5) Analýza mtDNA
1/28
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
29 Terms
proč je mtDNA supr
Známe kompletní sekvenci, neprobíhá rekombinace, mění se pouze procesem mutace, vysoká mutační rychlost, maternální dědičnost
proč je maternální dědičnost supr
Mater. dědičnost výrazně snižuje počet studovaných předků
iii. Předávání mtDNA pouze přes ženskou linii (“pramáN Eva”)
Koalescence
splývání linií mtDNA
množství mutací a stáří mtDNA
mtDNA se liší, díky náhodným mutacím (jsou předávány v dané linii)
čím je generace starší, em více mutací se v ní nahromadí
Kde v mtDNA mutace hledáme:
• kódující region x kontrolní region HVR (regulace replikace mtDNA, HVSI a HVSII)
kódující region
lze analyzovat pomocí RFLP, specifickou sadou restrikčních enzymů
kontrolní region
variabilita studována přímým sekvencováním
Co v mtDNA zkoumáme:
některé mutace se předávají na potomky vždy spolu = haplotyp = určují tedy dědičnou linii
• některé haplotypy jsou si podobnější (sdílejí stejné mutace) a ukazují na společného příbuzného předka – tyto haplotypy se sdružují do tzv. haploskupin
Haplotyp
• sestava mutací
• podobnější haplotypy sdílejí příbuznější jedinci
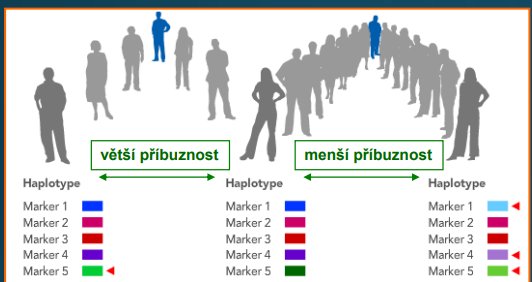
Haploskupina
• příbuzné haplotypy sdílejí stejné nebo podobné sestavy mutací
• tvoří haploskupinu
• mají svůj geografický původ (vzhledem k původnímu předkovi)
hledání společného předka
na základě haplotypů a haploskupin jsme schopni určit, ke které linii daní jedinci náleží (hledáme příbuznost na základě rozdílů a podobností) = identifikace společného předka
již víme - selekčně neutrální mutace – v čase se hromadí konstantní rychlostí = molekulární hodiny
podle počtu těchto mutací lze také odvodit stáří sledované linie, předka
problém
– stanovit podobnost mezi jedinci nemusí být úplně jednoduché -> vycházíme z mnoha změn na různých místech a také u většího počtu jedinců -> více sekvencí = nárůst geometrické řady počtu možných stromů -> stovky sekvencí (+ vícenásobné substituce,) -> vytvoření programu PAUP
tudíž
dokážeme odhadnout počet genetických změn pro každou skupinu, při znalosti průměrné mutační rychlosti na generaci je pak možné dopočítat počet generací ke společnému předku
Cann a kol.
analyzovali mtDNA 147 lidí, každý vzorek přidělen do jedné skupiny podle geografického původu
k analýze použili 12 restrikčních enzymů (průměrně připadlo 370 restrikčních míst na osobu)
• analýza pomocí PAUP
Výsledek 1
dva významné shluky – zástupci se sub-saharským předkem / ostatní regiony včetně Afriky -> s největší pravděpodobnose společný předek z Afriky -> v souladu s oběma modely původu: africký model počítá s ještě jedním mladším předkem: oba modely se tedy liší názorem na to, jak a kdy k tomu došlo
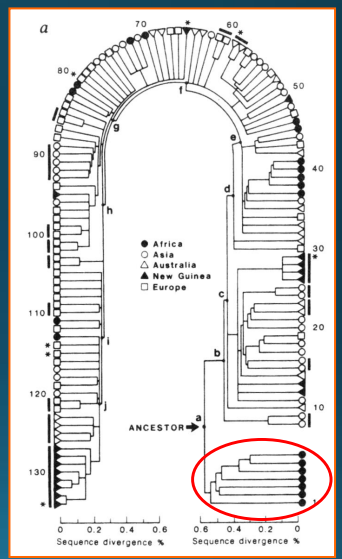
Výsledek 2
na základě odhadu četnose mutací v mtDNA a sledované variability pokus o výpočet, kdy došlo k oddělení od tohoto společného předka -> 2-4% rozdílů za milion let -> společný předek žil před 285 000 lety (2%) až 143 000 lety (4%) -> průměr 214 000 let
? Co se stalo s mtDNA předků, kteří v té době žili mimo Afriku?
podle všeho byl H. erectus, neanderthalensis aj. zcela nahrazen moderním člověkem bez vzájemného křížení
mitochondriální Eva
společný předek, který žil v Africe před zhruba 200 000 lety
i neexistovala v té době v Africe pouze jediná Eva, ale těchto Ev, předchůdkyň bylo zcela logicky určitě několik stovek nebo tisíce
Námitky proti modelu nahrazení
rozdělení mtDNA na africkou a neafrickou nemusí odrážet reálné rozdělení populací (neboli geografická asociace tohoto rozdělení nemusí představovat ono následné nahrazení expandující africkou populací)
Otázka interpretace genetických stromů (Alan Templeton, 1993, 1997, 1998)
1) získané genetické stromy nemusí odpovídat populačním stromům - avšak řada studií až do současnosti ukazuje na vysokou korelaci mezi genetickými a populačními údaji = námitku lze zamítnout
2) odlišnost afrických populací od ostatních nemusí být důsledkem nahrazení, ale příčinou může být omezený genový tok v důsledku prostorové izolace - jsou-li populace příliš vzdálené, neprobíhá genový tok a výsledek vypadá jako při nahrazení jedněch populací jiný = Templetonův model omezeného genového toku
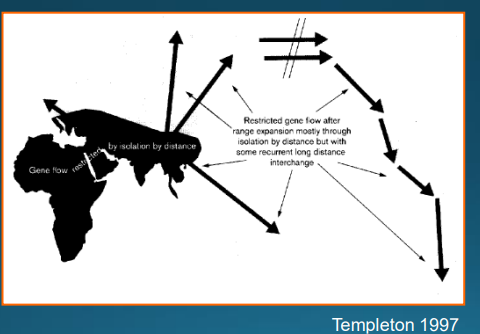
Templetonův model omezeného genového toku
dle Templetona neexistuje důkaz o expanzi africké populace mimo Afriku
africké stopy přítomné ve všech populacích jsou spíše důsledkem stálého genového toku mezi geograficky blízkými oblastmi v uplynulých několika stech tisících letech (africké znaky se tak do těchto populací rozšířily)
nové analýzy genetických stromů (kombinuje až 10 různých znaků) ukazují dle něj na několik migrací z Afriky s neustálým genovým tokem
první expanze proběhla před 1,7 miliony let – Homo erectus
druhá vlna před 800 až 400 000 lety
a) expanze Homo heidelbergensis (předek Neandrtálce) nebo
b) možná migrace jiného předka Neandrtálce a moderního člověka
třetí vlna před asi 150 000 lety
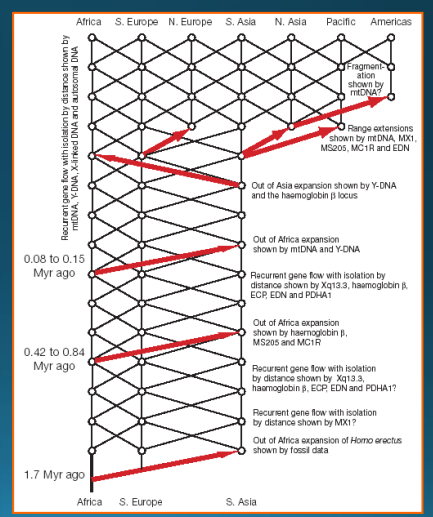
„Out of Africa again and again“
Templetonovy výsledky svědčící o opakovaném genovém toku (expanze z Afriky a následné křížení) v průběhu posledních 2 milionů let:
• podporují africké genetické kořeny = moderní člověk se mohl rozšířit z Afriky, avšak nejen odtud = Eva mohla žít kdekoliv
• do jisté míry podporuje multiregionální model – opakované migrace s genovým tokem vedou ke stejnému výsledku, tedy africkému původu, přestože je původ ve skutečnosti multiregionální
• plně však nepodporuje ani jeden ze dvou modelů = uznáván jako další model
Analýza chromozomu Y - výhody
Nehomologický úsek Y nerekombinuje
nabízí potvrzení výsledků studia mtDNA použitím jiných zdrojových dat
Různé odhady stáří společného předka dle chromozomu Y
???
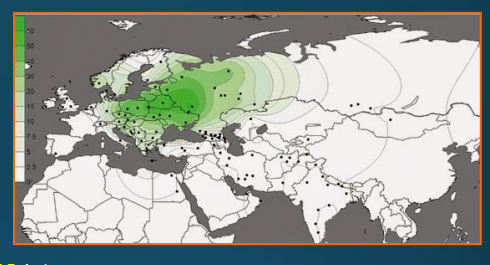
haploskupiny dle chromozomu Y
Dnes jednotlivé haploskupiny – např. haploskupina R
R1a – 2 centra výskytu
• Evropa • Centrální a Jižní Asie
Odlišná velikost populace mužů a žen
• ukazuje na to, že poměr mezi efektivní velikostí ženské a mužské populace (Nf /Nm) dosahoval hodnoty větší než 1,0
• a směrem do současnosti se zvyšuje v důsledku rychlejšího růstu hodnoty Nf
Odlišná velikost populace mužů a žen - proč
• nepoměr pohlaví jako reakce na nehostinné podmínky mimo Afriku zejména na kontinentální evropskou tundru
• ztráta významného podílu sběračství (vidíme u dnešních Inuitů – jen 10 % zajištěno sběrem)
• většinu potravy tak musí zajistit lov
• lov je nyní na větším území - musí se ulovit více zvěře (výpadek ze sběru) + zvířata se pohybují za potravou po větším území
= hlad, vyčerpání, nehody při lovu = zvyšuje se úmrtnost mladých mužů (například u dnešních Inuitů je výrazná převaha žen, v 19. st. 57 mužů na 100
atudium mtDNA a chromozom Y - závěr
výsledky a závěry analýzy mtDNA a chromozomu Y jsou navzájem kompatibilní = = můžeme je považovat za správné (včetně diploidních a jaderných znaků)
• společný předek moderních lidí žil v Africe
- Model afrického původu = OK
- Multiregionální model = OK + i jinde
Výsledky dalších analýz
diploidní znaky, jaderné geny -> nepodporují model nahrazení