Deel 2: de onderdelen van de cell (cellbio)
1/70
Earn XP
Description and Tags
dellen van de cell
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai | Chat |
|---|
No analytics yet
Send a link to your students to track their progress
71 Terms
Membraan
Basisstructuur van alle biologische membranen: een dubbele fosfolipidelaag met daarin eiwitten. Apolair karakter laat toe grote concentratieverschillen binnen/buiten de cel te onderhouden. Functioneert als actieve filter voor transport van stoffen.
glycocalyx
Stevig maar elastisch omhulsel rond de bacteriecel. Voornaamste functies: fysische bescherming van de protoplast en opvangen van de turgordruk (voorkomt openbarsten). Doorlaatbaar voor kleine moleculen — de plasmamembraan (niet de celwand) bepaalt de permeabiliteit. Hoofdcomponent: mureïne. Basis voor onderscheid tussen Gram-positieve en Gram-negatieve bacteriën.
celwand (bacterie)
Stevig maar elastisch omhulsel rond de bacteriecel. Voornaamste functies: fysische bescherming van de protoplast en opvangen van de turgordruk (voorkomt openbarsten). Doorlaatbaar voor kleine moleculen — de plasmamembraan (niet de celwand) bepaalt de permeabiliteit. Hoofdcomponent: mureïne. Basis voor onderscheid tussen Gram-positieve en Gram-negatieve bacteriën
mureïne
Heteropolysaccharide dat de hoofdcomponent vormt van de bacteriële celwand. Bestaat uit peptidoglycaanketens (polysaccharideruggengraat met peptidezijketens) die onderling door covalente bindingen verbonden zijn. Vormt één enkel reusachtig zakvormig molecule rond de cel. Bevat D-aminozuren — uniek voor bacteriën. Penicilline blokkeert de covalente koppeling tussen de ketens, waardoor de celwand te zwak wordt en de cel openbarst.
Gram positive
Bacteriën met één plasmamembraan omgeven door een zeer dikke mureïnecelwand. Bij de Gramkleuring wordt het kristalviolet-jodiumcomplex niet weggewassen door alcohol → kleuren blauw-paars. Gevoeliger voor penicilline door dikkere celwand.
gram negetive
Bacteriën met een dunne celwand gelegen in een periplasmatische zone tussen twee membranen (binnenste plasmamembraan + buitenste lipopolysaccharidembraan). Voorbeeld: Escherichia coli. Minder gevoelig voor penicilline door dunnere celwand.
celwand (fungi)
Bestaat grotendeels uit chitine, aangevuld met andere polysacchariden (bv. mannaan bij gisten, cellulose bij bepaalde groepen). Samenstelling varieert sterk per taxonomische groep. Bij hogere fungi vormt het geheel van celwanden het voornaamste structuurelement voor de macroscopische vorm van de fungus.
celwand (planten)
Karakteristiek kenmerk van plantencellen. Dikker, sterker en onbeweeglijker dan de glycocalyx van dierlijke cellen. Functies: fysische bescherming, opvangen turgordruk, aanhechting van cellen aan elkaar, vorming van transportkanalen, communicatie via plasmodesmata. Onderscheid tussen primaire celwand (flexibel, voor groei) en secundaire celwand (steviger, na groei). Opgebouwd volgens het principe van vezels (cellulose-microfibrillen) in een amorfe matrix (pectine, hemicellulose, glycoproteïnen).
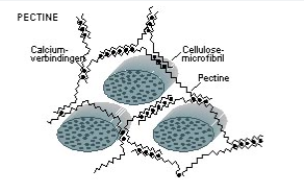
Pectine
Matrixpolysaccharide in de primaire plantencelwand, opgebouwd uit galacturonzuur en andere suikers. Heeft veel negatieve ladingen en grote affiniteit voor Ca²⁺. Neemt veel water op (geleiachtige toestand). Pectinemoleculen vormen netwerken via calciumbruggen. Vormt de lijmlaag tussen celwanden van naburige cellen. Lost op in warm water — verklaart waarom aardappelen gaar worden bij koken.
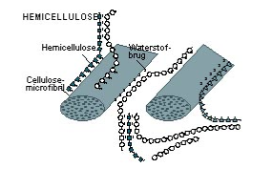
Hemicellulose
Vertakt polysaccharide in de primaire celwand, opgebouwd uit pentosen en hexosen. Bindt via H-bruggen aan cellulose-microfibrillen en vormt zo een driedimensionaal netwerk. Samenstelling varieert sterk per plantensoort en ontwikkelingsstadium.
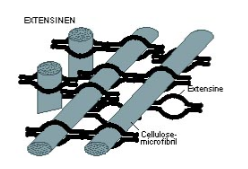
Glycoproteïnen
Eiwitten met suikergroepen in de celwandmatrix, bekend als extensinen. Hechten cellulose-microfibrillen aan elkaar. Bevatten veel hydroxyproline (verhindert α-helixvorming). Bij celgroei laten de eiwitten elkaar los zodat de microfibrillen kunnen verschuiven en de celwand kan meegroeien.
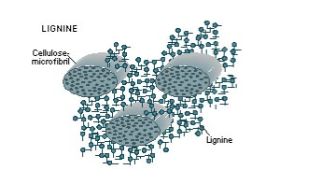
Lignine
Zeer onoplosbaar polymeer in de secundaire celwand van xyleemcellen, opgebouwd uit fenolderivaten die in drie dimensies polymeriseren. Vult de ruimte tussen cellulose-microfibrillen op. Maakt de celwand star: de cel kan niet meer groeien en sterft. Verhoogt de druksterkte enorm (draagt gewicht van bomen). Ontstond evolutionair ±400 miljoen jaar geleden toen planten het land begonnen te bewonen.
Cutine
Polymeer van vetzuren in de secundaire celwand van epidermiscellen. Vormt samen met was een waterondoordringbare laag aan de buitenkant van de epidermiscel.
Plasmodesmata
Dunne cytoplasmastrengen die vrijwel alle levende cellen in een hogere plant met elkaar verbinden doorheen de celwand. De plasmamembranen van twee naburige cellen lopen in elkaar over. Bevatten vaak een desmotubulus in het midden. Gevormd tijdens de aanleg van de nieuwe celwand bij celdeling. Zorgen voor sneller transport van stoffen tussen cellen dan via de plasmamembraan alleen mogelijk zou zijn.
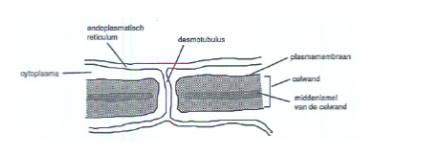
desmotubulus.
Dubbele membraanstructuur die door het midden van een plasmodesma loopt en het endoplasmatisch reticulum van twee naburige plantencellen met elkaar verbindt.
plasmamembraan
Membraan dat alle levende cellen omgeeft. Functioneert als actieve filter: transporteert selectief voedingsstoffen naar binnen en afvalstoffen naar buiten. Buitenste fosfolipidelaag bevat suikerresidu's (van glycolipiden en glycoproteïnen) die een rol spelen in celcommunicatie. Bij prokaryoten bevindt de elektronentransportketen zich op de plasmamembraan; bij eukaryoten op de cristae van de mitochondriën.
Kanaaljuncties
Juncties tussen dierlijke cellen via connexons: cilindervormige structuurtjes die hydrofiele kanaaltjes vormen tussen twee naburige cellen. Kunnen geopend en gesloten worden. Laten kleine moleculen (ionen, suikers, aminozuren, vitaminen) en elektrische impulsen van cel tot cel passeren. Proteïnen zijn te groot om erdoor te gaan.
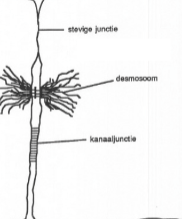
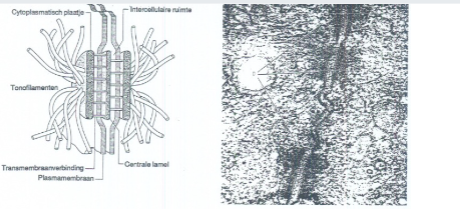
Desmosomen
Juncties met voornamelijk een adhesiefunctie: hechten naburige dierlijke cellen aan elkaar vast in een weefsel. Bestaan uit twee cytoplasmatische plaatjes en een centrale lamel, verbonden door proteïnebruggetjes doorheen de plasmamembranen. Tonofilamenten (microfilamenten van het cytoskelet) zijn verankerd op de plaatjes, waardoor de cytoskeletten van naburige cellen met elkaar verbonden worden.
Dichte juncties
Juncties waarbij de twee plasmamembranen via speciale proteïnen aan elkaar gekleefd zijn, zonder intercellulaire ruimte. Lopen helemaal rond de cel zodat geen vloeistof meer tussen de cellen kan passeren. Twee functies: (1) voorkomen terugdiffusie van stoffen via de intercellulaire ruimte (bv. in darmepitheel), (2) vormen een barrière binnen de membraan zelf zodat membraanproteïnen niet van de ene naar de andere kant van de cel kunnen diffunderen — behoudt zo de polariteit van de cel.
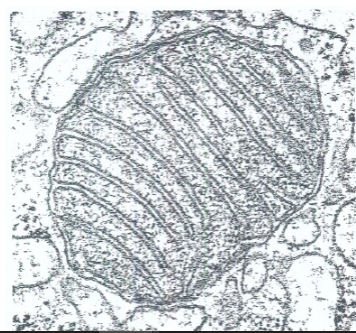
De mitochondriën
elangrijkste plaats van energieproductie: O2-ademhaling; Krebs-cyclus en elektronentransportketen; vetzuurafbraak (β-oxidatieweg)/ Mitochondriën zijn aanwezig in vrijwel alle eukaryote cellen en zorgen voor energieproductie via de O₂-ademhaling. Ze zijn ±1 μm groot (vergelijkbaar met een bacteriecel) en een cel kan er duizenden bevatten, afhankelijk van de energiebehoefte.Ze hebben een dubbel membraan: de binnenste membraan vormt instulpingen (cristae) waar de elektronentransportketen plaatsvindt. De matrix (stroma) is de locatie van de Krebs-cyclus, β-oxidatie van vetzuren en aminozuurmetabolisme. Hoe actiever de cel, hoe meer en hoe sterker ontwikkelde cristae.Mitochondriën bezitten eigen circulair DNA en 70S ribosomen (vergelijkbaar met prokaryoten), maar de meeste mitochondriale eiwitten worden gecodeerd door kern-DNA en in het cytoplasma gesynthetiseerd. Dit leidt tot extranucleaire erfelijkheid (overerving niet via Mendel).Vermenigvuldiging gebeurt door tweedeling.Volgens de endosymbiontische theorie (Lynn Margulis) zijn mitochondriën afkomstig van prokaryote endosymbionten die opgenomen werden in voorlopers van eukaryote cellen. Bewijs: circulair DNA, 70S ribosomen, elektronentransportketen op de binnenste membraan. Deze theorie is nu algemeen aanvaard
De plastiden
Groep van verwante organellen (chloroplasten, amyloplasten, chromoplasten) die allemaal afstammen van de proplastide. Aanwezig in planten en verantwoordelijk voor fotosynthese, zetmeelopslag en pigmentopslag.
proplastiden
Ongedifferentieerde voorlopers van alle plastidetypen. Aanwezig in meristeemcellen en voortplantingscellen. Alle andere plastiden kunnen hieruit ontstaan

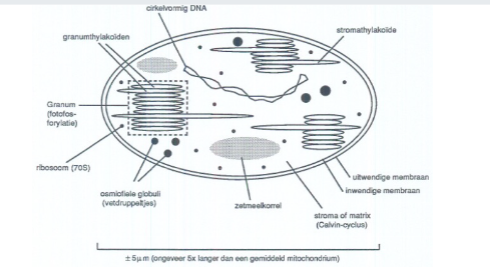
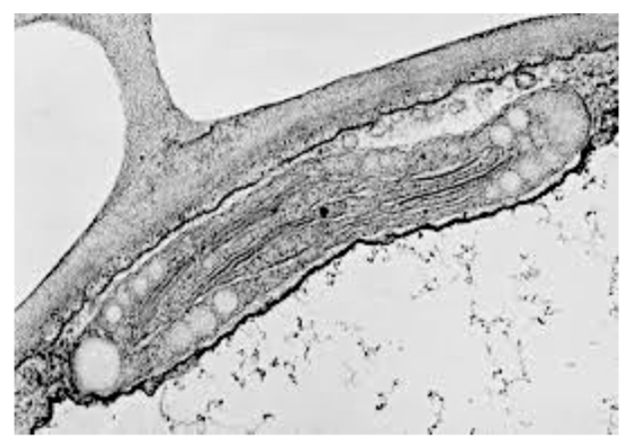
chloroplasten
Plastiden verantwoordelijk voor fotosynthese (H₂O + CO₂ → suiker + O₂). Bevatten chlorofyl (groen), zijn ±5 μm groot, en hebben een dubbele membraan + thylakoïdmembranen. Bevatten eigen circulair DNA en 70S ribosomen (extranucleaire erfelijkheid). Vermenigvuldigen door tweedeling.
amyloplasten
Kleurloze plastiden die zetmeel opslaan. Veel aanwezig in reserveweefsel (bv. aardappel). Functioneren ook als statolieten: zware partikels waarmee planten de zwaartekracht detecteren. Ontstaan alleen uit proplastiden.
chromoplasten
Gekleurde plastiden die carotenoïdpigmenten opstapelen (rood, oranje, geel). Voorbeelden: tomaten, wortelen, bloemen. Kunnen ontstaan uit proplastiden, amyloplasten én chloroplasten (bv. bij het rijpen van vruchten).
thylakoïdmembranen
Derde membraansysteem in chloroplasten, afkomstig van instulpingen van de binnenste membraan maar grotendeels daarvan afgesplitst. Locatie van de fotofosforylatie (opvangen lichtenergie → chemische energie + O₂-vrijstelling).
thylakoïdruimte
De afgesloten ruimte binnenin de thylakoïden (het lumen). Speelt een rol in de fotofosforylatie
stroma/matrix
De vloeistofruimte in de chloroplast buiten de thylakoïden. Locatie van de Calvin-cyclus: synthese van suiker uit CO₂ en H₂O met behulp van chemische energie uit de fotofosforylatie.
grana
Regelmatige stapeltjes van thylakoïden binnen de chloroplast. Locatie van de fotofosforylatie. Meervoud van granum.
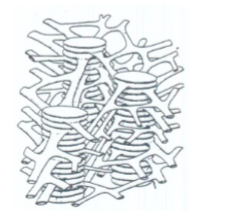
granumthylakoïden
Thylakoïden die gestapeld zijn binnen een granum. Onderdeel van de grana-structuur.
stromathylakoïden.
Thylakoïden die door het stroma lopen en de grana met elkaar verbinden. Ze verbinden het lumen van granumthylakoïden van verschillende grana met elkaar.

Het endoplasmatisch reticulum
Een netwerk van membranen dat een afgesloten ruimte vormt binnen de cel. Bestaat uit drie types: het R.E.R. (ruw, met ribosomen), het S.E.R. (glad, zonder ribosomen) en het T.E.R. (transitioneel, vormt transportvesikels). R.E.R. en S.E.R. zijn continu met elkaar. Het R.E.R. bestaat uit stapels afgeplatte cisternae, het S.E.R. uit een netwerk van fijne buisjes. Het R.E.R. is ook continu met de buitenste kernmembraan. Het ER is het startpunt van de secretieweg van de cel: transportvesikels vertrekken vanuit het T.E.R. naar het Golgi-apparaat, en vervolgens naar het plasmalemma of lysosomen. Het ER is ook de belangrijkste plaats voor membraansynthese: andere membranen in de cel groeien aan door incorporatie van membraanfragmenten afkomstig van het ER.
Het ruw endoplasmatisch reticulum
Bestaat uit stapels afgeplatte membraanzakjes (cisternae) bezet met ribosomen aan de buitenkant. Voornaamste functie: synthese van eiwitten bestemd voor secretie, het Golgi-apparaat of lysosomen. Deze eiwitten hebben een signaalsequentie aan hun N-terminus waardoor ze tijdens de synthese door de membraan naar het lumen worden getransporteerd; daarna wordt de signaalsequentie verwijderd. De meeste van deze eiwitten worden geglycosyleerd (= glycoproteïnen): suikers worden binnenin het ER gekoppeld aan serine/threonine (O-glycosylatie) of asparagine (N-glycosylatie). Cellen gespecialiseerd in secretie (bv. kliercellen van de pancreas, plasmacellen) bevatten zeer veel R.E.R. Het T.E.R. is een gespecialiseerd gedeelte aan de uiteinden van het R.E.R. waar transportvesikels worden afgesnoerd richting Golgi-apparaat

Het glad endoplasmatisch reticulum
synthese van lipiden; detoxificatiereacties/ Bestaat uit een netwerk van fijne buisjes, zonder ribosomen. Heeft twee hoofdfuncties. Ten eerste de synthese van lipiden: alle lipiden behalve vetzuren worden in het ER aangemaakt, inclusief sterolen zoals cholesterol en steroïdhormonen (bv. testosteron in Leydig-cellen). Ten tweede detoxificatie: apolaire toxische stoffen accumuleren gevaarlijk in celmembranen en worden door het S.E.R. omgezet naar polaire verbindingen die de cel kan uitscheiden. Het sleutelenzym hierbij is cytochroom P450, dat OH-groepen plaatst op de toxische stof, waarna andere enzymen extra polaire groepen (suikers, fosfaat) toevoegen. Het S.E.R. staat ook in voor de vorming van peroxisomen. In spiercellen is het S.E.R. gespecialiseerd als sarcoplasmatisch reticulum, dat Ca²⁺-ionen uit het cytoplasma verwijdert en opslaat
microsomen
Artefacten die ontstaan wanneer cellen worden gehomogeniseerd: de ER-membranen splitsen op in kleine vesikels (microsomen). Ze komen nooit voor in levende cellen. Ruwe microsomen zijn afkomstig van R.E.R. en zijn bezet met ribosomen (dus zwaarder). Gladde microsomen zijn afkomstig van S.E.R. en hebben geen ribosomen. De twee types kunnen gescheiden worden via centrifugatie op een dichtheidsgradiënt. Gladde microsomen zijn moeilijker zuiver te isoleren omdat ze lijken op vesikels van het plasmalemma of Golgi-apparaat.
Het Golgi-apparaat
Bestaat uit stapeltjes afgeplatte membraanzakjes (cisternae) zonder ribosomen. Ontvangt transportvesikels van het R.E.R. en verwerkt, modificeert en sorteert proteïnen voor verdere bestemming. Hoofdfuncties: modificatie van glycoproteïnen (suikergroepen aanpassen), productie van secretievesikels (naar buiten de cel) en aanleveren van enzymen aan lysosomen. Membraanrecyclage voorkomt dat het plasmalemma of lysosomen ongecontroleerd aangroeien.
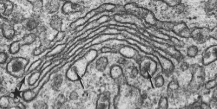
cisternae.
Afgeplatte membraanzakjes waaruit het Golgi-apparaat is opgebouwd. Hebben licht opgezwollen uiteinden waarvan vesikels afsnoeren.
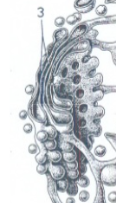
Golgi-vesikels.
Vesikels die afsnoeren van de uiteinden van de Golgi-cisternae. Verschillende types naargelang bestemming: secretievesikels (naar buiten de cel), primaire lysosomen (naar lysosomen), of vesikels voor celwandsynthese (bij planten en fungi).
Golgi-stapeltje
Eén stapel van Golgi-cisternae. Een cel kan er enkele tot vele honderden bevatten afhankelijk van het celtype.
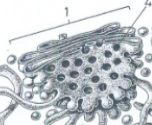
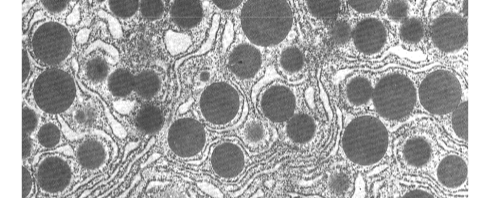
secretievesikels
Golgi-vesikels bestemd voor secretie buiten de cel. Migreren via microtubuli naar het plasmalemma, versmelten ermee en geven hun inhoud af aan de buitenwereld (exocytose). Bij planten en fungi ook verantwoordelijk voor aanvoer van celwandmateriaal.
zymogeen-granulen
Grote secretievesikels in kliercellen (bv. pancreas) die zymogenen bevatten. Worden als zodanig herkend op E.M.-foto's van secretiecellen.
Zymogenen
Inactieve precursoren van verteringsenzymen. Worden in inactieve vorm aangemaakt en uitgescheiden, en pas in het darmkanaal omgezet naar actieve verteringsenzymen. Dit voorkomt dat actieve verteringsenzymen per ongeluk schade aanrichten in de cel zelf.
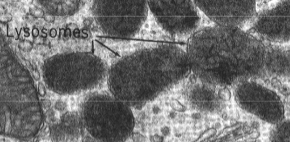
De lysosomen
Organellen gevormd door het Golgi-apparaat (en onrechtstreeks het R.E.R.). Functie: afbraak van macromoleculen tot bouwstenen ("de maag van de cel"). Bevatten hydrolytische enzymen (proteasen, nucleasen, lipasen, glycosidasen, enz.) met een zuur pH-optimum. De zure inhoud wordt onderhouden door protonpompen in het membraan (ATP-verbruik). Vrijgekomen bouwstenen gaan naar het cytoplasma voor hergebruik. Primaire lysosomen = nog niet actief, afkomstig van Golgi. Secundaire lysosomen = actief bezig met vertering, herkenbaar op E.M. aan dense, onregelmatige inhoud
autofagie.
Afbraak van eigen cellulair materiaal door lysosomen. Het eigen materiaal wordt omsloten en versmelten met primaire lysosomen. Onverteerbare resten worden uitgescheiden via exocytose of blijven circuleren in de cel
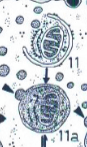
heterofagie.
Afbraak van vreemd materiaal dat via endocytose in de cel werd opgenomen. Het vreemde materiaal zit in endosomen die versmelten met lysosomen, waarna afbraak plaatsvindt.
endosomen
Vesikels gevormd aan het plasmalemma bij opname van materiaal van buiten de cel (endocytose). Versmelten met lysosomen voor verdere afbraak. Twee types: fagosomen (vast materiaal) en pinosomen (opgeloste moleculen).
endocytose
Het proces waarbij de cel materiaal van buiten opneemt via vorming van endosomen aan het plasmalemma. Omvat fagocytose (vast materiaal) en pinocytose (opgeloste moleculen). Onderdeel van heterofagie.
fagocytose
Opname van vast materiaal (bv. bacteriën) via het plasmalemma. De gevormde endosomen noemt men fagosomen. Deze versmelten met primaire lysosomen tot secundaire lysosomen waar het materiaal wordt afgebroken.
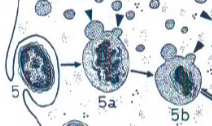
pinocytose
Opname van opgeloste moleculen via het plasmalemma. De gevormde endosomen noemt men pinosomen. Deze versmelten met primaire lysosomen voor verdere afbraak.
De peroxisomen
Organellen gevormd door afsnoering van het S.E.R. Omgeven door één enkele membraan (geen dubbele membraan zoals mitochondriën, geen interne membranen). Op E.M.-foto's herkenbaar aan kristallijn materiaal (vaak katalase-enzymen) — zo te onderscheiden van primaire lysosomen.
Voornaamste functie: oxidatieve afbraakreacties via oxidase-enzymen. Hierbij wordt O₂ opgenomen en H₂O₂ gevormd (toxisch bij overmaat). Andere substraten worden vervolgens afgebroken met behulp van dat H₂O₂ via peroxidase-enzymen. Overmaat H₂O₂ wordt geneutraliseerd door katalase: 2 H₂O₂ → 2 H₂O + O₂.
Worden samen met glyoxisomen ondergebracht in de groep van de microbodies.
![<p> Organellen gevormd door afsnoering van het S.E.R. Omgeven door één enkele membraan (geen dubbele membraan zoals mitochondriën, geen interne membranen). Op E.M.-foto's herkenbaar aan kristallijn materiaal (vaak katalase-enzymen) — zo te onderscheiden van primaire lysosomen.</p><p class="font-claude-response-body break-words whitespace-normal leading-[1.7]">Voornaamste functie: oxidatieve afbraakreacties via oxidase-enzymen. Hierbij wordt O₂ opgenomen en H₂O₂ gevormd (toxisch bij overmaat). Andere substraten worden vervolgens afgebroken met behulp van dat H₂O₂ via peroxidase-enzymen. Overmaat H₂O₂ wordt geneutraliseerd door katalase: 2 H₂O₂ → 2 H₂O + O₂.</p><p class="font-claude-response-body break-words whitespace-normal leading-[1.7]">Worden samen met glyoxisomen ondergebracht in de groep van de <strong>microbodies</strong>.</p>](https://assets.knowt.com/user-attachments/316a2db8-9f19-4a88-8f66-b04ec72ed6ff.png)
De glyoxisomen
Type microbody, uitsluitend aanwezig in planten (niet in dieren). Bevatten de enzymen van de glyoxylzuurcyclus waarmee vetten worden omgezet in suikers — iets wat dieren niet kunnen. Zeer actief in cellen van kiemende oliehoudende zaden (bv. pindanoten), waar reservevet wordt omgezet naar suiker voor de groeiende kiemplant. Op E.M.-foto's vaak duidelijk geassocieerd met vetdruppeltjes in de cel.
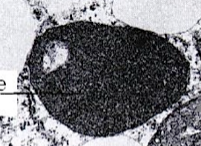
Reservepartikels
Onopgeloste polysacchariden of proteïnen in partikelvorm in het cytoplasma. Twee belangrijke types: glycogeenpartikels en proteïnelichaampjes. Glycogeenpartikels hebben een typische rozetvorm, zijn groter en heterogener dan ribosomen, en komen voor in lever- en spiercellen (energiereserve) en bij prokaryoten. Proteïnelichaampjes (protein bodies) komen voor in veel celtypes, vooral talrijk in zaden. Ze worden gevormd uit R.E.R.-strengen die opzwellen door opgestapeld reserveproteïne.
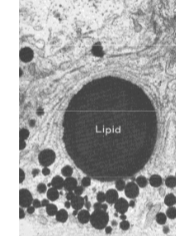
Vet- en oliedruppels (oleosomen)
Bevatten bijna uitsluitend triacylglycerolen en dienen als energiereserve of als reserve van lipideprecursoren. Op E.M.-foto's herkenbaar aan homogene, ronde vorm. Worden gevormd door het S.E.R. In adipocyten (vetcellen) kan de gehele cel uit één grote vetdruppel bestaan. Vetzuren uit vetdruppels worden afgebroken via β-oxidatie in de mitochondriën — op E.M.-foto's ziet men dan ook vaak een duidelijke associatie tussen vetdruppels en mitochondriën.
Kristallen
Kristallijne inclusies in veel celtypes, vaak proteïnekristallen met onbekende functie. Komen voor in vrijwel alle celorganellen, maar vooral frequent in peroxisomen. In plantencellen komen calciumoxalaat-kristallen veel voor: oxaalzuur is toxisch, maar door binding aan calcium wordt het onoplosbaar en dus onschadelijk. Calciumoxalaatkristallen bevinden zich vaak in de vacuole van plantencellen.

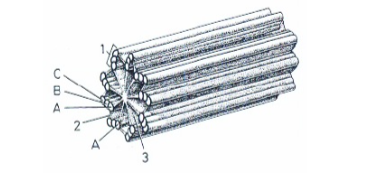
Microtubuli
Holle buisjes aanwezig in vrijwel alle eukaryote cellen. Opgebouwd uit het eiwit tubuline (α- en β-dimeren) die 13 protofilamenten vormen. Verantwoordelijk voor het cytoskelet, de spoelfiguur tijdens celdeling, en de inwendige structuur van ciliën en flagellen. Polymerisatie/depolymerisatie is een dynamisch en regelbaar proces. Inhibitoren: colchicine (blokkeert polymerisatie) en taxol (blokkeert depolymerisatie) — beide blokkeren de celcyclus bij de mitose.

tubuline
Globulair eiwit waaruit microtubuli zijn opgebouwd. Komt voor als α- en β-tubuline die samen dimeren vormen. Deze dimeren polymeriseren tot protofilamenten, waarvan 13 naast elkaar de wand van een microtubulus vormen.
protofilamenten
Lineaire polymeren van tubuline-dimeren. Dertien protofilamenten vormen samen de wand van één microtubulus. Op schuine doorsnede zijn er bijgevolg 13 tubulinemoleculen zichtbaar.
Microtubuli-organiserende centra
Structuren waaraan microtubuli met één uiteinde zijn verankerd en van waaruit ze worden opgebouwd. Voor het cytoskelet is dit het centrosoom (celcentrum), voor ciliën en flagellen is dit de basaalkorrel. In dierlijke cellen bevinden zich in het centrosoom twee centriolen.
Het cytoskelet
Driedimensionaal netwerk van filamenten dat vertrekt uit het celcentrum en zich door de hele cel vertakt. Bestaat uit microtubuli, actinefilamenten (microfilamenten) en — bij dierlijke cellen — intermediaire filamenten. Labiel en reversibel: wordt afgebroken bij het begin van de mitose en heropgebouwd aan het einde. Ligt aan de basis van cellulaire beweging (amoeboïde beweging, cytoplasmastroming, spiercontractie).
corticale microtubuli
Microtubuli die in plantencellen net onder de plasmamembraan lopen, parallel aan het celoppervlak. Sturen de oriëntatie van cellulosemicrofibrillen in de celwand.
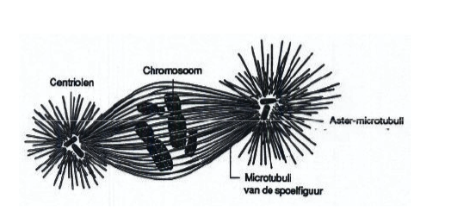
Centriolen
Structuren in het centrosoom van dierlijke en funguscellen, opgebouwd uit 9 tripletten van microtubuli. Staan loodrecht op elkaar. Functie: lokalisatie van het celcentrum en bepaling van de polariteit en het delingsvlak van de cel. Vermenigvuldigen door duplicatie voor de mitose. Structureel identiek aan basaalkorrels van ciliën/flagellen.
het pericentriolair materiaal
Dense substantie rond de centriolen die het eigenlijke microtubuli-organiserend centrum vormt. De microtubuli van het cytoskelet eindigen hier, niet bij de centriolen zelf. Ook aanwezig in plantencellen waar geen centriolen voorkomen.
Spoelfiguur
Structuur gevormd uit microtubuli tijdens de mitose, vanuit twee microtubuli-organiserende centra (centrosomen bij dieren, poolkapjes bij planten). Verantwoordelijk voor de scheiding van chromosomen. Wordt verder besproken bij celdeling.
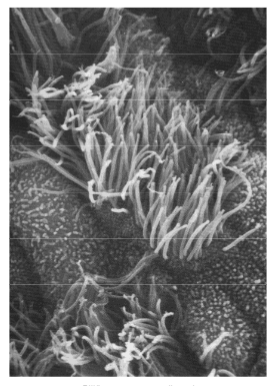
Ciliën
Korte cytoplasmatische uitstulpingen met een microtubuli-skelet (9 doubletten + 2 centrale singletten). Ingeplant op een basaalkorrel. Beweging ontstaat doordat microtubuliparen t.o.v. elkaar verglijden.
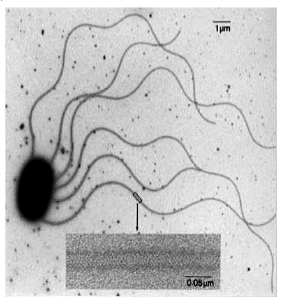
flagellen
Cytoplasmatische uitstulpingen, structureel identiek aan ciliën maar langer. Bij prokaryoten veel eenvoudiger: één enkel proteïne dat een draad vormt, aangedreven door een roterende moleculaire motor (stator + rotor).
basaalkorrel
Verankering van ciliën en flagellen, structureel vrijwel identiek aan een centriool (9 tripletten microtubuli). Functioneel uitwisselbaar met centriolen: centriolen kunnen evolueren naar basaalkorrels en omgekeerd.

vacuole.
Groot vesikel dat het grootste deel van rijpe plantencellen inneemt. Omgeven door de tonoplast. Functies: turgordruk voor strekkingsgroei, waterreserve, lysosomale afbraak van afvalstoffen, opslag van reservestoffen, pigmenten (anthocyanen) en defensieve stoffen (alkaloïden). Ontstaat door samenvoegen van kleine vacuolen in jonge cellen.
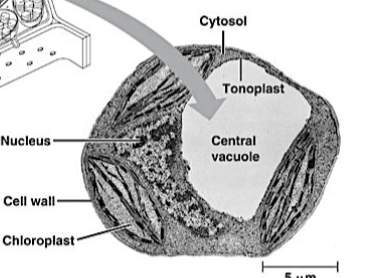
tonoplast
De enkelvoudige membraan die de vacuole omgeeft en afscheidt van het cytoplasma.
turgordruk
Osmotische druk opgebouwd in de vacuole door hoog gehalte aan opgeloste stoffen (±0,4–0,6 M). Trekt water aan via osmose en is de stuwende kracht achter de strekkingsgroei van plantencellen.
Ribosomen
Kleine partikels opgebouwd uit ribosomaal RNA en eiwitten. Komen vrij voor in het cytoplasma of gebonden aan het R.E.R. Eukaryote ribosomen: 80S (grote subeenheid 60S + kleine subeenheid 40S). Prokaryote ribosomen en die van mitochondriën/chloroplasten: 70S. Functie: eiwitsynthese.
Cytosol
Het vloeibare gedeelte van het cytoplasma. Bestaat uit een sterk geconcentreerde waterige oplossing van kleine moleculen (metabolische intermediairen) en macromoleculen (enzymen voor metabolisme).