Biology: Transport in animals
1/39
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced |
|---|
No study sessions yet.
40 Terms
8.1 Introduction to the circulatory system
8.1 Introduction to the circulatory system
Why do multicellular organisms need transport system
Multicellular organisms need internal transport systems like the circulatory system.
This is because, compared to single-celled organisms:
Multicellular organisms are larger, so the diffusion distance across their surface would be too large.
Multicellular organisms have a higher metabolic rate.
Multicellular organisms need to supply nutrients and oxygen rapidly to a larger number of active cells.
So overall diffusion alone would be too slow
Single and double circulatory systems
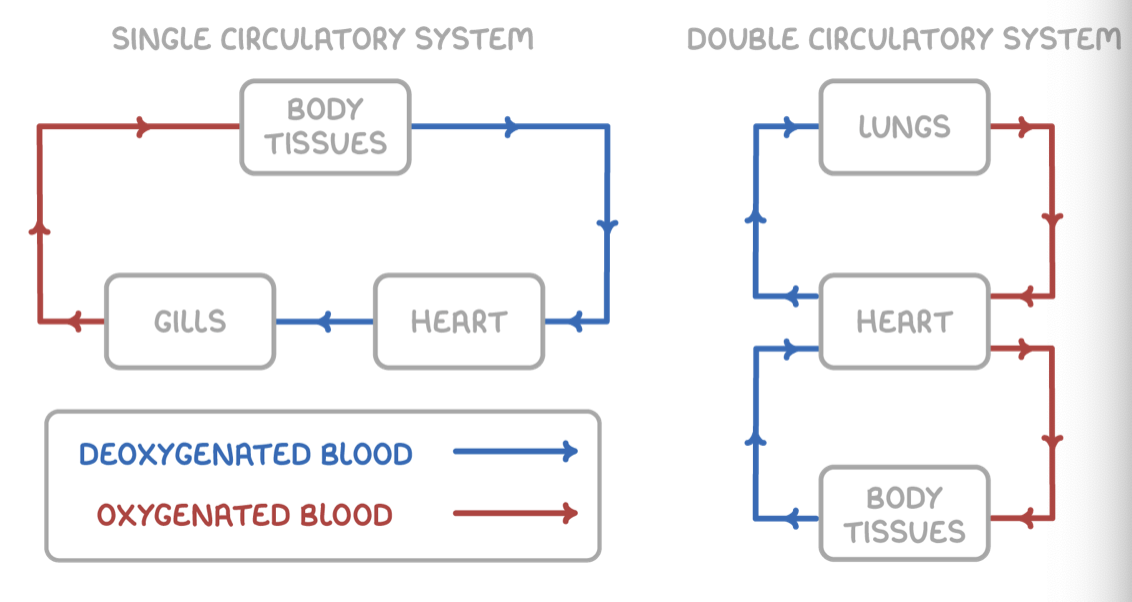
Fish circulatory system
This means that blood passes through the heart only once per circuit.
The single circulation in fish occurs as follows:
The heart pumps blood to the gills to pick up oxygen.
Blood flows directly from the gills to the rest of body to deliver oxygen.
Mammal circulatory system
This means that blood passes through the heart twice per circuit.
The double circulation in mammals occurs as follows:
The right side of the heart pumps deoxygenated blood to the lungs to pick up oxygen.
The left side of the heart pumps oxygenated blood to the rest of the body, which gives an extra push to deliver this blood more quickly to distant body tissues or those that require a high pressure, like the kidneys.
Open and closed circulatory systems
Vertebrates have closed circulatory systems:
Their blood is enclosed in vessels.
Arteries generally distribute oxygenated blood.
Veins generally return deoxygenated blood to heart.
Some invertebrates like insects have open systems:
Their blood flows freely through the body cavity.
Blood returns to heart through valves.
Blood doesn't just transport oxygen.
Mammalian circulatory system structure
Mammals have a closed, double circulatory system. The two main divisions of the mammalian circulatory system are the systemic system and the pulmonary system.
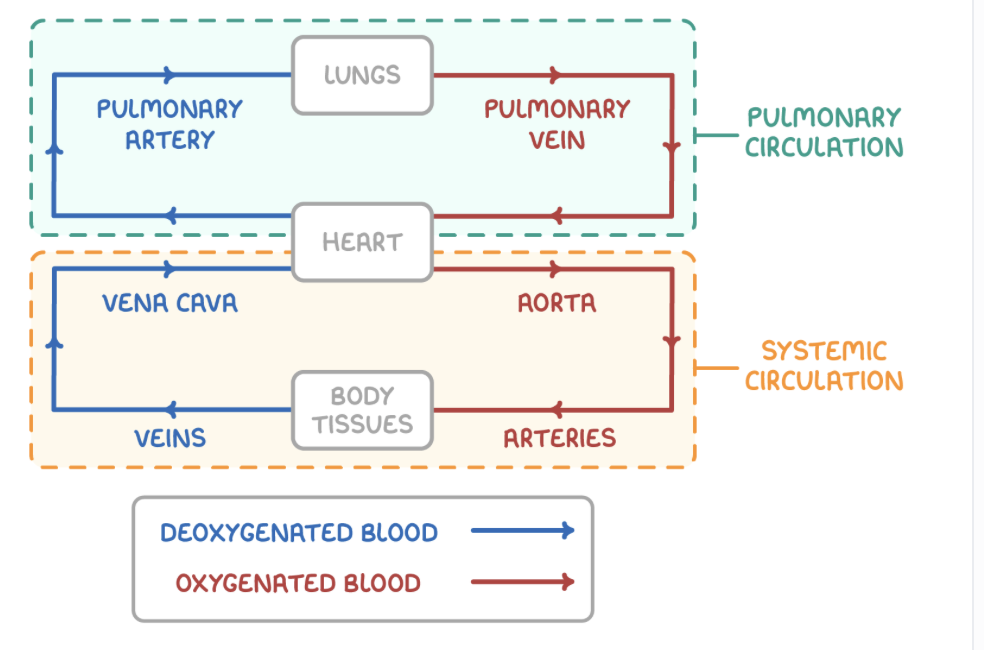
Systemic and Pulmonary circulatory system
Systemic circulatory system:
Oxygenated blood is pumped out of the heart via the aorta to most body tissues.
Deoxygenated blood is returned to the heart via the vena cava from the body tissues.
Pulmonary circulatory system:
Deoxygenated blood is pumped out of the heart via the pulmonary artery to the lungs.
Oxygenated blood is returned to the heart via the pulmonary vein from the lungs.
8.2 Blood vessels
8.2 Blood vessels
Blood vessels and their functions
There are five main types of blood vessels in the mammalian circulatory system.
These main types of blood vessels are:
Arteries - Carry blood away from the heart.
Arterioles - Carry blood from arteries into capillaries.
Capillaries - Site of diffusion between blood and body tissues.
Venules - Carry blood from capillaries into veins.
Veins - Return blood to the heart.
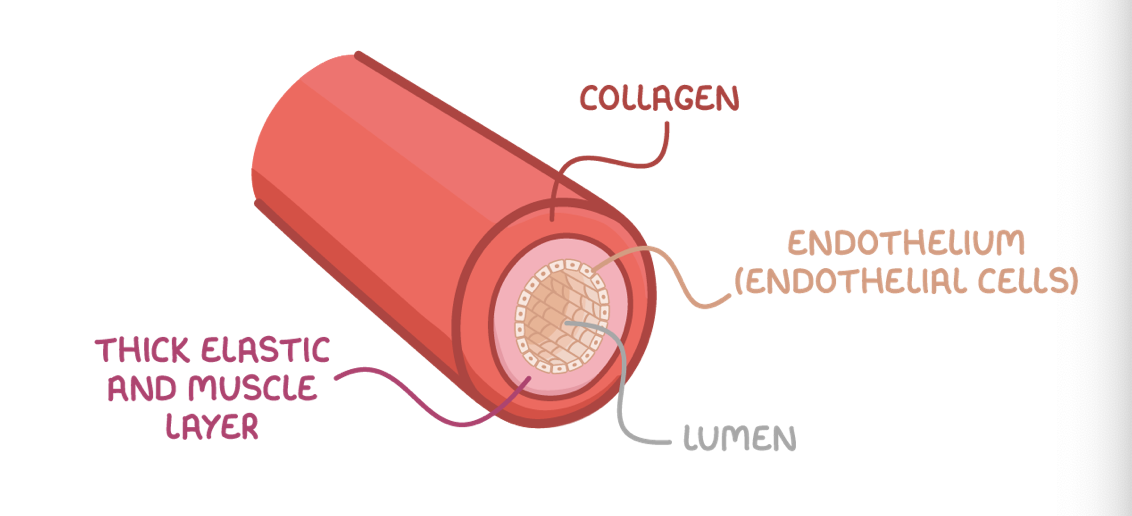
Arteries and arterioles
Adaptations of arteries:
Collagen - Provides strength to prevent the vessel from bursting and to maintain vessel shape.
Elastic fibres - Contain elastin that lets them stretch and recoil to minimise changes in pressure.
Thick smooth muscle layer - Contracts/relaxes to constrict/dilate the lumen and control blood flow.
Arterioles are smaller than arteries, but with a comparatively larger lumen. Their walls have more smooth muscle and less elastin as they do not need to withstand such high pressures.
Vasoconstriction and vasodilation
Vasoconstriction and vasodilation
The smooth muscle in arteries and arterioles means they are adapted to control the blood flow through them.
They do this through the following mechanisms:
Vasoconstriction - Smooth muscle contracts, constricting the blood vessel and decreasing blood flow.
Vasodilation - Smooth muscle relaxes, dilating the blood vessel and increasing blood flow.
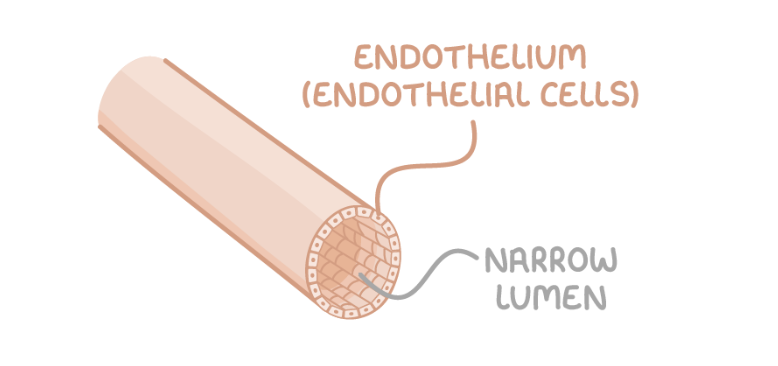
Capillaries
Capillaries form extensive networks between arterioles and venules, providing an area between blood and tissues where exchange of substances like gases and nutrients can occur.
Adaptations of capillaries:
Lumen is very narrow - This allows red blood cells to be close to body cells.
Walls are thin - Substances can be exchanged across a short distance by diffusion.
Highly branched - This provides a large surface area for diffusion.
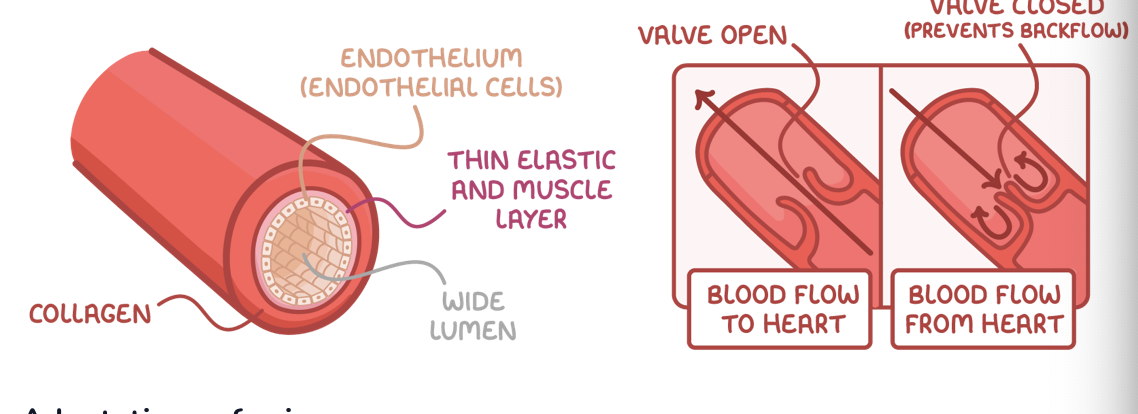
Veins and venues
Veins are adapted to carry blood towards the heart at low pressure.
Adaptations of veins:
Collagen - Provides strength to prevent the vessel from bursting and maintain vessel shape.
Little smooth muscle and elastic fibre - Not much is needed due to low blood pressure, and thinner walls allow veins to be easily compressed, aiding the flow of blood.
Valves - Pocket valves shut to prevent the backflow of blood when veins are squeezed by surrounding skeletal muscle.
The pocket valves in veins are similar in structure to valves in the heart, but pocket valves are the only type of valve controlled by skeletal muscle.
Venules are smaller than veins. They have very thin walls, and very little smooth muscle. Both venules and veins have valves.
8.3 Blood, Tissue Fluid and Lymph
8.3 Blood, Tissue Fluid and Lymph
Blood composition
Plasma - Mostly water, transports substances in solution.
Red blood cells - Carry oxygen.
White blood cells - Immune cells.
Platelets - Involved in clotting
Function of blood
Transports oxygen and carbon dioxide.
Transports nutrients from digestion.
Transports waste for excretion.
Transports hormones.
Transports food from storage.
Transports clotting factors.
Composition of tissue fluid
Tissue fluid (interstitial fluid) fills spaces between cells (interstitial space). It is the site of diffusion between blood and body cells, providing cells with nutrients and oxygen while removing waste products. It also helps fight infection as it forms part of the immune response.
Tissue fluid has the same composition as plasma, except:
Tissue fluid has no red blood cells.
Tissue fluid has fewer proteins.
Tissue fluid has fewer white blood cells.
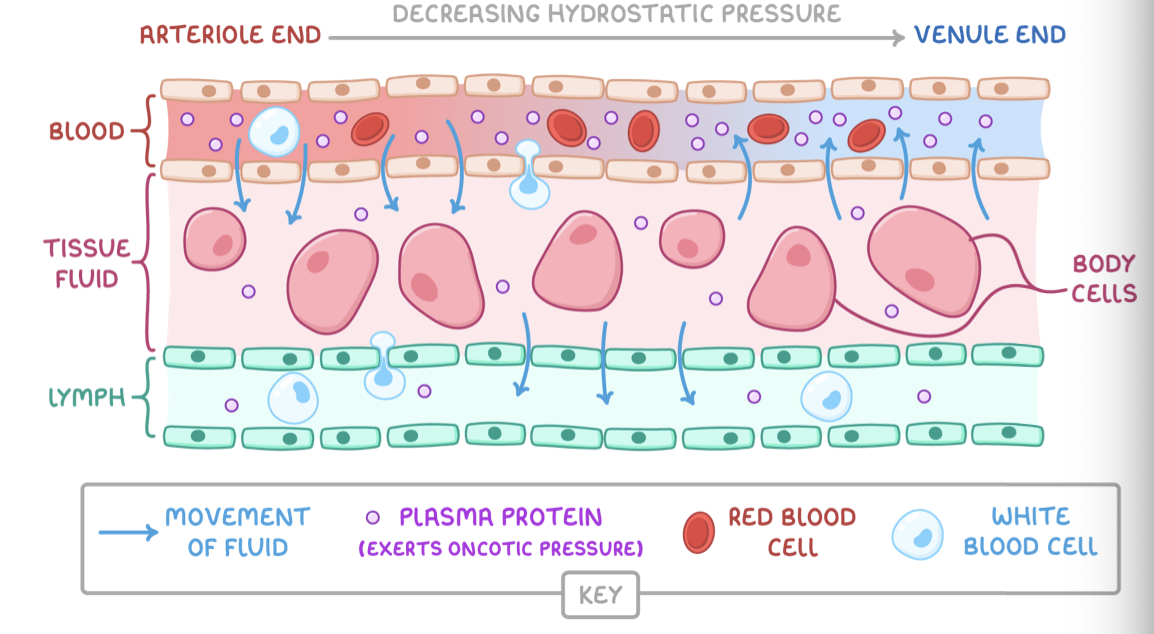
The formation of tissue fluid
Tissue fluid is formed from the blood flowing through capillaries.
At the arteriole end of capillaries:
A high hydrostatic pressure, exerted by the force of the heart pumping, forces fluid out of capillaries.
This forms tissue fluid surrounding body cells.
At the venule end of capillaries:
The hydrostatic pressure is lower.
Proteins in blood exert a high oncotic pressure, a type of osmotic pressure, in capillaries.
The water potential is lower in capillaries than in tissue fluid due to fluid loss.
Some tissue fluid moves back into capillaries by osmosis.
The composition of lymph
Lymph is the fluid that flows around the lymphatic system via lymph vessels.
Lymph has a similar composition to tissue fluid, except:
Lymph has less oxygen and nutrients.
Lymph has more fatty acids.
Lymph has more white blood cells (lymphocytes).
The formation and transport of lymph
Lymph is formed from tissue fluid.
It is formed and transported around the body as follows:
Some tissue fluid doesn't re-enter capillaries from tissue fluid.
This fluid instead drains into lymph vessels (lymph capillaries) forming lymph.
Lymph is transported through lymph vessels by muscle contractions.
Lymph is passed through lymph nodes to filter pathogens.
Lymph is eventually returned to the blood.
8.4 Haemoglobin
8.4 Haemoglobin
How haemoglobin transports oxygen
Oxygen enters the lungs from atmospheric air and then needs to be transported to body cells.
Oxygen is transported to body cells via the blood:
Red blood cells contain haemoglobin, which has 4 haem groups.
In the capillaries in the lungs, oxygen binds to iron in haem groups forming oxyhaemoglobin.
Each haemoglobin molecule can carry 4 oxygen molecules, one per haem group.
Oxyhaemoglobin can be transported via blood to respiring body tissues.
At body cells, oxygen dissociates from haemoglobin.
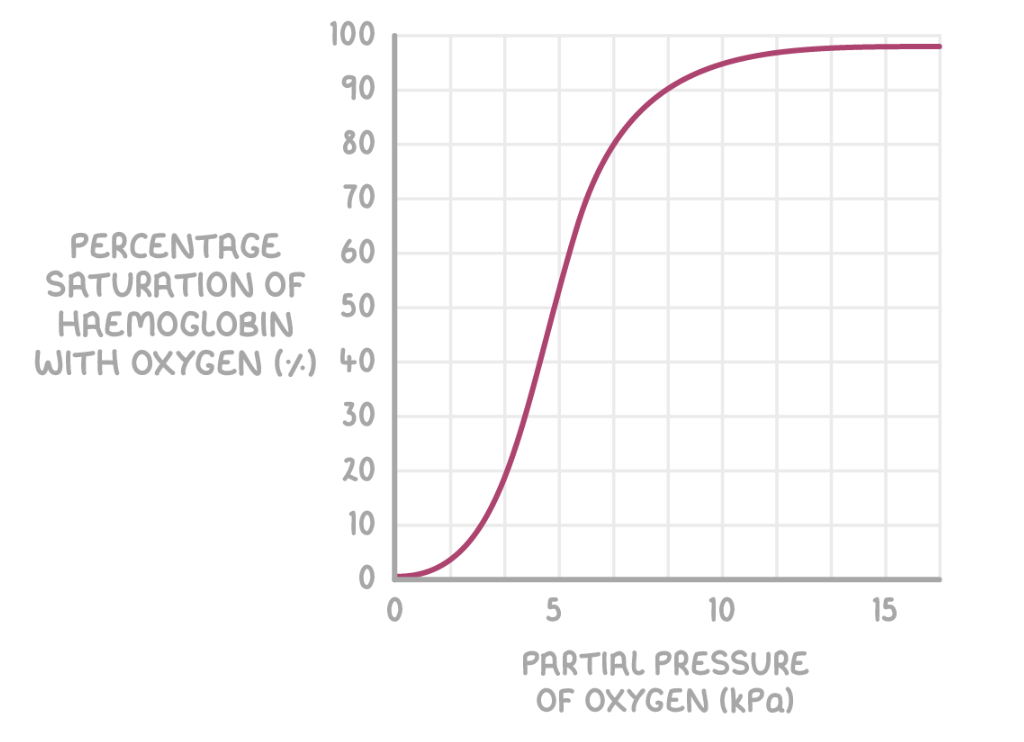
Factors affecting haemoglobin
Haemoglobin saturation with oxygen depends on the oxygen concentration, or partial pressure of oxygen (pO2). The units of partial pressure are kilopascals (kPa). Partial pressure can be shown on a graph called an oxygen dissociation curve.
pO2 affects haemoglobin saturation as follows:
Higher pO2 - Haemoglobin has a high affinity for oxygen and binds with it (e.g. in the lungs).
Lower pO2 - Haemoglobin has a low affinity for oxygen and releases it (e.g. at respiring body cells).
Cooperative nature of oxygen binding
Haemoglobin saturation with oxygen also changes depending on how much oxygen has bound to it.
The cooperative nature of oxygen binding means that:
When haemoglobin binds with one oxygen, it changes shape so it becomes easier to bind another oxygen.
When haemoglobin is mostly saturated with oxygen, it is harder for more oxygen to bind.
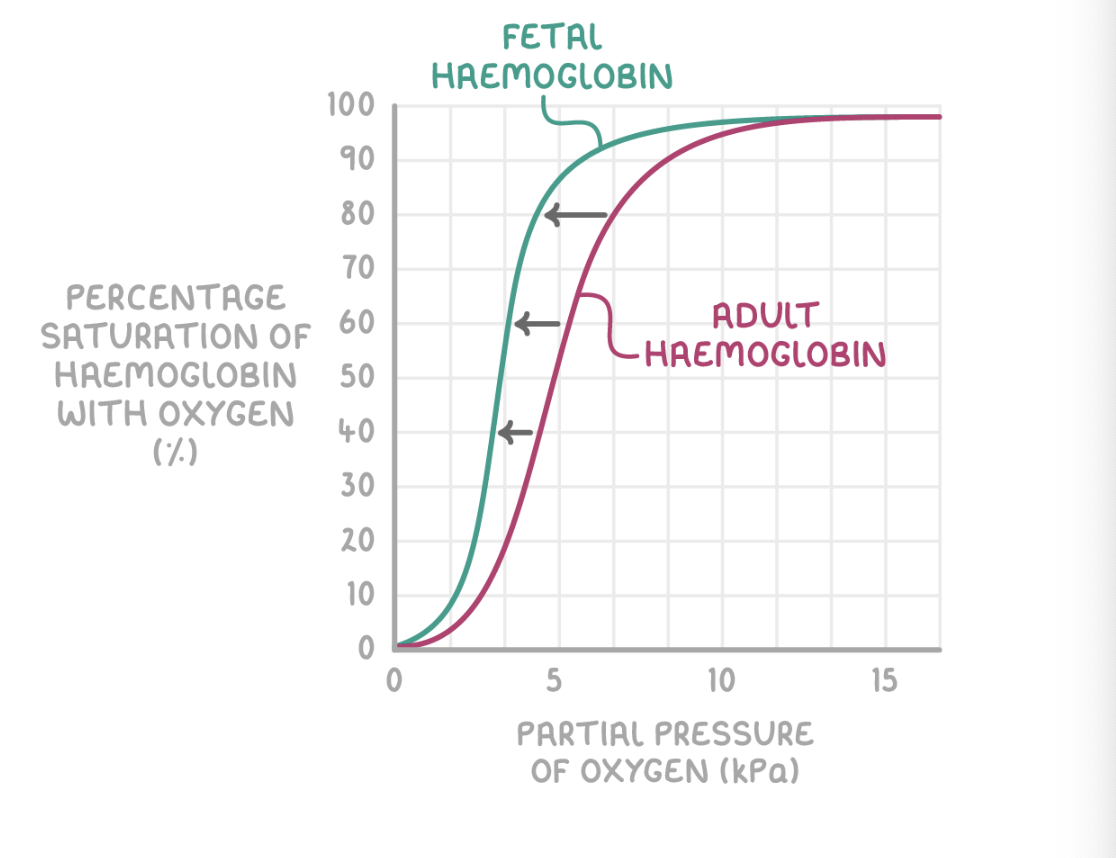
Differences between fetal and adult haemoglobin
Fetal haemoglobin differs from adult haemoglobin because:
The fetus needs to obtain oxygen from the mother's blood.
The fetal haemoglobin therefore has a higher oxygen affinity than the adult haemoglobin found in the mother's blood.
This allows the oxygen to dissociate from the mother's haemoglobin, and bind with haemoglobin in the fetal blood.
This ensures that the fetus gets enough oxygen to survive while it develops.
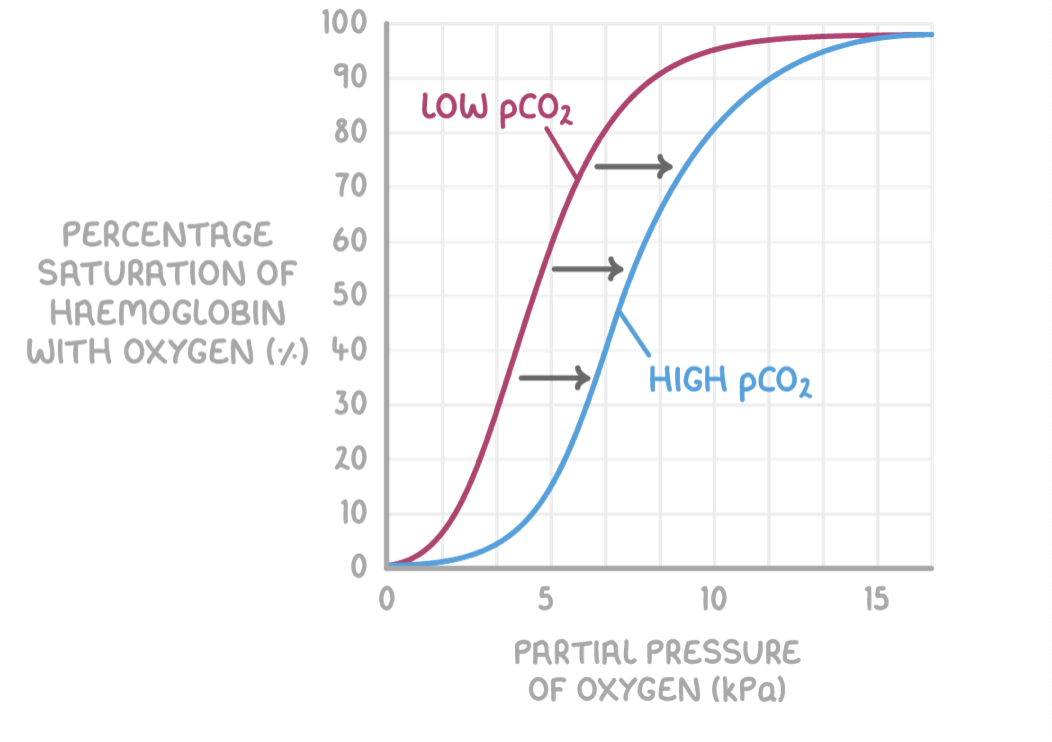
The Bohr effect
Haemoglobin also has a lower affinity for oxygen at higher partial pressures of carbon dioxide (pCO2)
pCO2 affects haemoglobin saturation as follows:
Higher pCO2 at respiring tissues causes haemoglobin to release oxygen.
This is because of the Bohr effect, which is the decreased affinity for oxygen in haemoglobin when carbon dioxide is present.
This means the oxygen saturation of haemoglobin is lower for a given pO2when pCO2 is higher.
This is needed because the active tissues that produce carbon dioxide require a lot of oxygen from blood. The Bohr effect shifts the oxygen dissociation curve to the right, as seen by the blue line in the graph above.
Transport of carbon dioxide in the bloodstream
It is beneficial for CO2 to be converted into another form in the blood. This is because this maintains a steep diffusion gradient between respiring tissues, which need to get rid of CO2, and the blood, which removes this CO2 to the lungs.
There are multiple ways CO2 is transported via the blood:
A small percentage of CO2 is transported dissolved in plasma.
Some CO2 enters red blood cells, and is transported bound to haemoglobin as carbaminohaemoglobin.
Most CO2 enters red blood cells and undergoes a reaction in the cytoplasm to form hydrogen carbonate ions.
Transport of Co2 as hydrogen carbonate ions
Once CO2 from respiration enters red blood cells, it undergoes some changes in the cytoplasm so it can be transported as hydrogen carbonate ions (also called bicarbonate ions, or HCO3-). This process also explains why the Bohr effect occurs.
At respiring tissues, which have a high pCO2, the following occurs:
CO2 reacts with water to form carbonic acid (H2CO3), catalysed by the enzyme carbonic anhydrase.
H2CO3 dissociates to H+ and HCO3-.
H+ binds to haemoglobin forming haemoglobinic acid, causing haemoglobin to release oxygen.
This prevents the blood from becoming too acidic (H+ ions reduce pH).
It also causes the Bohr effect: at high pCO2, haemoglobin releases oxygen so it can diffuse into respiring tissues.
HCO3- ions leave red blood cells and are transported via plasma, while chloride ions (Cl-) enter red blood cells.
This is called the chloride shift, and it maintains the charge balance by preventing excessive positive charge in red blood cells.
At the lungs, which have a low pCO2, the following occurs:
The low pCO2 causes HCO3- and H+ to reform CO2.
CO2 diffuses out of the body during expiration.
8.5 Heart structure
8.5 Heart structure
Heart chambers
The human heart is made up of four chambers: the left and right atria, and the left and right ventricles.
Differences between atria and ventricles:
Atria - The top chambers in the heart that collect blood from blood vessels (veins).
Ventricles - The bottom chambers in the heart that pump blood into blood vessels (arteries).
The left side of the heart contains oxygenated blood, and the right side contains deoxygenated blood. This forms two separate pumping mechanisms.
The heart has two separate pumping mechanisms because:
Blood pressure drops in the lungs as it flows through the capillaries.
A single pump would slow the blood flow to the body cells.
Two pumps increase the pressure before the blood circulates.
The septum separates the two sides of the heart, preventing oxygenated and deoxygenated blood from mixing.
Atrioventricular valves
The tricuspid valve is located between right atrium and right ventricle.
The bicuspid (mitral) valve is located between left atrium and left ventricle.
Both valves prevent backflow of blood into the atria when the ventricles contract.
Semi-lunar valves
These are located between the ventricles and the pulmonary artery and the aorta.
They prevent the backflow of blood into the ventricles when they relax.
Blood vessels associated with the heart
These blood vessels are:
The pulmonary vein - This moves oxygenated blood into the left atrium from the lungs.
The aorta - This moves oxygenated blood from the left ventricle to the body.
The vena cava - This moves deoxygenated blood into the right atrium from the body.
The pulmonary artery - This moves deoxygenated blood from the right ventricle to the lungs.
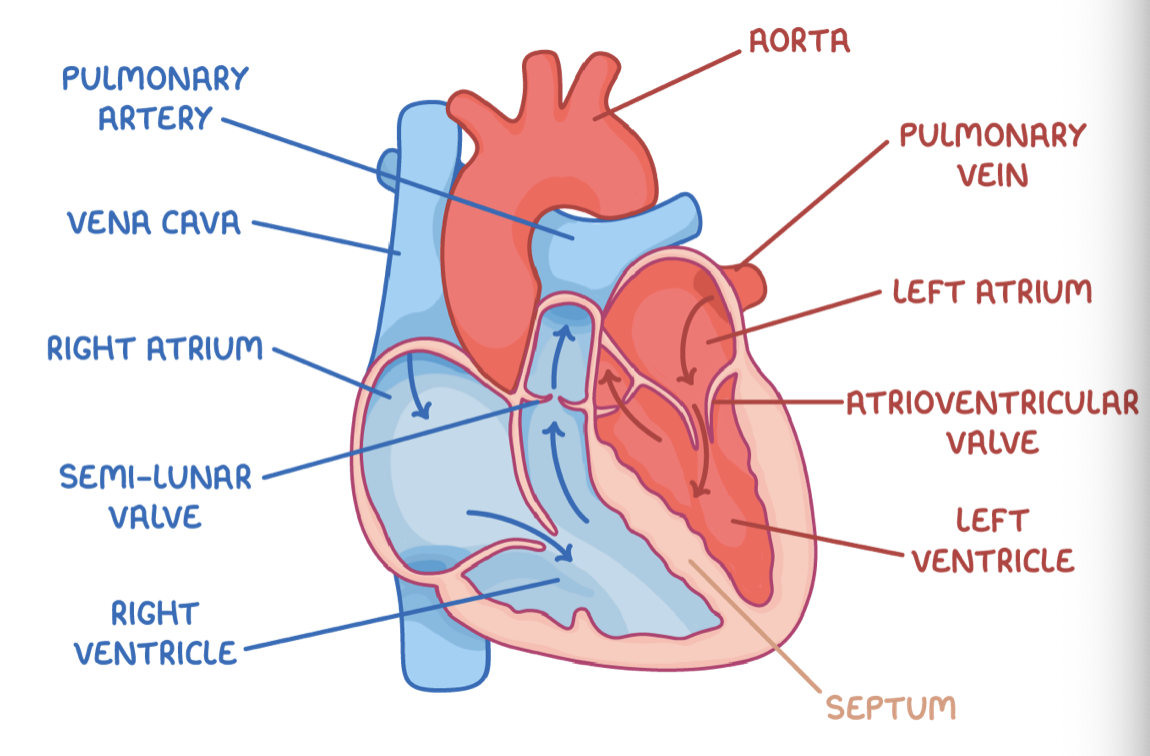
Heart structure relation to function
The ventricle walls are thicker than the atria walls
The ventricles have thicker walls with more muscle than the atria.
This is because:
The atria only need enough pressure to pump blood a short distance into the ventricles.
The ventricles need a lot of pressure to pump blood a long distance out of the heart to other organs.
The left ventricle wall is thicker than the right ventricle wall
The left ventricle has a thicker wall with more muscle than the wall of the right ventricle.
This is because:
The right ventricle only needs enough pressure to pump deoxygenated blood a short distance to the lungs.
The left ventricle needs a lot of pressure to pump oxygenated blood to the other more distant organs of the body.
8.6 Cardiac cycle
8.6 Cardiac cycle
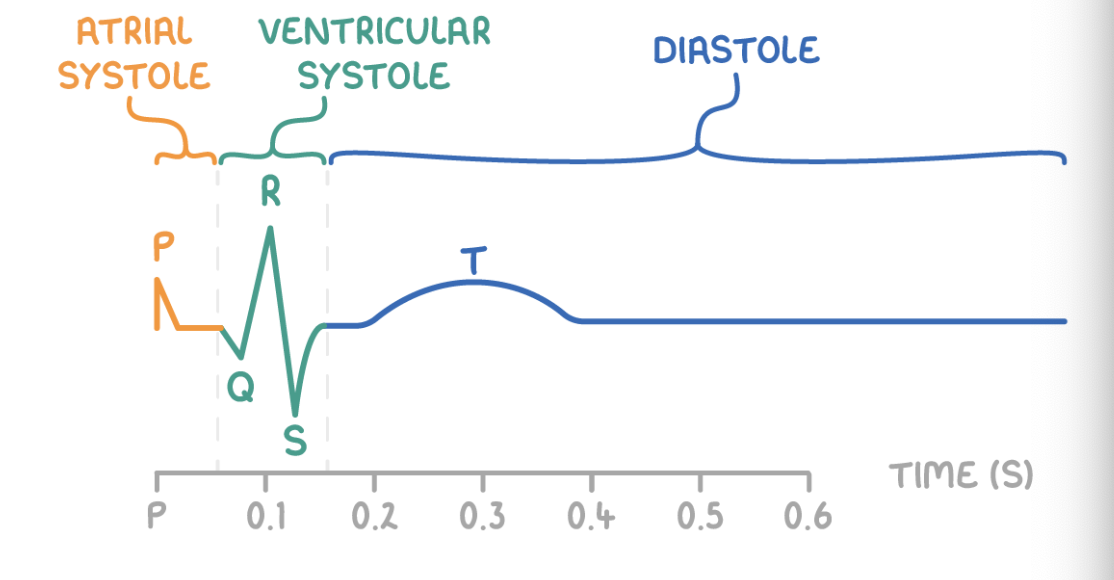
Stages of the cardiac cycle
Stage 1 - Atrial systole
The ventricles relax, and the atria contract.
This increases the atrial pressure.
The atrioventricular valves open.
Blood flows into the ventricles.
Stage 2 - Ventricular systole
The ventricles contract, and the atria relax.
The ventricular pressure increases.
The semi-lunar valves open, and the atrioventricular valves close.
Blood flows into the arteries.
Stage 3 - Diastole
The ventricles and atria relax.
The semi-lunar valves close.
Blood flows passively into the atria.
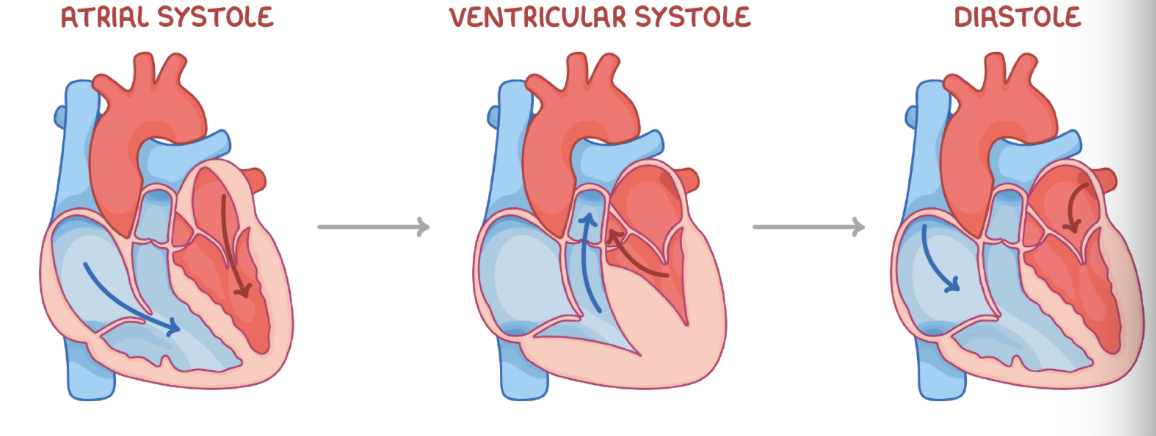
8.7 How the heartbeat is controlled
8.7 How the heartbeat is controlled
How the heartbeat is controlled
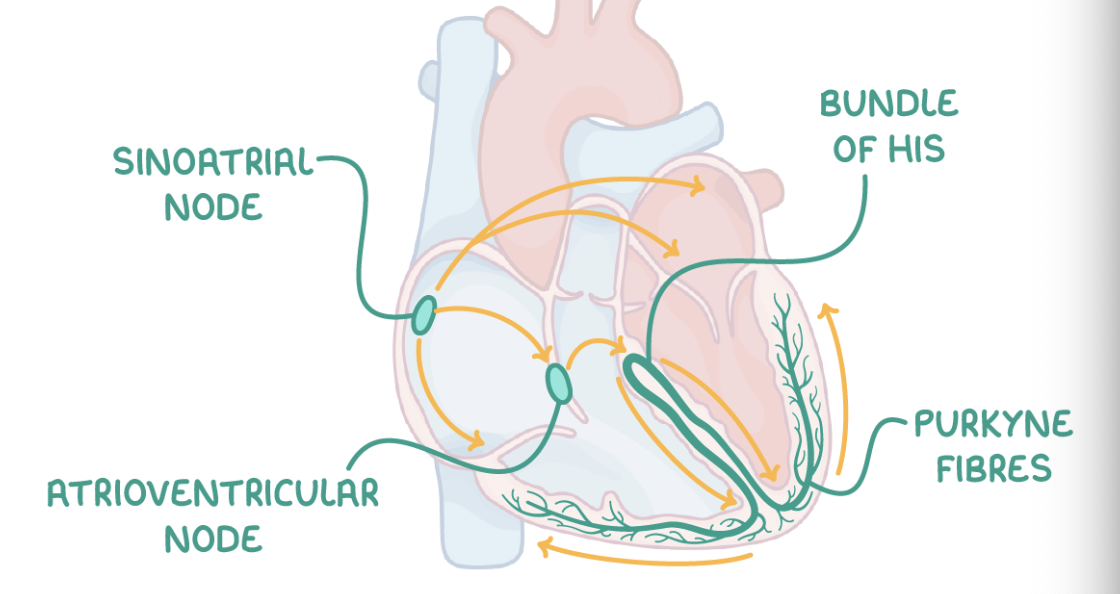
Cardiac muscle is myogenic, which means that the contraction of cardiac muscle initiates within the heart itself. The basic rhythm of the heart is maintained by a wave of electrical excitation.
This wave travels through the following structures in this order:
Sinoatrial node (SAN) - Initiates the heart beat by stimulating the atria to contract.
A layer of collagen fibres - Prevents direct electrical flow from atria to ventricles.
Atrioventricular node (AVN) - Picks up the electrical activity from the SAN and imposes a slight delay.
Bundle of His - Receives electrical activity from the AVN and conducts the wave of excitation to the apex (base) of the heart.
Purkyne fibres - These branch off the bundle of His, causing the right and left ventricles to contract from the bottom upwards.
How electrocardiograms monitor heart function
Electrocardiograms (ECGs) record the heart's electrical activity using electrodes.
An ECG shows the various stages of the cardiac cycle:
P wave - Atrial systole
QRS - Ventricular systole
T wave - Diastole
ECGs help diagnose abnormalities like:
Tachycardia - This is an abnormally rapid heart rate.
Bradycardia - This is an abnormally slow heart rate.
Ectopic heartbeats - This is extra heartbeats out of the normal rhythm.
Atrial fibrillation - This is the abnormally rapid and ineffective contraction of the atria.