Molekylær cellebiologi
1/25
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
26 Terms
Describe the general mechanism of GPCR activation and how GPCR signaling is terminated or desensitized.
GPCR-er er 7-transmembrane reseptorer som overfører ekstracellulære signaler til intracellulære signalveier via heterotrimere G-proteiner. G-proteinet består av Gα, Gβ og Gγ. I inaktiv tilstand er Gα bundet til GDP, og Gαβγ-komplekset ligger ved plasmamembranen.
Når ligand binder GPCR, endrer reseptoren konformasjon og fungerer som en GEF for Gα. Det betyr at GPCR får Gα til å slippe GDP, slik at GTP kan binde. Da dannes Gα-GTP, og både Gα-GTP og Gβγ kan regulere effektorproteiner som adenylyl cyclase, PLCβ, fosfodiesteraser eller ionekanaler.
Signalet avsluttes på flere nivåer. Gα hydrolyserer GTP til GDP, og dette akselereres av RGS-proteiner. Aktiv GPCR kan fosforyleres av GRK, som gjør at arrestin binder reseptoren. Arrestin hindrer videre G-proteinkobling og kan føre til internalisering av reseptoren. I tillegg fjernes second messengers, for eksempel ved at PDE bryter ned cAMP/cGMP, eller at Ca²⁺ pumpes ut av cytosol.
Process line:
Ligand → GPCR aktiveres → GDP byttes til GTP på Gα → Gα-GTP/Gβγ aktiverer effektorer → RGS, GRK/arrestin og fjerning av second messengers avslutter signalet
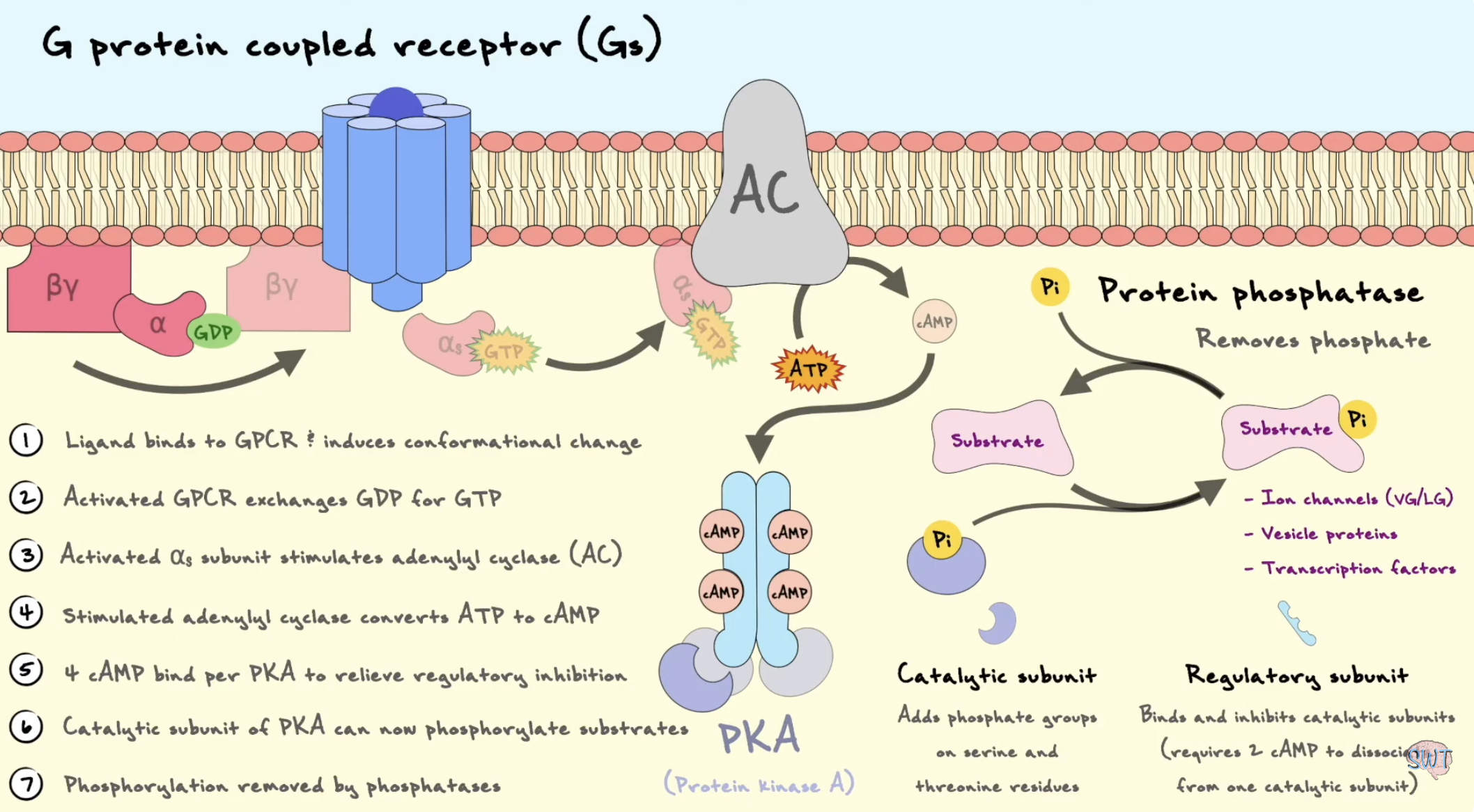
Describe how a Gs-coupled GPCR can regulate cAMP, PKA and gene expression.
Når et ligand aktiverer en Gs-koblet GPCR, fungerer reseptoren som GEF for Gαs. Gαs slipper GDP og binder GTP. Aktiv Gαs-GTP stimulerer adenylyl cyclase i plasmamembranen. Adenylyl cyclase omdanner ATP til cAMP.
cAMP fungerer som en second messenger og aktiverer PKA, protein kinase A. I inaktiv tilstand holdes PKA sine katalytiske subenheter bundet av regulatoriske subenheter. Når cAMP binder de regulatoriske subenhetene, frigjøres de katalytiske subenhetene og blir aktive.
PKA kan fosforylere målproteiner i cytosol, men kan også gå inn i kjernen. Der kan PKA fosforylere CREB, en transkripsjonsfaktor. Fosforylert CREB binder CRE-sekvenser i DNA og rekrutterer koaktivatorer, slik at transkripsjon av målgenene øker. Dermed kan et signal utenfra cellen føre til endret genuttrykk.
Process line:
Gs-GPCR → Gαs-GTP → adenylyl cyclase → cAMP ↑ → PKA aktiveres → CREB fosforyleres → genuttrykk endres
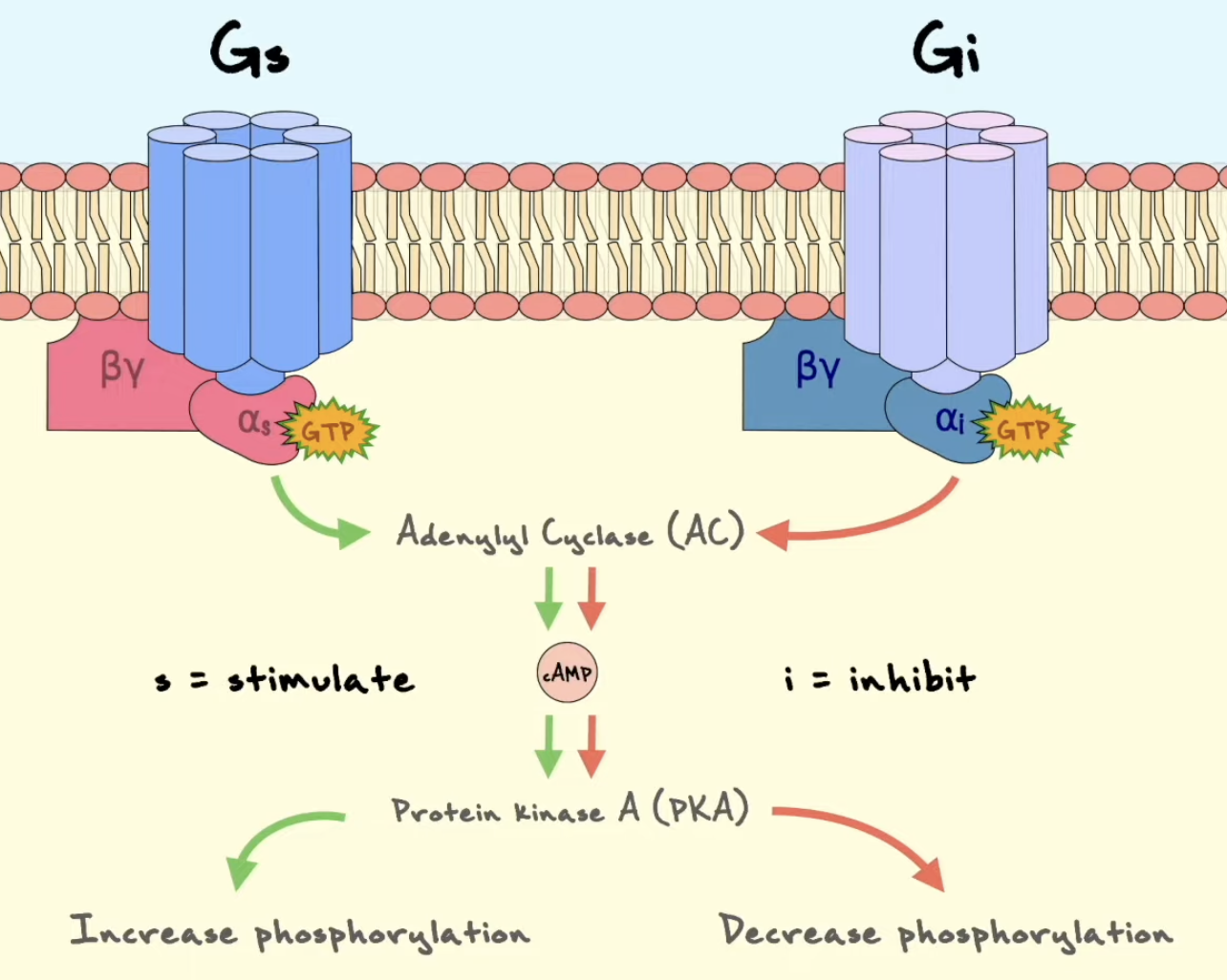
Explain how Gi-coupled GPCR signaling affects cAMP levels, and contrast this with Gs-coupled signaling.
Ved Gi-koblet GPCR-signalering aktiveres Gαi når GPCR fungerer som GEF og stimulerer GDP-GTP-utveksling. Aktiv Gαi-GTP hemmer adenylyl cyclase. Dette gir redusert produksjon av cAMP fra ATP.
Når cAMP-nivået faller, aktiveres mindre PKA. Dermed blir det mindre fosforylering av PKA-målproteiner, inkludert eventuelle transkripsjonsfaktorer som CREB. Gi-signalering virker derfor ofte som en brems på cAMP/PKA-responser.
Dette står i kontrast til Gs-signalering, der Gαs-GTP stimulerer adenylyl cyclase, slik at cAMP øker og PKA aktiveres. Gs og Gi kan derfor gi motsatt regulering av samme second messenger-system.
Process line:
Gs → adenylyl cyclase aktiveres → cAMP øker → PKA aktiveres
Gi → adenylyl cyclase hemmes → cAMP synker → PKA-aktivitet reduseres
Explain how cholera toxin and pertussis toxin affect GPCR/cAMP signaling. Include how cholera toxin enters the cell, the target cell type, and the molecular target.
Koleratoksin virker særlig på tarmepitelceller. Det er et AB-toksin. B-subenhetene binder GM1-gangliosid, et glykolipid i plasmamembranen. Dette gjør at toksinet tas opp ved endocytose. Toksinet transporteres videre retrogradt via endosomer og Golgi til ER, og A-subenheten kommer derfra over i cytosol. A-subenheten er enzymatisk aktiv.
I cytosol ADP-ribosylerer A-subenheten Gαs. Dette hemmer Gαs sin evne til å hydrolysere GTP til GDP. Dermed blir Gαs låst i aktiv GTP-bundet form. Aktiv Gαs stimulerer adenylyl cyclase kontinuerlig, slik at cAMP øker kraftig. I tarmepitel fører dette til PKA-aktivering, økt Cl⁻-sekresjon og vannsekresjon til tarmlumen. Resultatet er kraftig vandig diaré.
Pertussistoksin påvirker Gαi. Det ADP-ribosylerer Gαi slik at Gαi ikke kan aktiveres av GPCR. Gαi forblir dermed inaktivt og kan ikke hemme adenylyl cyclase. Resultatet er at cellen mister Gi-bremsen på cAMP-produksjon.
Kort skille:
Koleratoksin: Gαs låses aktiv → adenylyl cyclase konstant aktiv → cAMP opp.
Pertussistoksin: Gαi låses inaktiv → adenylyl cyclase hemmes ikke → cAMP opp.
Viktig presisering:
Begge toksinene ADP-ribosylerer en Gα-subenhet, men effekten blir ulik fordi de modifiserer ulike G-proteiner og ulike funksjonelle steder: koleratoksin ødelegger av-knappen på Gαs, mens pertussistoksin hindrer på-knappen på Gαi.
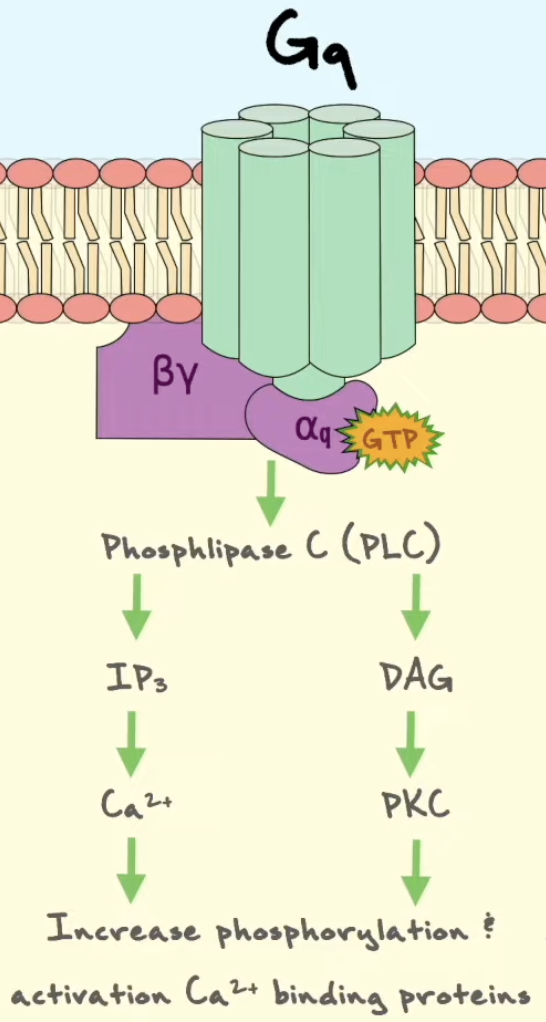
Describe how a Gq-coupled GPCR activates the IP3/DAG/Ca²⁺ pathway.
Når et ligand aktiverer en Gq-koblet GPCR, fungerer reseptoren som GEF for Gαq. Gαq slipper GDP og binder GTP. Aktiv Gαq-GTP aktiverer PLCβ, phospholipase Cβ, i plasmamembranen.
PLCβ spalter membranlipidet PIP2 til to second messengers: IP3 og DAG. IP3 er vannløselig og diffunderer gjennom cytosol til ER. Der binder IP3 til IP3-styrte Ca²⁺-kanaler i ER-membranen. Kanalene åpnes, og Ca²⁺ frigjøres fra ER til cytosol.
DAG blir værende i plasmamembranen. Sammen med økt Ca²⁺ kan DAG aktivere PKC, protein kinase C. Ca²⁺ kan også aktivere andre signalveier, særlig via calmodulin.
Process line:
Gq-GPCR → Gαq-GTP → PLCβ aktiveres → PIP2 spaltes til IP3 + DAG → IP3 åpner Ca²⁺-kanaler i ER → Ca²⁺ øker i cytosol → PKC og calmodulin-veier aktiveres
Explain how Ca²⁺ activates calmodulin and CaM-kinases, and how calcium signaling is terminated.
Ca²⁺ fungerer som en second messenger fordi cytosolisk Ca²⁺ normalt holdes lavt, men kan øke raskt ved signalering. Når Ca²⁺ frigjøres fra ER, kan det binde calmodulin, som er et Ca²⁺-sensorprotein. Når Ca²⁺ binder calmodulin, endrer calmodulin form og kan aktivere målproteiner.
Et viktig mål er Ca²⁺/calmodulin-dependent protein kinase, særlig CaMKII. CaMKII kan aktiveres av Ca²⁺/calmodulin. Ved gjentatte Ca²⁺-pulser kan CaMKII-subenheter autofosforylere hverandre. Dette gjør at CaMKII kan forbli aktiv en kort periode selv etter at Ca²⁺ faller. Dette kalles en form for molecular memory.
Ca²⁺-signalet avsluttes ved at Ca²⁺ fjernes fra cytosol. SERCA pumper Ca²⁺ tilbake til ER. PMCA pumper Ca²⁺ ut av cellen. Na⁺/Ca²⁺-exchanger bidrar også til å fjerne Ca²⁺, og mitokondrier kan ta opp Ca²⁺ via MCU. Samtidig kan GPCR-signalet stoppes ved GRK/arrestin på reseptornivå og RGS-proteiner på G-proteinnivå.
Process line:
Ca²⁺ ↑ → calmodulin aktiveres → CaMKII aktiveres → Ca²⁺-pulser kan gi autofosforylering/memory → SERCA, PMCA, Na⁺/Ca²⁺-exchanger og MCU fjerner Ca²⁺
Smell relies on olfactory GPCRs. Describe how an odorant molecule activates signal transduction in olfactory receptor neurons, and explain the role of cyclic nucleotide-gated ion channels.
I luktesansen binder et odorant molecule til en olfactory GPCR på en olfaktorisk sansecelle. Reseptoren aktiverer det spesifikke G-proteinet Golf. Aktiv Gαolf-GTP stimulerer adenylyl cyclase, som øker nivået av cAMP.
I olfaktoriske nevroner virker cAMP direkte på cyclic nucleotide-gated cation channels i plasmamembranen. Når cAMP binder kanalene, åpnes de, og Na⁺ og Ca²⁺ strømmer inn i cellen. Dette depolariserer membranen og kan starte et nervesignal. Ca²⁺ som kommer inn, kan også åpne Ca²⁺-aktiverte Cl⁻-kanaler. Fordi Cl⁻-konsentrasjonen er relativt høy inne i olfaktoriske sanseceller, kan Cl⁻ strømme ut og forsterke depolariseringen.
Process line:
Odorant → olfactory GPCR → Golf → adenylyl cyclase → cAMP ↑ → cAMP-gated cation channels åpner → Na⁺/Ca²⁺ inn → depolarisering → Ca²⁺-aktiverte Cl⁻-kanaler forsterker signalet
Vision relies on photoreceptor GPCR signaling. Describe how light activates rhodopsin signaling in rod photoreceptors, and explain how cyclic nucleotide-gated ion channels are affected.
I stav-fotoreseptorer er rhodopsin en lysfølsom GPCR. I mørke er cGMP-nivået relativt høyt, og cGMP-styrte kationkanaler er åpne. Da strømmer Na⁺ og Ca²⁺ inn i fotoreseptoren.
Når lys absorberes, isomeriseres 11-cis retinal til all-trans retinal inne i rhodopsin. Dette endrer rhodopsin og aktiverer G-proteinet transducin. Aktiv Gαt-GTP aktiverer cGMP phosphodiesterase, PDE. PDE bryter ned cGMP til GMP, slik at cGMP-nivået faller.
Når cGMP faller, lukkes de cGMP-styrte kationkanalene. Mindre Na⁺ og Ca²⁺ strømmer inn, og fotoreseptoren hyperpolariseres. Dette reduserer frigjøring av neurotransmitter. Dette er motsatt av lukt: i lukt åpner cAMP kanaler og depolariserer cellen, mens i syn fører lys til redusert cGMP, kanal-lukking og hyperpolarisering.
Process line:
Lys → 11-cis retinal blir all-trans retinal → rhodopsin aktiveres → transducin → PDE → cGMP brytes ned → cGMP-gated channels lukker → hyperpolarisering → mindre neurotransmitterfrigjøring
Receptor tyrosine kinases can activate MAP-kinase signaling and promote cell division. Describe the pathway from ligand binding to changes in gene expression and G1/S progression.
RTK-er aktiveres ofte av growth factors eller mitogens. Ligandbinding fører vanligvis til dimerization av reseptorene, slik at de cytosoliske tyrosine kinase domains aktiveres og fosforylerer hverandre på tyrosinrester. Dette kalles trans-autophosphorylation.
De fosforylerte tyrosinene fungerer som bindingssteder for signalproteiner med SH2 eller PTB domains, og reseptoren blir en signaling platform. Adapterproteinet Grb2 kan binde reseptoren og rekruttere Sos, som er en RasGEF. Sos aktiverer Ras ved å stimulere GDP-GTP-utveksling, slik at Ras-GTP dannes.
Aktiv Ras-GTP aktiverer MAPK-kaskaden: Raf/MAPKKK → MEK/MAPKK → ERK/MAPK. Aktiv ERK går inn i kjernen og fosforylerer transkripsjonsfaktorer som aktiverer immediate early genes, inkludert gener som fører til Myc-uttrykk.
Myc stimulerer uttrykk av gener som fremmer G1-progresjon, særlig G1 cyclins. G1-cykliner aktiverer G1-Cdks, som fosforylerer Rb. Fosforylert Rb slipper E2F, og E2F aktiverer gener som trengs for G1/S transition, DNA-replikasjon og S-fase.
Process line:
Growth factor → RTK dimerization → trans-autophosphorylation → SH2/PTB proteins → Grb2/Sos → Ras-GTP → Raf → MEK → ERK → Myc → G1 cyclins → G1-Cdk → Rb-P → E2F → G1/S
Explain how Ras is normally regulated by RasGEFs and RasGAPs, and why Ras mutations can cause cancer.
Ras er en liten monomeric GTPase som fungerer som en molekylær bryter. Ras er inaktiv med GDP og aktiv med GTP.
RasGEFs, for eksempel Sos, skrur Ras på ved å stimulere frigjøring av GDP, slik at GTP kan binde. RasGAPs skrur Ras av ved å stimulere Ras sin GTPase activity, slik at Ras hydrolyserer GTP til GDP.
Kreftaktiverende Ras-mutasjoner er ofte point mutations i eller nær det catalytic site i Ras. Disse mutasjonene svekker GTP-hydrolysen. Ras blir derfor værende som constitutively active Ras-GTP, selv når RasGAP er til stede.
Dette gir vedvarende aktivering av Raf-MEK-ERK, som øker uttrykk av Myc, G1 cyclins og andre proliferasjonsgener. Mutert Ras kan virke dominant gain-of-function, fordi én mutert allel kan produsere nok aktiv Ras-GTP til å drive signalering.
Process line:
RasGEF/Sos → Ras-GTP ON
RasGAP → Ras-GDP OFF
Ras point mutation → GTP hydrolysis fails → Ras-GTP stays ON → MAPK signaling ↑ → proliferation ↑
Describe how RTK-Ras-MAPK signaling is normally terminated or limited, and explain why scaffold proteins and feedback regulation are important.
RTK/Ras/MAPK-signalering må begrenses for at vekstsignaler ikke skal bli for sterke eller langvarige.
På reseptornivå kan RTK-signalet dempes ved dephosphorylation av fosfotyrosiner, endocytosis, receptor downregulation eller degradering. Når fosfotyrosinene fjernes, mister SH2/PTB proteins bindingsstedene sine, og signaleringskomplekset svekkes.
På Ras-nivå stimulerer RasGAPs hydrolyse av GTP til GDP, slik at Ras-GDP dannes og Raf ikke lenger aktiveres. På MAPK-nivå kan protein phosphatases defosforylere Raf, MEK, ERK eller ERK sine målproteiner.
Negative feedback kan også dempe tidligere trinn i signalveien. Scaffold proteins kan holde Raf, MEK og ERK sammen i en modul. Dette gir raskere og mer spesifikk signalering og reduserer crosstalk, særlig fordi celler har flere parallelle MAPK-moduler.
Process line:
RTK defosforyleres/internaliseres → SH2/PTB-binding forsvinner → RasGAP slår av Ras → phosphatases slår av Raf/MEK/ERK → feedback begrenser signalet
Compare normal RTK-Ras-MAPK signaling with oncogenic Ras signaling. Why does oncogenic Ras bypass normal growth factor control?
Ved normal RTK/Ras/MAPK-signalering trenger cellen et ekstracellulært signal, for eksempel en growth factor eller et mitogen. Dette aktiverer RTK, gir dimerization, trans-autophosphorylation, rekruttering av SH2/PTB-proteiner og aktivering av RasGEF/Sos. Ras aktiveres midlertidig som Ras-GTP og skrus deretter av av RasGAPs.
Ved oncogenic Ras er Ras selv mutert. Mutasjonen svekker evnen til å hydrolysere GTP til GDP. Dermed blir Ras værende som aktiv Ras-GTP, selv uten aktiv RTK og uten vekstfaktor.
Forskjellen er altså at normal Ras er avhengig av upstream signaler, mens onkogen Ras fungerer som en konstant aktiv “på-bryter”. Dette gir vedvarende Raf-MEK-ERK signaling, økt Myc, økte G1 cyclins, Rb-fosforylering og E2F-aktivitet. Derfor kan cellen få proliferasjonssignal selv når omgivelsene ikke gir klarsignal til celledeling.
Process line:
Normal: growth factor → RTK → Ras-GTP midlertidig → RasGAP slår av
Onkogen: Ras mutation → GTP hydrolysis fails → Ras-GTP konstant → MAPK aktiv uten growth factor
A mitogen stimulates a cell to enter the cell cycle. Explain how extracellular growth signals can be connected to cyclin-Cdk activity, Rb phosphorylation and E2F activation.
Et mitogen eller en growth factor kan stimulere cellecyklusinngang ved å aktivere RTK/Ras/MAPK-signalering. Når RTK aktiveres, dannes et signaleringskompleks som aktiverer Ras-GTP via Grb2/Sos. Ras aktiverer MAPK-kaskaden Raf → MEK → ERK.
Aktiv ERK går inn i kjernen og fosforylerer transkripsjonsfaktorer. Dette aktiverer immediate early genes, der Myc er et viktig eksempel. Myc-proteinet fungerer som transkripsjonsfaktor og øker uttrykk av gener som trengs for cellevekst og G1-progresjon, særlig G1 cyclins.
G1 cyclins, for eksempel cyclin D, binder Cdk4/6 og danner aktive G1-Cdk complexes. Disse fosforylerer Rb-proteinet. Ufosforylert Rb binder og hemmer E2F. Når Rb fosforyleres, frigjøres E2F.
Aktiv E2F stimulerer uttrykk av gener som trengs for S-phase entry, inkludert G1/S cyclins, S cyclins, DNA-replikasjonsproteiner og E2F selv. Dette gir positive feedback-loops som driver cellen forbi restriction point og inn i S-fase.
Process line:
Mitogen → RTK → Ras → Raf → MEK → ERK → Myc → G1 cyclins/Cdk4/6 → Rb phosphorylation → E2F release → G1/S cyclins + S-phase genes → cell cycle entry
Cadherins play central roles in development and tissue organization. Describe how cadherin-mediated cell-cell adhesion works, how cadherins are linked to the cytoskeleton, and how this contributes to tissue organization.
Cadheriner er vanligvis single-pass transmembrane adhesion proteins som medierer Ca²⁺-dependent cell-cell adhesion. De ekstracellulære domenene har flere cadherin repeats, og Ca²⁺-binding gjør domenet mer stivt og rod-like. Dette gjør at cadheriner kan binde cadheriner av samme eller nært beslektet type på naboceller. Dette kalles homophilic binding.
Én cadherin-cadherin-binding er relativt svak, men mange cadheriner samles i clusters, slik at adhesjonen blir sterkere. Dette kalles ofte Velcro principle. Cadherin-clustering kan reguleres av Rho-family GTPases.
I adherens junctions kobles klassiske cadheriner indirekte til actin cytoskeleton via p120-catenin, β-catenin, α-catenin og vinculin. I epitelceller kan dette danne et adhesion belt like under den apikale siden, der aktinnettverket kobles mellom naboceller. Dette gir mekanisk sammenheng i vevet og bidrar til celleform, morfogenese og vevsorganisering.
Process line:
Ca²⁺ binder cadherin → homophilic cadherin binding → cadherin clusters/Velcro → catenins/vinculin → actin linkage → adhesion belt → tissue organization
Explain how cadherin junctions can function as tension sensors, and why cadherin expression is important for cell sorting, EMT and cancer invasion.
Cadherin-baserte adherens junctions kan fungere som tension sensors. Når mekanisk kraft virker på junctionen, kan α-catenin strekkes ut og eksponere bindingssteder for vinculin. Vinculin rekrutterer flere koblinger til actin, slik at junctionen blir sterkere når den belastes.
Cadheriner bidrar også til cell sorting. Celler som uttrykker samme cadherintype binder bedre til hverandre enn til celler med andre cadheriner. Dette hjelper celler å sortere seg i riktige vev under utvikling.
I kreft og utvikling er E-cadherin viktig. Tap eller nedregulering av E-cadherin svekker epitelial cell-cell adhesion. Transkripsjonsfaktorer som Snail og Twist kan redusere E-cadherin-uttrykk og fremme epithelial-mesenchymal transition, EMT. Da mister celler epiteliale egenskaper, blir mer migrerende og kan lettere invadere omkringliggende vev.
Process line:
Tension → α-catenin strekkes → vinculin/actin rekrutteres → junction styrkes
Snail/Twist ↑ → E-cadherin ↓ → EMT → migrasjon/invasjon ↑
Compare adherens junctions, desmosomes, focal adhesions and hemidesmosomes. Include the adhesion molecules, cytoskeletal linkage and main function.
Adherens junctions er cell-cell anchoring junctions. De bruker classical cadherins og kobles via catenins/vinculin til actin filaments. De er viktige for celleform, vevsorganisering og dynamisk mekanisk kobling mellom naboceller.
Desmosomes er også cell-cell anchoring junctions, men de bruker desmosomale cadheriner og kobles til intermediate filaments, ofte keratin i epitelceller. De gir sterk mekanisk motstand i vev som utsettes for strekk og slitasje.
Focal adhesions er cell-matrix junctions. De bruker integrins som binder ECM, for eksempel fibronectin, og kobles intracellulært til actin filaments via blant annet talin og vinculin. De er dynamiske og viktige for cellemigrasjon, mekanosensing og signalering via FAK/Src.
Hemidesmosomes er stabile cell-matrix junctions i epitel. De bruker særlig α6β4 integrin, binder laminin i basal lamina og kobles intracellulært til keratin intermediate filaments via plectin og BP230.
Kort husk:
Adherens = cadherin + actin.
Desmosome = cadherin + intermediate filaments.
Focal adhesion = integrin + actin.
Hemidesmosome = integrin + intermediate filaments.
Epithelial cells express matrix receptors that attach them to the basal lamina. Describe ECM, basal lamina, integrins, focal adhesions, hemidesmosomes, and outside-in/inside-out activation.
Extracellular matrix, ECM, er et nettverk av proteiner og polysakkarider utenfor cellene. ECM gir mekanisk støtte, organiserer vev og kan regulere signalering. Basal lamina er et spesialisert ECM-lag under epitelceller. Den inneholder blant annet laminin, type IV collagen, nidogen og perlecan/proteoglycans. Basal lamina gir mekanisk støtte, bidrar til epitelorganisering, fungerer som selektiv barriere/molekylært filter og kan fungere som scaffold ved vevsreparasjon.
Integrins er de viktigste reseptorene for celle-matrix-adhesjon. De er α/β transmembrane heterodimers. Utenfor cellen binder de ECM-komponenter som fibronectin, collagen eller laminin. Inne i cellen kobles de til cytoskjelettet og signalproteiner.
Ved outside-in activation binder ECM-ligand integrinet utenfra. Integrinet går fra bøyd/inaktiv til utstrakt/aktiv form, og cytoplasmatiske bindingssteder eksponeres. Da kan talin og vinculin koble integrinet til actin.
Ved inside-out activation kommer signalet fra innsiden av cellen. Intracellulære signalproteiner, for eksempel via Rap1-RIAM, rekrutterer talin til integrinets β-hale. Dette øker integrinets affinitet for ECM.
I focal adhesions kobles integriner til actin filaments og aktiverer signalering via FAK/Src. I hemidesmosomes kobles α6β4-integrin til keratin intermediate filaments og binder laminin i basal lamina.
Process line:
ECM/basal lamina → integrin aktiveres → talin/vinculin binder β-tail → cytoskeleton kobles på → focal adhesion eller hemidesmosome → adhesjon + signalering
Animal cell locomotion depends on actin regulation and focal adhesions. Describe the main steps in cell crawling and the key proteins involved.
Cellemigrasjon skjer i en syklus med protrusion, attachment, traction og rear detachment.
Først danner cellen protrusjoner i fronten, særlig lamellipodia. Dette drives av actin polymerization. Rac GTPase aktiverer Arp2/3 complex, som lager forgrenede actin-nettverk og skyver membranen fremover.
Deretter fester cellen seg til ECM gjennom integrins i focal adhesions. Integriner binder matrix utenfor cellen og kobles til actin inne i cellen via talin, vinculin og andre adapterproteiner. FAK/Src signaling bidrar til regulering av focal adhesions.
Så trekkes cellekroppen fremover ved traction. Dette skjer når Rho GTPase stimulerer actin stress fibers og myosin II-basert kontraksjon. Til slutt løsnes adhesjoner bak i cellen, slik at cellen kan bevege seg videre.
Retningen kan styres av chemotaxis, ECM-sammensetning eller adhesjonsmolekyler som ICAMs/VCAMs på endotel.
Process line:
External cue → Rac → Arp2/3 → lamellipodium → integrin/focal adhesion → Rho/myosin II traction → rear detachment → cell movement
Describe the main non-anchoring junctions and adhesion systems: tight junctions, gap junctions, plasmodesmata, selectins and ICAM/VCAM-mediated leukocyte adhesion.
Tight junctions forsegler rommet mellom epitelceller og begrenser paracellular transport. De bidrar også til apical-basal polarity ved å hindre at membranproteiner diffunderer fritt mellom apikal og basolateral membran. Viktige proteiner er claudins, occludins og scaffold-proteiner.
Gap junctions finnes hos dyreceller og danner kanaler mellom cytoplasma i naboceller. De bygges av connexins som danner connexons. De tillater passasje av små molekyler og ioner, og er viktige for koordinert signalering, for eksempel i hjertemuskel.
Plasmodesmata finnes i planteceller og danner cytoplasmatiske kanaler gjennom celleveggen. Funksjonelt ligner de gap junctions fordi de muliggjør direkte celle-celle-kommunikasjon.
Selectins medierer svak og transient adhesjon, særlig mellom leukocytter og endotel. De binder spesifikke oligosakkarider på glycoproteins/glycolipids og gjør at leukocytter kan rulle langs blodåreveggen. Deretter aktiveres integriner på leukocytten, for eksempel LFA-1, som binder ICAM-1 på endotelceller. Dette gir sterk adhesjon og gjør at leukocytten kan krype ut av blodkaret.
Process line:
Selectin weak binding → leukocyte rolling → integrin activation → LFA-1 binds ICAM-1 → firm adhesion → crawling/extravasation
How do DNA regulatory proteins control transcription of specific eukaryotic genes?
Eukaryot transkripsjon reguleres av DNA regulatory proteins/transcription factors som binder spesifikke cis-regulatory DNA sequences. Slike sekvenser kan være enhancers, silencers, insulators eller promoter-nære regulatoriske elementer. Promoteren er området der general transcription factors, Mediator og RNA polymerase II danner et pre-initiation complex.
Activators binder ofte enhancers, som kan ligge langt unna målgenet, noen ganger hundretusener av basepar unna. Ved DNA looping kan enhancer-bundne activators komme i fysisk kontakt med promoterområdet. Activators kan rekruttere Mediator, co-activators, chromatin remodeling complexes, histone-modifying enzymes og histone chaperones. Dette kan øke tilgjengeligheten av DNA og stabilisere transkripsjonsmaskineriet ved promoteren.
Mediator er et multiproteinkompleks som formidler kontakt mellom enhancer-bundne activators og det basale transkripsjonsmaskineriet ved promoteren. Det bidrar til rekruttering/stabilisering av RNA polymerase II, general transcription factors og pre-initiation complex, og kan bidra til aktivering eller re-initiering av transkripsjon.
Repressors kan hemme transkripsjon ved å konkurrere med activators om DNA-bindingssteder, binde activators og maskere deres activation domain, eller rekruttere co-repressors. Co-repressors kan rekruttere HDACs, histone methyltransferases og DNA methylases, som kan bidra til heterochromatin formation og redusert transkripsjon.
Combinatorial control betyr at det er den samlede effekten av activators og repressors, og deres konsentrasjon i cellen, som avgjør om et gen uttrykkes.
Process line:
Cis-regulatory element → activators/repressors binder → DNA looping/cooperativity → Mediator/co-regulators/chromatin remodeling → pre-initiation complex/RNA pol II reguleres → gen på eller av
How can transcription regulators open or compact chromatin?
I eukaryote celler er DNA pakket rundt histoner i nucleosomes, som danner chromatin. Dette begrenser tilgangen for transcription factors og RNA polymerase II. Derfor må chromatin ofte omorganiseres før et gen kan uttrykkes.
Activators kan rekruttere ATP-dependent chromatin remodeling complexes, som flytter, løsner eller omorganiserer nucleosomes. Dette kan eksponere promoter- eller enhancerregioner. Nucleosome breathing kan gi midlertidige åpninger i chromatin, slik at DNA-bindende regulatoriske proteiner kan få tilgang. Når de først binder, kan de rekruttere flere proteiner og stabilisere en mer åpen chromatin state.
Activators kan også rekruttere histone acetyltransferases, HATs. HATs acetylerer histonhaler, svekker interaksjonen mellom histoner og DNA og gjør chromatin mer tilgjengelig. Histone acetylation kan også gjøre det lettere for histone chaperones å fjerne eller bytte ut histoner.
Repressors virker ofte motsatt. De kan rekruttere co-repressors, som igjen rekrutterer HDACs. HDACs fjerner acetylgrupper fra histoner og gjør chromatin mer kompakt. Repressors kan også rekruttere histone methyltransferases, som kan legge repressive histonmerker.
DNA methylation av cytosin i CpG-sekvenser kan bidra til tettere chromatin. Methyl-CpG-binding proteins kan binde metylerte CpG-er og rekruttere co-repressors og histonmodifiserende enzymer. Dette kan etablere stabil genavstenging.
Process line:
Activator → chromatin remodeling/HAT/histone chaperones → åpent chromatin/euchromatin → transkripsjon opp
Repressor → HDAC/histone methyltransferase/DNA methylation → kompakt chromatin/heterochromatin → transkripsjon ned
How can combinations of transcription factors generate spatial gene expression patterns during development?
Romlig genuttrykk under utvikling oppstår ofte gjennom combinatorial control, der en cis-regulatory module integrerer signalet fra flere activators og repressors. Det er altså ikke én transcription factor alene som bestemmer uttrykket, men kombinasjonen av hvilke regulatorer som er til stede, hvor mye det finnes av dem, og hvilke bindingssteder modulen inneholder.
Drosophila even-skipped, Eve, er et klassisk eksempel. Tidlig i Drosophila-utvikling dannes et syncytial blastoderm, der mange kjerner deler samme cytoplasma. Transkripsjonsregulatorer danner gradienter langs anterior-posterior-aksen. Ulike kjerner “leser” derfor ulike konsentrasjoner av regulatoriske proteiner.
Eve-genet uttrykkes i syv striper. Dette skyldes at Eve-genet har flere regulatory modules, der hver modul kontrollerer én stripe. For Eve stripe 2 fungerer Bicoid og Hunchback som activators, mens Giant og Krüppel fungerer som repressors. Stripe 2 uttrykkes bare der Bicoid/Hunchback er tilstrekkelig høye, samtidig som Giant/Krüppel ikke dominerer.
Dette viser at en enhancer/regulatory module kan fungere som et tolkningssystem for posisjonsinformasjon. Hvis activator/repressor-kombinasjonen er riktig, aktiveres transkripsjon. Hvis repressors er til stede i feil område, blir genet av. Eve stripe 2 kan også studeres eksperimentelt ved å koble stripe 2-regulatorisk modul til et LacZ reporter gene, slik at man ser hvor modulen driver genuttrykk.
Process line:
Morfogengradienter → ulik TF-konsentrasjon i ulike kjerner → cis-regulatory module leser kombinasjonen → Eve stripe 2 på bare i riktig område
How can epigenetic mechanisms establish cell memory?
Cell memory betyr at en celle opprettholder sitt genuttrykksmønster og sin differensierte identitet gjennom senere celledelinger. Dette er viktig fordi en differensiert celle må “huske” hvilke gener som skal være aktive og inaktive etter mitose.
Cell memory kan etableres gjennom transcriptional circuits, særlig positive feedback loops, der en transcription factor aktiverer sitt eget uttrykk eller aktiverer andre faktorer som opprettholder samme genprogram. Mange Hox genes kan for eksempel bidra til langvarig celleminne ved å aktivere sin egen transkripsjon, og Hox-regulering er knyttet til Polycomb- og Trithorax-regulert chromatinstruktur.
Epigenetiske mekanismer stabiliserer slike genprogrammer. DNA methylation kan arves etter DNA-replikasjon. Når DNA kopieres, blir den nye tråden først umetylerte. Maintenance DNA methyltransferase kan gjenkjenne hemimetylerte CpG-steder og metylere den nye tråden, slik at metyleringsmønsteret videreføres.
Histone modifications kan også bidra til epigenetisk arv. Under DNA-replikasjon kan modifiserte histoner fordeles til datter-DNA, og reader-writer proteins kan gjenopprette samme histonmodifikasjoner på nye histoner. Pensum beskriver at datterceller kan arve både spesifikke chromatin modifications og DNA methylation patterns.
Polycomb group proteins bidrar ofte til å holde gener av, mens Trithorax group proteins bidrar til å holde gener aktive. Sammen gjør transcriptional circuits, DNA methylation, histone modifications og chromatin states at celler kan bevare identiteten sin etter celledeling.
Process line:
Cell fate signal → transcription factor network → positive feedback → DNA methylation/histone marks/Polycomb-Trithorax → chromatin state vedlikeholdes → cell identity arves
How is mRNA processed and regulated after transcription in eukaryotic cells?
I eukaryote celler dannes først pre-mRNA, som må prosesseres før det kan eksporteres og translateres. Post-transkripsjonell regulering kan påvirke både mRNA-stabilitet, proteinmengde og hvilke proteinvarianter som produseres.
Først får pre-mRNA en 5’ cap, vanligvis en 7-methylguanosine cap. Denne beskytter mRNA mot nedbrytning, bidrar til kjerneeksport og er viktig for translation initiation gjennom cap-binding proteins.
Deretter skjer splicing, der introns fjernes og exons settes sammen av spliceosome. Splice activators og splice repressors kan regulere hvilke splice sites som brukes. Ved alternative splicing kan ulike exons inkluderes eller ekskluderes, slik at ett gen kan gi flere mRNA-varianter og dermed flere proteinisoformer.
I 3’-enden skjer RNA cleavage and polyadenylation. CPSF gjenkjenner polyadenyleringssignalet, og CstF bidrar til kløyving. Deretter legges en poly-A tail til. Poly-A tail beskytter mRNA, påvirker mRNA lifetime og kan regulere translation efficiency.
mRNA editing kan endre RNA-sekvensen etter transkripsjon. ADAR kan utføre A-to-I editing, mens editosome-mediert editing kan gi C-to-U-endringer. Slike endringer kan påvirke kodende sekvens, stabilitet eller funksjon.
mRNA kan også reguleres av kjemiske modifikasjoner, for eksempel m6A, som kan påvirke mRNA-stabilitet og nedbrytning.
Process line:
pre-mRNA → 5’ cap → spliceosome/splice regulators → alternative splicing → CPSF/CstF cleavage → poly-A tail → RNA editing/m6A → modent og regulert mRNA
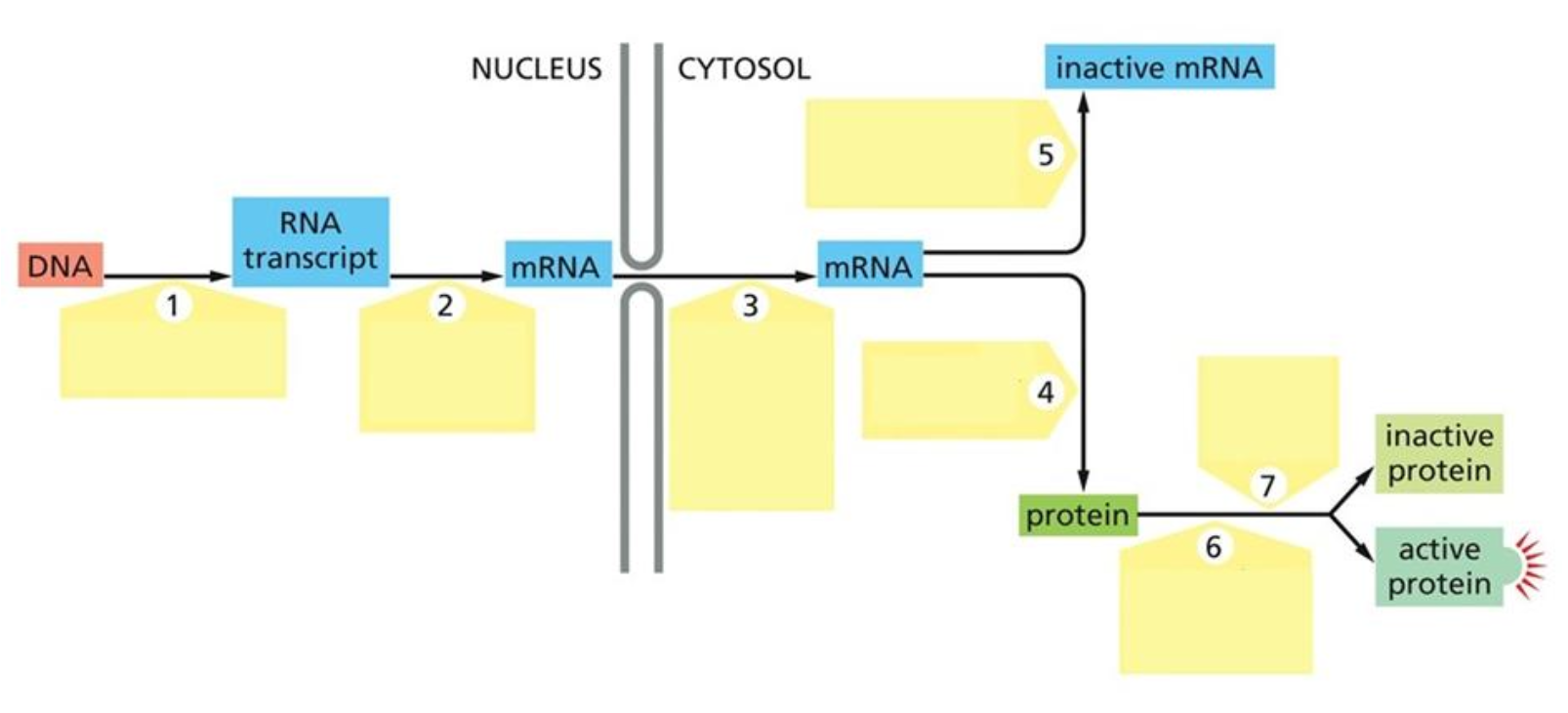
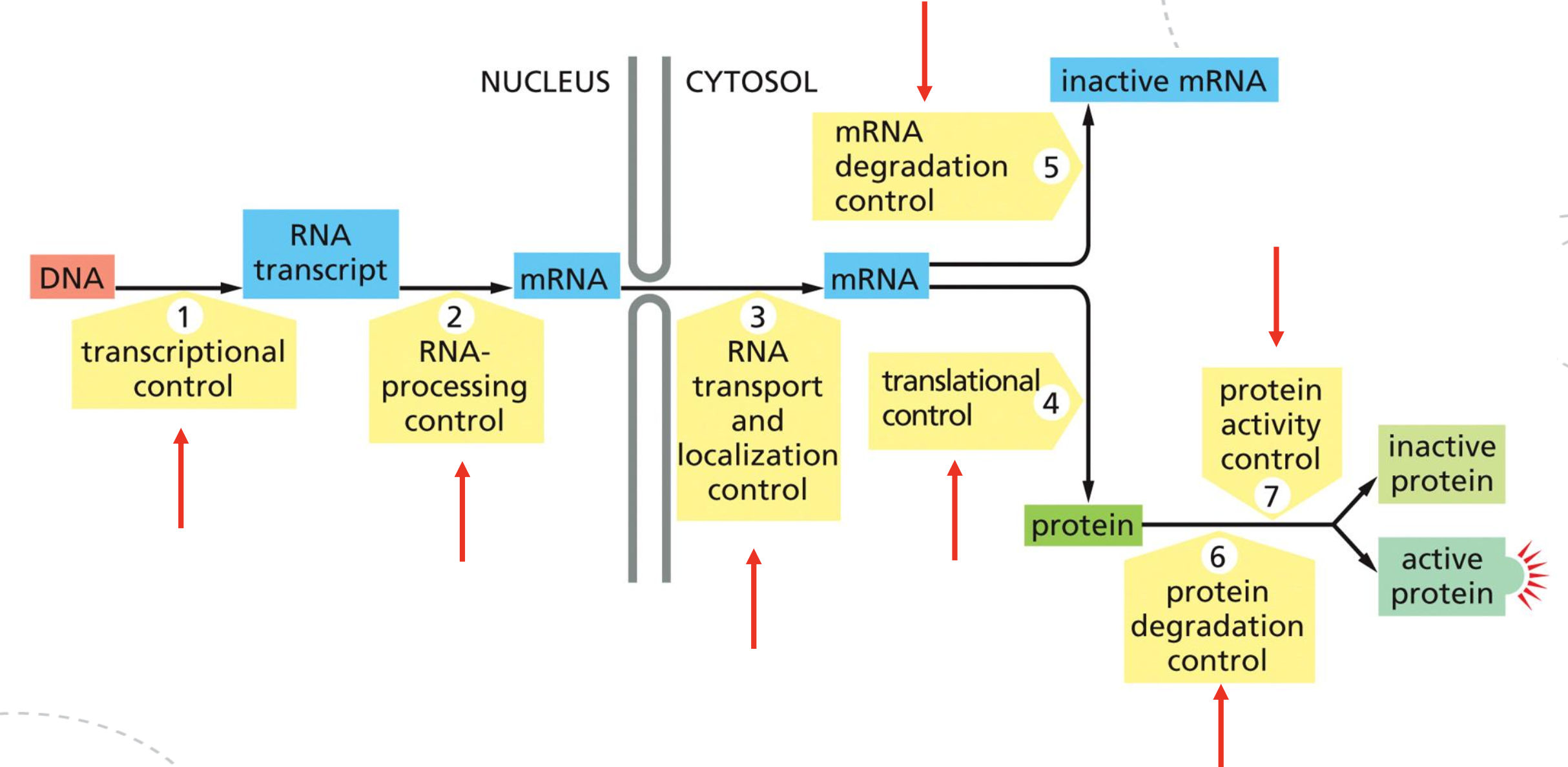
Where can gene expression be regulated between DNA and active protein? Explain the seven control points.
Eukaryot genuttrykk kan reguleres på flere nivåer fra DNA til aktivt protein.
1. Transcriptional control: regulerer om genet transkriberes og hvor mye RNA som lages. Involverer transcription factors, activators, repressors, cis-regulatory elements, promoter, Mediator, RNA polymerase II og chromatin.
2. RNA-processing control: regulerer hvordan pre-mRNA modnes. Involverer 5’ capping, splicing, alternative splicing, 3’ cleavage/polyadenylation og RNA editing.
3. RNA transport/localization control: regulerer eksport fra kjernen og hvor mRNA lokaliseres i cytosol. Involverer RNA-binding proteins, nuclear export, cytoskeleton transport og lokal translasjon.
4. Translational control: regulerer hvor effektivt mRNA translateres til protein. Involverer translation initiation, 5’ cap-binding complex, eIF2 cycle, IRES, alternative start sites og miRNA.
5. mRNA degradation control: regulerer mRNAets levetid. Involverer poly-A tail, 5’ cap, deadenylation, decapping, exonucleases, P-bodies og miRNA/Argonaute/RISC.
6. Protein degradation control: regulerer hvor lenge proteinet overlever. Involverer ubiquitin, polyubiquitination, proteasome, phosphorylation og SCF-Cullin/F-box.
7. Protein activity control: regulerer om proteinet er aktivt eller inaktivt. Involverer folding, chaperones, cofactors, phosphorylation, proteolytic processing, allosteric regulation og protein complexes.