Lecture 17- Ionotropic Receptors
1/26
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced | Call with Kai |
|---|
No study sessions yet.
27 Terms
Ionotropic receptors vs metabotropic- MEMORIZE PIC
ionotropic: NT binds, ion channel opens, done
happens on a ms timescale
effects are limited by the type of ion channel it is
they have low affinity for nt binding molecules because they only want to produce EPSPs in response to big bolus’s of molecules
Metabotropic- g protein couples- involves a number of different reactions
is not itself an ion channel
communicator that links NT eC to g protein IC
look at pic
activation of pathways take a lot longer so effecrs take longer to happen
termination of effects is slower too bc g protein can still be active after NT unbinds
located some distance from active zone
they already dont see large amounts of NT, so they will only be activated when acetylcholine leaves the cleft
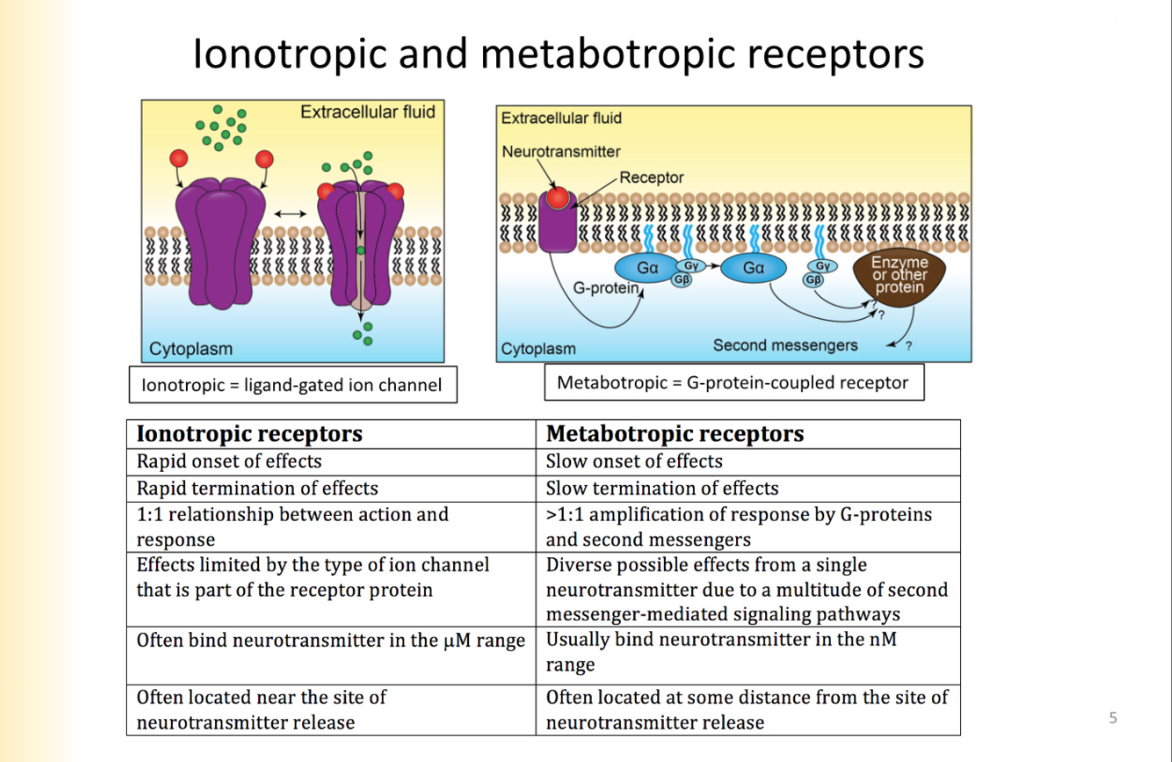
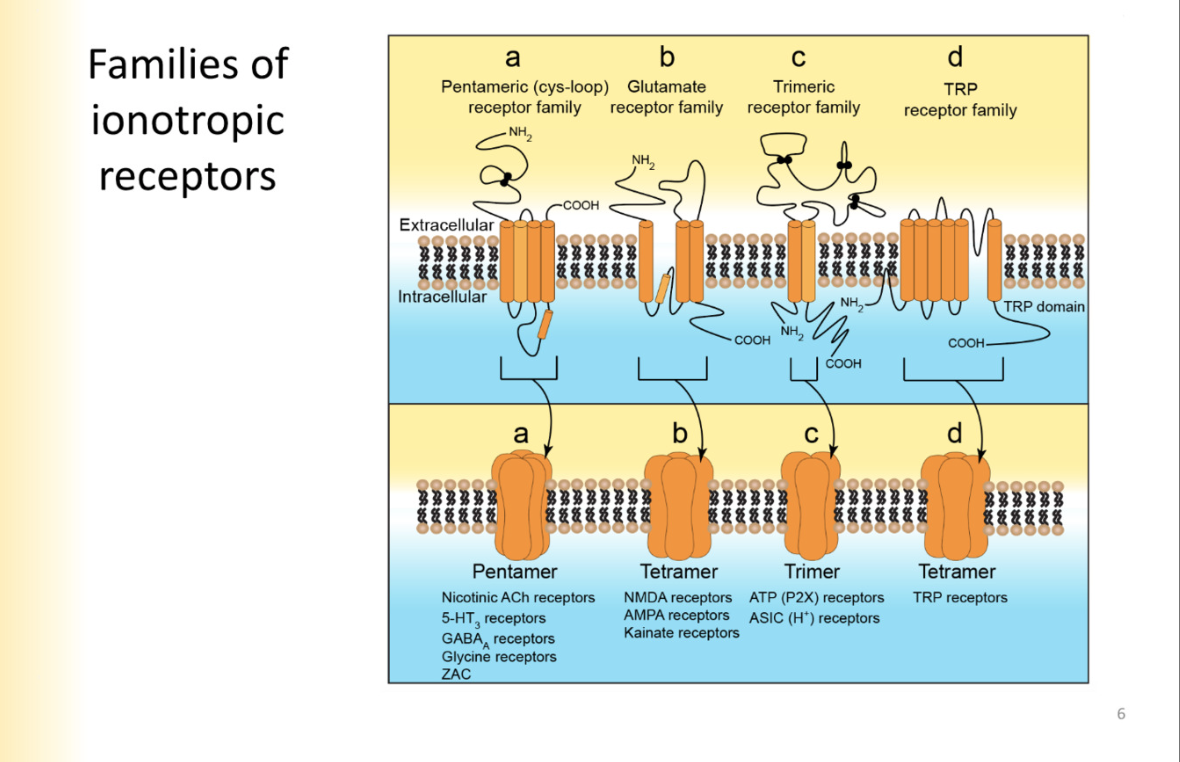
Families of ionotropic receptors
are ion channels
Pentameric channels - nicotinic ACH receptors, 5-HT receptors, GABAa receptors, Glycine, and zinc activated receptors
all comprised of 5 subunits
Trimers- ATP RECEPTORS acid sensing ion channels
(Tetramers)Glutamate Receptor Family- NMDA, AMPA, Kainate
TRP Receptor Family (Tetramers)- TRP receptors - transient receptor potential- not involved in synaptic transmission
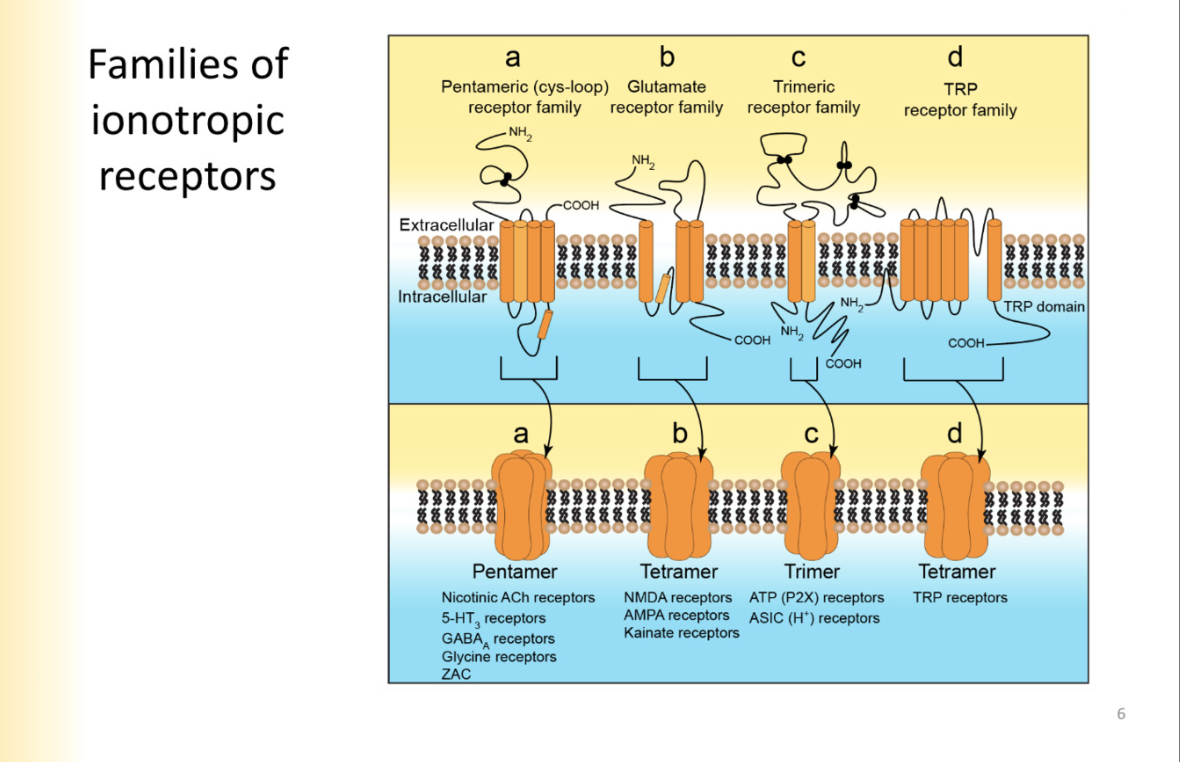
TRP Tetramer family vs Glutamate Receptor family
TRP tetramers have 6 subunits- p loop between 5 and 6
Glutmate- 3 complete transmembrane segments
second segment hooks into the membrane
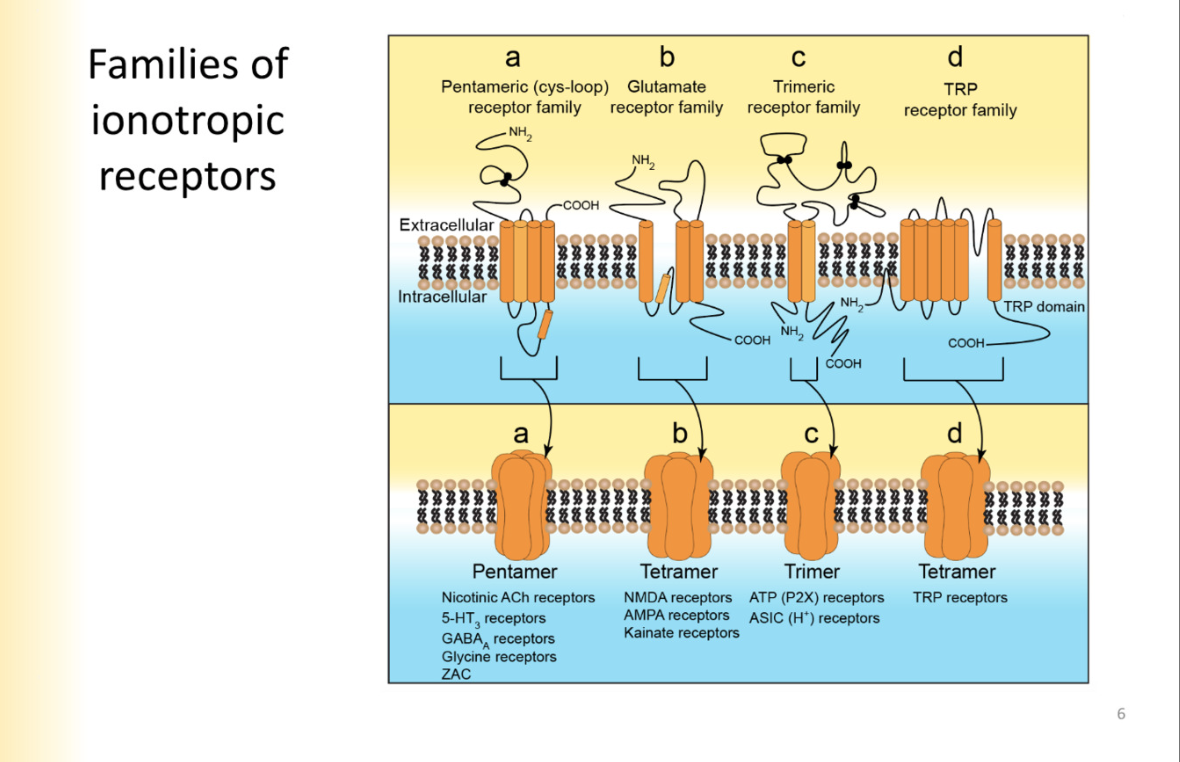
Trimers description
elaborate EC configuration of the protein, but only have 2 full transmembrane segments
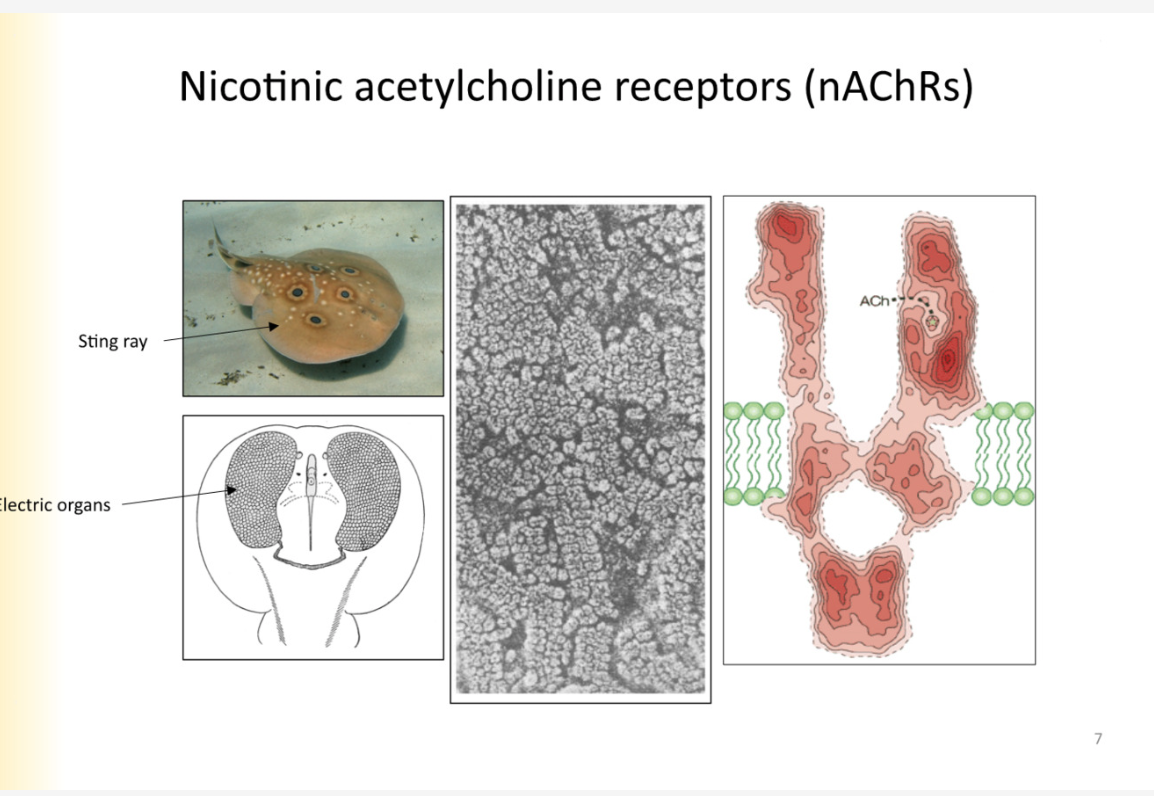
Nicotinic ACH receptors- stingray
stingrays uses ach to generate electricity to zap prey
electric organs are filled with ACH receptors
finding this was essential to classify ACH receptor - need a lot of protein in pure form
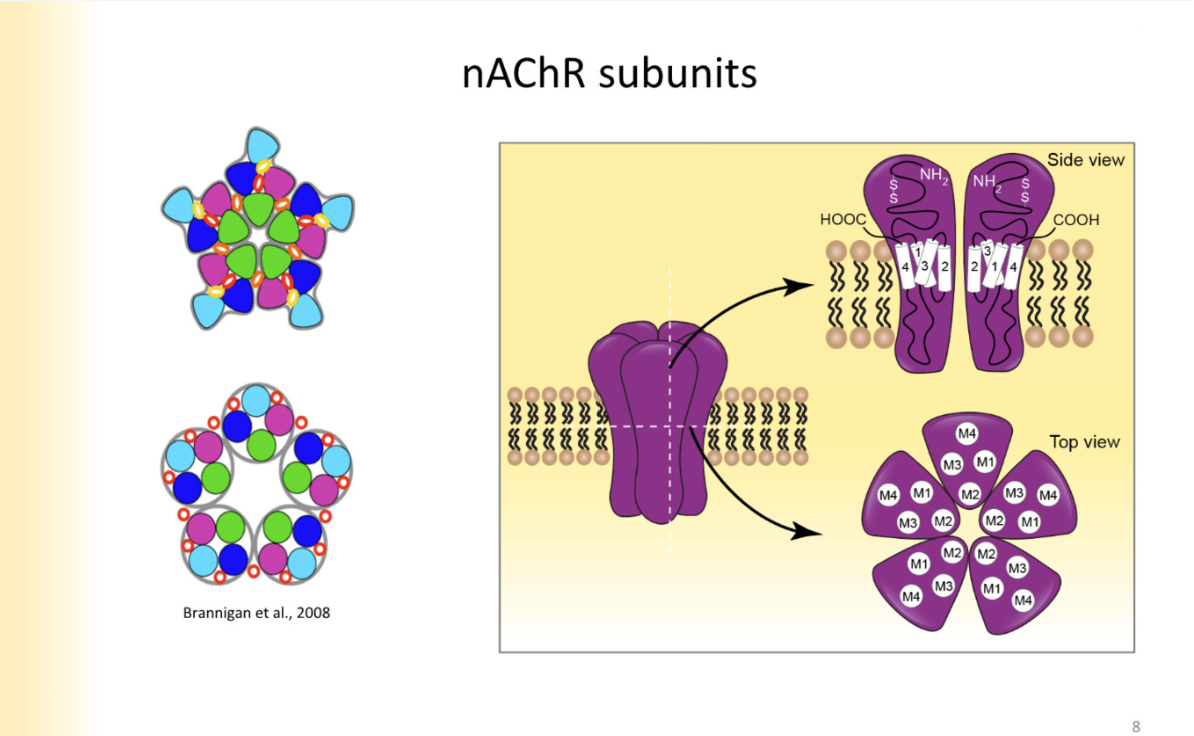
nACHR structure
top down view of closed channels- colors are diff transmembrane segments
4- transmembrane segments make up one subunit of it
M2 segments face thre pore of the channel- instead of pore lining segment
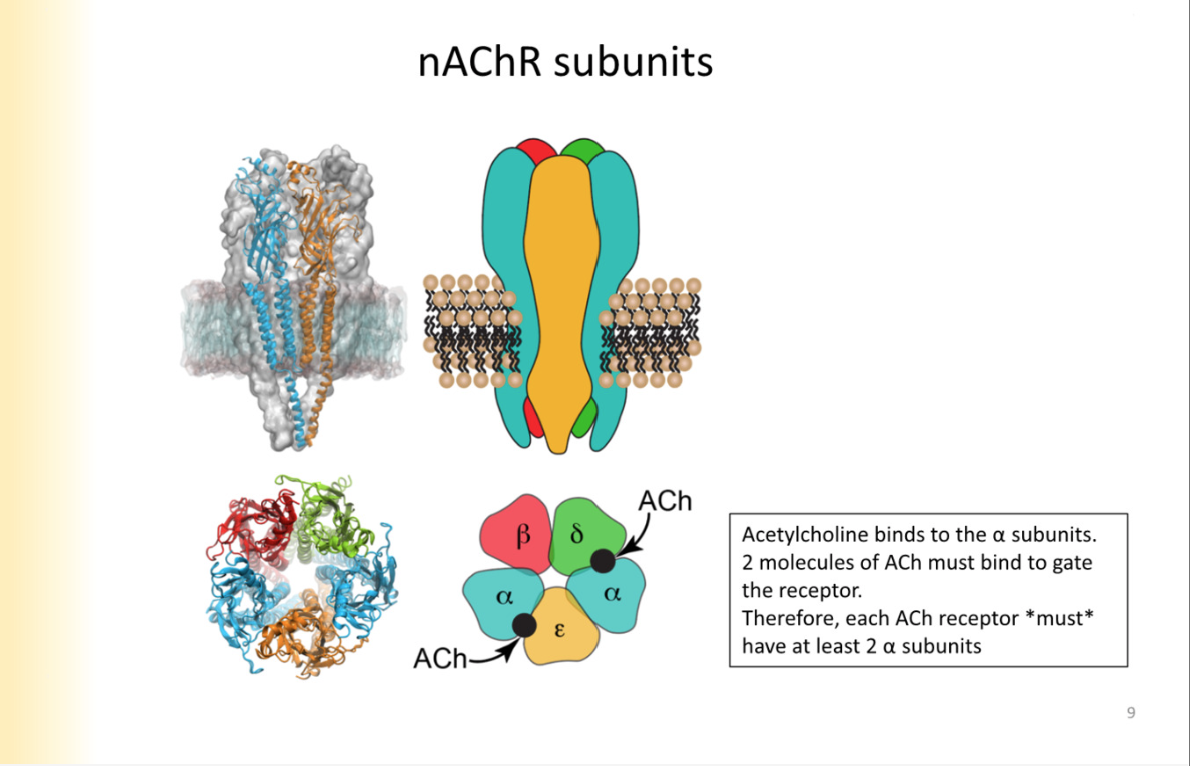
acHR subunits
alpha, beta, delta, epsilon
in order for receptor to open, two molecules of ACH have to bind on ALPHA SUBUNITYS ONLY- every receptor has to have atleast two alpha subunits
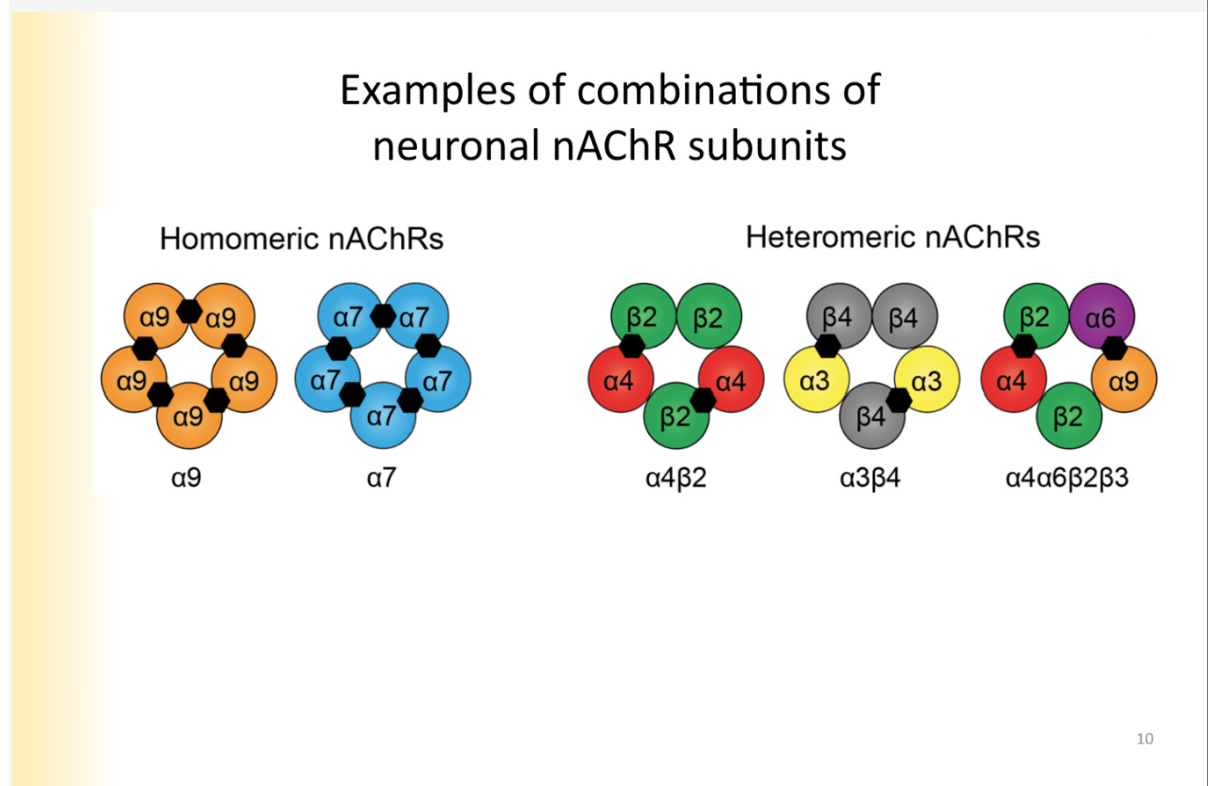
Examples of combinations of nACHR units
Homomeric nACHR’s - all made of the same subunits
heteromeric- not all of the subunits are the same
*still have to have 2 alphas
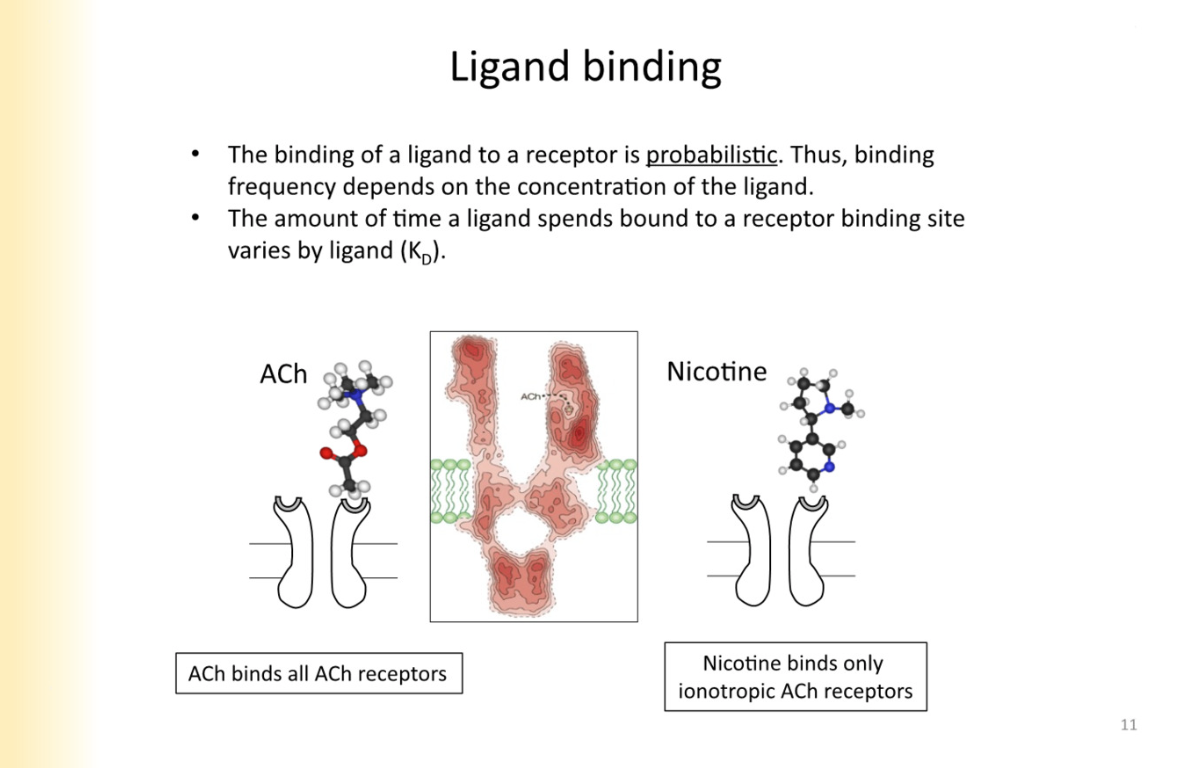
Ligand Binding - what does binding frequency depend on
ligand binding to receptor is purely probabalistic - depends on the concentration of the ligand
nicotine only binds to ionotropic ACH recpetors - specific agonist for it
ligand not always bound to receptor— it is on and off binding
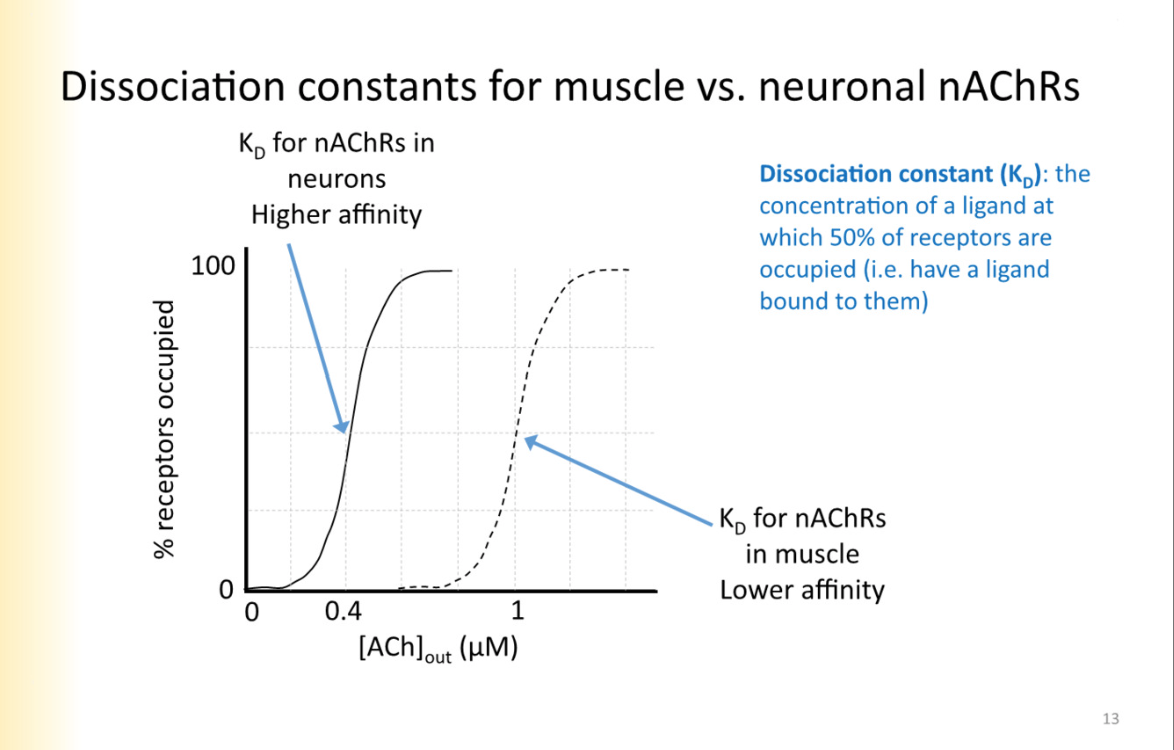
Kd information
measure of the strength binding
D= dissasociation
Greater kd, the more dissociating ligand from receptor
Kd tells you concentration that will induce binding at 50% of the receptors
when Conc=kd
Unbinding rate/binding rate
PIC: higher Kd- requires more concentration for 50% of receptors to be occupied
Agonist - ligand
compound that elicits the same biological effects as the naturally occuring (endogenous) ligand when it binds to receptor
nACHR agonists- nicotine
produces same EPSP
Antagonist - ligand
a compound that reduces or eliminates the effect of an agonist when bound to a receptor
ex. curare or alpha bungarotoxin- reducing the effect of the agonist or eliminating
compeitive antagonist
binds to the same site as an agonist(orthosteric binding) but does not activate the receptor. This reduces or prevents activation of the channel by an agonist
effects can be overcome by putting alot of agonist- outcompeting
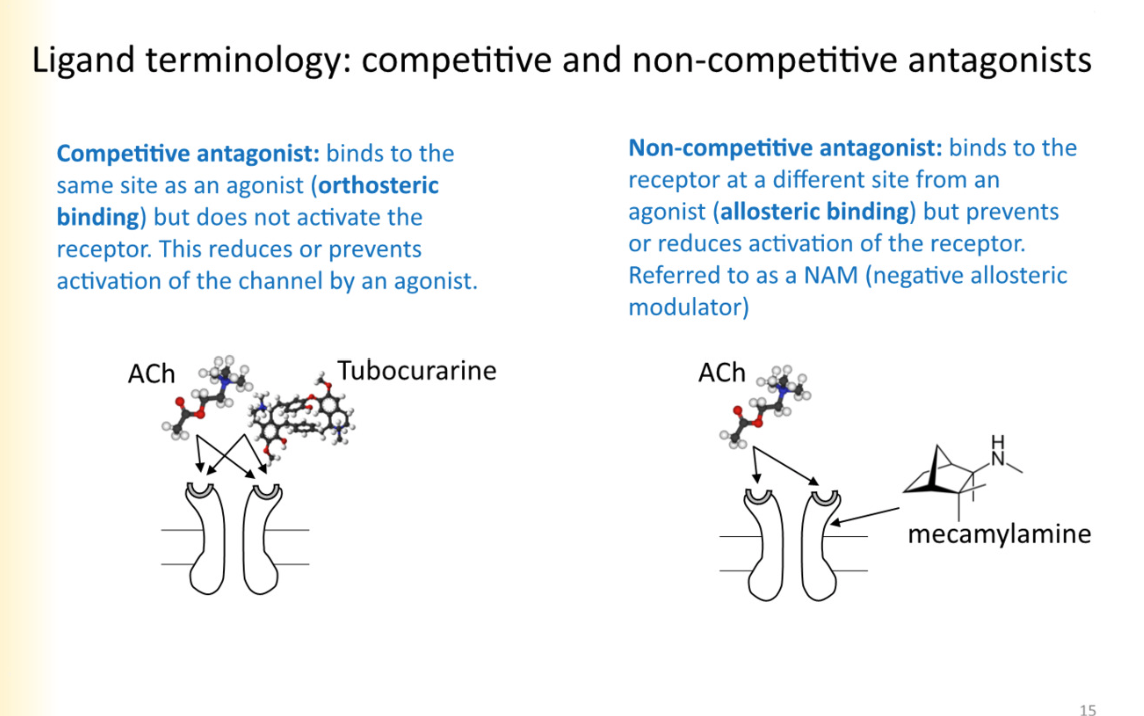
Non-competitive antagonist
Allosteric binding
binds to the receptor at a different binding site from an agonist, but prevents or reduces activation of the receptor
*negative allosteric modulator (NAM)
reversible vs irreversible antagonist
reversible: non covalently binds to the receptor, so can come off the receptor by “washing off”
non-reversible: binds covalently - so cannoy be washed off
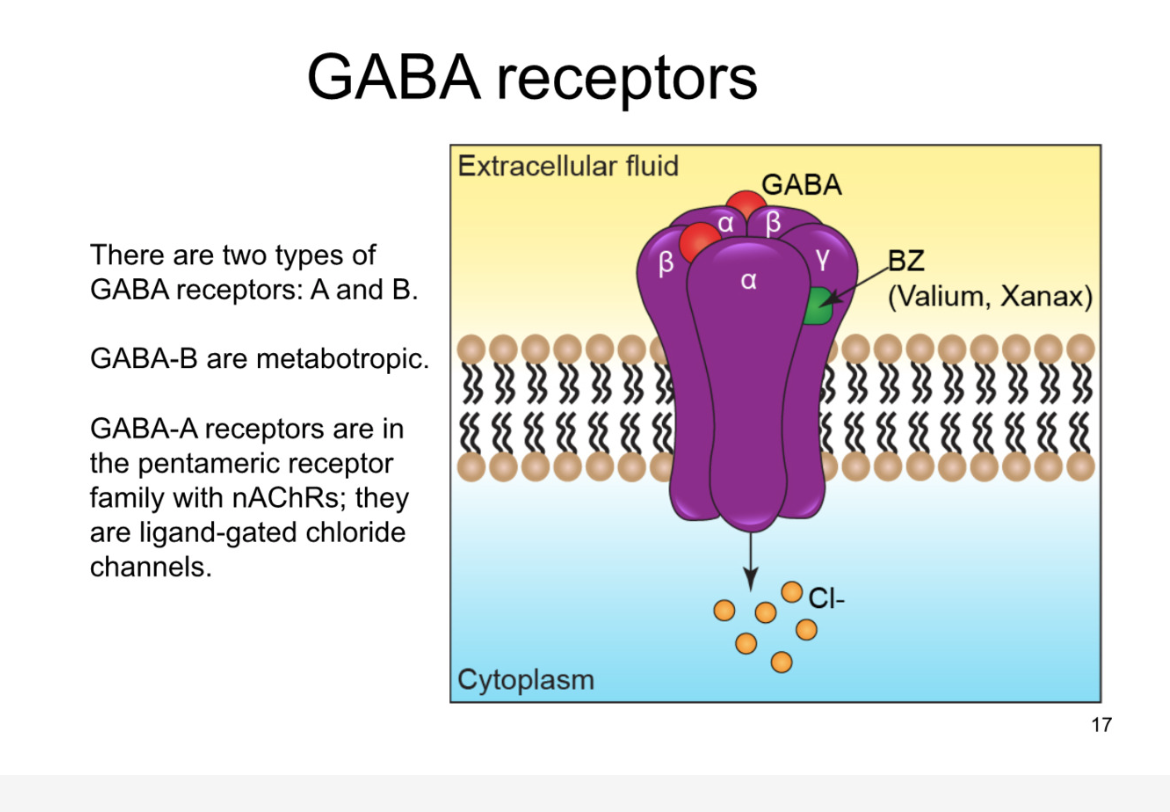
GABA recpetors
ionotropic GABA receptors are GABA A receptors
5 subunits, 4 transmem
in pentameric family
GABA activated chloride channels
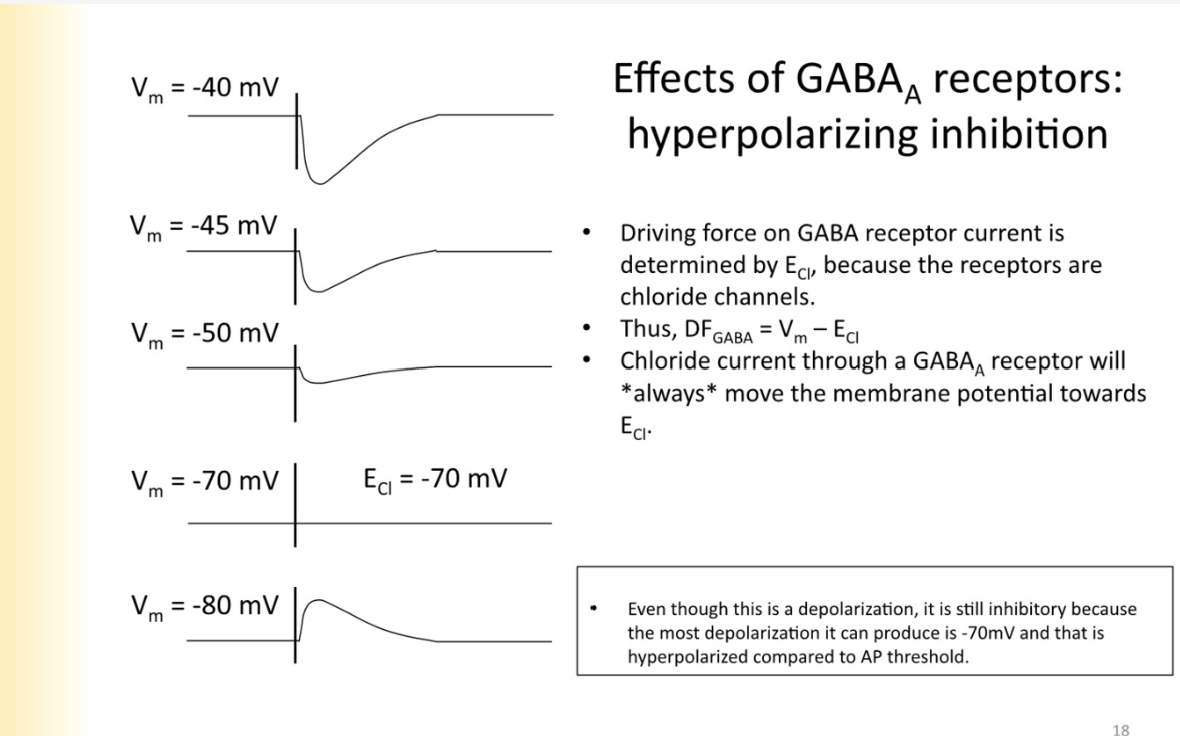
Effects of GABA receptors - what is driving force on these receptors determined by - why is it always inhibitory
equilibrium potential is affected by driving force on the cl- ions
equilibrium potential of cl- is the one that changes most
DF= VM-Ecl
eq CL=-70
-40mV- Df increasing, increase current inward
at -80 mV you would see a membrane depolarization- cl- efflux out of cell
STILL IS INHIBITORY — the most depolarized the cell can get from it is -70mV
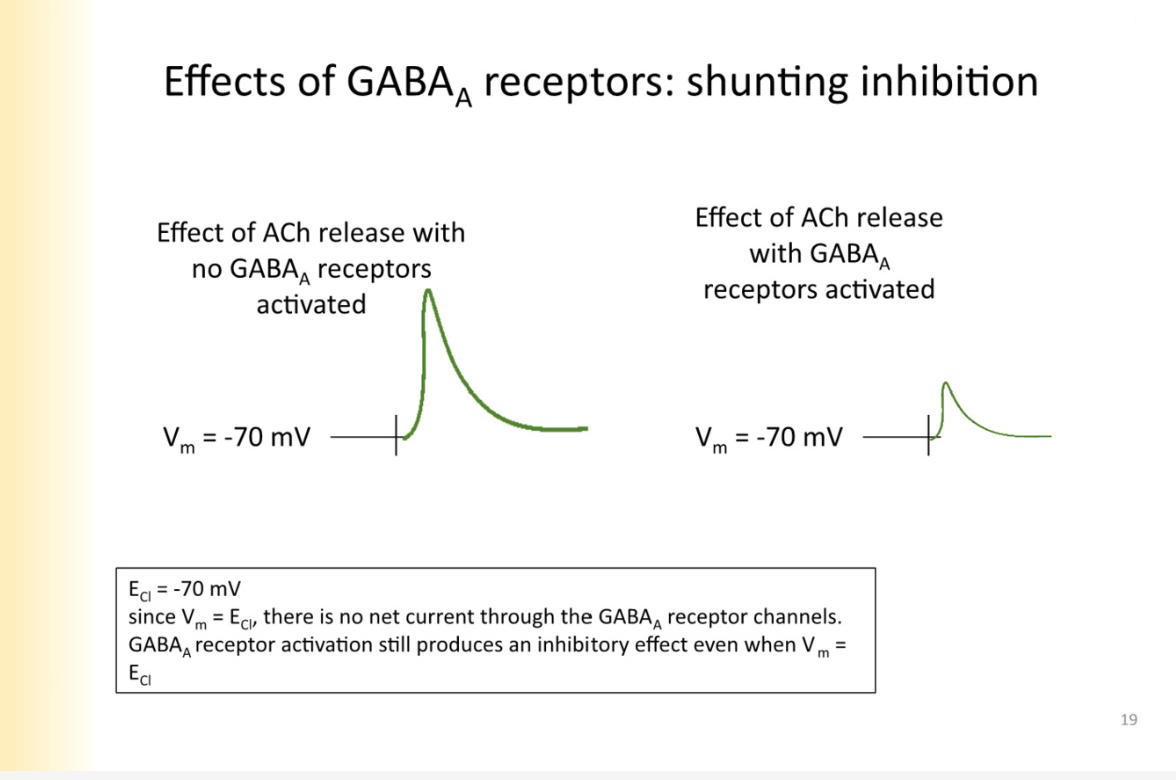
Shunting inhibition
open GABA receptors are providing another place for current to go and for ions to cross membrane
LOWERING resistance of the cell by opening ion channels
if put ach on cell when GABA receptors are activated it still shunts- WHY?
the membrane potential at any given moment is determined by the permeability of ions the cell is permeable to
as long as GABA receptors open, they have the -70 MV influence on cell
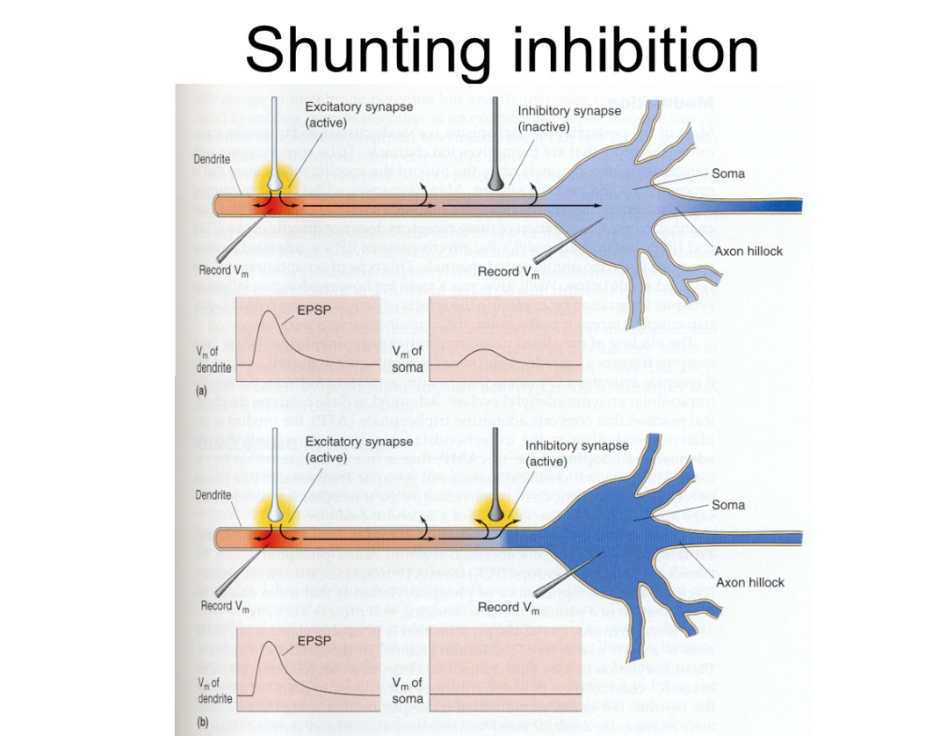
Shunting inhibition shown on neuron
inhibitory synapses right near the soma- strong on memebrane potential bc they do not have to travel very far
EPSP in dendrite and smaller in SOMA if nothing stops it
if you activate inhibitory synapse- excitatory synapse produces same epsp in dendrite, but no change in membrane potential in soma because opposite inhibitory response
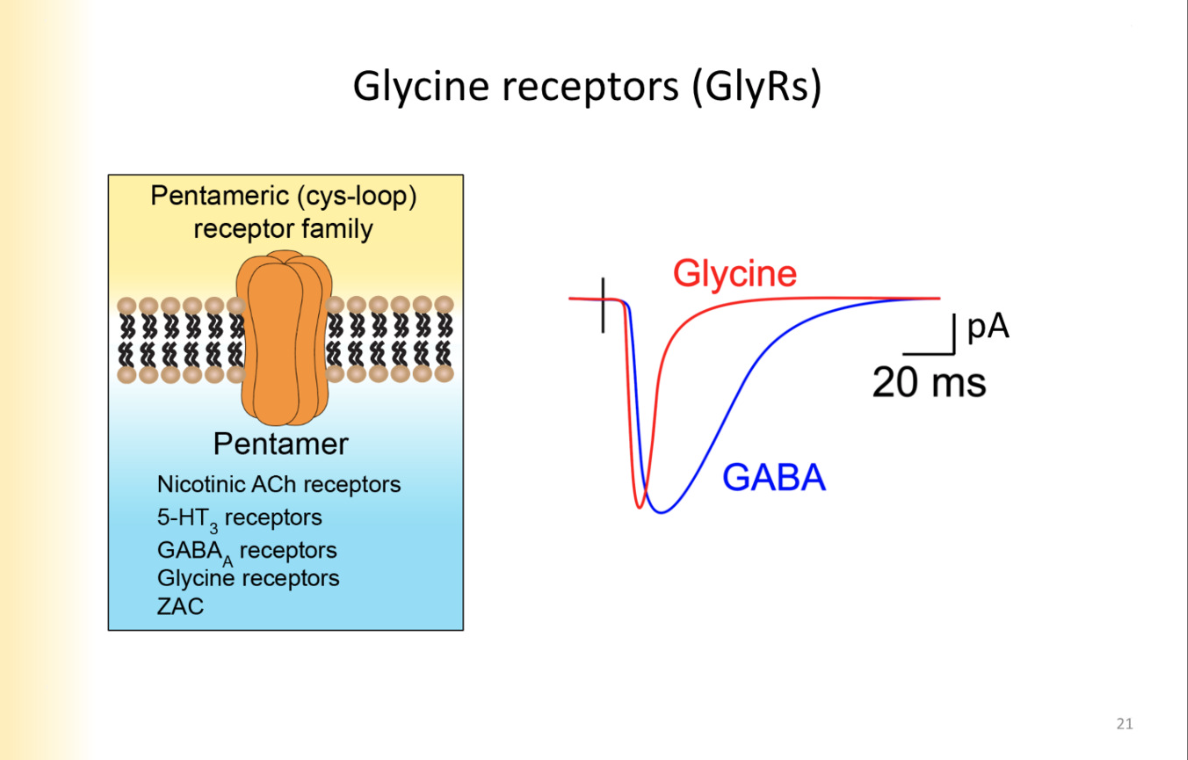
Glycine receptors
Glycine receptors are also chloride channels
same family, same structure
OPEN FASTER and shuts off faster
activated by glycine
Glutamate receptor family - explain structure of channel and types of receptors that exist within this family
Tetramers - 4 subunits to them
extracelluarly- 2 layers of globs
clamshell type structures with binding sites within them
EC portion- has NT binding sites and places where modifying agents can bind to them
NMDA, AMPA and kainate- receptors are ALSO activated by these compounds
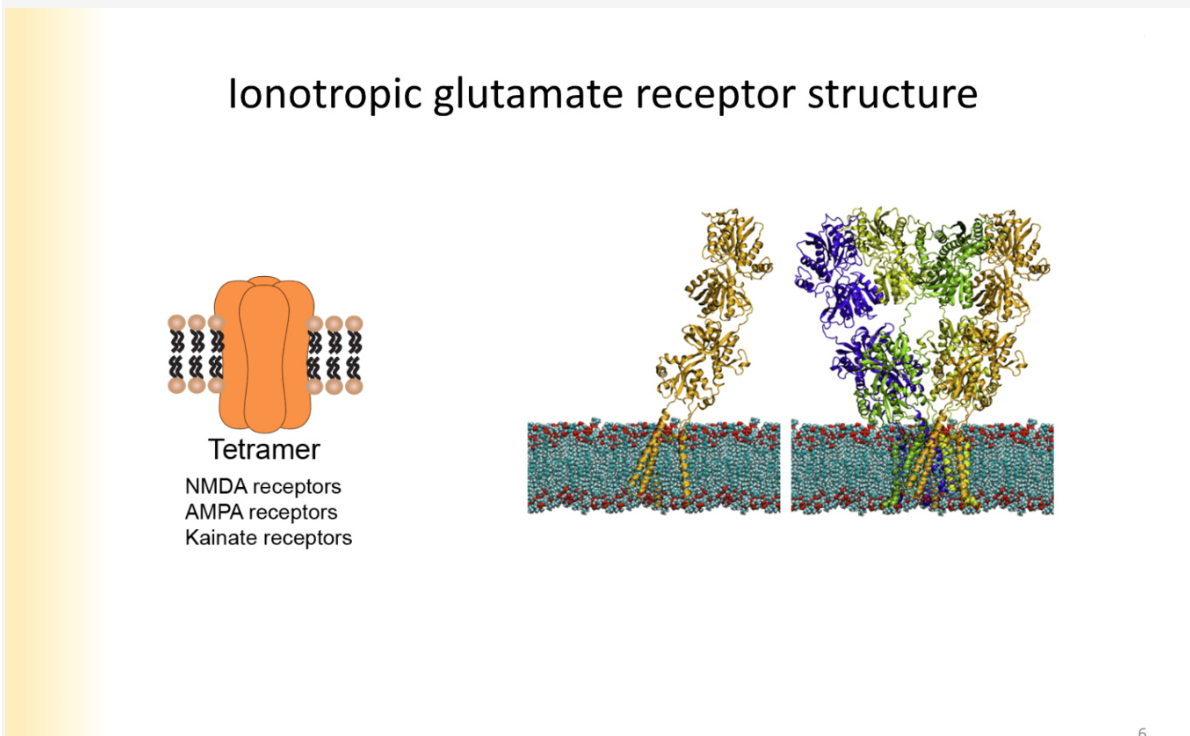
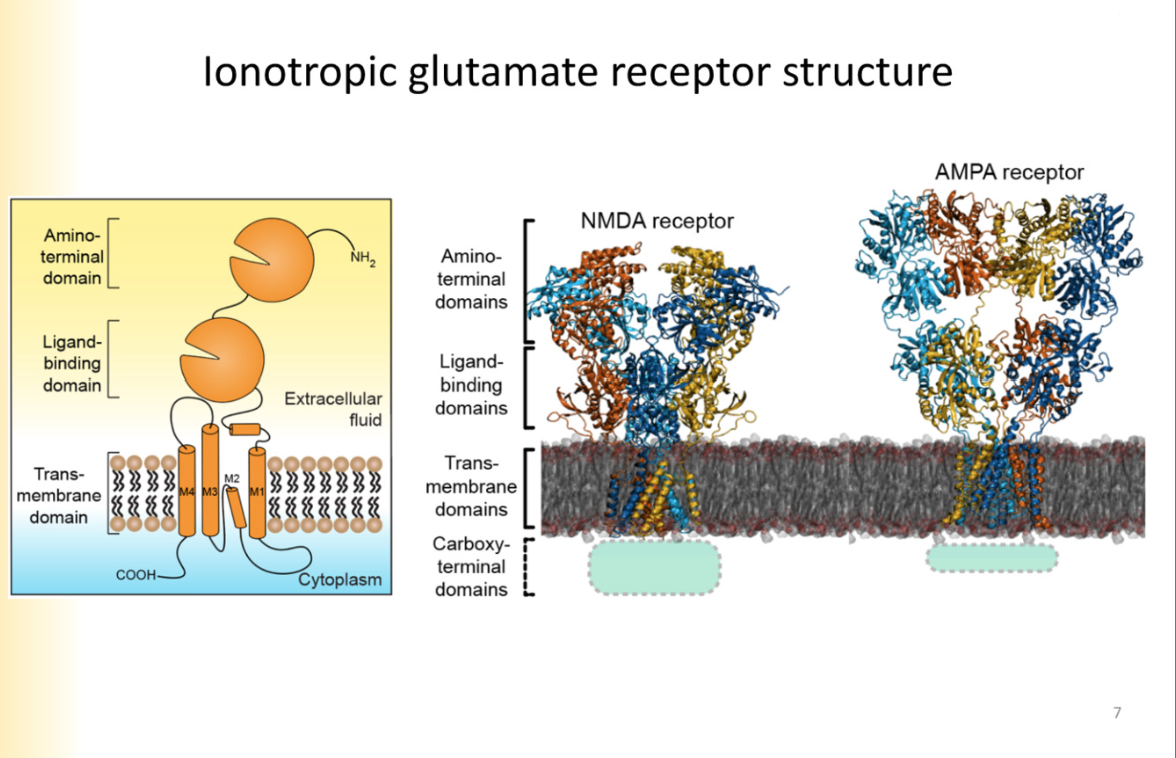
Structure of glutamate receptors
3 transmembrane segments- M2 segments creeps into membrane
represents a P loop- pore lining region
clamshell 1- ligand binding
clamshell 2- amino terminal domain is where modulating agents bind
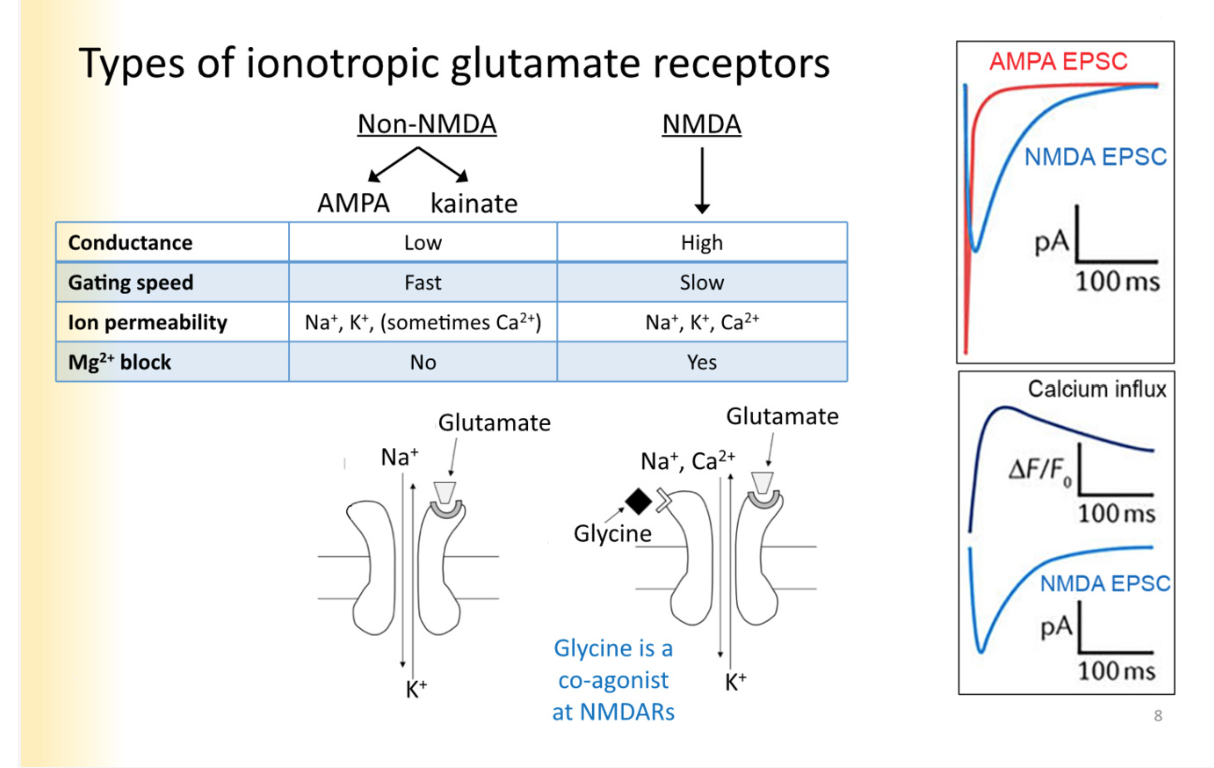
Types of ionotropic glutamate receptors - why is glycine not acknowledged in glutamate receptors and separate ones are called glycine receptors
Non NMDA vs NMDA
COMPARE THEM LOOK AT PICTURE
AMPA- fast, excitatory neurotransmission
low conductance
na, k, ca- sometimes ca2+ permeability
No Mg2+ block
at most active zones, reacting immediately to release of glutamate
NMDA
high conductance
slow gating speed
Na, k, ca- significant ca2+ permeability
yes mg2+ block
ca2+ flouresces when its bind to
AT REST, NMDA receptors are blocked by magnesium
pore of channel has mg2+ ion in it
Need two agonists to respective binding sites to activate- glutamate and glycine
Why do you focus on glutamate and ignore glycine: in most synapses, glycine is abundant. There is enough glycine to be bound to active sites- just waiting on glutamate
Why do glycine recpetors separately exist?
at post synaptic glycine synapses where glycine exists, glycine receptors only react to released glycine from vesicle and not free calcium all around the cell
Mg2+ block of glutamate receptors
At negative membrane potentials, there is Mg2+ in the pore
expelled upon depolarization
when inside of the cell is negative, mg2+ is drawn into pore, when depolarizes, repulsion happens
REFER TO PIC
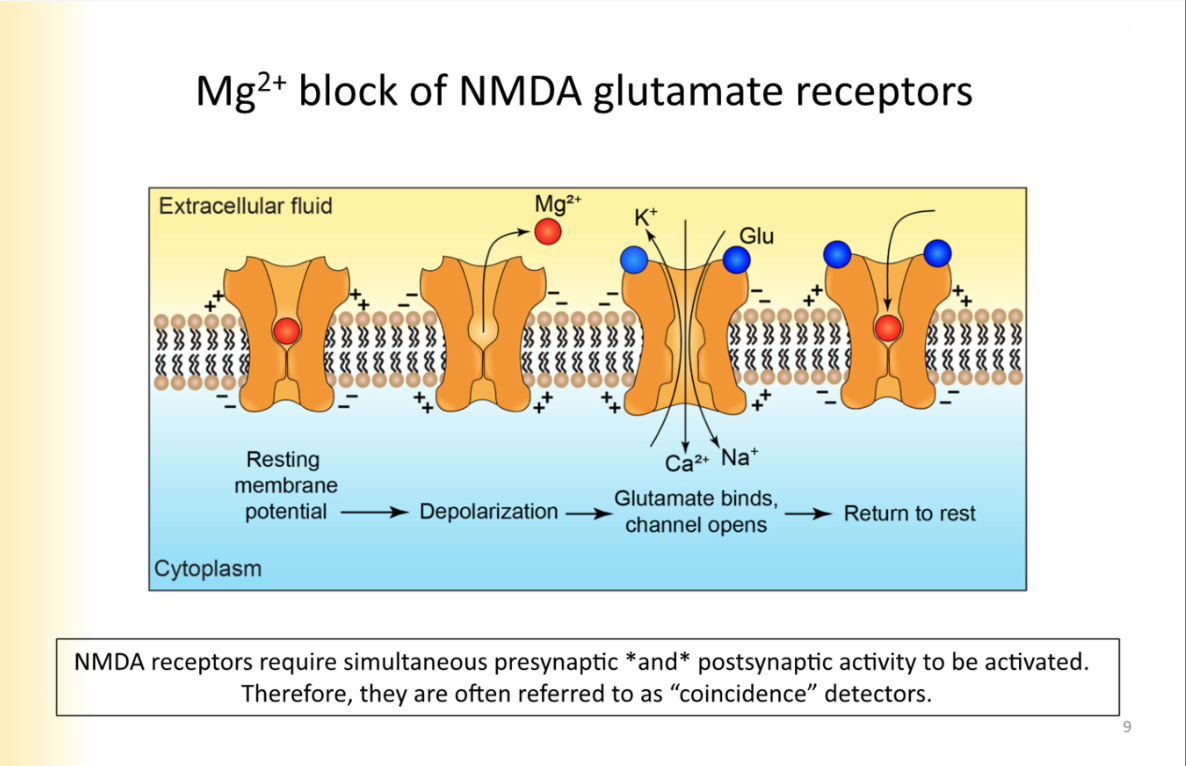
What presynaptic and post synaptic activity has to happen for NMDA glutamate receptors to work
glutamate is released from the synapse (however there is still MG2+ block)
Need multiple glutamate releases, and AMPA receptors at the same synapse
Glutamate binds to AMPA receptors
EPSP produced, post synaptic neuron is depolarized
Mg2+ block relieved
second glutamate release- NMDA receptors can work
IV plot of NMDA receptors and Mg2+ block- one with EC magnesium and one withh
without- similar to AMPA receptor
current determined by DF
reversal potential 0MV- calcium eq potential positive
WITH Mg2+
negative potentials, has mag2+ block
even if it has glutamate, receptor will not pass any current
by -10MV, all Mg2+ block gone, cell would pass the same current as if there were no mg2= at all
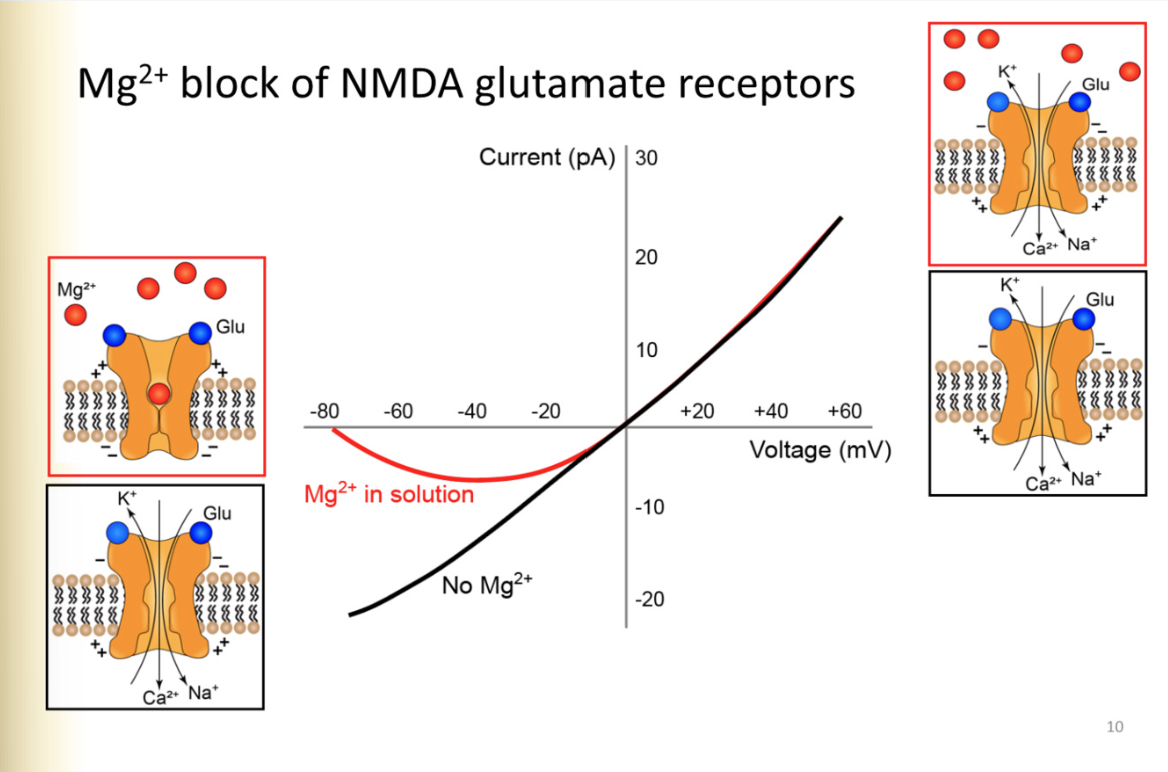
How does mg2+ presence affect current
little mg2+, a lot of current through NMDA receptor, and vice versa