Cerebellum
1/14
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai | Chat |
|---|
No analytics yet
Send a link to your students to track their progress
15 Terms
3 key facts about the cerebellum's size, neurone count, and position.
10% of total brain volume (anatomically small), but 80% of total brain neurones due to the massive granule cell population; essential for sampling/processing enormous amounts of information
Strategically positioned at the brainstem (pons level), allowing it to sample all ascending spinal cord signals plus motor and sensory nuclei.
Not crossed; cerebellar projections control the same side of the body (unlike the cerebral cortex)
Why is feedback control alone insufficient for smooth movement, and how does the cerebellum solve this?
Problem with feedback only: Sensory feedback (e.g. muscle spindle reflex) takes time — neural conduction + processing delay means the correction arrives after the body has already moved further. This causes overcorrection → oscillation → shaky, unstable movement.
Cerebellar solution — feedforward control: The cerebellum uses synaptic plasticity (LTP/LTD) based on previous movement success/failure to build internal models that predict the future state of the body and pre-calculate force/timing before the movement completes. It is proactive, not reactive.
What are the two main streams of afferent input to the cerebellum?
Periphery (sensory & execution):
• Proprioception (muscles/joints → limb position)
• Motor command copy (from spinal cord)
• Reflex feedback (involuntary spinal responses)Cerebral cortex (intent & context):
• Goal-oriented planning (high-level 'why')
• Motor concept (sequence of movement, from premotor cortex)
• Motor command (direct from pre-central gyrus)
• Spatial awareness (parietal/visual cortex → space and motion context)
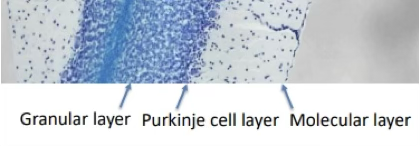
Describe the 3 layers of the cerebellar cortex.
Granular layer (innermost): densely packed granule cells; 80% of all brain neurones are granule cells
Purkinje cell layer (middle): large soma ('tree-shaped'), very extensive flat 2D dendritic tree (allows maximal packing) extending into the molecular layer
Molecular layer (outermost): sparse; where parallel fibres run horizontally and Purkinje dendrites extend up
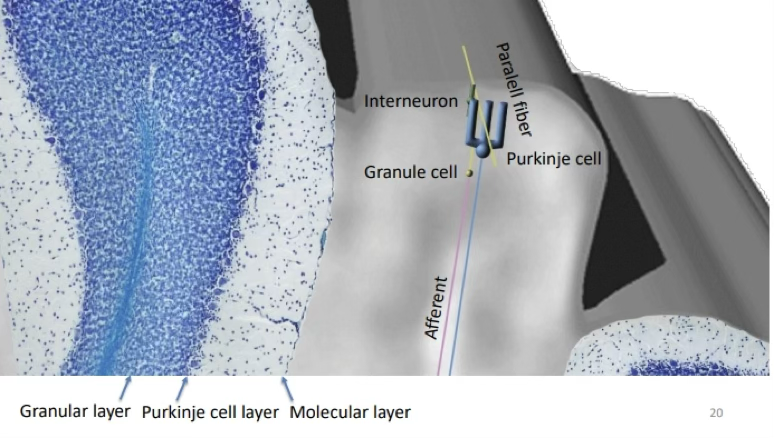
Describe the cerebellar circuit as shown in this figure.
Afferent inputs (mossy fibres) project onto granule cells in the granular layer
Convergence
Granule cell axons rise into the molecular layer and bifurcate into parallel fibres running horizontally
Parallel fibres pass perpendicularly through the flat dendritic trees of Purkinje cells, synapsing on them. One parallel fibre can contact thousands of Purkinje cells sequentially due to the extensive packing of Purkinje cells
Divergence
Interneurones (inhibitory) synapse onto granule or Purkinje dendrites to modulate signals
Purkinje cells send the final processed signal out of the cortex to the deep cerebellar nuclei
Convergence
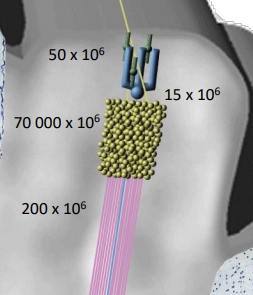
What does this figure illustrate about divergence in the cerebellar circuit?
Shows the extraordinary scale of divergence in the cerebellar cortex:
• ~50 million mossy fibre inputs enter
• ~70 billion granule cells (massive expansion)
• ~15 million Purkinje cells receive from granule cells
• ~200 million output fibres leave via deep cerebellar nuclei
Overall circuit:
Convergence (mossy fibres to granule cells)
Divergence (granule cells to purkinje cells)
Convergence (purkinje cells to deep cerebellar nuclei)
The granule cell layer acts as a massive parallel processing expansion layer, allowing the cerebellum to represent and compare enormous combinations of input patterns.
What are the unique properties of Purkinje cells and what does the cat paw experiment show?
Properties: Purkinje cells are inhibitory (GABA), spontaneously active at ~20 Hz, and receive very few active connections in adults (most parallel fibre synapses are silent).
Cat paw experiment:
Stimulating area below thumb (blue) → increases firing to ~40 Hz
Stimulating wrist below pinky (green) → decreases firing in blue area to ~10 Hz
Opposite areas produce opposite effects because the stimulated area drives inhibitory interneurones that suppress the antagonist area
This directional tuning is learned — disrupting normal input breaks it down
This was done by stimulating neurones in those 2 areas simultaneously with nonsense input (random input → random responses)
What is the area that regulates cerebellar learning? How?
The inferior olivary nucleus sends axons into the cerebellum as climbing fibres:
A single climbing fibre wraps around and 'climbs' one Purkinje cell's entire dendritic tree; this is one of the strongest synaptic connections in the entire nervous system
Highly active when learning new, difficult motor skills
When a climbing fibre fires, it triggers a massive Ca²⁺ influx into the Purkinje cell
This Ca²⁺ changes the strength of parallel fibre synapses (induces plasticity (LTD))
The cerebellum updates its motor command based on error signals, hence “learning”
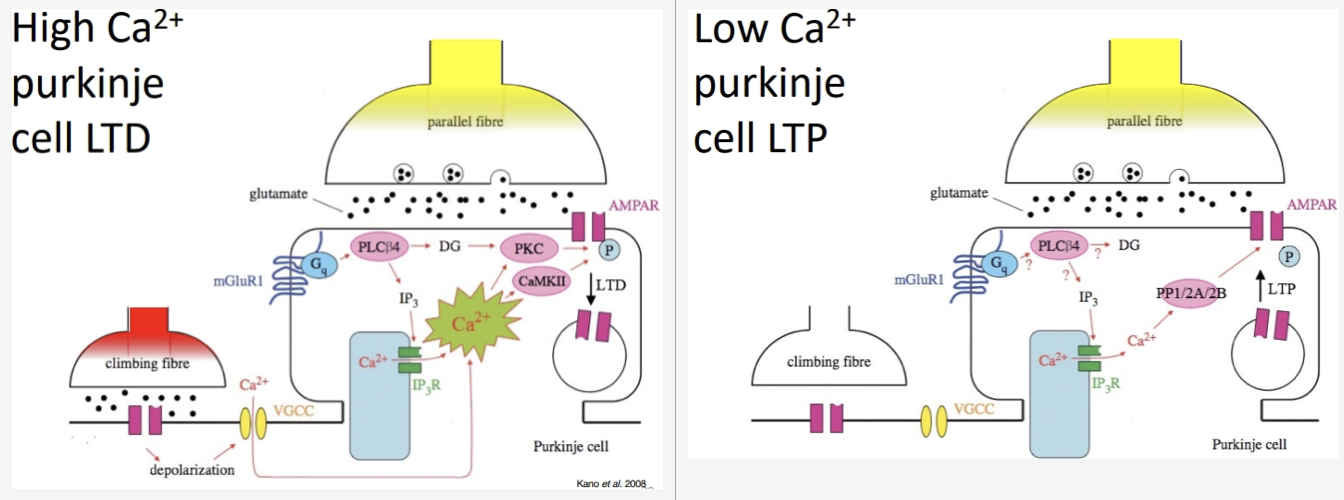
What happens when climbing fibres are active simultaneously vs. not simultaneously with parallel fibres?
Parallel fibre active + climbing fibre active simultaneously:
Climbing fibre causes a massive depolarisation, leading to huge Ca2+ influx
This triggers the removal of AMPA receptors from the post-synaptic purkinje membrane
Outcome: LTD at the parallel fibre–Purkinje synapse (and LTP at the interneurone)
That motor pattern is associated with an error, and thus weakened
Parallel fibre active + climbing fibre NOT active:
Significantly less depolarisation and minimal Ca2+ influx
In this basal state, the cell inserts more AMPA receptors into the post-synaptic membrane
Outcome: LTP at the parallel fibre–Purkinje synapse (and LTD at the interneurone)
That motor pattern was successful, and thus strengthened
Summary: The cerebellum learns by suppressing 'wrong' synapses (those co-active with errors) and strengthening 'right' ones, leaving a refined, efficient motor command.
How do interneurones and Purkinje cells work synergistically in cerebellar learning?
The two work in opposite but complementary ways to suppress wrong motor commands.
Both mechanisms converge to suppress the wrong pathway from two angles simultaneously.
What is the efferent output pathway of the cerebellum and where does it project?
Purkinje cells project to deep cerebellar nuclei (inside white matter), the actual output points. From there, signals reach 3 main destinations:
Motor nuclei in the brainstem: posture, balance, eye movements
Spinal cord ventral horn: direct influence on motor neurones for execution
Primary motor cortex (via thalamus): voluntary fine motor control
Organisation is closed-loop and reciprocal: any area that sends input to the cerebellum also receives processed output back from it.
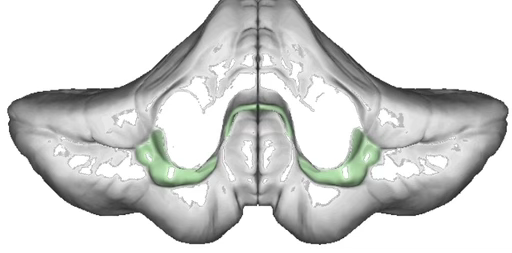
What is the vestibulocerebellum’s role?
Evolutionarily oldest part of the cerebellum; has no deep cerebellar nucleus, but rather the Purkinje cells project directly to the vestibular nuclei in the brainstem.
Function: feedforward control of gaze via the Vestibulo-Ocular Reflex (VOR):
Vestibular system detects head movement
Vestibulocerebellum predicts required compensatory eye movement
Sends proportionate feedforward signal to eye muscles
What happens when the vestibulocerebellum’s damaged?
Damage (e.g. stroke):
The feedforward prediction becomes faulty → eyes overshoot/undershoot → unstable visual field
Severe balance issues because the body can no longer proactively compensate for head movements
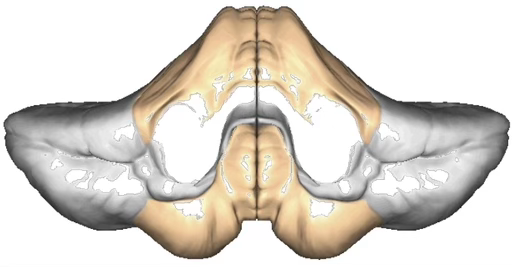
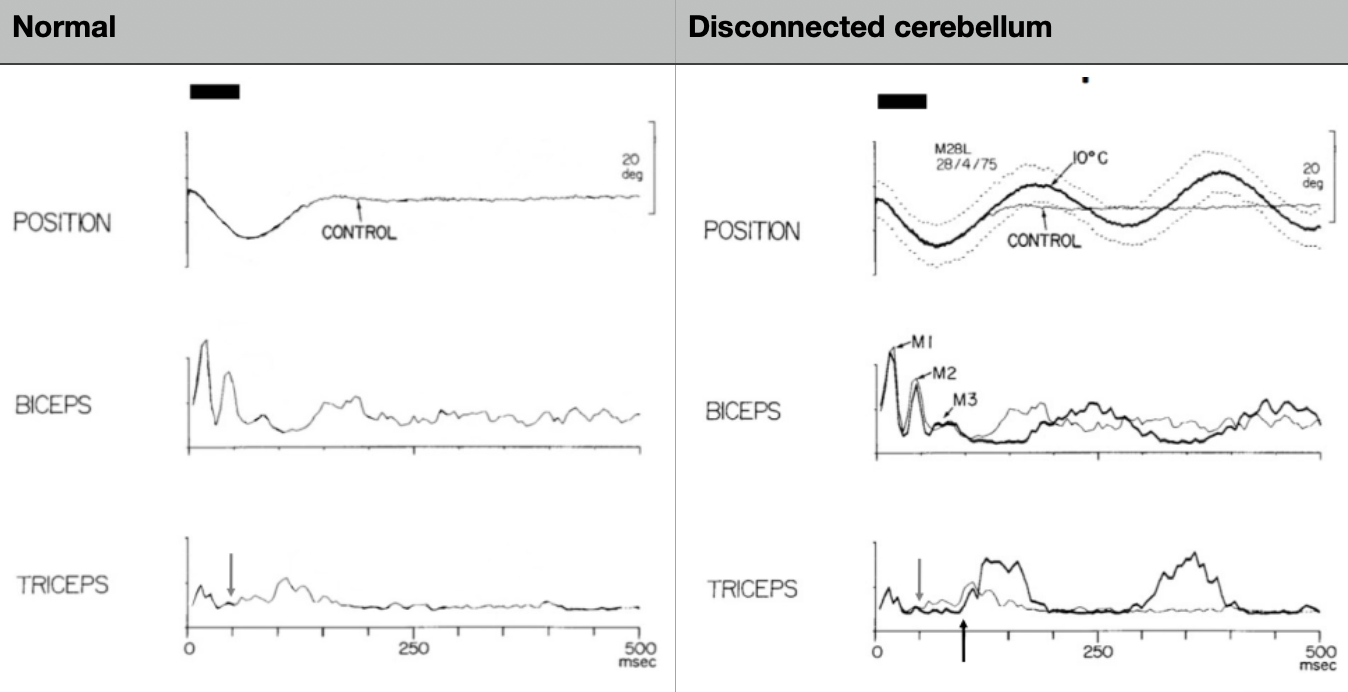
What does the monkey weight experiment show about the spinocerebellum?
The spinocerebellum receives massive proprioceptive/somatosensory data from the spinal cord to coordinate movement stability.
Monkey experiment (Vilis & Hore 1977):
Normal: when a weight is added, bicep stretch reflex fires AND spinocerebellum simultaneously sends a preparatory contraction to the triceps as a 'brake'
With cerebellum cooled (disconnected): stretch reflex still occurs (spinal) BUT no feedforward signal to triceps → bicep contracts unopposed → arm overshoots, then oscillates wildly
Conclusion: Feedback reflex alone is insufficient; feedforward braking is mandatory for smooth stable movement. Without it → dysmetria and tremor.
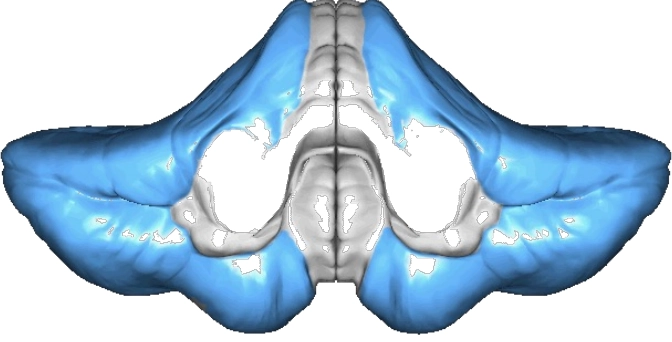
What is the cerebrocerebellum, and what does its experiment with the monkey show?
The largest part of the cerebellum with 2 zones:
Intermediary zone (overlapping with spinocerebellum): both cortical and spinal input → voluntary movement requiring proprioceptive regulation
Far lateral zone: only cortical connections → function unknown
Experiment: monkey trained to click a button; scientist occasionally pushes the finger
Normal: cerebrocerebellum sends predictive stabilising signal → minimal oscillation
Disconnected: no feedforward signal → oscillations/tremor whenever active movement is made
Conclusion: feedforward signals are mandatory to suppress oscillations. Without them, every voluntary movement is trapped in a slow, oscillating feedback loop that is too slow to produce smooth, human-like movement.