HC3: Bacterial invasion
1/20
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No analytics yet
Send a link to your students to track their progress
21 Terms
Invasive bacteria
= bacteriën die niet alleen “lokaal” een infectie geven, maar door barrières heen breken en dieper het lichaam in kunnen gaan (bijvoorbeeld in het bloed, organen etc.) → daardoor ernstige/systemische ziekten veroorzaken
Listeria
→ kan vanuit de darm het lichaam binnendringen en via bloed verspreiden (veroorzaakt meningitis (hersenvliesontsteking))
→ bij zwangere sepsis (bloedvergiftiging)Salmonella
→ kan vanuit darm het lichaam in en via bloed verspreiden, veroorzaakt (para)tyfusShigella
→ infecteert vooral darmwand en veroorzaakt heftige ontstekingDysenterie = ernstige diarree, vaak met bloed/slijm en buikkrampen
Yersinia
→ klassieke verwekker van de pest
Invasion in host cell
= induction of phagocytosis-like processes in normally non-phagocytic cells by rearrangement of the cytoskeleton
Cytoskeleton of eukaryotic cells:
→ bestaat uit 3 types proteine filamenten
actin filaments (microfilamenten)
Dikte: Het dunst (ongeveer 7 nm).
Bouwsteen: Bestaat uit strengen van het eiwit actine.
Functie: Ze zijn flexibel maar sterk. Ze bepalen de vorm van het celoppervlak en zijn cruciaal voor celbeweging (zoals het kruipen van cellen) en spiercontractie. Bij gastheer-pathogeen interacties zie je vaak dat bacteriën (zoals Listeria) actine "kapen" om zich door de cel voort te bewegen.
intermediate filaments (intermediaire filamenten)
Dikte: Middelste maat (ongeveer 10 nm).
Bouwsteen: Varieert per celtype (bijvoorbeeld keratine in epitheelcellen of vimentine).
Functie: Ze werken als een soort trekkabels en geven de cel mechanische sterkte. Ze beschermen de cel en de celkern tegen openscheuren wanneer er fysieke druk of rek op komt te staan. Ze zijn heel stabiel en touwachtig gevlochten.
microtubules (microtubuli)
Dikte: Het dikst (ongeveer 25 nm).
Bouwsteen: Holle buisjes gemaakt van tubuline-dimeren
Functie: Ze fungeren als de snelweg van de cel waarover transporteiwitten (zoals kinesine en dyneïne) blaasjes en organellen verplaatsen. Daarnaast vormen ze de spoelfiguur tijdens de celdeling om chromosomen uit elkaar te trekken. Veel virussen gebruiken deze microtubuli-snelweg om na binnenkomst snel naar de celkern te reizen.
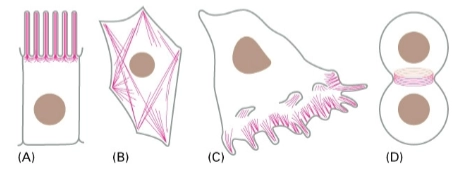
Actinefilamenten:
→ verschillende structuren in de cel
Microvilli
= vingerachtige uitsteeksel aan oppervlak van bijvoorbeeld darmepitheelcellen → actine vormt hier stijve, parallelle bundels die deze uitsteeksels omhooghouden om celoppervlak te vergrotenContractile bundles
= bundels van actine in cytoplasma die samen met motorcellen kunnen samentrekken (werken als spieren) → vormveranderingSheet-like and fingerlike protrusions
= wanneer een cel gaat ‘lopen’ of kruipenSheet-like (lamellipodia) zijn platte, brede uitlopers.
Fingerlike (filopodia) zijn dunne uitsteeksels waarmee de cel de omgeving aftast.
Contractile ring
= tijdens celdeling → vormt actine samen met myosine een ring in midden van de cel, deze ring trekt steeds strakker tot cel in 2en spiltst
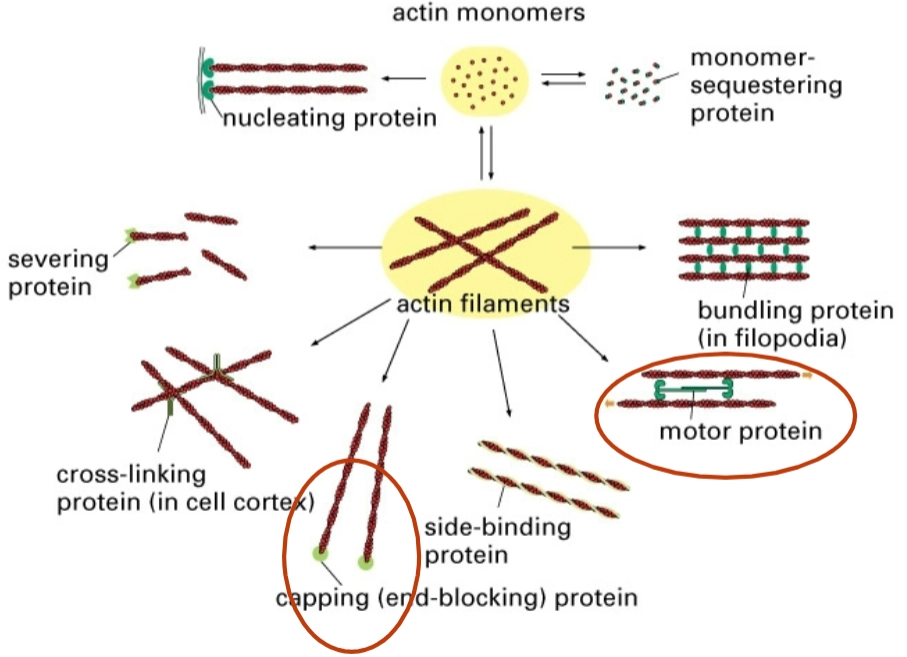
Actin Binding Proteins (ABP)
→ de eiwitten die bepalen wat er met het actine gebeurt:
Monomer-sequestering protein: Actine start als losse bouwstenen (actin monomers). Dit eiwit bindt aan de losse bouwstenen zodat ze tijdelijk niet aan een filament kunnen vastplakken. Zo houdt de cel een voorraadje achter de hand.
Nucleating protein: Helpt bij de start (nucleatie) van een nieuwe actineketen. Het zet de eerste bouwstenen in elkaar, waarna de keten snel kan groeien.
Severing protein: Dit eiwit knipt bestaande actinefilamenten in stukken. Hierdoor wordt het netwerk vloeibaarder óf ontstaan er juist meer losse uiteinden waar weer nieuwe groei kan plaatsvinden.
Cross-linking protein (in cell cortex): Knoopt actinefilamenten in een soort driedimensionaal web/netwerk aan elkaar, vlak onder het celmembraan (de celcortex). Dit geeft de cel stevigheid.
Capping (end-blocking) protein: Dit eiwit gaat als een soort 'dopje' op het uiteinde van een actinefilament zitten. Hierdoor stopt de groei of de afbraak aan die kant; het stabiliseert de lengte.
Side-binding protein: Bindt langs de zijkant van het filament om het te stabiliseren (bijvoorbeeld tropomyosine).
Motor protein: (Omcirkeld in het rood) Dit is meestal myosine. Het loopt over het actinefilament heen om blaasjes te transporteren of om actinefilamenten langs elkaar heen te trekken (samentrekking).
Bundling protein (in filopodia): (Ook rood omcirkeld) Zorgt ervoor dat actinefilamenten strak parallel aan elkaar gaan liggen, wat zorgt voor de stevige sprieten (filopodia) die je ook bij Dia 1 onder (C) zag.
Actin polymerization and de-polymerization
= shape and movement of cells
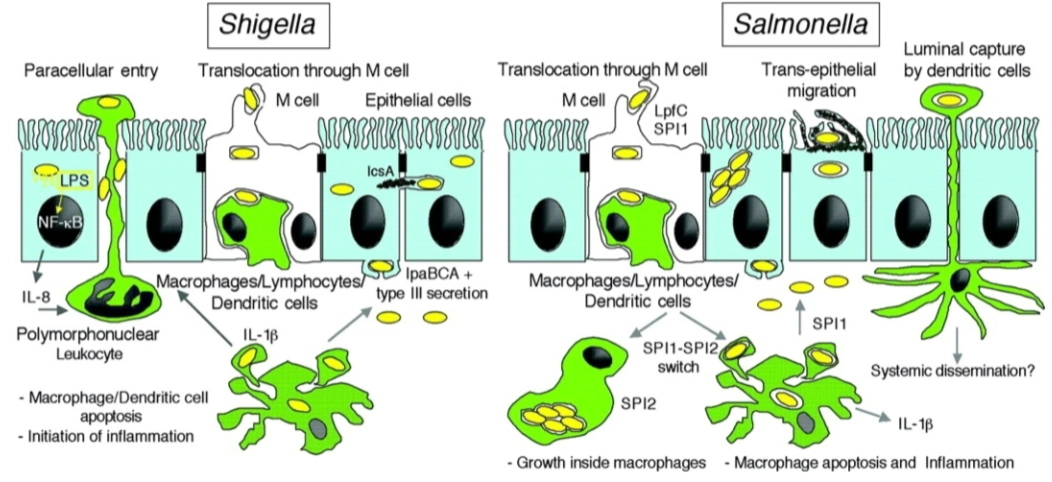
Entry mechanisms used by invasive bacteria:
→ Zipper-mechanism & Trigger-mechanism
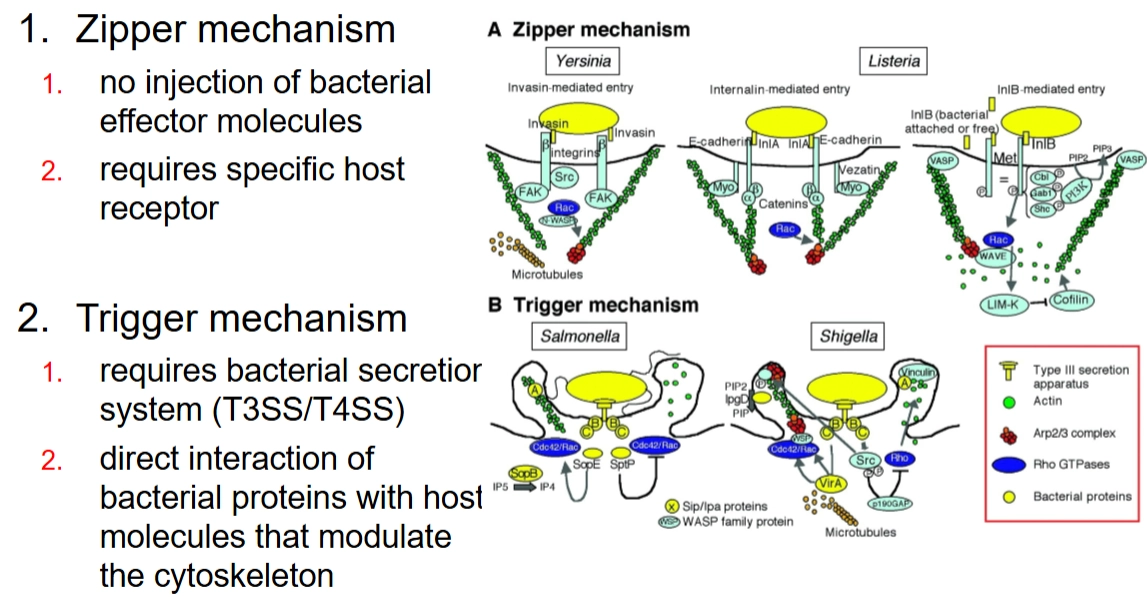
Zipper-mechanism
→ beperkte/bescheiden veranderingen in cytoskeletHoe het werkt: Dit mechanisme verloopt via de buitenkant van de cel. De bacterie heeft specifieke eiwitten op zijn oppervlak (bijv. invasine of internaline) die heel sterk binden aan normale receptoren (zoals cadherines of integrines) op de gastheercel.
Het effect: Er is in eerste instantie geen direct contact met de binnenkant van de cel. Maar doordat de eiwitten van de bacterie aan de receptoren binden, activeert de gastheercel zijn normale signaalroutes voor cel-celadhesie. Dit zet lokaal het actine-skelet in gang: het membraan kruipt langzaam als een "ritssluiting" om de bacterie heen (phagocytic cup closure), totdat de bacterie volledig is ingesloten en naar binnen wordt getrokken (bacterial internalization).
Voorbeelden: Listeria monocytogenes en Yersinia enterocolitica.
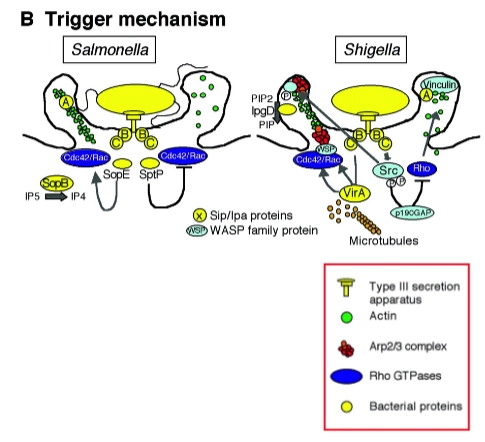
Trigger-mechanism
→ enorme/massale cytoskelet veranderingenHoe het werkt: Dit is een veel agressievere methode. De bacterie maakt direct contact met het actine-skelet aan de binnenkant van de cel. Dit doet hij door bacteriële effector-moleculen (eiwitten) rechtstreeks de gastheercel in te spuiten via een moleculaire injectienaald, zoals een Type III of Type IV secretiesysteem (T3SS/T4SS).
Het effect: Deze ingespoten eiwitten activeren direct en lokaal de nucleating proteins en andere actine-regulatoren (die we in de vorige dia's zagen). Hierdoor explodeert de actine-polymerisatie onder het membraan. Het celmembraan begint enorm te golven en uit te stulpen (ruffling and folding of membrane). De bacterie wordt door deze grote golven van het membraan als het ware overspoeld en naar binnen geslokt.
Voorbeelden: Salmonella enterica en Shigella flexneri.
→ doel is een ALTERED CELL SHAPE OR MOVEMENT
→ hierdoor reorganiseert de gastheercel zijn actinefilamenten zo dat de bacterie veilig naar binnen kan glippen
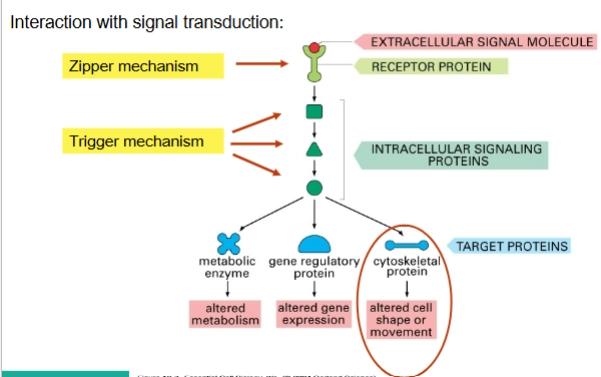
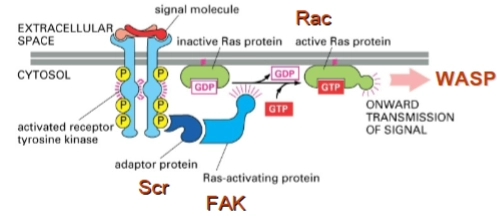
Zipper mechanism vs. Trigger mechanism
De Zipper-route: De gele pijl laat zien dat het Zipper-mechanisme helemaal bovenaan begint, bij de Receptor protein. De bacterie doet alsof hij een lichaamseigen extracellular signal molecule is en activeert de receptor vanaf de buitenkant. De normale cascade (de stapjes naar beneden) wordt daarna netjes doorlopen.
De Trigger-route: De drie gele pijlen laten zien dat het Trigger-mechanisme de bovenkant (de receptor) volledig overslaat. Omdat de bacterie zijn effector-eiwitten direct naar binnen spuit, grijpt hij direct in op de Intracellular signaling proteins (zoals de zogenaamde Rho-GTPases: Rac, Cdc42 of RhoA).
Eukaryote cell-oppervlakte receptoren
Ion-channel-linked receptor
G-protein-linked receptor
Enzyme-linked receptor
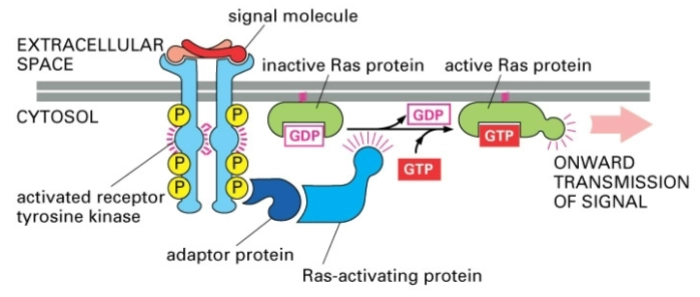
Andere receptor
Tyrosine kinase
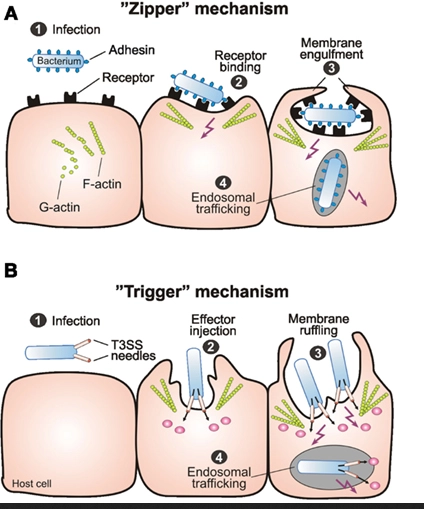
Zipper mechanism: 3 successive steps
Contact and adherence (Contact en hechting)
Wat er gebeurt: De bacterie landt op de gastheercel. Een eiwit op de bacterie (het bacterial ligand) bindt heel specifiek aan een receptor op het celmembraan van de gastheer.
Belangrijk detail: In deze allereerste fase doet het actineskelet van de gastheercel nog helemaal niets (independent of actin skeleton). De binding zorgt er wel voor dat al die receptoren naar elkaar toe schuiven en gaan groeperen op specifieke, stevige plekken in het membraan (receptor clustering in lipid rafts). Zie het als een groepje mensen dat zich verzamelt rondom een attractie.
Phagocytic cup formation (Vorming van de 'beker')
Wat er gebeurt: Doordat de receptoren nu massaal geclusterd en geactiveerd zijn, sturen ze een alarmsignaal naar de binnenkant van de gastheercel.
Belangrijk detail: Dit signaal activeert de bestaande communicatieroutes van de cel die normaal het cytoskelet regelen. De cel wordt nu gedwongen om op die specifieke plek actine te polymeriseren (nieuwe actine-steunbalken bouwen). Hierdoor stulpt het membraan naar buiten toe uit (membrane extension) en vormt zich een kommetje of cup (phagocytic cup) dat de bacterie begint te omhelzen.
Phagocytic cup closure and retraction (Sluiting en intrekking)
Wat er gebeurt: De randen van de 'beker' groeien over de bacterie heen totdat ze elkaar raken en samensmelten. De rits is nu dicht: de bacterie zit opgesloten in een blaasje (een vacuole) binnen in de cel.
Belangrijk detail: Om dit blaasje nu daadwerkelijk dieper de cel in te trekken, moet de gemaakte actine-structuur weer worden afgebroken (actin de-polymerization). Het weghalen van de actine-steunbalken zorgt ervoor dat het blaasje met de bacterie erin soepel naar binnen zakt (retraction).
Kort samengevat: Binden (zonder actine) → Omhelzen (actine opbouwen) → Insluiten & Binnenhalen (actine weer afbreken).
Zipper mechanism → polymerisatie van actine na invasie van bacterie
FAK (Focal Adhesion Kinase) & Src kinase
Wat het is: FAK is een eiwit dat normaal gesproken aan de binnenkant van het celmembraan zit op plekken waar de cel vastgrijpt aan zijn omgeving (focal adhesions).
De actie: Wanneer een bacterie aan een receptor bindt (zoals bij het Zipper-mechanisme), activeert dit het eiwit Src kinase. Src kinase is een soort enzymatische schakelaar: het plakt een fosfaatgroep op FAK (FAK is phosphorylated). Hierdoor rekruteert en activeert FAK de volgende eiwitten in de keten.
Cdc42 / Rac (Rho-GTPases)
Wat het is: Dit zijn moleculaire 'aan/uit'-schakelaars binnen in de cel die specifiek de dynamiek van het cytoskelet aansturen.
De actie: Zodra FAK geactiveerd is, geeft deze het signaal door aan Cdc42 en Rac.
Cdc42 zorgt specifiek voor de vorming van filopodia (vingerachtige uitsteeksels).
Rac zorgt voor lamellipodia (brede, golvende membraanuitstulpingen).
Link met Trigger-mechanisme: Sommige bacteriën spuiten effectoren in die Cdc42 of Rac direct aanzetten, waardoor ze Src en FAK dus slim overslaan.
N-WASP (Neural Wiskott-Aldrich Syndrome Protein)
Wat het is: Dit is een zogenaamd 'effector-eiwit' dat reageert op de actieve schakelaars.
De actie: De geactiveerde Cdc42 bindt aan N-WASP. Hierdoor verandert N-WASP van vorm: het klapt open en verandert van een inactieve toestand naar een actieve toestand. Eenmaal opengevouwen kan N-WASP direct de 'bouwmachine' van de cel activeren.
ARP2/3 complex (Actin-Related Proteins)
Wat het is: Dit is de ultieme nucleating protein (de bouwmachine) die we in de allereerste dia zagen.
De actie: N-WASP activeert het ARP2/3-complex. Dit complex landt op een al bestaand actinefilament en begint daar een nieuwe zijtak te bouwen onder een specifieke hoek van 70 graden. Dit zorgt voor een explosieve groei van een vertakt actinenetwerk vlak onder het membraan, wat het membraan naar buiten duwt.
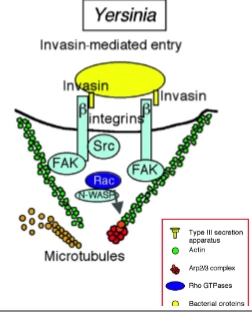
Voorbeelden Zipper-mechanism:
Yersinia
Yersinia
Stap 1: Binding aan de integrinereceptor
• Wat er gebeurt: Yersinia heeft een eiwit op zijn buitenkant dat invasine heet. Dit eiwit bindt super strak aan de $\beta$-keten van integrinereceptoren op het oppervlak van de gastheercel.
• De truc van de bacterie: Normaal gesproken gebruikt de cel deze integrines om zich stevig vast te zetten aan de buitenwereld, de extracellulaire matrix (E-matrix). Yersinia misbruikt deze 'ankerplaats' dus als landingsbaan.Stap 2: Het alarmsignaal en actinebouw
• Wat er gebeurt: Doordat invasine aan de integrines bindt, worden die receptoren massaal naar elkaar toe getrokken (clustering). Dit activeert een kettingreactie van enzymen die fosfaatgroepen plakken op specifieke aminozuren (Tyrosine-phosphorylation events).
• De moleculaire cascade: Dit activeert exact de domino-steentjes die we net zagen: FAK, Src en de Rho-GTPase Rac. Rac geeft het signaal door aan N-WASP, en die zet de bouwmachine (het ARP2/3-complex) aan het werk om massaal actine te polymeriseren (actin polymerization).Stap 3: Reorganisatie en opsluiting
• Wat er gebeurt: Door al die nieuwe actine-steunbalken verandert de vorm van de cel lokaal compleet (rearrangement cytoskeleton). Het celmembraan begint zich als een ritssluiting om de bacterie heen te vouwen (membrane traffic).
• De ontknoping: De randen van het membraan smelten samen: de 'beker' sluit zich (closure phagocytic cup). Yersinia zit nu veilig opgesloten in een blaasje binnen in de gastheercel.
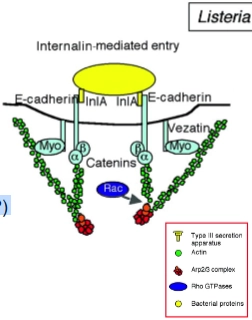
Voorbeelden Zipper-mechanism:
Listeria
Stap 1: Binding aan E-cadherine
• Wat er gebeurt: Listeria heeft een eiwit op zijn oppervlak genaamd Internalin A (InlA). Dit eiwit bindt specifiek aan E-cadherine op de gastheercel.
• De truc van de bacterie: E-cadherine is een receptor die epitheelcellen normaal gesproken gebruiken om heel strak aan elkaar te plakken (adherens junctions). Listeria doet alsof hij een buurcel is en grijpt zo de cel vast.Stap 2: Vorming van de 'beker' (en de missende schakels)
• Wat er gebeurt: Aan de binnenkant van de cel zit aan E-cadherine een heel complex van eiwitten vast: de catenines. Deze catenines vormen de fysieke brug tussen de receptor en het actineskelet.
• De antwoorden op de vraagtekens:
◦ Actin polymerization (regulated by?): Dit wordt gereguleerd door de Rho-GTPase Cdc42 (en het eiwit Arp2/3). Listeria dwingt via de catenines de cel om hier lokaal actine-steunbalken op te bouwen om het membraan rond de bacterie te duwen.
◦ Activation myosin motor (regulated by?): Dit wordt gereguleerd door Src kinase en receptorfosforylering. Myosine (het motor-eiwit dat we in de allereerste dia zagen) is hier nodig om spanning op het actinenetwerk te zetten, zodat het membraan strak om de bacterie heen trekt.Stap 3: Sluiting en intrekking
• Wat er gebeurt: Net als bij Yersinia sluit de 'beker' zich om de bacterie heen (closure). Het membraan smelt samen en Listeria wordt in een blaasje (vacuole) mee naar binnen getrokken (retraction), diep het cytoplasma in.
Listeria: Internalin B
→ de bacteriën hoeven hier zelf niet de cel in te ‘graven’
→ te zien door een extra mechanisme dat Listeria bevat:
= InlB en de Met-receptor
Listeria heeft nóg een troef: Naast Internalin A (InlA, voor E-cadherine) heeft Listeria ook een eiwit genaamd Internalin B (InlB) op zijn oppervlak.
De receptor: InlB bindt heel specifiek aan de Met-receptor op de gastheercel.
De normale functie: Normaal gesproken is Met de receptor voor Hepatocyte Growth Factor (HGF), een signaalstof die cellen aanzet tot groei, overleving en beweging. Als HGF bindt, activeert de cel signaalroutes (zoals PI3-kinase) die het actineskelet reorganiseren. Listeria doet zich met InlB dus gewoon voor als een groeifactor om diezelfde actine-reorganisatie te kapen.
Trigger mechanism: 4 successive stages
Fase 1: Pre-interaction stage (Vóór het contact)
• Wat er gebeurt: De bacterie zwemt richting de gastheercel. De bacteriële effector-eiwitten (de 'wapens') liggen al klaar in het bacteriële cytoplasma. De moleculaire injectienaald, het Type III Secretie Systeem (TTSS), zit al in elkaar in de celwand van de bacterie.
• Regulatie: De uitscheiding van deze eiwitten is streng onderdrukt totdat de bacterie exact op de juiste plek is. De bacterie activeert de transcriptie van deze TTSS-genen door omgevingsfactoren te scannen zoals de juiste pH en de hoeveelheid vrije zuurstof (zodat hij weet dat hij in de darmen zit). Pas wanneer er daadwerkelijk fysiek contact is met het membraan van de doelwitcel, gaat de sluis open: secretion!!
Fase 2: The interaction stage (De injectie)
• Wat er gebeurt: De bacterie landt op het membraan en vormt een zogenaamd signaling platform. De injectienaald (TTSS) doorboort het eukaryote celmembraan. Dit doet de bacterie door specifieke eiwitten, zogenaamde translocons, in het membraan van de gastheercel te duwen. Dit translocon vormt de poort waar de rest van de effectoren doorheen kan stromen.
• Het verschil:
◦ Salmonella gebruikt de translocon-eiwitten SipB en SipC.
◦ Shigella gebruikt de translocon-eiwitten IpaB en IpaC.
Fase 3: Formation of a macropinocytic pocket (De actine-explosie)
Dit is de meest dynamische fase. De geïnjecteerde eiwitten kapen de Rho-GTPases (Cdc42/Rac) die we eerder bespraken om een enorme golf in het membraan (ruffling) op te wekken. Hierin verschillen de twee bacteriën strategisch:Salmonella-strategie:
• SopB en SopE: Dit zijn de effectoren die direct de gastheer-schakelaars Cdc42 en Rac aanzetten. Hierdoor start een explosieve polymerisatie van actine via het Arp2/3-complex.
• SipA: Dit eiwit bindt direct aan de nieuw gevormde actinefilamenten en stabiliseert ze, zodat de cel de steunbalken niet direct weer kan afbreken. De uitstulping blijft zo stevig overeind staan om de bacterie te omsluiten.Shigella-strategie:
• VirA en IpaC: VirA grijpt in op de microtubuli (de grotere transportkabels). Samen met IpaC activeren ze Cdc42/Rac en het enzym Src om actinegroei te forceren.
• IpgD: Dit is een heel slim enzym. Het breekt een membraan-lipide af. Hierdoor laat het actineskelet los van het membraan direct boven zich (disconnecting from subcortical membrane). Omdat de 'ankerplaatsen' weg zijn, wordt het celmembraan super flexibel en slap, waardoor er nóg grotere golven en flappen ontstaan om de bacterie heen.
Fase 4: Actin depolymerization (Het sluiten van de zak)
• Wat er gebeurt: De bacterie is nu overspoeld door het membraan en zit binnen in een grote blaas (een macropinosoom). Net als bij het Zipper-mechanisme moet de cel nu de 'chaos' opruimen en de gemaakte actine-steunbalken weer snel afbreken (depolymerisatie) om de rust te herstellen en de bacterie dieper de cel in te trekken.
• Het verschil:
◦ Salmonella gebruikt SptP: Dit eiwit werkt als een antagonist (tegenhanger) van SopE. Het zet de schakelaars Cdc42 en Rac weer netjes 'UIT'. De actinebouw stopt direct en de cel keert terug naar zijn normale vorm.
◦ Shigella gebruikt IpaA: Dit eiwit bindt aan vinculine (een belangrijk eiwit dat actine aan het membraan koppelt) en remt dit af. Hierdoor stort het lokale actine-netwerk direct in elkaar.
Samenvatting Zipper- en Trigger-mechanism
Na het Zipper- of Trigger-mechanism → 2 survival strategieën van de bacterie
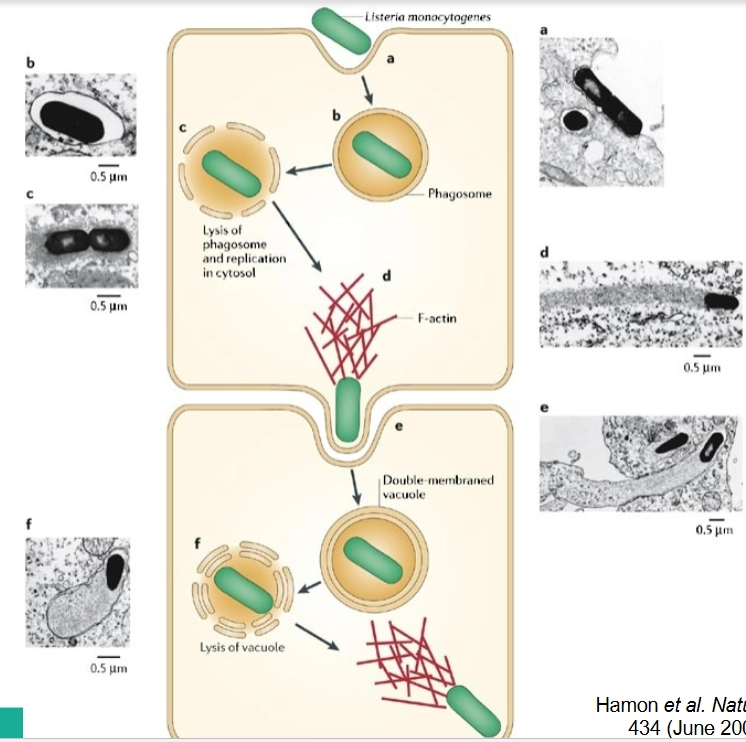
Cytosolic Pathogens = Sommige bacteriën weigeren in het blaasje te blijven. Ze breken het membraan van de vacuole heel snel af en ontsnappen naar het cytoplasma (de celvloeistof).
Voordeel: In het cytoplasma is een overvloed aan voedingsstoffen (zoals suikers en aminozuren) te vinden voor snelle vermenigvuldiging. Bovendien kunnen ze hier het actineskelet kapen (zoals we eerder zagen) om zich voort te bewegen.
Voorbeelden: Listeria monocytogenes en Shigella flexneri.
Vacuolar Pathogens = Andere bacteriën kiezen ervoor om juist binnenin de vacuole te blijven wonen. Normaal gesproken fuseert zo'n vacuole met een lysosoom (een blaasje vol maagzuur en afbraakenzymen) om de bacterie te doden. Deze bacteriën manipuleren de celcommunicatie om die fusie te blokkeren of te veranderen. Ze bouwen de vacuole om tot een veilige, op maat gemaakte 'bunker' waarin ze kunnen groeien.
Voorbeeld: Salmonella enterica (die een Salmonella-containing vacuole of SCV vormt).
Cytosolic pathogens: moleculaire stappen
Stap 1: Lysis of the vacuole (Het blaasje opblazen)
De bacterie zit vast in de vacuole. Om hieruit te breken, scheiden ze specifieke eiwitten (toxines) uit die gaten boren in het eukaryote membraan:
Listeria gebruikt LLO en PlcA/PlcB: LLO (Listeriolysin O) vormt poriën in het membraan. PlcA en PlcB zijn fosfolipases (enzymen die vetten in het membraan afbreken). Samen lossen ze het blaasje op.
Shigella gebruikt IpaB en IpaC: Deze eiwitten (die ook al hielpen bij de invasie) vormen poriën in het vacuole-membraan, waardoor dit destabiliseert en uit elkaar valt.
Stap 2: Intracellular replication (Vermenigvuldiging)
Zodra ze vrij in het cytoplasma zwemmen, beginnen Listeria en Shigella zich razendsnel te delen. Ze maken optimaal gebruik van de aanwezige nutriënten van de gastheer.
Stap 3: Actin-based motility (De actineraket)
Dit is het meest spectaculaire onderdeel, waarbij de bacteriën de Actin Binding Proteins misbruiken die we in je allereerste vraag bespraken:
De bacteriën produceren een specifiek eiwit aan één kant van hun pool (hun 'achterkant'):
Listeria gebruikt ActA.
Shigella gebruikt IcsA (ook wel VirG genoemd).
Deze bacteriële eiwitten rekruteren en activeren direct het Arp2/3-complex (de actine-bouwmachine) van de gastheercel.
Er vindt een explosieve polymerisatie van actine plaats aan de achterkant van de bacterie. Dit vormt een groeiende staart van vertakt actine (een actin comet tail). De continue groei van deze staart duwt de bacterie als een raket met hoge snelheid door het cytoplasma heen.
Stap 4: Intercellular spread (Infectie van de buurcel)
De bacterie schiet met zo'n vaart door de cel dat hij keihard tegen het buitenmembraan van de gastheercel botst. Het membraan stulpt hierdoor heel ver naar buiten uit en vormt een lange uitstulping (protrusion) die diep in de buurcel dringt.
De buurcel denkt dat het een normaal blaasje is en slikt deze uitstulping in. De bacterie zit nu in een dubbel-membraan vacuole in de buurcel. De bacterie gebruikt vervolgens weer zijn LLO/fosfolipases om ook dit dubbele membraan kapot te maken.
Het grote biologische voordeel: De bacterie kan zich nu van cel naar cel verspreiden zonder dat hij ooit de cel hoeft te verlaten. Hierdoor blijft de bacterie volledig onzichtbaar voor de antistoffen en immuuncellen die buiten de cellen op de loer liggen!
Vacuole → maken van een hostile environment door de gastheer
= door de volgende barrières
Poor nutrient content: Er is bijna geen eten (suikers/aminozuren) binnenin het blaasje.
Low pH: De cel pompt protonen naar binnen om het blaasje extreem zuur te maken.
Antibacterial peptides & Oxygen radicals: Er worden giftige eiwitten en agressieve zuurstofradicalen (ROS) in gegooid die de bacterie proberen te oxideren en kapot te bijten.
→ om deze omgeving te overleven moeten de bacteriën kiezen uit de volgende strategieën:
Adaptation to the hostile conditions: Gewoon incasseren. Sommige bacteriën hebben een aangepaste, supersterke celwand die simpelweg bestand is tegen het zuur en de zuurstofradicalen.
Alteration of the hostile vacuole: De boel saboteren. De bacterie produceert enzymen zoals peroxidase en catalase die de giftige zuurstofradicalen direct neutraliseren. Daarnaast blokkeert hij actief de fusie met het lysosoom.
Escape to the cytosol: Het blaasje opblazen en maken dat je wegkomt (de tactiek van Listeria en Shigella).
→ bij keuze 3 heeft de bacterie evolutionaire voordelen:
Space to grow (replicate!): In het cytoplasma is alle ruimte om ongestoord te delen.
Access to nutrients: De celvloeistof zit bomvol met suikers, ATP en aminozuren. Het is één groot buffet voor de bacterie.
→ sommige bacteriën bewegen hierbij door de cel met behulp van actine
Samenvatting van de verschillende pathogenen besproken:
Yersinia
Listeria
Shigella
Salmonella
Pathogeen | Voorkeurslocatie in weefsel | Effect op Macrofagen | Gevolg voor gastheer |
|---|---|---|---|
Yersinia | Extracellulair (buiten de cel) | Verlamt ze (Yops), voorkomt dat hij opgegeten wordt. | Lokale infectie van lymfeklieren. |
Listeria | Intracellulair (in de cel) | Overleeft en groeit ín de macrofaag. | Verspreiding via immuuncellen naar bloed/hersenen. |
Shigella | Intracellulair (in het epitheel) | Doodt macrofagen, lokt PMN's uit die de darmwand slopen. | Ernstige, lokale darmwandbeschadiging (dysenterie). |
Salmonella | Intracellulair (in vacuole) | Schakelt over op SPI2 om te groeien ín de macrofaag. | Kan systemisch worden (verspreiding door hele lichaam). |
(ook nog een afbeelding van Yersinia & Listeria)