PNS ir raumenų fiziologija
1/145
There's no tags or description
Looks like no tags are added yet.
Name | Mastery | Learn | Test | Matching | Spaced |
|---|
No study sessions yet.
146 Terms
audinys, kurio ląstelės membraninis potencialas geba kisti susidarant veikimo potencialui
Kas yra jaudrusis audinys?
· Nervinis audinys (išskyrus glijos ląsteles)
· Raumeninis audinys
Jaudriųjų audinių pvz
Veikimo potencialas sklinda tik jaudriuosiuose audiniuose. Padirginus šiuos audinius įvyksta staigūs membraninio (ramybės) potencialo (MP) pokyčiai, kaip pasekmė susidaro veikimo potencialas;
Jaudriųjų ląstelių membranos pasižymi atrankiu pralaidumu tam tikriems jonams
Jaudriesiems audiniams specifiškos savybės
Ląstelės membrana yra sudaryta iš dvigubo fosfolipidų sluoksnio, kuriame yra įterptinių baltymų.
Ląstelės membranos sandara
integralieji - baltymo molekulės perveria membraną; sudaro jonų kanalus;
periferiniai - išsidėsto išorinėje ar vidinėje membranos pusėje - pvz. išorėje receptoriai, vidinėje pusėje tarpininkai - G baltymai;
ATPazės - naudoja energiją.
Membranos baltymai
Baltymai, turintys vandens poras
Kas yra jonų kanalai?
bet kokia elektrinė membranos būsena (teigiama ar neigiama), bet nelygi 0 mV.
Kas yra poliarizacija?
MP tampa mažiau neigiamas nei ramybėje.(-70mV -> -50mV)
Kas yra depoliarizacija?
po depoliarizacijos membrana grįžta į ramybės potencialo būseną.(vėl elektroneigiamėja)
Kas yra repoliarizacija?
membrana tampa labiau poliarizuota (MP elektroneigiamenesnis nei ramybėje) (-70mV -> -90mV)
Kas yra hiperpoliarizacija?
Adekvatūs - prie jų poveikio jaudrieji audiniai evoliucijos eigoje yra prisitaikę (evoliuciškai susiformavo taip, kad receptoriai yra prisitaikę priimti dirginimą, pvz.: regos receptoriai reaguoja į šviesos bangos ilgį, klausos - prisitaikę priimti tam tikrą garso bangą);
Neadekvatūs - prie jų audiniai nėra prisitaikę (receptorius evoliucijos eigoje neprisitaikė, bet vis tiek geba reaguoti, pvz.: didelis triukšmas gali sukelti skausmą - nocireceptoriai, nors ir neprisitaikę priimti garso bangą, tačiau ypač stiprus garsas sukelia skausmo pojūtį.)
Dirgiklių klasifikacija pagal fiziologinę reikšmę
fiziniai (mechaniniai, temperatūros, garso, šviesos, elektriniai;
cheminiai (įvairios cheminės medžiagos);
fiziniai - cheminiai (pH, osmosinio slėgio, elektrolitų sudėties pokyčiai).
Dirgiklių klasifikacija pagal prigimtį
ikislenkstinis (nesukelia atsako - VP (veikimo potencialo) - nors sukelia depoliarizaciją; audinys nereaguoja į dirgiklius);
slenkstinis (silpniausias dirgiklis, kuris sukelia VP.);
viršslenkstinis (stipresnis už slenkstinį, sukelia VP.) Dar skirstomas:
submaksimalus - kiek didesnis už slenkstinį; juo dirginant raumuo nesusitraukia maksimaliai
maksimalus - sukelia maksimalų susitraukimą; juo dirginant raumuo susitraukia maksimaliai
supramaksimalus - didesnis už maksimalų dirgiklis, tačiau raumuo stipriau jau nebesusitraukia, o atvirkščiai - mes galime jam taip pakenkti
Dirgiklių klasifikacija pagal stiprumą
tai aplinkos ar vidinis organizmo veiksnys, kuris pasiekęs tam tikrą laipsnį ir greitį (slenkstį), sužadina audinio ar organo veiklos bei būklės pakitimus
Kas yra dirgiklis?
procesas, kai audinys ar organas veikiamas dirgikliais. Dirginimo metu vyksta ląstelės membranos depoliarizacija, pasiekiama slenkstinė įtampa ir randasi veikimo potencialas
Kas yra dirginimas? Kas vyksta jo metu?
ląstelės, audinio ar organo gebėjimas reaguoti į dirginimą
Kas yra dirglumas/jaudrumas?
· Atrankus laidumas. Atrankiais jonų kanalais per membraną keliauja tik tam tikri jonai. Tai priklauso nuo jono dydžio ir gebėjimo prisijungti vandens molekules - daugiau jų prisijungęs jonas tampa didesnis ir negali praeiti per porą;
· Funkcinis specifiškumas. Pasyvieji jonų kanalai atsakingi už membranos ramybės potencialą, cheminiai jonų kanalai lemia sinapsinio potencialo susidarymą neurone, dėl elektrinių jonų kanalų susidaro ir plinta veikimo potencialas VP;
· Jonų kanalai gali būti pasyvūs ir aktyvūs,
· Būna išsidėstę tam tikrose membranos vietose.
Jonų kanalų savybės
· pasyvūs - visada atviri, jonai jais juda nuolat pagal elektrocheminį gradientą (iš didesnės konc. į mažesnę). Dėl jonų judėjimo palaikomas ląstelės membraninis potencialas (ramybės potencialas).
· aktyvūs - ramybėje yra uždari, turi dažnai turi tam tikras struktūras pvz vartus (įtampai jautrūs - ramybėje uždari, o laikui jautrūs - ramybėje atviri), kurie atsidaro, kai yra tam tikros sąlygos. Skiriami cheminiai, elektriniai ir mechaniniai.
Jonų kanalų skirstymas
atsidaro, kai kinta membranos elektrinis krūvis (t.y. kai ląstelė padirginama) - poliarizacija, pvz.: elektriniai Na+ arba K+ jonų kanalai. Ramybės būsenos jaudriosios ląstelės - membranos elektriniai kanalai yra uždari. Susidarant veikimo potencialui, keičiasi įtampa abipus membranos - tai skatina vienus elektrinius jonų kanalus atsidaryti, kitus užsidaryti. Vartams atsivėrus jonai juda kanalu. Dėl jonų judėjimo per šiuos kanalus keičiantis krūviams formuojasi veikimo potencialas VP (nervinis impulsas). Jis sklinda per visą aksono ilgį, kol pasiekia sinapsę.
Apibūdink elektrinius jonų kanalus
Ramybėje uždari, atsidaro, kai prie kanalo jungiasi cheminė medžiaga (mediatorius). Kanalas atsiveria ir juo laisvai gali judėti jonai. Neuromediatoriai kaip acetilcholinas ir gama aminosviesto rūgštis jungiasi prie neurono membranos cheminių kanalų (receptorių) ir šie atsiveria. Yra sinapsėse, per juos perduodamas sinapsinis potencialas iš vieno neurono į kitą į postsinapsinę dalį.
Apibūdink cheminius jonų kanalus
būdingi kraujagyslių sienelei, dažniausiai tai Ca2+ jonų kanalai, jie atsidaro kraujagyslei atsipalaidavus, o jai susitraukus - užsidaro, valdomi mechaniškai;
Apibūdink mechaninius jonų kanalus
· Aktyvaciniai (įtampai jautrūs) vartai - ramybėje uždari.
· Inaktyvaciniai (laikui jautrūs) vartai - ramybėje atviri.
Jonų kanalų vartų skirstymas
· Įtampai jautrūs vartai atsiveria depoliarizuojantis ląstelės membranai
· Laikui jautrūs inaktyvaciniai vartai užsiveria praėjus tam tikram laiko tarpui nuo elektrinų kanalų atsidarymo ir sustabdo Na jonų judėjimą kanalais
Natrio vartų skirstymas
· pasyvūs jonų kanalai išsidėstę visame neurono membranos paviršiuje,
· cheminiai jonų kanalai daugiausia išsidėstę dendritų ir neurono kūno membranoje,
· elektriniai kanalai vyrauja aksono stormenyje, visame nemielininio aksono membranos paviršiuje ir mielininio aksono Ranvjė sąsmaukose.
Jonų kanalų išsidėstymas neurono membranoje
Pasyvioji pernaša gali vykti tiek pasyviaisiais, tiek aktyviaisiais jonų kanalais
Per ką gali vykti pasyvioji pernaša?
Kai kurios dujų molekulės (CO2, O2, N2) arba krūvio neturinčios molekulės (pvz etanolis) pasyviai difunduoja per membraną nedalyvaujant baltymų molekulėms
Ar baltymų molekulės visada dalyvauja pernašoje?
potencialų skirtumas abipus membranos ramybės metu, kai ląstelės pamatinė membrana išorėje įelektrinta teigiamai, o viduje - neigiamai. Susidaro dėl pasyviųjų kanalų.
Kas yra membraninis (ramybės) potencialas? Kodėl jis susidaro?
Nejaudriųjų -90 mV; jaudriųjų -70 mV
Koks nejaudriųjų ląstelių membraninis potencialas? Jaudriųjų?
Nejaudriosios ląstelės turi tik K kanalus
Kodėl skiriasi jaudriųjų ir nejaudriųjų ląstelių membraninis potencialas?
stambios baltymų molekulės ir anijonai ląstelės viduje
Kas sukelia neigiamą krūvį ląstelėje?
Membranos laidumas tam tikriems jonams priklauso nuo jonų kanalų skaičiaus ir dydžio
Nuo ko priklauso membranos laidumas jonams?
Na+ vid. 15 mmol/l; už 150 mmol/l
K+ vid. 150 mmol/l; už 5 mmol/l
Cl- vid. 10 mmol/l; už 125 mmol/l
Na, K, Cl jonų viduląstelinė ir užląstelinė koncentracija
Koncentracijos gradientas Na+ jonams nukreiptas tokia pat kryptimi kaip elektrinis potencialas, todėl susidaro labai stiprus elektrocheminis gradientas, skatinantis Na+ jonus judėti į ląstelę.
Kas skatina Na+ judėti į ląstelę?
Neuronų membrana yra atrankiai laidi K+, Na+, Cl- jonams ir nelaidi stambiems baltymams bei ląstelės viduje esantiems anijonams.
Neuronų membranos laidumas
Koncentracijos gradientas skatina K judėti iš ląstelės (iš didesnės konc. į mažesnę), išorinė ląstelės membranos pusė tampa dar teigiamesnė. K vis difunduojant iš ląstelės elektrinis potencialas didėja. Susitelkus dideliam K kiekiui, K ima judėti elektrinės jėgos kryptimi (iš didesnio krūvio į mažesnį – iš išorės į vidų) atgal į ląstelę. Elektrocheminė jėga, dėl kurios vyksta K judėjimas per membraną, nėra didelė, nes jėgų kryptys nukreiptos į skirtingas puses (cheminė į išorę, skatina K judėti iš ląstelės, elektrinė į vidų), bet laidumas K didelis. Susidaro elektrocheminis gradientas. Koncentracijos gradientas Na jonams nukreiptas tokia pat kryptimi kaip ir elektrinė jėga (į vidų), tad susidaro stiprus elektrocheminis gradientas, skatinantis Na jonus judėti į ląstelės vidų. Na jonų elektrocheminis gradientas yra labai stiprus, tačiau membrana nėra labai laidi Na. Todėl jeigu ramybės potencialas yra -70 - K juda iš ląstelės, o Na į ląstelę. Membranoje esantis baltymas Na+/K+ siurblys/ATPazė aktyviai prieš elektrocheminį gradientą (naudodamas ATP energiją) į ląstelę grąžina 2 K+ ir iš ląstelės išneša 3 Na+. Šis baltymas nedalyvauja susidarant pačiam MP, bet yra svarbus palaikant jį normos ribose.
Membraninio potencialo palaikymas
Pasyviais
Kuriais kanalais palaikomas membraninis potencialas?
nervinio impulso susidarymas aksono stormenyje ir tolimesnis plitimas nervine skaidula iki sinapsės; tai staigus ramybės potencialo pokytis nuo -70 iki +30 ir vėl -70.
Kas yra veikimo potencialas?
slenkstinis dirgiklis
Būtina sąlyga veikimo potencialui
Veikimo potencialas kyla neurono aksono stormenyje, kur yra daugiausia elektrinių jonų kanalų.
Kur kyla veikimo potencialas? Kodėl?
Veikimo potencialas susidaro, kai sinapsinis potencialas iš dendritų ir neurono kūno pasiekia aksono stormenį, sumuojasi ir membraninis potencialas tampa teigiamesnis -> padirginus nervinę ląstelę elektros srove membranos potencialas teigiamėja, įvyksta depoliarizacija. Depoliarizuojantis aksono stormeniui, atsiveria elektriniai Na+ kanalai ir membranos laidumas natriui labai padidėja. Pagal elektrocheminį gradientą Na+ juda į ląstelę. Dirginant slenkstiniu dirgikliu, neurono membranos depoliarizacija pasiekia slenkstinę įtampą (apie –55 mV) ir kyla veikimo potencialas – staigus ramybės potencialo pokytis. Membranos depoliarizacijai pasiekus slenkstį, atsiveria dar daugiau elektrinių Na+ jonų kanalų, membrana dar labiau depoliarizuojasi ir skatina atsiverti dar daugiau Na+ kanalų. Tai vadinama teigiamu grįžtamuoju ryšiu. Depoliarizacija baigiasi užsidarius laikui jautriems elektrinių Na+ kanalų vartams ir atsivėrus elektriniams K+ jonų kanalams. Elektriniai K+ jonų kanalai lėtai reaguoja į depoliarizaciją. Jie ima atsiverti tik veikimo potencialui pasiekus piką. Depoliarizacija baigiasi, kai K+ pradeda skverbtis iš ląstelės. Silpnėjant Na+ jonų srautui į ląstelę ir didėjant K+ jonų srautui iš ląstelės, membranos potencialas vėl tampa neigiamas - repoliarizacija
Veikimo potencialo susidarymas ir plitimas nervine skaidula
Įvykus membranos repoliarizacijai, daugelio neuronų membranų lėtieji K+ kanalai dar kurį laiką lieka atviri. Tuomet K+ jonai vis dar plūsta iš neurono – membranos potencialas tampa neigiamesnis nei ramybės metu. Šis procesas vadinamas hiperpoliarizacija. Hiperpoliarizacijos pabaigoje visi K+ kanalai jau būna uždari.
Ką daro K kanalai įvykus repoliarizacijai?
Kilus veikimo potencialui, kito veikimo potencialo sukelti neįmanoma - neuronas yra nejaudrus. Didžioji dalis Na+ jonų kanalų tuo metu yra neaktyvi ir negali atsiverti, nors ir veikia stiprus dirgiklis. Dauguma K+ jonų kanalų tuo metu yra atviri. Tai vadinama absoliučiu refrakteriniu (nejaudros) periodu.
Kas yra absoliutus refrakterinis periodas?
Kai užsidaro visi K kanalai
Kada ląstelė grįžta į ramybės būklę?
Veikia pagal ,,Jėgos dėsnį'' - silpnas dirgiklis sukelia silpną depoliarizaciją; stiprus dirgiklis sukelia stipresnę. Na jonų judėjimas vyksta, bet slenksčio nėra. Nėra VP, todėl ir nesklinda aksonu. Nėra refrakterinio periodo, nes nėra hiperpoliarizacijos, VP.
Vietinės depoliarizacijos (dietinio potencialo) ypatumai
Veikimo potencialas - ,,viskas arba nieko'' - yra slenkstinis dirgiklis, potencialas susidaro; nėra - nėra VP. Vietinė depoliarizacija - ,,jėgos'' dėsnis;
Veikimo potencialas - yra slenkstis; vietinė depoliarizacija - nėra slenksčio;
Veikimo potencialas - sklinda visa aksono membrana iki sinapsės; vietinė depoliarizacija nesklinda;
Veikimo potencialas pasižymi refrakteriškumu, vietinėje depoliarizacijoje nėra refrakterinio periodo.
Palygink veikimo potencialą su vietine depoliarizacija
Motorinių neuronų 0,001 s (1 ms)
Griaučių skersaruožių raumenų 0,02-0,03 s (20-30 ms)
Širdies 0,3 s (300 ms)
Refrakterinio periodo trukmė
didžioji dalis Na jonų kanalų yra neaktyvūs, nors ir veikia stiprus dirgiklis, dauguma K jonų kanalų tuo metu yra atviri
Jonų kanalai refrakteriniu periodu
Po absoliutaus refrakterinio periodo neurone gali kilti kitas veikimo potencialas, bet tik jei neuronas bus dirginamas gerokai stipresniu už slenkstinį dirgikliu. Taip yra todėl, kad dalis Na+ kanalų dar vis neaktyvūs, o dalis K+ kanalų vis dar atviri.
Ar po refrakterinio periodo gali kilti kitas veikimo potencialas? Kodėl?
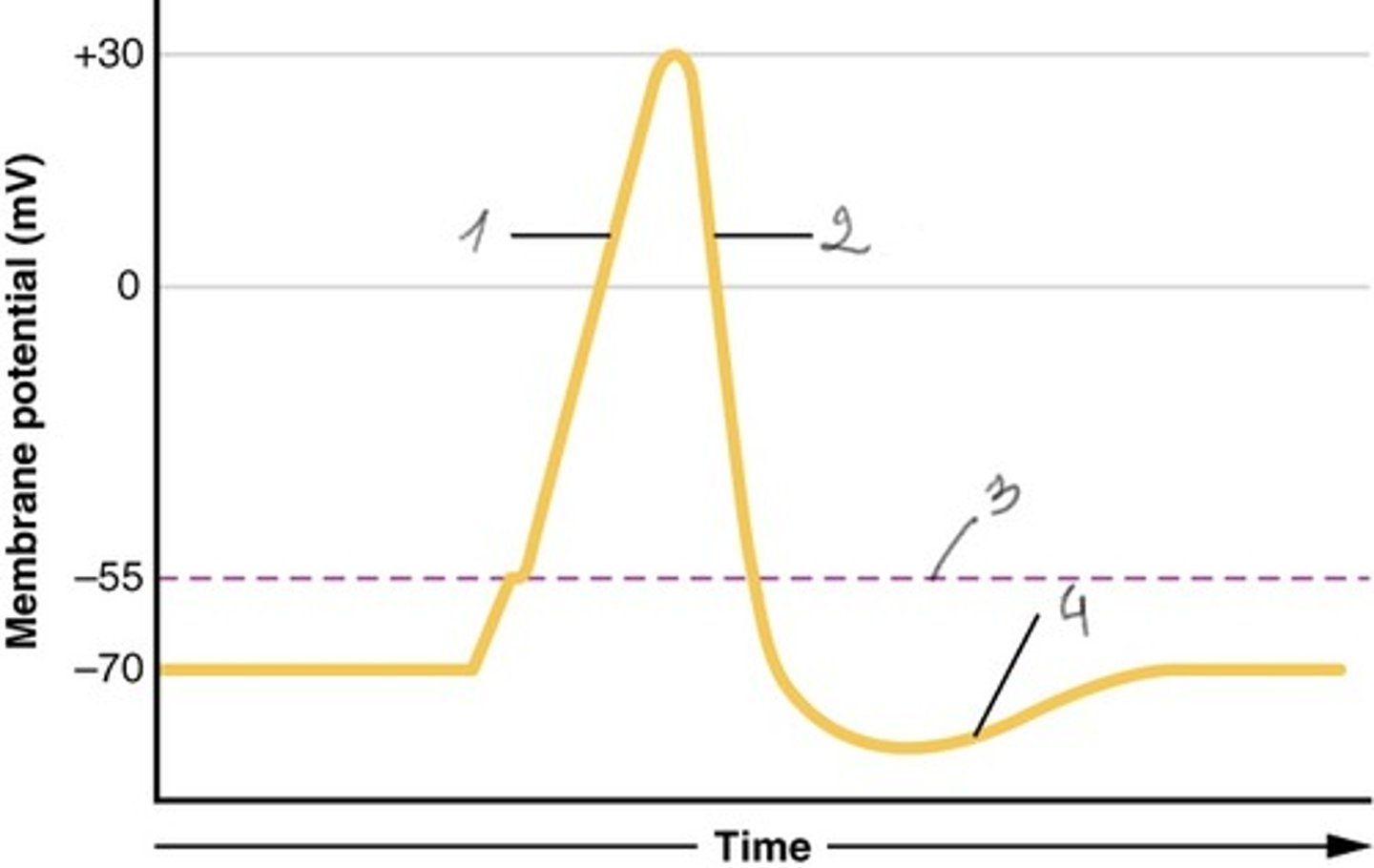
Veikimo potencialo kreivė
1. Depoliarizacija
2. Repoliarizacija
3. Slenkstinė įtampa
4. Hiperpoliarizacija
Elektriniai Na+ ir K+ jonų kanalai išsidėstę visame aksono membranos paviršiuje. Tarp sudirgintos ir nesudirgintos aksono membranos dalies susidaro ratinės srovės: veikimo potencialas susidaro neurono aksono stormenyje ir pirmiausia vyksta depoliarizacija, Na juda į ląstelę sukeldamas krūvių pasikeitimą (inversiją), ląstelės vidus teigiamėja. Gretima membranos dalis vis dar neigiama, yra santykinėje ramybėje. Prasideda vietinė depoliarizacija, nes Na+ kanalai nepakankamai atsidarę, tarp sujaudintos ir nesujaudintos aksono sričių susidaro "ratinės" srovės (srovė kinta iš + į -). Po kurio laiko ta membranos vieta, kurioje vyko depoliarizacija, tampa repoliarizuota, nes veikimo potencialas pasiekia piką. Užsidarius Na+ el. kanalų laikui jautriems vartams atsidaro K el. kanalai. K+ iš ląstelės -> įvyksta krūvių inversija ir vidus tampa neigiamas, o depoliarizuojasi gretima sritis, kuri prieš tai buvo vietinės depoliarizacijos stadijoje. Grįžimo nėra, membrana absoliučiame refrakteriniame periode; todėl nervinis impulsas gali sklisti tik viena kryptimi - link sinapsių.
Nervinio impulso sklidimas nemielinine skaidula
Aksonas padengtas mielino sluoksniu, kas padidina membranos varžą, sumažina jonų laidumą per membraną. Ranvjė sąsmaukose yra susikaupę elektriniai Na+ ir K+ jonų kanalai. Nervinis impulsas plinta tiktai Ranvjė sąsmaukomis, nes tik čia yra elektriniai Na+ ir K+ jonų kanalai. Depoliarizacijai įvykus vienoje Ranvjė sąsmaukoje, Na+ plūsta į tą sąsmauką sukeldami krūvių inversiją (+ viduje, - išorėje), o greta esanti Ranvjė sąsmauka yra dar nesujaudinta (vidinė membranos pusė -, išorinė +). Tarp sujaudintos ir nesujaudintos sąsmaukų susidaro "ratinė" srovė, kuri plinta panašiai kaip ir nemielininėje nervinėje skaiduloje. Būdingas šuoliškas/saltatoriškas plitimas (srovės juda peršokdamos meilininį dangalą), panaudoja mažiau energijos, plinta greičiau
Nervinio impulso sklidimas mielinine skaidula
Mielinas yra labai geras nervinio impulso izoliatorius (kelis tūkstančius kartų sumažina laidumą), todėl nervinis impulsas mielininiu aksonu gali plisti tiktai nemielinizuotomis aksono dalimis (Ranvjė sąsmaukomis)
Kodėl mielininėse skaidulose nervinis impulsas plinta tik Ranvjė sąsmaukomis?
· aksono tipo - ar mielininis, ar nemielininis (mielininiu greičiau)
· elektrinių jonų kanalų tankio aksono membranoje - kuo tankiau kanalai išsidėstę, tuo greičiau plis impulsas, nes greičiau susidarys depoliarizacija
· aksono skersmens - kuo didesnis skersmuo, tuo mažesnė varža, tuo greičiau plinta nervinis impulsas.
Nuo ko priklauso nervinio impulso plitimo greitis?
1. Vientisumo dėsnis – nervinis impulsas plinta tik vientisa (nepažeista) nervine skaidula
2. Izoliuotumo dėsnis – nervinis impulsas plinta izoliuotai (negali ,,peršokti‘‘ nuo vienos nervinės skaidulos į kitą)
3. Abipusio laidumo dėsnis – nervinis impulsas skaidula gali plisti abejomis kryptimis (bet tik iki sinapsės).
Nervinio impulso sklidimo dėsniai
laikina grįžtama būsena, kai jaudrioji sistema nėra mirusi, bet neatlieka jai būdingų funkcijų.
Kas yra parabiozė?
Ją galima sukelti parabiotikais (pvz chloroformas, fenolis, kalio chloridas, novokainas). Parabiotikas inaktyvuoja Na kanalus. Parabiotikas - nebūtinai cheminė medžiaga, gali būti ir fizinis reiškinys, pvz.: šaltis (šalčiu galime paveikti jaudriąją sistemą taip, kad ji nebeatliks savo funkcijų).
Parabiozės taikymo sritis - anestezija.
Kas gali sukelti parabiozę? Kur ji taikoma?
Išlyginamoji fazė - vienodas raumens atsakas į įvairaus stiprumo dirgiklius
Paradoksinė fazė - silpnesnis dirgiklis (dirginamas nervas) sukelia stipresnį raumens atsaką nei stipresnis dirgiklis
Slopinamoji fazė - parabiotikas visiškai blokuoja NI, joks dirgiklis nebesukelia raumens atsako.
Prabiozė - grįžtamasis procesas, todėl grįžta iš slopinamosios į išlyginamąją, tačiau grįžtant į normalią būklę, atsistato atbuline eiga (3, 2, 1)
Parabiozės fazės
tai funkcinė jungtis tarp neuronų arba tarp neurono ir efektoriaus
Kas yra sinapsė?
Cheminė sinapsė yra sudaryta iš dviejų dalių: presinapsinė ir posinapsinė dalys. Tarp jų sinapsės plyšys.
Presinapsinė dalis vadinama sinapsės gumbeliu - tai aksono sustorėjimas (ten jis praranda mieliną); jame susikaupusios sinapsinės puslytės, kuriose yra neuromediatoriai.
Presinapsinėje membranoje yra labai daug elektrinių Ca2+ jonų kanalų, kurie vaidina labai svarbų vaidmenį depoliarizuojant presinapsinę membraną.
Posinapsinėje membranoje išsidėstę neuromediatoriui specifiniai receptoriai.
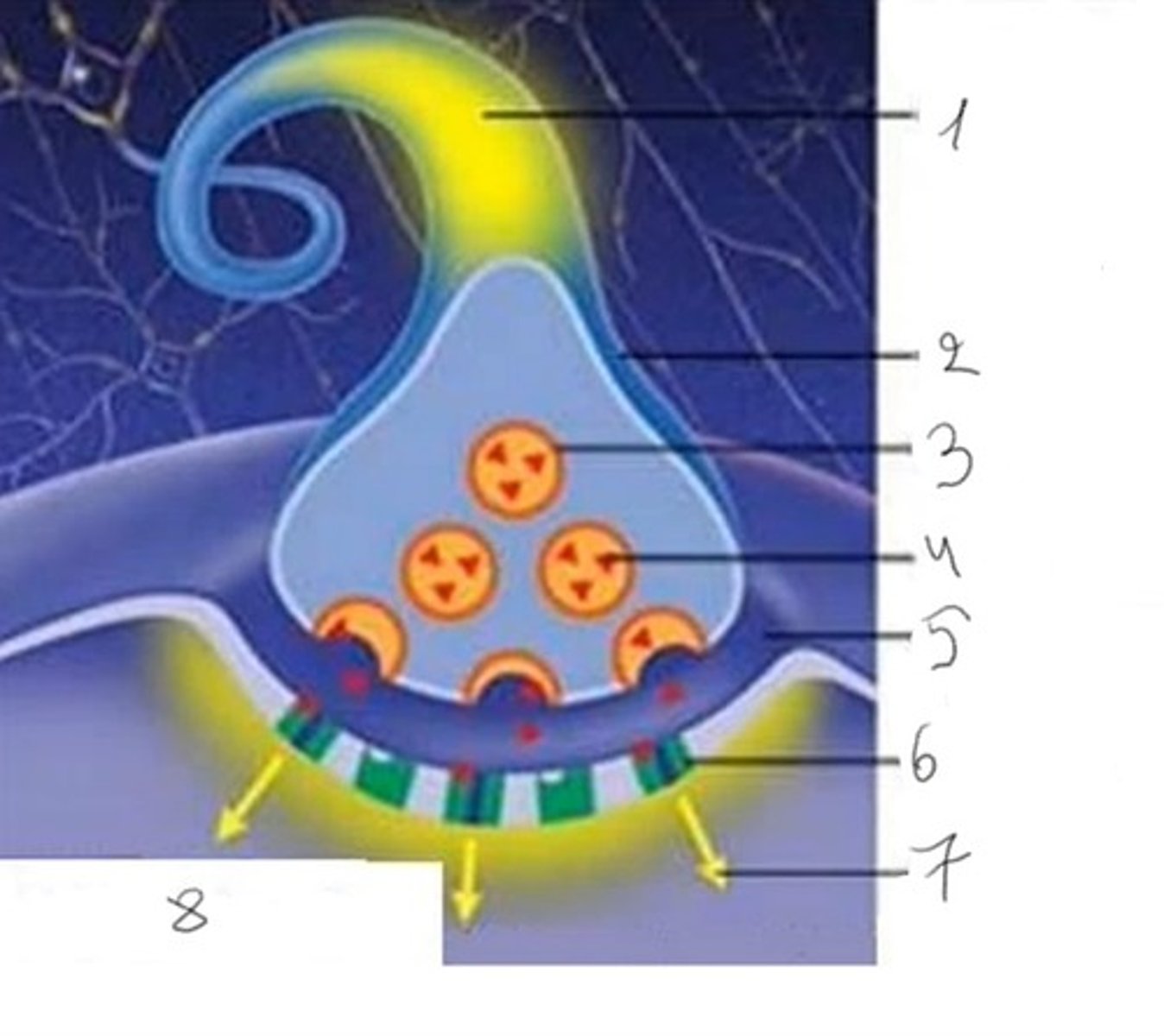
Sinapsės dalys
1. Nervinis impulsas
2 Presinapsinio neurono terminalė
3. Vezikulė
4. Mediatorius
5. Sinapsinis plyšys
6. Receptorius
7. Posinapsinis impulsas
8. Posinapsinė membrana
Cheminės
Elektrinės
Sinapsių skirstymas pagal signalų perdavimo pobūdį
Centrinės (galvos ir nugaros smegenyse)
Periferinės (jungtis tarp neurono-raumens, neurono-liaukos; pvz raumens galinė plokštelė)
Cheminių sinapsių skirstymas pagal išsidėstymą NS
Jaudinamosios
Slopinamosios
Cheminių sinapsių skirstymas pagal veikimo potencialo susidarymą posinapsinėje dalyje
Aksodendritinės - aksonas ir dendritas
Aksosomatinės - aksonas ir kito neurono kūnas
Dendrodendritinės
Aksoaksoninės - aksonas ir aksonas
Yra sinapsių, kur aksonas sudaro kolateralę ir sudaro sinapsę su savo paties kūnų - pvz Renšou ląstelės
Cheminių sinapsių skirstymas pagal centrinės sinapsės vietą
Cholinerginės
Adrenerginės
Dopaminerginės
Serotoninerginės
Peptidinerginės
Cheminių sinapsių skirstymas pagal neuromediatorių
Cholinerginiai - acetilcholinas
Adrenoerginiai - noradrenalinas
Dopaminerginiai - dopaminas
Serotoninerginiai - serotoninas (susiję su sąmoninga veikla, mokymusi, atmintimi, dėmesiu, motyvacija, tikslų siekimu)
GABA (gama aminosviesto rūgštis) - pagrindinis galvos smegenyse slopinantis mediatorius, glicinas
Glutamatas - siejamas su jaudinimo procesais NS
Neuromediatorių pvz
Sinapsėje susidaro sinapsinis potencialas (taip perduodamas impulsas). Kai nervinis impulsas pasiekia gumbelį, depoliarizuojama membrana, atsiveria elektriniai Ca jonų kanalai, ir jie ima judėti link gumbelio. Ca jonai, patekę į gumbelį, skatina pūslytes judėti link presinapsinės membranos ir susilieti su presinapsine membrana. Mediatorius, kuris yra pūslyčių viduje, egzocitozės būdu išsilieja į sinapsinį plyšį. Mediatoriai jungiasi prie specifinių postsinapsinėje membranoje esančių receptorių. Poveikis gali būti dvejopas (priklauso nuo mediatoriaus ir jonotropinių/metabotropinių receptorių).
Sinapsinio potencialo susidarymas
Jonotropiniai receptoriai tiesiogiai atveria jonų kanalą, kai prisijungia neuromediatorius. Jonotropinės sinapsės paplitusios ten, kur reikalingas greitas impulso perdavimas: klausos, regos, motorinėse sistemose. Tai integralinis/ kanalinis baltymas (glikoproteinas), jame yra vadinamas kanalas - pora. Šis kanalas ramybėje uždaras, bet kai prie jo prisijungia neuromediatoirus, jis atsiveria (cheminis jonų kanalas), jonai juda pagal konc. gradientą (vieni į ląstelę, kiti - iš jos).
Apibūdink jonotropinius receptorius
Metabotropiniai receptoriai prisijungę neuromediatorių tiesiogiai neatidaro jonų kanalų, o juos valdo per antrinį signalo perdavėją (tarpininką) – G baltymą. Tarpininkas aktyvina jonų kanalus ir jonai juda į ląstelę. Dažnai per tarpininkus perduotas nervinis impulsas yra sustiprinamas. Tai integralinis baltymas, tačiau jame nėra poros (kanalo), jis negali atsiverti prie jo prisijungus mediatoriui. Kai mediatorius jungiasi prie jo (susidaro receptoriaus-mediatoriaus kompleksas), kinta konformacija ir suaktyvinamas G baltymas (išsidėstęs vidinėje membranos pusėje, sudarytas iš subvienetų: alfa – aktyvioji G baltymo dalis, beta, gama), kurio alfa subvienetas toliau aktyvina adenilatciklazę ir yra paleidžiama tarpininkų grandinė (ciklizanF, inozitoltrifosfatas, Ca2+ jonai). Netiesiogiai aktyvinamas cheminis jonų kanalas, jis atsiveria ir jonai juda.
Apibūdink metabotropinius receptorius
Jonotropinis greitesnis
Kuris veikimas - jonotropinis ar metabotropinis - yra greitesnis?
Jonai, judėdami kanalais postsinapsinėje membranoje sukelia postsinapsinį potencialą
Kas sukelia postsinapsinį potencialą?
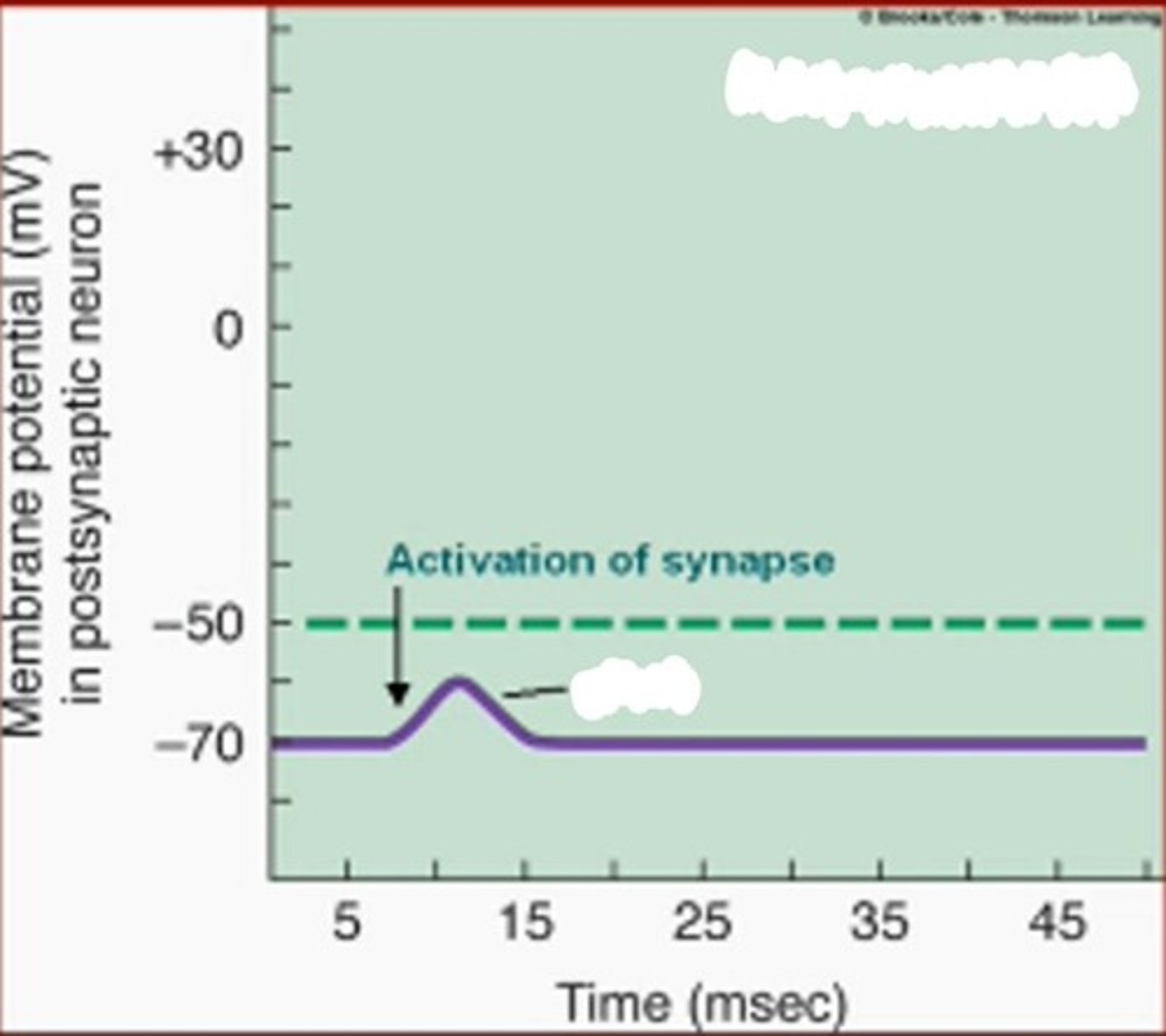
Jei sinapsėje skiriasi jaudinamasis mediatorius (pvz., acetilcholinas), tokiam mediatoriui prisijungus prie specifinio receptoriaus posinapsinėje membranoje, atsidarius cheminiam jonų kanalui, į posinapsinę dalį plūsta Na+, tai sukelia posinapsinės membranos depoliarizaciją. Membraninis potencialas tampa nebe toks neigiamas, aksono stormenyje kyla VP ir sklidimas vyksta toliau (jei pasiekiama slenkstinė įtampa).
Apibūdink jaudinamąjį potencialą
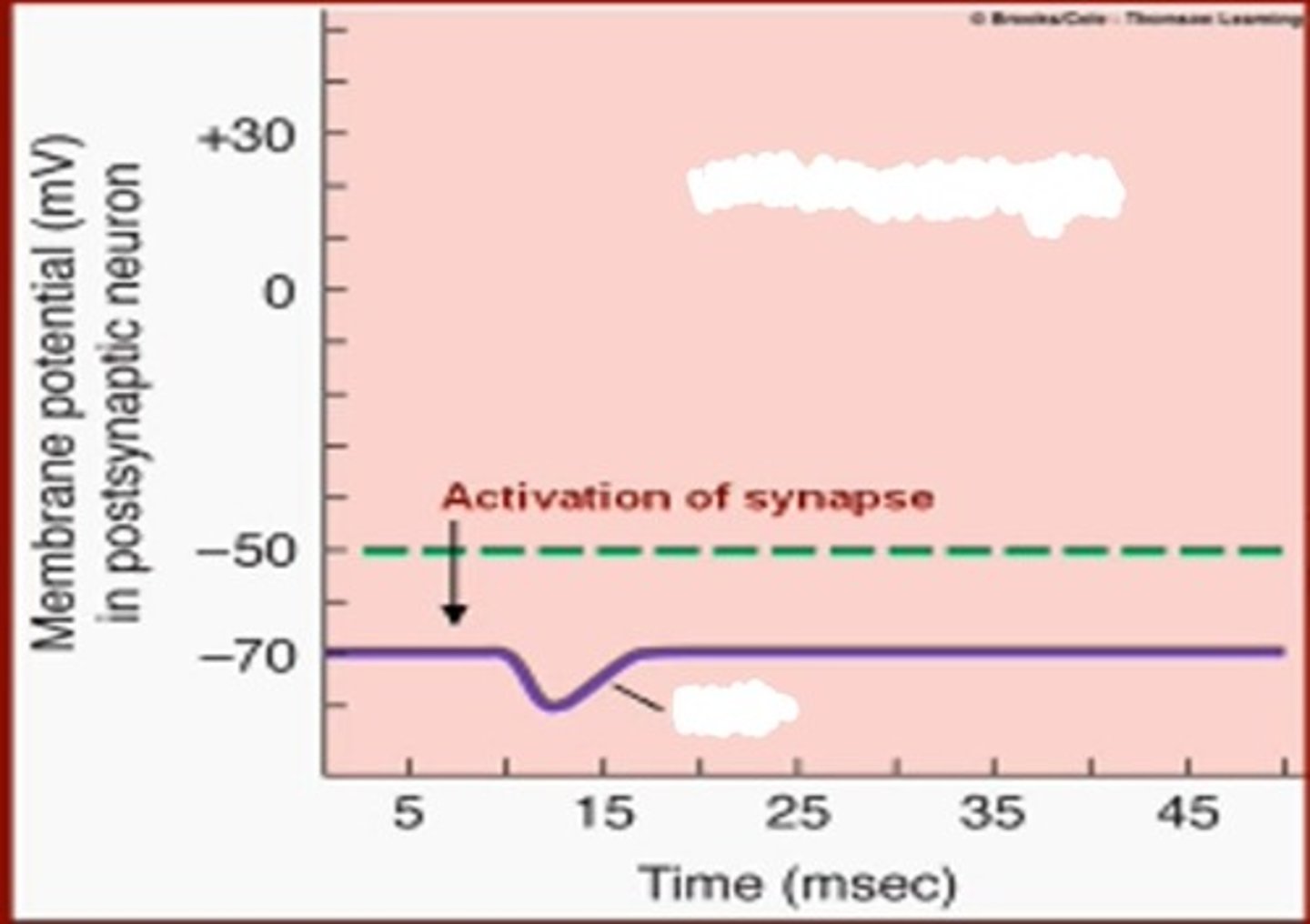
Jei sinapsėje skiriasi slopinamasis mediatorius (pvz., gama-aminosviesto rūgštis), tai toks mediatorius, prisijungęs prie specifinių receptorių posinapsinėje membranoje ir sužadinęs cheminių jonų kanalų atsidarymą (net dalyvaujant G baltymui ir kitam tarpininkui), sukelia K+ judėjimą iš posinapsinės ląstelės ir Cl- judėjimą į posinapsinę membraną. Posinapsinės membranos membraninis potencialas tampa labiau neigiamas nei buvo ramybėje (hiperpoliarizacija - slopinimas). Postsinapsiniame neurone potencialo nebėra ir impulsas nebesklis.
Apibūdink slopinamąjį potencialą
Slopinamasis arba jaudinamasis
Postsinapsinio potencialo poveikis būna ...
Jaudinamasis potencialas
Slopinamasis potencialas
Klasikiniai (mažamolekuliniai) neuromediatoriai
Neuropeptidai - stambiamolekuliniai junginiai; CNS mediatoriai
Neuromediatorių grupės
Acetilcholinas (Ach)
Amino rūgštys (GABA (gama-amino sviesto), glicinas, glutamatas)
Monoaminai (noradrenalinas, adrenalinas, dopaminas, serotoninai, histaminas)
Tirpūs dujiniai neuromediatoriai (azoto oksidas (NO) ir anglies monoksidas (CO)) - dėl jų lakumo tyrinėjimai labai sudėtingi (veikia centrines sinapses). Atrasti neseniai.
Daugiausiai veikia jonotropinius receptorius (bet glutamatas - metabotropinius).
Klasikinių neuromediatorių pvz. Kuriuos receptorius jie veikia?
Pogumburio peptidai (pvz., somatostatinas)
Hipofizės peptidai (pvz., STH, beta-endorfinai)
Žarnyno ir smegenų peptidai (pvz., medžiaga P, cholecistokininas, gastrinas, nervų augimo faktorius, enkefalinai)
Iš kitų audinių (pvz., ANGII (angiotenzinas II) - atsakingas už pirminį troškulį, kalcitoninas, bradikininas, oksitocinas - meilės hormonas)
neuropeptidų pvz.
Tiesiogiai nesukelia sinapsinio potencialo. Išsiskiria sinapsėje su klasikiniais mediatoriais. Veikia klasikinių neuromediatorių veikimo trukmę ir intensyvumą - moduliatoriai. Veikia metabotropinius receptorius, todėl būtinas G baltymas: G-baltymo alfa-subvienetas tiesiogiai aktyvina jonų kanalus, aktyvina tarpininkų sintezę (cAMP, IP3...), aktyvinančių fermentų sistemas ir metabolizmą; jungiasi prie DNR ir veikia genų ekspresiją. Peptiderginių sinapsių mediatoriai pasižymi santykinai lėtu veikimu (lyginant su klasikiniais mediatoriais, kurie jungiasi prie jonotropinių receptorių)
Neuropeptidų veikimo mechanizmas
Posinapsinio potencialo vertė nedidelė, jis slenksčio dažniausiai nepasiekia. Ypač centrinėse sinapsėse veikimo potencialas susidaro retai, nes slenkstis nepasiekiamas.
Kodėl vyksta sumacija postsinapsinėje dalyje?
Laiko ir erdvės
Sumacijos tipai
Sinapsėje kyla keletas sinapsinių potencialų, kurie eina vienas po kito serijomis. Tuomet jie susisumuoja, pasiekiama slenkstinė įtampa, slenkstinė membraninio potencialo vertė, prasideda depoliarizacija posinapsinio aksono stormenyje ir kyla veikimo potencialas. Ir viena sinapsė gali sukelti VP.
Laiko sumacija
Keliose sinapsėse tuo pačiu metu kyla sinapsinis potencialas. Atkeliauja sinapsiniai potencialai mažiausiai 2 sinapsėmis, posinapsinėje dalyje sumuojasi sinapsiniai potencialai. Pasiekę slenkstį sukelia slenkstinę įtampą aksono stormenyje, atsiranda VP, kuris sklinda iki kitos aksono sinapsės.
Erdvės sumacija
Širdies skersaruožiuose raumenyse;
Lygiuosiuose raumenyse;
Smegenų glijos audiniuose.
Kur paplitusios elektrinės sinapsės?
· Nervinis impulsas plinta plyšinėmis jungtimis (angl. gap junctions)
· Kanalas sudarytas iš dviejų puskanalių - koneksonų
· Koneksonas sudarytas iš 6 vienodų subvienetų - koneksinų
· Nėra mediatorių
Elektrinių sinapsių sandara
Nes nėra mediatorių, jonai jungtimis plinta dėl krūvių skirtumo
Kodėl elektrinės sinapsės greitesnės?
Impulsas gali plisti visomis kryptimis
Kuria kryptim plinta elektrinės sinapsės?
Cheminėse sinapsėse postsinapsinis potencialas ir VP plinta dėl mediatorių, o elektrinėse nėra mediatorių, nervinis impulsas čia plinta dėl elektrinių jėgų poveikio (dėl elektrinių krūvių pokyčio, elektrolitų).
Cheminėse sinapsinis potencialas perduodamas dėl jonų koncentracijos gradiento atsivėrus kanalams (joniniams, metaboliniams), o elektrinėse jonai plinta dėl elektrinių krūvių skirtumo žymiai greičiau.
Cheminėse NI perduodamas viena kryptimi nuo presinapsinės į postsinapsinę sritį, elektrinėse NI gali plisti visomis kryptimis (raumuo veikia kaip vienetas)
Cheminės labiau paplitusios, elektrinės retesnės, bet vyrauja tam tikruose audiniuose (širdies skersaruožių raumenų, lygiųjų raumenų, epiteliniame, CNS neuroglijoje, tinklainėje)
Cheminė lėtesnė, nes reikia laiko kol depoliarizuosis presinapsinė membrana, atsidarys elektriniai Ca 2+ jonų kanalai, puslytės susilies su presinapsine membrana, išsilies mediatorius. Elektrinėse impulsas plinta be jokių trukdžių, todėl greitesnės
Palygink chemines ir elektrines sinapses
Raumens ir nervo jungtis
Kas yra galinė plokštelė?
Somatinės nervų sistemos impulsai motoriniu neuronu pasiekia nervo ir raumens jungtį, kurioje kyla sinapsinis potencialas. Kiekvieną griaučių raumens skaidulą inervuoja vienas α motorinio neurono aksonas. Vienas α motorinis neuronas inervuoja daug raumeninių skaidulų, nes jo aksonas daug kartų šakojasi, sudarydamas periferines chemines sinapses. Prieš pat galinę plokštelę aksonas netenka mielino dangalo ir suskyla į plonesnes šakeles. Kiekviena šakelė sudaro sustorėjimą - sinapsės gumbelį, kuriame išsidėsčiusios sinapsinės pūslytės. Jose susikaupęs neuromediatorius acetilcholinas (Ach). Tarp sinapsės gumbelio ir raumens skaidulos sarkolemos yra tarpas - sinapsės plyšys. Sarkolemos invaginacijos į raumens skaidulą sudaro T-vamzdelių sistemą, kuriose yra terminalinių sarkoplazminio tinklo cisternų, pilnų Ca2+ jonų
Nervo-raumens jungties struktūra
· Kūno judesių formavimas (mechanika)
· Kūno padėties palaikymas (statika)
· Tuščiavidurių organų tūrio reguliavimas (šlapimo pūslės, kraujagyslių)
· Pernašos funkcija (širdies siurblio - kraujo pernaša, virškinamojo trakto peristaltika - maisto pernaša)
· Šilumos gamyba - dalyvauja termoreguliacijoje (drebulys)
Svarbiausios raumenų fiziologinės funkcijos
sarkomerai, kuriuose ir vyksta raumens susitraukimas
Raumeninės skaidulos struktūrinis vienetas
Ca jonai, veikiant Ca siurbliui (Ca ATPazei), grąžinami į sarkoplazminį tinklą. Manoma, kad Ca siurblys atplėšia Ca jonus nuo baltymo parvalbumino, kuris labai greitai sujungia tam tikrą dalį Ca jonų, išsiskyrusių ir sarkomplazminio tinklo. Sarkoplazmos lasivųjų Ca jonų koncentracijai sumažėjus iki 10^-7 mol/l, tropomiozinas užblokuoja aktyviąsias aktino vietas. Tarp raumenų gijų (miofibrilių) esant pakankamai ATP, raumuo atsipalaiduoja. Veikiant Ca siurbliams, Ca jonai sugrąžinami į sarkoplazminio tinklo išilginius vamzdelius ir galinėse talpyklose surišami su baltymu kalsekvestrinu. Dirbant Ca siurbliams tiesiogiai naudojama ATP (pirminė aktyvioji pernaša). Stingant ATP, skersiniai miozino tilteliai išlieka prisijungę prie aktino stipriosios jungties būsenos, nes susidaro rigoro kompleksas. Todėl griaučių raumenims būdingas rigor mortis (pomirtinis sustingimas).
Raumens atsipalaidavimas
α judinamasis neuronas sujaudinamas. VP plinta judinamojo neurono aksonu į raumenines skaidulas. Nervo ir raumens sinapsėje veikimo potencialas sukelia acetilcholino (Ach) išsiskyrimą, kuris lemia galinės plokštelės potencialo susidarymą postsinapsinėje membranoje. Galinės plokštelės potencialas sukelia sarkolemos VP, kuris plinta raumeninės skaidulos paviršiumi ir patenka į skersinių vamzdelių T sistemą. VP T sistema plinta iki membranos potencialo valdomų elektrinių Ca kanalų (DHP prijungiamųjų baltymų), kurie mechaniniu būdu per pėdos struktūrą atidaro sarkoplazminio tinklo Ca kanalus. Atsidarius Ca kanalams, Ca2+ koncentracijos gradiento kryptimi difunduoja iš sarkoplazminio tinklo į sarkoplazmą, supančią miofibriles, ir daug kartų didina viduląstelinę Ca2+ koncentraciją (nuo 10^-7 iki 10^-5 mol/l), ypač padidėja Ca2+ koncentracija prie aktino ir miozino siūlelių. Dauguma Ca2+ labai greitai sukimba su troponinu C, pakinta viso troponino baltymų komplekso struktūra, pašalinamas slopinamasis troponino I poveikis, per troponiną T perduodamas signalas tropomiozinui, ir visas troponino-tropomiozino kompleksas pasislenka į įdubą tarp dviejų aktino grandinių, užleisdamas vietą miozino galvutei (dalis Ca sukimba su parvalbuminu). Skersiniai tilteliai tarp miozino ir aktinio sukamaisiais judesiais sukelia sarkomerų ir visos raumeninės skaidulos susitraukimą ir jėgos didėjimą, jiems padeda raumeninės ląstelės citoskeletas.
Raumens skaidulos susitraukimas
viena troponino C molekulė prisijungia iki 4 Ca2+
Kiek Ca jonų gali prisijungti viena troponino C molekulė?
Ca2+ koncentracijos padidėjimas sarkoplazmoje sukelia griaučių raumens skaidulos susitraukimus.
Kas sukelia raumens skaidulos susitraukimus?
miozinas, aktinas, tropomiozinas, troponinas, ATP bei Ca2+ jonai.
Kurios molekulės dalyvauja slenkamųjų filamentų mechanizme?
Storieji filamentai sudaryti iš daugelio miozino molekulių. Viena miozino molekulė turi judrią galvutę ir uodegą. Miozino galvutė formuoja skersinius tiltelius su aktinu. Miozino uodega geba svertiškai judėti. Miozino galvutės ir uodegos judesiai būtini susidarant skersiniams tilteliams. Miozino galvutėje yra ATP-aziniu aktyvumu pasižyminti sritis, gebanti prisijungti ATP. Prisijungus ATP ir jai hidrolizuojantis į ADP ir neorganinį fosfatą, miozino galvutė aprūpinama energija. Kita miozino galvutės sritis pasižymi gebėjimu jungtis prie aktino.
Apibūdink storojo filamento struktūrą
Plonasis filamentas sudarytas iš baltymų: aktino, tropomiozino ir troponino. Aktinas - pagrindinis plonojo filamento komponentas. Aktino subvienatai, susijungę į grandinėlę ir susivynioję tarpusavyje, sudaro dvigubą spiralę. Kiekviename aktino subvienete yra specifinė sritis (aktyvusis centras), gebanti prijungti miozino galvutę. Prie aktino siūlų paslankiomis jungtimis yra prisitvirtinusios dvi tropomiozino grandinės. Nedirbančiame raumenyje tropomiozinas uždengia aktino aktyviuosius centrus, todėl miozinas negali jungtis prie aktino ir negali susidaryti skersiniai tilteliai. Kad miozino galvutės galėtų jungtis prie aktino, tropomiozino grandinės turi pasislinkti. Tai tampa įmanoma dėl plonojo filamento baltymo troponino. Troponinas tolygiais tarpais yra išsidėstęs išilgai visos tropomiozino grandinės. Pats troponinas nesugeba pastumti tropomiozino grandinės. Tam būtini Ca2+ jonai, kurie išsiskiria iš sarkoplazminio tinklo galinių cisternų ir prisijungia prie troponino. Tai sukelia troponino ir tropomiozino komplekso erdvinius pokyčius, tropomiozino grandinė pasislenka nuo aktyvių aktino grandinės centrų, gebančių prijungti mioziną.
Apibūdink plonojo filamento struktūrą