species and thus have high biomass per unit area
most lakes
most lakes fall somewhere between these two extremes
ecologists do not necessarily consider eutrophication lakes to be in better conditions to oligotrophic lakes
bottom up control of a food chain plays into the categories of eutrophication and oligotrophic
in lakes with lots of biomass at the bottom of the trophic pyramids, and thus high rates of photosynthesis, there tend to be larger amounts of biomass at each trophic level, making the lakes eutrophic
in lakes with few phytoplankton and low rates of photosynthesis and productivity, we find very little standing biomass across all trophic levels, making the lakes oligotrophic
what limnologists have observed in lakes
they have observed that lakes can shift from oligotrophic to more eutrophic with the addition of nutrients to the water, usually in the form of nitrogen or phosphorus runoff form terrestrial sources
it appears that lake food chains can be controlled from the bottom up extr
complications to our food chain predictions
extreme levels of anthropogenic nutrient additions can complicate our food chain predications
when nutrient input gets too high, phytoplankton blooms can limit sunlights ability to penetrate water, leading to lower overall levels of photosynthesis
part of this effect also occurs because at such high abundances, the phytoplankton overwhelm the herbivores, who are unable to keep with eating them
this leads to masses of decomposition phytoplankton and the decomposition process takes up more oxygen
lower oxygen levels from reduced photosynthesis and increases decomposition reduce the performance of some of the consumer species, thereby further reducing consumption of phytoplankton and leading to lower abundance and biomass at high trophic levels
evidence for top down control
HSS suggests that predators and parasites of the herbivores control the herbivores populations and keep them at lower levels
other research has extended the HSS theory to suggest that control by competition or prediction alternates trophic levels on the food chains, with competition always working as the control at the trophic levels, followed by predation at the trophic levels below this, and then by competition at the next level down
food densities of primary produces, and chains with odd numbers of levels have low densitites of primary producers
herbivores in top down control
with herbivores population in check, the plants they eat proliferates until completion checks their growth, making the world green
the HSS argument has several problems
first would herbivores really eat plants down to tiny brown nubs?
plants have many defences against herbivores, suggesting that not all plant material is equal in terms of trnasferring energy to the primary consumers
food webs adding complexity
one of the biggest shortcomings of food chains that is that simple “chain” model leaves out so many possible feeding interaction
of course it is possible to generalize the chains to include many species at a single trophic level, but then the model ignores
omnivores in food webs
species that acquire energy by consuming across multiple trophic levels are called omnivores
omnivore is so common that species that feed only from one trophic level are singled out as trophic specialists
food chains omitted the concept of omnivore in the service of simplicity, but only ores are more the rule than the exception
some organisms eat such a varied diet that we could never assign them to a specific trophic level
what the food chain model also fails to recognize
the food chain model fails to recognize that organisms consume alternative foods at different stages of their lives
developmental changes are referred to as ontogenetic diet shifts
food chains overlook an essential concept of all of biology and ecology
everything that lives will eventually die, and the energy contained in once living creatures is recycled into energy for another creature to use
this death-to-life concept is pretty clear in the arrows that moves up the food chains
the return of their energy to the chain involves a whole suite of addio al species called detrivores and decomposers that feed on dead plant and animal parts called detritus
complex trophic model
these all led ecologists to a more complex trophic model called a food web
a graphical representation of the energetic feeding relationship amount of species that coexist in a community
examples of food webs
each line in a food web represents a trophic link, meaning an instance of consumption in this complex net of relationship
using a food web instead of a food chain address some of the concerned raise earlier about the inclusion of omnivory and ontogenetic diet shifts
food webs can include cannibalism or looping, in which animals eat their own kind
a food web that depicts dertivtores and decomposeres will have arrows point downward, representing energy flowing toward dertivores
moving beyond trophic levels